Development of the Citrus Longhorned Beetle Anoplophora chinensis (Cerambycidae: Coleoptera) on Artificial Diet and Chilling Effect on Their Life Cycle Completion
Hai Nam Nguyen, Ki-Jeong Hong

TL;DR
Researchers found that chilling Anoplophora chinensis larvae at specific temperatures and durations improves their pupation success when reared on an artificial diet.
Contribution
The study introduces optimized chilling regimes and artificial diet protocols for successful laboratory rearing of Anoplophora chinensis.
Findings
Pupation success increased with longer chilling periods, reaching 55% after 16 weeks at 10 °C.
Developmental durations ranged from 34 to 89 weeks depending on the number of chilling cycles.
Adults reared on artificial diet had significantly lower body weight compared to field-collected individuals.
Abstract
This study examined how cold treatment (chilling) affects the development of Anoplophora chinensis larvae reared on an artificial diet and their life cycle completion. Larvae were exposed to low temperatures (5 °C and 10 °C) for various durations and then returned to warm conditions to monitor pupation. Results showed that longer chilling periods increased pupation success: 55% pupated after 16 weeks at 10 °C, compared with 16.7% after 12 weeks and none after 9 weeks. Some larvae required one to three chilling cycles to complete their life cycle, with development times ranging from 34 to 89 weeks. The body weight of adults emerging from the artificial diet was significantly lower than those collected in the field. Overall, the study demonstrates that both chilling duration and temperature are critical for successful pupation, offering insights that may support scalable rearing of this…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1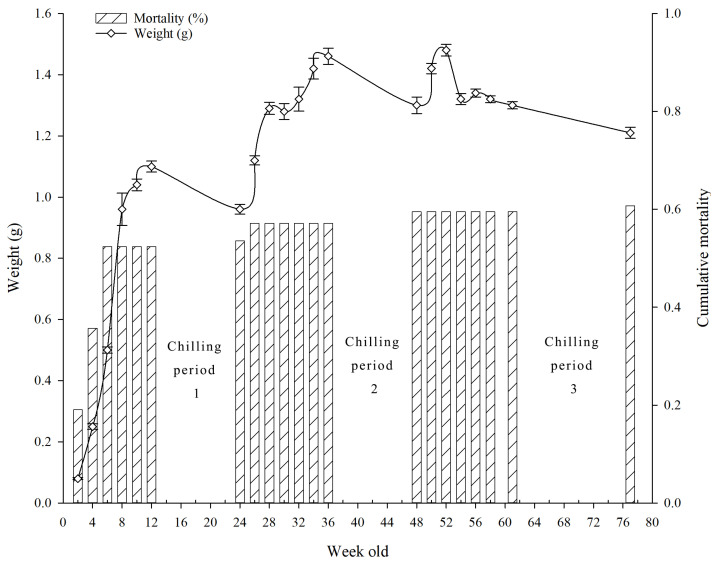 Figure 2
Figure 2| Ingredient | Supplier | Unit | Volume | Remark |
|---|---|---|---|---|
| Distilled water | IS Chemical, Busan, Republic of Korea | mL | 900 | Solvent |
| Agar powder | Chemicals Duksan, Incheon, Republic of Korea | g | 30 | Gelling agent |
| Wheat germ | Bob’s Red Mill, Milwaukie, OR, USA | g | 65 | Proteins, lipids |
| Sucrose | Sigma-Aldrich, St. Louis, MO, USA | g | 18 | Carbohydrates |
| Casein from bovine | Sigma-Aldrich, St. Louis, MO, USA | g | 9.4 | Proteins, amino acids |
| Yeast extract | Sigma-Aldrich, St. Louis, MO, USA | g | 25 | Proteins, amino acids |
| Cellulose powder 38 μm | Fujifilm Wako, Osaka, Japan | g | 121 | Fiber |
| Wesson salt | Thermo Fisher Scientific, Waltham, MA, USA | g | 0.9 | Mineral mix |
| Vitamin mix * | NOW Foods, Bloomingdale, IL, USA | capsule | 4 | Vitamins |
| Sodium benzoate | Sigma-Aldrich, St. Louis, MO, USA | g | 2 | Preservative |
| Citric acid | Sigma-Aldrich, St. Louis, MO, USA | g | 2 | Preservative |
| Sorbic acid | Sigma-Aldrich, St. Louis, MO, USA | g | 1.6 | Preservative |
| Tetracycline | Sigma-Aldrich, St. Louis, MO, USA | g | 0.2 | Antibiotic |
- —National University Development Project of Suncheon National University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForest Insect Ecology and Management · Coleoptera: Cerambycidae studies · Insect Pest Control Strategies
1. Introduction
The citrus longhorned beetle—CLB (Anoplophora chinensis Forster, 1771; Coleoptera: Cerambycidae)—is currently recognized as one of the most serious invasive wood-boring pests in Europe and Asia. A. malasiaca is regarded as a synonym of A. chinensis and was frequently cited in earlier Japanese literature [1]. Morphological variation and the presence of multiple COI haplotypes within the species reflect considerable genetic diversity among populations from different geographic regions [1,2,3]. This polyphagous xylophagous beetle is native to East Asia, particularly China, Japan, and Korea, as well as several Southeast Asian countries, including Vietnam, Malaysia, Indonesia, and the Philippines [1,4]. Following initial incursions into Europe in the early 2000s, primarily linked to imported bonsai and nursery stock [2,4,5], A. chinensis remains recorded in several European countries as of late 2025, including Italy, France, Croatia, Germany, the Netherlands, Poland, Slovenia, and Slovakia. In Switzerland, the species has been declared eradicated, while scattered reports also exist from Austria, Belgium, Denmark, Estonia, Finland, Lithuania, and Sweden [5,6]. Species distribution models and climate variables consistently indicate that the CLB has the potential to expand its global range well beyond its native distribution. Europe and North America are identified as regions at particularly high risk. Climate change is projected to increase the extent of suitable habitats, especially at higher latitudes [7,8].
CLB attacks a wide range of broadleaf hosts, including citrus, maple, willow, poplar, and chestnut, causing severe damage through larval tunneling in woody tissues [4,9]. Its life cycle includes oviposition beneath host tree bark, prolonged larval tunneling within woody tissues, pupation in the trunk or root system, and adult emergence during the summer months. This species is considered highly destructive, as larvae bore deeply into the xylem and cambial layers, thereby weakening structural integrity and often resulting in host tree mortality [4].
Early Japanese studies established that the CLB has a multiyear life cycle lasting up to three years, with larval survival strongly influenced by temperature regimes [10]. A previous study further demonstrated the feasibility of rearing larvae on cellulose-based artificial diets, although growth was slower compared to natural hosts [11]. Subsequent work confirmed citrus, ornamental, and forest trees as preferred hosts, with adult females exhibiting higher fecundity and survival [12,13]. Building on these foundations, more recent studies in Europe and China have refined artificial diet formulations and climate control protocols. Favaro et al. (2017) developed a general diet suitable for multiple invasive cerambycids, showing that CLB can complete development but with reduced efficiency relative to A. glabripennis Motschulsky, 1854 [14]. Cai et al. (2016) demonstrated that combining artificial diets with controlled temperature regimes (20–30 °C) enables successful rearing from egg to adult [15], while other works highlighted the critical role of temperature, with development arrested at ≤10 °C or ≥35 °C [16,17].
Collectively, these studies underscore both the feasibility and the challenges of mass rearing the CLB, emphasizing that reliable colonies require specialized diets and strict environmental regulation [14,15,17]. Larval development in A. chinensis is strongly influenced by exposure to low temperatures [17]. Overwintering larvae can extend their life cycle under fluctuating thermal regimes, with chilling periods helping to synchronize development with seasonal cycles [10,11]. Larvae enter developmental arrest at ≤10 °C and resume growth when temperatures rise, indicating a facultative diapause-like response [10,15]. Chilling is critical for CLB larvae [17]; without this period, development is disrupted, survival rates decline, and adult emergence becomes irregular. Although artificial diets can sustain continuous development under optimal conditions (20–30 °C), the absence of chilling often reduces fitness and longevity [10,15].
Despite evidence that exposure to low temperatures (≈10 °C for ~12 weeks) is critical for A. chinensis larvae to complete their life cycle [14,17], several uncertainties remain. Previous conclusions were based on limited experimental designs and did not assess whether shorter or longer durations could also be effective. Moreover, the precise temperature thresholds remain uncertain. To address this gap, the present study expands the range of chilling durations and temperature gradients, aiming to refine our understanding of diapause termination and improve rearing protocols.
In this study, we evaluated the development of A. chinensis under artificial diet conditions and examined the effects of chilling duration and temperature on the completion of their life cycle. Our objective was to address the above-mentioned gap in order to refine artificial rearing protocols and strengthen pest management strategies for this destructive longhorn beetle.
2. Materials and Methods
2.1. Insect Sources
CLB adults were manually collected from willow trees, Salix alba L. (Malpighiales: Salicaceae), growing along the Dongcheon River in Suncheon-si, Jeollanam-do, Republic of Korea (34.982874 N, 127.489675 E), with a total of 26 females and 27 males during June and July of 2023 and 2024 (Table 1).
Adults collected from the wild were transferred to the breeding room and weighed for further comparisons. The insects were paired in plastic vessels (10 × 15 × 20 cm) or mass-reared in breeding cages (40 × 40 × 40 cm) at a density of approximately 4–5 pairs to obtain eggs. Water was provided in a 5 oz cup containing soaked cotton on top. Fresh willow twigs served as food, and larger twigs (1–2 cm in diameter; 15 cm long) were placed for oviposition. Food and oviposition substrate were replaced daily. The insects were maintained in the breeding room at a temperature of 24–26 °C under a photoperiod of 16L:8D, with RH ranging from 40–60%.
The eggs were harvested by gently detaching them from beneath the host plant bark after peeling the twigs. Subsequently, they were surface-sterilized with 5% sodium hypochlorite solution, rinsed with distilled water, and individually placed into a Combi Cryo-Box (SCB4500, 81-hole type; Ecocell, Gyeonggi, Republic of Korea) or a Corning Falcon tissue culture dish (flat bottom 96 wells type, Sigma-Aldrich, St. Louis, MO, USA) to prevent larval cannibalism after hatching (Figure 1).
2.2. Artificial Diet Preparation
The diet was modified from the AG2 diet described by Keena (2005) [18] (Table 2). Agar powder was mixed with distilled water and heated to boiling. Wheat germ, sucrose, sodium benzoate, and citric acid were added at boiling, while the remaining ingredients were incorporated after cooling for 5 min. All components were then blended for 3 min. The diet was poured into 1 oz or 5 oz plastic cups, and lids were closed once the diet had cooled to prevent dehydration. The prepared diet was stored in a refrigerator until use.
2.3. Development of A. chinensis Larvae on the Artificial Diet
Neonate larvae, upon hatching, were transferred into 1 oz artificial diet cups via a small perforation created on the diet surface to facilitate larval entry, and each cup was labelled. Diet cups were replaced at two-week intervals or earlier if signs of fungal or bacterial contamination were observed. Prior to transfer into fresh diet cups, larvae were surface-sterilized by immersion in a 5% sodium hypochlorite solution for 5 s, followed by rinsing in distilled water for an additional 5 s. At six weeks of age, the larvae were relocated to larger 5 oz diet cups to accommodate growth.
In 2023, a total of 84 newly hatched larvae were used to examine the development of C. chinensis on an artificial diet. Larval weights were recorded at two-week intervals. All larvae were reared under identical conditions for 12 weeks before being placed in an incubator (VS-1203P1-150-0, Vision Scientific, Daejeon, Republic of Korea) set at 10 °C under dark conditions to stimulate pupation (RH: 20–40%). Following the chilling period, larvae were returned to the rearing room (T = 26 ± 2 °C; RH = 40–60%; 16L:8D photoperiod) and maintained for an additional 12 weeks. Larvae that pupated during this interval were recorded, whereas those that did not pupate were subjected to a second 12-week chilling treatment at 10 °C. After the second chilling, larvae were again reared for 12 weeks under the same room conditions; individuals that still failed to pupate were then exposed to a third chilling treatment, during which the temperature was set at two levels (5 °C and 10 °C) for 16 weeks. The dates of pupation and adult emergence were recorded, and adult body weight was measured 7 days post-emergence to allow comparison with wild-collected adults.
2.4. Determining the Effect of Chilling Duration and Chilling Temperature on A. chinensis
The larvae used in this experiment were reared from eggs laid by adults collected in 2024. The diet and rearing methods followed those described above. Because the number of larvae of the same age was insufficient, chilling treatments could not be performed simultaneously. Instead, larvae that had reached 12 weeks of age and attained a body weight of at least 1 g were sequentially placed into cold chambers (VS-1203P1-150-0, Vision Scientific, Daejeon, Republic of Korea) set at 10 °C, with 15–20 individuals per cohort. An additional treatment at 5 °C was conducted with 20 individuals for a duration of 16 weeks. The remaining larvae (17 individuals) were maintained in the rearing room (as above) without exposure to cold treatment.
After accumulating cold exposure durations for 9, 12, 14, 16, and 19 weeks at the two temperature levels, age-matched cohorts were transferred back to the rearing room (T = 26 ± 2 °C; RH = 40–60%; 16L:8D photoperiod) for further observation. The new diet cup was replaced immediately after larvae were reintroduced to the rearing room and subsequently every 2 weeks (as above protocol). The number of pupae obtained under each experimental condition was recorded until larvae had been maintained in the rearing room for an additional 12 weeks, in order to calculate pupation percentage.
2.5. Data Analysis
Adult body weight was compared using one-way ANOVA after confirming normality with the Shapiro–Wilk test. Pearson’s chi-square test was used to assess the association between pupation rate and duration of cold treatment. When significant differences were identified, Fisher’s Exact Test was applied for each pairwise comparison, and the resulting p-values were compared with the adjusted significance level (α = 0.05) following Benjamini and Hochberg’s method [19]. Data analysis was performed using Minitab v22 (Minitab, LLC, State College, PA, USA) and Excel (Microsoft, Redmond, WA, USA).
3. Results
A. chinensis was able to develop on an artificial diet. Larval mortality increased sharply during the first 6 weeks (>50%) and then stabilized. After 12 weeks, the mean larval weight reached approximately 1.10 g, rising to 1.46 g after 36 weeks. Certain individuals required up to three cold treatments before completing the immature stages (Figure 2).
The pupation percentage after the first and second chilling was only 7.7–12.9%, whereas in the third treatment, 60% of larvae developed into adults. Development from larval hatching to adult emergence in CLB was completed after one, two, or three chilling periods, corresponding to 34.43, 55.93, and 88.65 weeks, respectively (Table 3).
The body weight of CLB adults reared on an artificial diet was significantly lower than that of wild-collected adults for both females (p = 0.001) and males (p = 0.036). On the artificial diet, females reached a mean weight of 1.06 g, compared with 1.28 g for individuals collected from the wild. For males, mean weights were 0.67 g on the artificial diet and 0.79 g in the wild (Table 4).
The duration of cold treatment did not affect CLB mortality (χ^2^ = 2.785, df = 5, p = 0.73), but it had a statistically significant effect on pupation percentage after chilling (χ^2^ = 22.699, df = 5, p < 0.01). The percent of larvae pupated under the 16-week chilling treatment was 55%, which was significantly higher than the shorter chilling treatments of 12 weeks, where pupation reached only 16.7%. No individuals pupated under the 9-week treatment. Interestingly, one individual pupated in the non-chilled cohort but failed to develop into an adult (Table 5).
Chilling temperature also had a marked effect on the pupation percentage of CLB. In both the 2023 and 2024 experiments, no individuals pupated following exposure to 5 °C. In contrast, at 10 °C, 55% of chilled CLB larvae pupated in the 2023 experiment and 60% in the 2024 experiment, with subsequent adult emergence. These differences were statistically significant (p = 0.002 in the 2023 experiment and p < 0.001 in the 2024 experiment) (Table 6).
4. Discussion
The present study demonstrates that A. chinensis can complete development on artificial diets, albeit with notable constraints. Larval mortality was high during the first six weeks (>50%) before stabilizing, and growth remained slow, with mean larval weights reaching 1.10 g at 12 weeks (prior to the first chilling). These results are consistent with earlier work on A. chinensis [14,15] and A. malasiaca [11,20], which showed that artificial diets can sustain development but with lower performance compared to natural hosts, and with elevated mortality, particularly among younger instars. In contrast, recent studies emphasized the critical role of temperature in artificial rearing success, demonstrating that fluctuating temperature regimes substantially improved larval survival relative to constant conditions [15,17]. Keena and Richards (2022) further noted that the ecological origin of A. chinensis populations may influence survivorship, suggesting that genetic or geographic variation could modulate rearing outcomes [17].
The significantly lower adult weights observed in our study (females: 1.06 g vs. 1.28 g in the wild; males: 0.67 g vs. 0.79 g) reinforce the conclusion that current artificial diets compromise physiological performance, echoing previous reports of reduced fitness and fecundity under laboratory conditions [14]. Cai (2016) similarly found female weights ranging from 0.715–1.464 g and male weights from 0.500–1.129 g across diets and generations, with sawdust-based diets generally producing heavier females [15]. Favaro et al. (2017) likewise documented reduced adult mass and fecundity in laboratory-reared cohorts compared with natural hosts, underscoring that diet composition strongly influences developmental outcomes [14]. Importantly, previous studies revealed that larvae of certain cerambycid species may perform better on drier diets [17,21], a pattern consistent with the outcomes of sawdust-based diets reported by Cai (2016) [15].
Our study did not include direct nutritional analysis, but previous work on cerambycids and other wood-boring beetles has highlighted the importance of protein content, carbohydrate balance, and moisture levels in supporting larval growth and adult fitness [18,22]. This indicates that both nutrient composition and diet texture play key roles. The current diet appears more suitable for later instars, as survival rates were relatively high thereafter. However, comprehensive studies are still needed to determine the optimal nutrient composition and rearing conditions for A. chinensis, particularly during the early instars.
For many insect species, diapause and cold exposure are dialectically interconnected, a relationship that is particularly evident in species from temperate and cold regions [23,24,25,26]. These insects enter diapause to enhance their survival during harsh winter conditions, yet they also require the cold period itself to break dormancy and resume development in the warmer spring [23,27]. Empirical evidence demonstrates that numerous species are unable to complete their life cycle without experiencing winter chilling [23,27,28,29]. Understanding the nature of this relationship provides valuable insights into species distribution, informs the development of control strategies, and facilitates the successful rearing of these insects under laboratory conditions.
Cold treatment emerged as a critical factor influencing pupation success in A. chinensis. While mortality was unaffected by cold duration, pupation percentage increased significantly with longer chilling periods. Pupation was 55% after 16 weeks of chilling, compared with only 16.7% after 12 weeks, and no pupation after 9 weeks. These results align with Adachi (1994) [10] and Keena and Richards (2022) [17], who emphasized the necessity of chilling for pupation cues, but extended their findings by demonstrating that multiple chilling cycles may be required. In contrast to its sister, A. glabripennis generally requires only a single winter chilling period to complete its life cycle [14,18]. Indeed, pupation percentages after the first and second chilling were low (7.7–12.9%), whereas the third treatment yielded 60% adult emergence. This variability highlights heterogeneity in diapause intensity among individuals, a phenomenon also noted in A. malasiaca, where overwintering strategies are flexible, and development can extend across multiple seasons [10]. While diapause has been reported in A. chinensis [10], we did not measure physiological or endocrine markers to verify this developmental arrest. Larvae in this study were subjected to chilling after reaching 12 weeks of age, a time point chosen to standardize physiological development as suggested previously for A. glabripennis [14,18]. However, it remains uncertain whether 12 weeks is sufficient for diapause stabilization in A. chinensis. Longer pre-chilling durations (e.g., 14–20 weeks) may allow greater accumulation of energy reserves and endocrine adjustments, potentially enhancing the effectiveness of chilling in terminating diapause of A. chinensis. Future studies should therefore examine the optimal larval age at the onset of chilling to refine rearing protocols.
Temperature thresholds further shaped pupation outcomes. No individuals pupated at 5 °C, whereas 55–60% pupated at 10 °C, with subsequent adult emergence. These results clarify that diapause termination requires moderate chilling rather than extreme cold, consistent with Cai et al. (2016) [15], who showed that development arrests at ≤10 °C but resumes under controlled regimes of 20–30 °C. Together, these findings suggest that although chilling is essential, its effectiveness depends on both duration and temperature, with 10 °C representing a functional threshold for pupation cues in A. chinensis. A limitation of the present study is that only two chilling temperatures (5 °C and 10 °C) were tested. Consequently, the precise threshold for diapause termination could not be determined, and intermediate temperatures (e.g., 7–9 °C) may also be effective. Subsequent work may benefit from testing a broader set of chilling conditions to refine the temperature requirements for pupation cues in A. chinensis.
Cold exposure clearly plays a critical role in the development of A. chinensis, and longer periods of chilling may be favored by the species. Our results suggest that extended chilling (e.g., 16 weeks) promotes pupation, yet the underlying mechanism remains uncertain. It is unclear whether prolonged cold directly facilitates successful pupation or whether secondary effects, such as diet desiccation, contribute to improved outcomes [17,21]. Nevertheless, diapause in A. chinensis appears to be facultative, allowing some individuals to complete development without experiencing a cold winter [10,15]. In this study, we observed one larva that successfully pupated without chilling, similar to the findings of Cai et al. (2016) [15] and Adachi (1994) [10]. This flexibility helps explain why A. chinensis is primarily adapted to temperate and subtropical climates [1,4,6], yet shows a tendency to expand into higher latitudes under climate change scenarios [7,8], where longer winters may be required. At the same time, the species persists in warmer regions of southeastern Asia [4], highlighting its broad ecological tolerance and invasive potential.
Notably, low-temperature treatment of A. chinensis resulted in a strongly female-biased sex ratio among emerged adults after the first and second chilling treatments. Although our sample size was limited, similar patterns have been reported in other cerambycid species. Cerambyx welensii Küster, 1845 (Coleoptera: Cerambycidae) showed variation in sex ratio and adult size under different ecological conditions [30], and twig-girdling beetles in the Onciderini tribe exhibited sex ratio shifts linked to seasonality and climate [31]. These findings support the hypothesis that low-temperature exposure may act as a cue affecting sex differentiation in CLB, warranting further study with larger cohorts.
Overall, comparative evidence for A. chinensis highlights the dual challenges of artificial rearing: elevated early larval mortality and reduced adult fitness on artificial diets, along with complex diapause break requirements that necessitate carefully calibrated chilling regimes. While much remains to be done to improve rearing performance, these findings refine our understanding of CLB biology and provide practical guidance for establishing laboratory colonies. Our results underscore the importance of integrating diet optimization with precise thermal management and appropriate chilling duration into rearing protocols of this species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lingafelter S.W. Hoebeke E.R. Revision of the Genus Anoplophora (Coleoptera: Cerambycidae)Entomological Society of Washington Washington, DC, USA 2002235
- 2Iwaizumi R. Arimoto M. Kurauchi T. A study on the occurence and fecundity of white spotted longicorn, Anoplophora malasiaca (Coleoptera: Cerambycidae)Res. Bull. Plant Prot. Jpn.201450915
- 3Ohbayashi N. Ogawa J. Su Z.-H. Phylogenetic analysis of the Lamiine genus Anoplophora and its relatives (Coleoptera, Cerambycidae) based on the mitochondrial COI gene Spec. Bull. Jpn. Soc. Coleopterol. Tokyo 20137309324
- 4Haack R.A. Hérard F. Sun J. Turgeon J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective Annu. Rev. Entomol.20105552154610.1146/annurev-ento-112408-08542719743916 · doi ↗ · pubmed ↗
- 5Hérard F. Maspero M. History of discoveries and management of the citrus longhorned beetle, Anoplophora chinensis, in Europe J. Pest Sci.20199211713010.1007/s 10340-018-1014-9 · doi ↗
- 6EPPO Global Database Distribution of Anoplophora chinensis Available online: https://gd.eppo.int/taxon/ANOLCN/distribution(accessed on 9 January 2026)
- 7Zhang L. Wang P. Xie G. Wang W. Impact of climate change on the distribution of the citrus longhorned beetle Anoplophora chinensis (Coleoptera: Cerambycidae) and its host plants, Castanea mollissima and Castanea seguinii: A predictive analysis using optimized Max Ent models J. Econ. Entomol.202511874675910.1093/jee/toae 30839832258 · doi ↗ · pubmed ↗
- 8Zhou Y. Ge X. Liu J. Zou Y. Guo S. Wang T. Zong S. Climate change effects on the global distribution and range shifts of citrus longhorned beetle Anoplophora chinensis J. Appl. Entomol.202214647348510.1111/jen.12996 · doi ↗