Revision of the Genus Ranacris You & Lin, 1983 (Orthoptera: Acrididae: Catantopinae), with Proposal of a New Synonym
Zhongke Lv, Jun Cai, Benyong Mao, Xun Wang, Hong Song, JoVonn G. Hill, Xiongyan Yin, Hanqiang Wang, Jianhua Huang

TL;DR
This paper revises the genus Ranacris and proposes a new synonym based on morphological and statistical analysis of species within the genus.
Contribution
The paper proposes a new junior synonym and clarifies the taxonomy of the genus Ranacris using morphometric analysis.
Findings
R. yunnanensis is a junior synonym of R. jinpingensis.
Morphometric analysis confirmed the statistical significance of distinguishing characters in Ranacris.
Ranacris shows a close relationship with Menglacris despite being in different subfamilies.
Abstract
The genus Ranacris You & Lin, 1983, is revised based on the examination of types and non-type material and the comparison of qualitative and quantitative characters. A new junior synonym is proposed: R. yunnanensis Mao, Ren & Ou, 2011 = R. jinpingensis Zheng, Lin, Deng & Shi, 2015, syn. nov. Ranacris You & Lin, 1983, is a small genus in the subfamily Habrocneminae with an undefined phylogenetic position, exhibiting in previous molecular studies a distant relationship with the genera in the same subfamily but an extremely close relationship with the genus Menglacris Jiang & Zheng, 1994, in another subfamily, Catantopinae. In this study, a taxonomic revision of Ranacris was conducted based on the examination of types and non-type material and the comparison of qualitative and quantitative characters. A morphometric analysis was carried out to test the statistical significance of the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2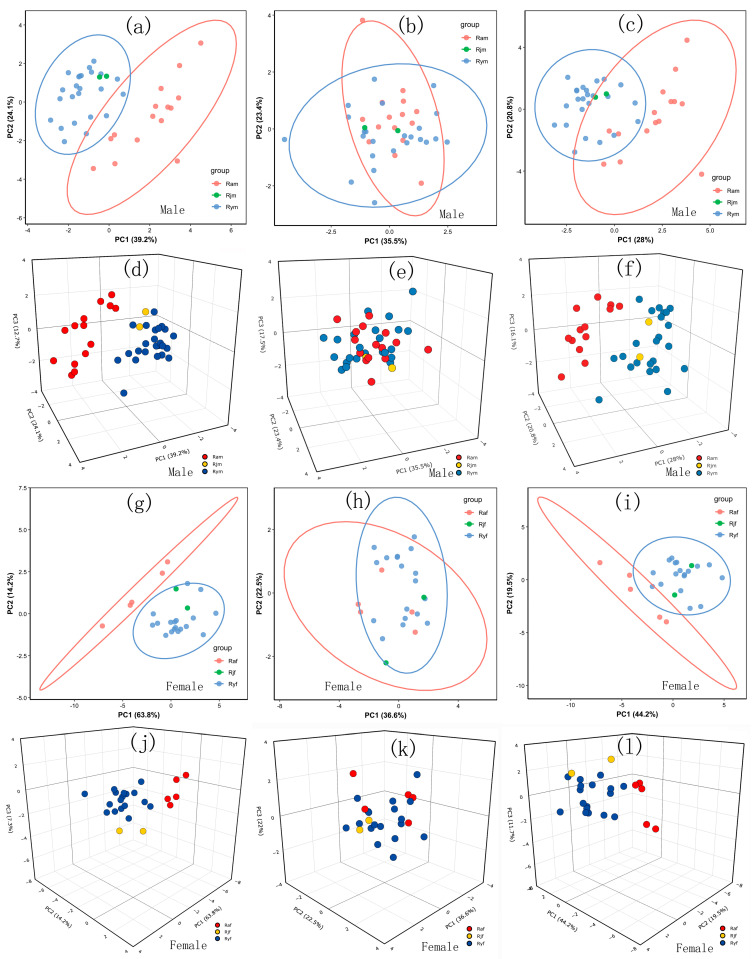 Figure 4
Figure 4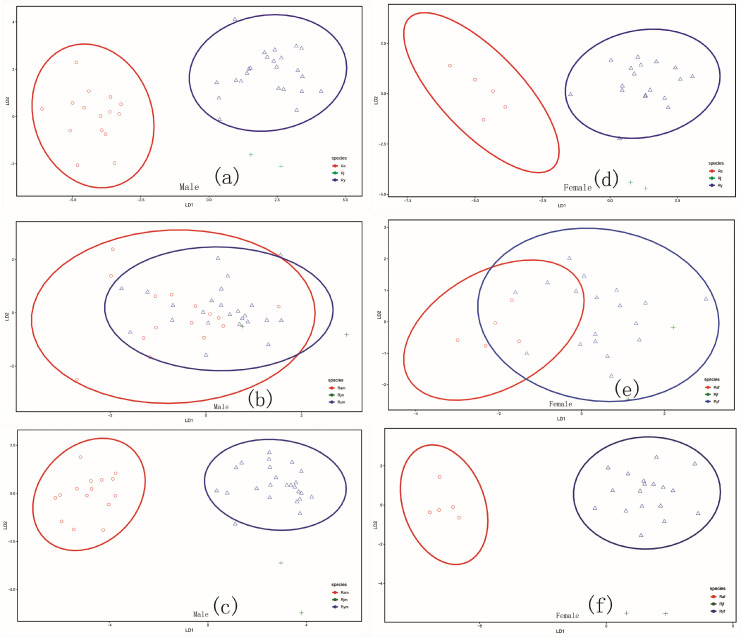 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8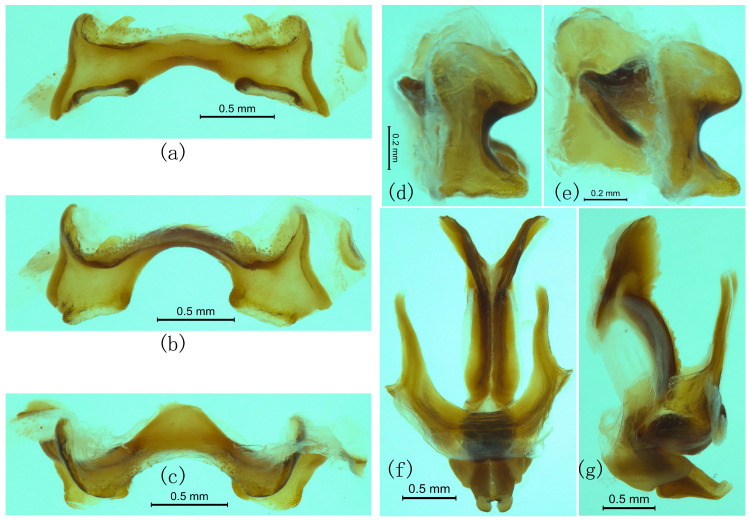 Figure 9
Figure 9| Species Pairs | BL | PNL | HFL | HFW | IOD | LDE | TDE | SOFL |
|---|---|---|---|---|---|---|---|---|
| Ram–Rym | W = 47 | W = 196 | W = 327 | W = 322 | W = 297 | W = 338 | W = 320 | W = 354 |
| Ram–Rjm | W = 15 | W = 4 | W = 26 | W = 23 | W = 22 | W = 29 | W = 25 | W = 30 |
| Rym–Rjm | W = 43 | W = 0 | W = 15 | W = 12 | W = 11 | W = 19 | W = 23 | W = 38 |
| Species pairs | PZL | MZL | IOD/LDE | LDE/TDE | LDE/SOFL | PZL/MZL | HFL/HFW | |
| Ram–Rym | W = 227 | W = 135 | W = 209 | W = 213 | W = 117 | W = 239 | W = 161 | |
| Ram–Rjm | W = 10 | W = 2 | W = 21 | W = 17 | W = 4 | W = 26 | W = 16 | |
| Rym–Rjm | W = 4 | W = 4.5 | W = 15 | W = 26 | W = 15 | W = 36 | W = 29 |
| Species Pairs | BL | PNL | HFL | HFW | IOD | LDE | TDE | SOFL |
|---|---|---|---|---|---|---|---|---|
| Raf–Ryf | W = 28 | W = 73 | W = 82 | W = 88 | W = 77 | W = 85 | W = 54 | W = 90 |
| Raf–Rjf | W = 8 | W = 8 | W = 10 | W = 10 | W = 8 | W = 10 | W = 8 | W = 10 |
| Ryf–Rjf | W = 36 | W = 15 | W = 25 | W = 22 | W = 7 | W = 30 | W = 18 | W = 25 |
| Species pairs | PZL | MZL | IOD/LDE | LDE/TDE | LDE/SOFL | PZL/MZL | HFL/HFW | |
| Raf–Ryf | W = 75 | W = 65 | W = 49 | W = 62 | W = 4 | W = 44 | W = 46 | |
| Raf–Rjf | W = 8 | W = 6 | W = 1 | W = 8 | W = 1 | W = 6 | W = 4 | |
| Ryf–Rjf | W = 21.5 | W = 13 | W = 6 | W = 20 | W = 17 | W = 22 | W = 19 |
| Species | Male | Female | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination Based on Full Data | Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination Based on Full Data | |||||||||||||
| Ram | Rym | Rjm | Ram | Rym | Rjm | Ram | Rym | Rjm | Ram | Rym | Rjm | Ram | Rym | Rjm | Ram | Rym | Rjm | |
| Ram | 15 | 0 | 0 | 7 | 4 | 0 | 15 | 0 | 0 | 5 | 0 | 0 | 5 | 2 | 0 | 5 | 0 | 0 |
| Rym | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 2 |
| Rjm | 0 | 24 | 0 | 8 | 20 | 1 | 0 | 24 | 0 | 0 | 18 | 0 | 0 | 16 | 1 | 0 | 18 | 0 |
| Species | Male | Female | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination based on Full Data | Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination Based on Full Data | |||||||
| Ram | Rym | Ram | Rym | Ram | Rym | Raf | Ryf | Raf | Ryf | Raf | Ryf | |
| Ram | 16 | 0 | 5 | 4 | 17 | 0 | 7 | 1 | 4 | 1 | 7 | 0 |
| Rym | 1 | 24 | 12 | 20 | 0 | 24 | 0 | 17 | 3 | 17 | 0 | 18 |
| Species | Male | Female | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination Based on Full Data | Discrimination Based on Measurements | Discrimination Based on Ratio Indices | Discrimination Based on Full Data | |||||||
| Ram | Rym | Ram | Rym | Ram | Rym | Raf | Ryf | Raf | Ryf | Raf | Ryf | |
| Ram | 15 | 0 | 7 | 4 | 15 | 0 | 5 | 0 | 5 | 2 | 5 | 0 |
| Rym | 0 | 26 | 8 | 22 | 0 | 26 | 0 | 20 | 0 | 18 | 0 | 20 |
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOrthoptera Research and Taxonomy · Subterranean biodiversity and taxonomy · Spider Taxonomy and Behavior Studies
1. Introduction
Ranacris You & Lin, 1983, was established with R. albicornis You & Lin, 1983, as the type species [1] and three known species to date [2]. It was initially classified in the family Catantopidae sensu Li & Xia, 2006 [3], and not assigned to a specific subfamily. Subsequently, it was placed in the subfamily Ranacridinae You et al., 1990 [4], which was later synonymized with the tribe Mesambrini Brunner von Wattenwyl, 1893, within the subfamily Catantopinae Brunner von Wattenwyl, 1893 [5,6]. Mao et al. [7] described the second species of the genus, R. yunnanensis. Later, the third species, R. jinpingensis Zheng, Lin, Deng & Shi, 2015 [8], was added to the genus. Menglacris Jiang & Zheng, 1994, is a genus in the subfamily Habrocneminae [2,3,9]. While Ranacris and Menglacris belong to different subfamilies in the modern classification scheme of Acrididae [2,3,10], mitogenomic and morphological evidence suggests a closer phylogenetic relationship between them [11].
When described, both R. yunnanensis and R. jinpingensis were compared with R. albicornis to reveal the distinguishing characters for each species. According to the original description by Mao et al. [7], R. yunnanensis can be distinguished from R. albicornis by (1) the more distinct sculptures and punctures on the body surface, especially on the dorsum of pronotum; (2) the more pronounced carinate lateral margins of vertex between eyes; (3) the nearly straight anterior and median transverse sulci of pronotum; (4) the peltate supra-anal plate of male vs. the triangular one in R. albicornis; (5) the distinctly smaller ratio of prozona length to metazona length in males.
The distinguishing characteristics of R. jinpingensis from R. albicornis proposed by Zheng et al. [8] are (1) the medially concave anterior margin of vertex; (2) the absent fastigial foveolae; (3) the absent furculae in males; (4) the medially triangularly protuberant posterior margin of subgenital plate in females; (5) the smaller ratio of prozona length to metazona length in males.
During the identification of specimens of Ranacris collected from Fenshuiling, Jinping County, Yunnan Province, China, we compared the three described species of the genus and found that R. yunnanensis and R. jinpingensis share the same type locality and exhibit no significant morphological differences.
In this study, the type specimens of all three known species of Ranacris were examined, and additional specimens from the type localities were analyzed to assess the morphological variation. A detailed morphological comparison was conducted to confirm whether the diagnostic characters proposed in the original references were really significantly different among species, and the taxonomic status of the Ranacris species was reassessed. Based on these findings, we provide a comprehensive revision of Ranacris and propose a new junior synonym, i.e., Ranacris yunnanensis Mao, Ren & Ou, 2011 = Ranacris jinpingensis Zheng, Lin, Deng & Shi, 2015, syn. nov.
2. Materials and Methods
2.1. Material Acquisition
This study is based on the specimens, including types, deposited in the insect collections of the Central South University of Forestry and Technology (CSUFT), the Biological Sciences Museum at Dali University (BMDU) and the Museum of Zoology at Shaanxi Normal University (SNU), respectively (for detailed information, see the “Material examined” section for each species). All photographs were taken using a Nikon D600 digital camera (Nikon Inc., Tokyo, Japan) or a Leica DFC 5500 system (Leica Microsystems Inc., Wetzlar, Germany), and the images were stacked using Helicon Focus version 6.0. The terminology for morphology follows Uvarov [12] and Storozhenko et al. [13].
2.2. Measurement Acquisition
A total of 233 specimens, including 21 of R. albicornis, 208 of R. yunnanensis and 4 of R. jinpingensis, were examined, among which 20 of R. albicornis, 42 of R. yunnanensis and 4 of R. jinpingensis were selected for morphometric analysis. Ten quantitative characters (absolute measurements) were measured according to the following criteria using a digital vernier caliper or using an ocular micrometer under a stereomicroscope. In addition, five ratio indices were computed from the measured data.
BL: body length—the distance from the apex of the fastigium to the tip of the abdomen along the longitudinal axis of the body (excluding the valvula in females).
HFL: hind femur length—the distance from the base of the hind femur to the apex along its longitudinal axis.
HFW: hind femur width—the maximum width of the hind femur along its transverse axis.
IOD: interocular distance—the minimum distance between the inner margins of the eyes.
LDE: longitudinal diameter of the eyes—the maximum length of the eyes along their longitudinal axis.
MZL: metazona length—the distance from the posterior transverse sulcus of the pronotum to the rearmost point of the posterior margin of the pronotum along the median keel. When the posterior margin of the pronotum is medially concave, MZL is measured from the midpoint of the posterior transverse sulcus to the intersection of the median keel (extended posteriorly) with the line connecting the posterolateral extremities of the posterior margin of the pronotum (excluding the median notch).
PNL: pronotum length—the distance from the frontmost point of the anterior margin of the pronotum to the rearmost point of the posterior margin along the median keel. When the anterior and posterior margins of the pronotum are medially concave, PNL is measured as the distance between the intersection points of the extension line of the median keel of the pronotum and the two tangent lines of the pronotum through the anterior and posterior margins.
PZL: prozona length—the distance from the frontmost point of the anterior margin of the pronotum to the posterior transverse sulcus of the pronotum along the median keel. When the anterior margin of the pronotum is medially concave, PZL is measured as the distance from the intersection point of the extension line of the median keel of pronotum and the connecting line between the frontmost points of the anterior margin of the pronotum to the median point of the posterior transverse sulcus of the pronotum.
SOFL: length of subocular furrow—the distance from the lower margin of the eyes to the base of the mandible along the subocular furrow.
TDE: transverse diameter of eyes—the maximum width of eyes along its transverse axis.
2.3. Data Analysis
The mean and standard deviation of the absolute measurements were computed using R functions “mean()” and “sd()” in the R packages of “base” and “stats”, respectively. The two-sample Wilcoxon test (also known as “Mann-Whitney” test) was performed using the R function “wilcox.test()” in the “stats” package because some variables do not follow a normal distribution. The principal components analysis (PCA) was performed using the R function “prcomp()” in the “stats” package. The plots based on the principal components (PC1, PC2 and PC3) coordinates from the PCAs were drawn using the “ggplot2” and “plotly” packages. The linear discriminant analysis was conducted using the R function “lda()” in the “MASS” package, and the score plots were drawn using the “ggplot2” package. All computations and operations were implemented in R 4.5.2 (https://www.r-project.org, accessed on 5 March 2026).
3. Results
3.1. Comparison of Qualitative Distinguishing Characters Among Ranacris Species
Re-examination of the type series and additional material demonstrates that none of the diagnostic characters proposed for R. yunnanensis and R. jinpingensis are discrete; all vary continuously within populations (Table 1; Figure 1 and Figure 2).
3.2. Morphometric Analysis of Quantitative Characters
According to the original description, both R. yunnanensis and R. jinpingensis could be distinguished from the reference species, R. albicornis, by a quantitative character, the ratio of prozona length to metazona length (PZL/MZL) in male [7,8]. However, the measured data in this study (Tables S1 and S2) demonstrates that the mean of PZL/MZL in males is 2.49 ± 0.44 for R. albicornis, 2.39 ± 0.53 for R. yunnanensis and 2.23 ± 0.04 for R. jinpingensis, respectively (Table S3), far smaller than the values provided in the original references [1,7,8]. The females also have similar values of PZL/MZL (Table S4). The Wilcoxon test for quantitative characters demonstrates that PZL/MZL again shows no significant differences in both males and females across the three species (Table 2 and Table 3).
Comparison of qualitative distinguishing characters among Ranacris species. (a–c) Pronotum in dorsal view. (d–f) Head and pronotum in lateral view. (g–l) Vertex in dorsal view. (m–o) Supra-anal plate of male in dorsal view. (a,d,g–j,m–o) R. albicornis. (g,m) Holotype male of R. albicornis. (n,o) Paratype male of R. albicornis. (b,e,k) Holotype male of R. yunnanensis. (c,f,l) Holotype male of R. jinpingensis.
Comparison of qualitative distinguishing characteristics among Ranacris species (continued). (a–c) Supra-anal plate of male in dorsal view. (d–s) Fastigial foveola. (t–v) Subgenital plate of female. (a,d–k,t) R. albicornis. (b,l–q) R. yunnanensis. (b) Holotype male of R. yunnanensis. (u) Paratype female of R. yunnanensis. (c,r) Holotype male of R. jinpingensis. (s) Paratype male of R. jinpingensis. (v) Paratype female of R. jinpingensis.
Most of the measurements and ratio indices in both male and female show no significant difference between R. albicornis and R. jinpingensis, as well as between R. yunnanensis and R. jinpingensis (p > 0.05; Table 2 and Table 3; Figure 3), except the LDE and SOFL in males between R. albicornis and R. jinpingensis (p = 0.04 and 0.03, respectively; Table 2), the PNL in males (p = 0.02; Table 2) and the body length in females between R. yunnanensis and R. jinpingensis (p = 0.03; Table 3). However, seven of the ten measurements in both male and female show significant differences between R. albicornis and R. yunnanensis (p < 0.01 in males and p = 0.04, 0.01, 0.001, 0.02, 0.003, 0.0008, 0.03 in females, respectively; Table 2 and Table 3). No ratio index in males shows a significant difference between any species pair, and only LDE/SOFL in females shows a significant difference between R. albicornis and R. yunnanensis (p = 0.002; Table 3).
As for the distribution pattern of measured values, there is overlap in all measurements between most species pairs (Tables S1 and S2; Figure 3). The mean body length of R. yunnanensis in both males and females is larger than that of R. albicornis (Tables S3 and S4), with a significant difference in males (p = 0.0001; Table 2) but not in females (p = 0.22; Table 3). However, the extremum ranges of body length are similar between R. albicornis and R. yunnanensis and highly overlap (Tables S3 and S4; Figure 3a,f). Among the ten measurements, R. albicornis has six extreme ranges in males and nine in females with the largest mean. While the male R. jinpingensis has the largest mean pronotum length and prozona length, the male R. albicornis is still larger than the male R. yunnanensis in terms of these two values. The male R. albicornis has the smallest mean only in terms of the metazona length. In summary, R. albicornis is slightly larger than R. yunnanensis and R. jinpingensis in most measurements.
The principal components analysis (PCA) demonstrates that the first two or three principal components explain a high percentage of the variance (Figure 4), and the measurements can distinguish unambiguously between R. albicornis and R. yunnanensis, but cannot distinguish R. jinpingensis from R. yunnanensis. The two R. jinpingensis samples fall completely within the 95% confidence interval of R. yunnanensis in the PCAs based on measurements and full data (Figure 4a,c,d,f,g,i,j,l). The ratio indices have no resolution in discriminating among species (Figure 4b,e,h,k), and the combined data’s resolution is slightly reduced due to the poor performance of the ratio indices (Figure 4c,f,i,l).
The linear discriminant analysis (LDA) indicates that both measurements and the full data can perfectly discriminate between R. albicornis and R. yunnanensis (Figure 5a,d,c,f), but the ellipse of the 95% confidence interval for R. jinpingensis cannot be calculated due to too few points (Figure 5). Just as in PCAs, the ratio indices cannot accurately discriminate any species from others in LDAs again (Figure 5b,e; Table 4). Since R. jinpingensis differs significantly from both R. albicornis and R. yunnanensis in only a few measurements and ratio indices, we performed the LDAs again, assigning the R. jinpingensis samples to R. albicornis and R. yunnanensis, respectively. When the samples of R. jinpingensis were assigned to R. albicornis, one male sample was discriminated based on the measurements as R. albicornis with a posterior probability of 0.7857, but the other male sample was discriminated as R. yunnanensis with a posterior probability of 0. 5247 (Table 5 and Table S5). While the female samples of R. jinpingensis were all correctly discriminated as R. albicornis based on measurements, one female sample of R. yunnanensis was simultaneously incorrectly discriminated as R. albicornis (Table 5 and Table S6). The discrimination for both male and female based on full data was completely correct (Table 5, Tables S5 and S6). When the samples of R. jinpingensis were assigned to R. yunnanensis, the discriminations for both male and female based on measurement and full data were all correct (Table 6, Table S7 and S8). The proportion of incorrect discrimination was always very high based on ratio indices (Table 5 and Table 6).
Score plots of the principal components analysis. (a–f) Male. (g–l) Female. (a,d,g,j) Score plots of PCAs based on the ten measurements. (b,e,h,k) Score plots of PCAs based on the five ratio indices. (c,f,i,l) Score plots of PCAs based on the full data. The codes for the species are as follows: Ram—male R. albicornis, Rym—male R. yunnanensis, Rjm—male R. jinpingensis, Raf—female R. albicornis, Ryf—female R. yunnanensis, and Rjf—female R. jinpingensis.
Plot of discriminant analysis of Ranacris spp. (a–c) Male. (d–f) Female. (a,d) Plots drawn from ten measurements. (b,e) Plots drawn from the five ratio indices. (c,f) Plots drawn from full data. The codes for the species are the same as in Figure 4.
3.3. Taxonomy
3.3.1. Genus Ranacris You & Lin, 1983
Ranacris You & Lin, 1983: 257 [1]; Otte, 1995: 334 [10]; Yin et al., 1996: 608 [14]; Jiang & Zheng, 1998: 188 [15]; Li & Xia, 2006: 637 [3]; Mao et al., 2011: 152 [7]; Zheng et al., 2015: 177 [8]; Storozhenko, 2018: 56 [6]; Zhang et al., 2023: 18 [11].
Type species: Ranacris albicornis You & Lin, 1983, by original designation.
Generic diagnosis. Body medium-sized. Head shorter than pronotum, face distinctly reclinate in profile. Antennae filiform, reaching or exceeding the posterior margin of pronotum. Pronotum tectiform, with slightly raised median keel and distinct lateral keels. Prosternal spine conical and straight; apex pointed. Lateral lobes of meso- and metasterna broadly separated throughout. Tegmen vestigial, hind wing absent. Hind femora with upper basal lobe of outer surface distinct longer than lower one, upper median keel sparsely serrate and denticulate apically, and lower genicular lobe rounded. Hind tarsi normal, arolium developed. Tympanal organ large and nearly rounded. In males, the tenth abdominal tergite broadly split in the middle, with a small arciform furcula at each side of the split; supra-anal plate triangular; subgenital plate short, conical, with apex acute and tilted upwards. Cerci conical. Upper valvula straight.
Distribution. China (Guangxi, Yunnan).
3.3.2. Species. Ranacris albicornis You & Lin, 1983
(Figure 6, Figure 7a and Figure 8a–g)
Ranacris albicornis You & Lin, 1983 [1]: 258; You et al., 1990: 70 [4]; Otte, 1995: 334 [10]; Jiang & Zheng, 1998: 189 [15]; Li & Xia, 2006: 638 [3]; Mao et al., 2011: 152, 154 [7]; Zheng et al., 2015: 179 [8]; Zhang et al., 2023: 7, 19, 21 [11].
Type locality: China (Pingxiang, Guangxi).
Material examined. Holotype ♂, the peak of Daqingshan, Pingxiang County, Guangxi, China, 1000 m, 13 October 1980, leg. Rizhao Lin, Hongxian Ji, Qijing You (SEMCAS, 14510043); paratypes, 2♂, data the same as holotype (SEMCAS, 14510044, 14510045). Other material examined: 8♂, 2♀, Diding Nature Reserve, Nanpo Township, Jingxi County, Guangxi, China, 800–900 m, 8 August 2010, leg. Jianhua Huang; 5♂, 3♀, Diding Nature Reserve, Nanpo Township, Jingxi County, Guangxi, China, 23.1207806° N, 105.9738116° E, 20 August 2025, leg. Zhongke Lv.
Habitus and closeups of Ranacris albicornis You & Lin, 1983. (a–c) Holotype male. (a,b) In dorsal and lateral views. (c) Labels (The Chinese in the lower left is the Chinese name of R. albicornis, and that in the upper right is the detailed type locality). (d–w) Closeups of male. (d–f) Frons. (g–i) Antenna. (g,h) In dorsal and lateral views. (i) Apex. (j) Left tegmen. (k) Meso- and metasterna. (l–o) Fore leg in inner, outer, dorsal and ventral views. (p–s) Middle leg in outer, inner, dorsal and ventral views. (t–w) Hind femur in dorsal, outer, ventral and inner views. (x,y) Female in dorsal and lateral views.
Habitus and closeups of Ranacris spp. (a) Left tegmen of female R. albicornis. (b) Left tegmen of male R. yunnanensis. (c–f) Tegmen of female R. yunnanensis. (g–l) Types of R. yunnanensis. (g,h) Holotype male in dorsal and lateral view. (i) Labels (The Chinese in the lower label is the detailed type locality and the name of the collector). (j,k) Paratype female in dorsal and lateral view. (l) Labels (The Chinese in the lower lable is the detailed type locality and the name of the collector). (m–o) Types of R. jinpingensis. (m,n) Holotype male in dorsal and lateral views. (o) Labels (The Chinese in the upper left is the Chinese name of R. jinpingensis, and that in the upper right is the detailed type locality).
Male genitaliae of Ranacris albicornis and R. yunnanensis. (a–g) R. albicornis. (a–e) Epiphallus in dorsal, dorsofrontal, frontal, lateral and ventrolateral views. (f,g) Phallic complex in dorsal and lateral views. (h–n) R. yunnanensis. (h–l) Epiphallus in dorsal, dorsofrontal, frontal, lateral and ventrolateral views. (m,n) Phallic complex in dorsal and lateral views.
Morphology.
Male. Body medium-sized, well proportioned. Head shorter than pronotum, as long as or slightly longer than half of the pronotum length. Vertex slightly convex in lateral view, distinctly longitudinally carinated, either thickly or finely, along the lateral margins between eyes, and transversely carinated slightly before the narrowest part between eyes, with a distinct fine median keel which becomes obsolete on the hind half; fastigium hexagonal, strongly reclined forwards with anterior margin distinctly concave; foveola small, triangular (Figure 2h), or rectangular (Figure 2f,g), or indistinct (Figure 2d,e,i), or even nearly absent (Figure 2j,k). Antennae filiform, 25-segmented, distinctly exceeding the posterior margin of pronotum. Frons broad and flat, with a rounded fine sulcus at each side of the frontal ridge below the antennal socket and an oval prominence between the sulcus and the antennal socket. Frontal ridge deeply and broadly sulcate nearly throughout, lateral margins distinct, straight and parallel. Facial keels distinct and nearly straight. Clypeus inversely trapezoidal, with lateral margins slightly concave or bearing a small tubercle in the middle, anterior margin broadly and triangularly emarginate (Figure 6d–f). Eyes elliptical, with vertical diameter about 1.3–1.7 times the horizontal diameter and about 1.4–1.7 times the length of the subocular furrow. Subocular furrow broadly depressed. Genae with a distinct and straight vertical carina at or slightly behind the extension line of the longitudinal axis of eyes (Figure 1d). Pronotum tectiform, densely and coarsely punctured, with anterior margin slightly narrower than posterior margin and nearly straight (Figure 1a) or sometimes minutely concave in the middle (Figure 1g,h); lateral keels distinct, slightly incurved at the anterior transverse sulcus; median keel distinct throughout; all of the three transverse sulci distinct and interrupting the median keel, with the anterior and posterior ones straight and the median one posteriorly circular-arc-shaped; posterior transverse sulcus situated far behind the middle of pronotum, and prozona length about 2.10–3.08 times of metazona length. Meso-, metanotal and abdominal tergites distinctly carinate medially, with the meso-, metanotal and the first two abdominal tergites raised tectiformly. Interspace of the lateral lobes of mesosternum trapezoidal, with its maximum width about 1.77 times its length. Interspace of the lateral lobes of metasternum inversely trapezoidal, with its width distinctly narrower than that of its lateral lobes. Tegmina vestigial and hind wing absent. Hind femora well-proportioned and distinctly exceeding the apex of abdomen, with its length approximately 4.00–5.02 times its maximum width. Arolium developed, nearly as long as the length of claw. Supra-anal plate triangular, much longer than broad, with lateral margins bisinuate and elevated in the basal third; sometimes the lateral margins deeply emarginate subapically to form a mucronate apex (Figure 1m–o and Figure 2a); dorsum broadly longitudinally sulcated throughout and interrupted by a transverse carina and raised area. Cerci long, conical, broad and slightly compressed basally, covered with long pubescence; apex sharply pointed.
Epiphallus not divided into two symmetric halves; lateral plates broad, with lateral margins slightly oblique inwards and concave in the middle; anterior projections large, with bluntly rounded apices; ancorae small and triangular, slightly curved ventrally with apices pointed; posterior projections short and conical; bridge moderately broad, with anterior and posterior margins nearly straight in dorsal view, and posterior half strongly and crateriformly depressed in the middle in frontal and lateral views (Figure 8d,e); lophi large and rectangular in frontal view, obliquely situated along the inner margins of lateral plate; oval sclerites present. Phallic complex robust; basal valves of penis large and broad; zygoma broad; apodemes moderately long and robust, not reaching the apices of the basal valves of penis; valves of cingulum and apical valves of penis upcurved and conical with pointed apex in lateral view.
Female. Body larger than male, slightly plump. Eyes with vertical diameter about 1.50–1.77 times the horizontal diameter and about 1.04–1.27 times the length of subocular furrow. Prozona length about 2 times the metazona length. Interspace of the lateral lobes of mesosternum with its maximum width 1.58 times its minimum width and twice its length. Hind femora length approximately 4.37–4.42 times its width. Ovipositor moderately long; dorsal valvulae slightly robust, with apex strongly narrowed, sharply pointed and slightly curved upward; dorsolateral margins of the dorsal valvulae densely denticulate; ventral valvulae slightly slender, broad basally, gradually narrowed apically, with apex sharply pointed and slightly curved downwards. Subgenital plate quadrangular, with posterior margin triangularly protruding in the middle.
Coloration.
Body castaneous to buffy. Labrum and clypeus brown to yellowish brown. Frons buffy below median ocellus, and dark brown (Figure 6d) to yellowish brown (Figure 6e, f) above median ocellus. Antennae black to dark brown, with a distinct pale annulation near the apex (Figure 6g–i). Vertex with a broad long-trapezoidal dark brown stripe in the middle. Genae black to blackish brown behind the posterior margin of the subocular furrow. Pronotal dorsum with an indistinct V-shaped dark brown stripe both before and behind the median transverse sulcus; postocular band black. Meso-, metanotum and the first abdominal tergite with a broad and black longitudinal stripe in the middle and at each side, respectively; mesosternum black with a pair of large oval yellowish brown maculations at the anterior half (Figure 6k). Fore and middle legs buffy with some distinct or indistinct black transverse bands. Hind femora dark brown; outer surface with three distinct and one indistinct transverse black maculations (Figure 6u) and dorsal surface with four indistinct transverse dark maculations at the corresponding positions (Figure 6t); ventral surface with outer half black variegated with yellowish brown and inner half reddish brown. Hind tibiae reddish, basal half yellowish to dark brown with a distinct or indistinct yellowish brown annulation near the base. Abdomen black on the lateral surface; sternum has broad longitudinal black stripe in the middle.
Measurements. Body length: ♂, 17.40–23.03 mm, ♀, 23.18–29.21 mm; pronotum length: ♂, 4.70–5.70 mm, ♀, 6.50–7.78 mm; tegmen length: ♂, 1.18–1.24 mm, ♀, 1.21–1.33 mm; hind femur length: ♂, 12.25–13.98 mm, ♀, 15.39–18.26 mm.
Distribution. China (Guangxi).
3.3.3. Ranacris yunnanensis Mao, Ren & Ou, 2011
(Figure 7b–o, Figure 8h–n and Figure 9).
Ranacris yunnanensis Mao, Ren & Ou, 2011: 305 [7].
Ranacris jinpingensis Zheng, Lin, Deng & Shi, 2015: 178 [8]. Syn. nov.
Type locality: China (Fenshuiling Nature Reserve, Jinping County, Yunnan).
Material examined. Ranacris yunnanensis: Holotype ♂, Fenshuiling Nature Reserve, Jinping County, Yunnan Province, China, 1650 m, 24 July 2006, leg. Benyong Mao; paratypes: 59♂, 115♀, data the same as holotype except a larger altitude range of 1650–1850 m and more than one collectors: Benyong Mao, Zizhong Yang, Jishan Xu, Haoyu Liu, Qiqi Wu, and Juntong Lang (BMDU except 2♂, 2♀ in SEMCAS); fifteen 15♂, 13♀, data the same as paratypes (BMDU); 1♂, Fenshuiling Nature Reserve, JinPing County, Yunnan, China, 103.222° E, 22.854° N, 14 September 2024, leg. Ting Luo, Yanting Qin; 2♂, 2♀, Fenshuiling, JinPing County, Yunnan, China, 103.222° E, 22.854° N, 17 September 2024, leg. Ting Luo, Yanting Qin (CSUFT). Ranacris jinpingensis: holotype ♂, Fenshuiling, JinPing County, Yunnan Province, China, 7 September 2012, leg. Xun Bian; paratypes: 1♂, 2♀, data the same as holotype (SNU).
Morphology.
This species has no significant difference from R. albicornis in all qualitative characters, except the body size of R. albicornis is slightly larger than that of R. yunnanensis. However, the body length of R. albicornis measured in this study is slightly smaller than that of R. yunnanensis (Tables S1–S4). Epiphallus with the apex of the funnelform depression of bridge directed upwards (Figure 8k,l). There is no significant difference between R. yunnanensis and R. jinpingensis, including the male genitalia (Figure 8h–n and Figure 9).
Measurements. Body length: ♂, 18.60–24.00 mm, ♀, 23.20–29.00 mm; pronotum length: ♂, 4.80–5.44 mm, ♀, 5.92–7.18 mm; tegmen length: ♂, 0.90–1.04 mm, ♀, 1.33–1.41 mm; hind femur length: ♂, 11.50–12.90 mm, ♀, 14.00–16.10 mm.
Distribution. China (Jinping, Yunnan).
Remarks. The type localities of R. yunnanensis and R. jinpingensis are the same place, Fenshuiling Nature Reserve, Jinping County, Yunnan Province, China. These two species show no significant differences in qualitative characters, as well as in nearly all measurements and ratio indices, according to the results of the morphometric analysis. Therefore, we treated R. jinpingensis as a junior synonym of R. yunnanensis herein.
Male genitalia of Ranacris jinpingensis. (a–e) Epiphallus in dorsal, dorsofrontal, frontal, lateral and ventrolateral views. (f,g) Phallic complex in dorsal and lateral views.
4. Discussion
Body size is associated with many aspects of an organism’s biology, including local adaptations to different climatic conditions, female fecundity and male mating success, and so on [16]. Generally, the increase in altitude constrains growth rates and development times, leading to a smaller adult body size at high elevations than in lowlands [17,18,19,20,21]. R. albicornis inhabits altitudes between 700 and 1000 m, and R. yunnanensis occurs at high elevations between 1600 and 1950 m. Therefore, the larger body size of R. albicornis than R. yunnanensis can be explained at least partially by the hypothesis of “local adaptations to different climatic conditions” [16].
Usually, the body length of grasshoppers correlates positively with body size, i.e., a species with a larger body size generally has a larger body length correspondingly. However, in this study, as mentioned in the Results section, R. albicornis shows a larger body size but a smaller body length than R. yunnanensis in both males and females. How does this inconsistency occur? As far as we know, at least three factors may influence the measured value of body length: the accurate recognition of mature nymphs, the preserving method of specimens and the biological attribute of abdomen in female grasshoppers. (1) Ranacris species have vestigial tegmina completely covered by the lateral lobe of pronotum (Figure 6j), leading to difficulty in accurately discriminating between adult and mature nymph. While the sclerotization of integument and the development of genitalia can be used to help determine the adulthood of the samples, it is still difficult to completely ensure the accuracy of the determination because there is no explicit standard for the maturity of the integument and genitalia, and the determination is mainly based on personal experience. If a slightly high proportion of mature (last-instar) nymphs are wrongly recognized as adults and included in the measured samples, the measured values will decrease to some extent. (2) Some measured samples of R. albicornis collected in 2010 are shrunken strongly, especially the abdomen, due to dehydration when preserved in absolute ethyl alcohol, and these individuals generally exhibit smaller measured values, suggesting that these individuals may be mature nymphs and probably result in a substantial decrease in body length in R. albicornis. Dehydration with absolute ethyl alcohol will not only substantially decrease the body length of nymphs but also slightly shrink that of adults. For the purpose of preserving molecular material and the convenience of preventing the specimens from decaying in the field, all specimens of R. albicornis and the types of R. jinpingensis were preserved in absolute ethyl alcohol. However, the measured samples of R. yunnanensis, including partial types, were all poisoned via ethyl acetate in the field and dried naturally at ordinary temperatures, and this treatment has a much smaller influence on the body shrinkage. Since dehydration via absolute ethyl alcohol mainly acts on the highly retractile abdomen, body length becomes the most affected measurement, resulting in a smaller body length in R. albicornis than in R. yunnanensis. (3) The abdomen of female grasshoppers is biologically linked to the reproductive state (e.g., abdomen extension for oviposition). Individuals in the period of oviposition exhibit a much longer body length than in normal conditions. According to our observations, the time of collecting the examined specimens of R. albicornis may be during the early stages of eclosion, whereas the large number of recorded types of R. yunnanensis indicates it was at the peak of reproduction [7]. Of course, there is another possibility that the body length of R. yunnanensis may be larger than that of R. albicornis, but this requires further confirmation. Regardless, the total body length should be used carefully or even ignored sometimes as a diagnostic character, especially for closely related species, due to its substantial intraspecific variation [22].
When R. yunnanensis and R. jinpingensis were described, the authors indicated that male R. albicornis had a significantly larger ratio value of PZL/MZL than R. yunnanensis and R. jinpingensis. However, the remeasured values of PZL/MZL from the holotype male and two paratype males of R. albicornis show that it is significantly smaller (only 2.6–3.1; Table S1) than that (3.5–3.7) presented by You & Lin [1] in the original reference. In addition, the means of PZL/MZL of R. yunnanensis and R. jinpingensis in males are also significantly smaller (Table S3) than those presented in the original reference [7,8]. No significant difference in PZL/MZL was detected between any species pair in both male and female (Tables S3 and S4). Why did such an inconsistency occur? Besides inherent errors from different measurers, the use of different definitions or criteria for measurements may be another important factor contributing to such inconsistency. There are different definitions for a few absolute measurements of body parts. For example, Uvarov [12] defined the body length as “from apex of head to tip of abdomen”, but Mao et al. [7] and Hill [23] defined it as “dorsally from the fastigium vertices to the distal end of the genicular lobe of caudal femur in a parallel plane with the abdomen”. Hochkirch [22] illustrated the criteria for 27 measurements (excluding body length due to its sensitivity to shrinkage) and defined the metazona length in the figure as “the length from posterior transverse sulcus to the end of the median keel of pronotum”, and this was followed by Mao et al. [7]. While there is no clear definition for any measurement in the representative monographs of grasshopper taxonomy in China [3,24,25,26,27,28], most Chinese acridologists understand the metazona length as “the distance from the posterior transverse sulcus of pronotum to the rearmost point of the posterior margin of pronotum along the median keel” and accept Uvarov’s definition of body length [12]. Hochkirch’s definition of metazona length [22] means a smaller measured value, and this will result in a larger ratio value of PZL/MZL. Therefore, it is extremely important for all acridologists to use a unified set of criteria for the absolute measurements. The body length should be defined as the distance from one point of the soma to another one but not that of the appendages. We think Mao et al.’s [7] and Hill’s [23] definitions do not meet the essential implication for body length. Similarly, the metazona length should include the most distant point but not the bottom of the notch when the posterior margin of the pronotum is concave in the middle, and the pronotum length should equal the sum of prozona length and metazona length.
While Ranacris was initially described as apterous [1], this study demonstrates that Ranacris species have vestigial tegmina (Figure 6j and Figure 7a–f), and this finding further confirms the close relationship between Ranacris and Menglacris revealed by molecular evidence in a previous study [11]. Because the sole morphological diagnosis of aptery previously used to define Ranacris no longer exists and the degree of reduction in tegmen from micropterous to apterous condition is not satisfactory for generic distinction in some groups due to the substantial variation even within species [29,30,31], it is reasonable to place the two genera within the same subfamily or tribe, or even to merge Ranacris and Menglacris into one genus after a comprehensive revision.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1You Q.J. Lin R.Z. Description of a new genus and new species of Catantopidae from Guangxi Province, China Entomotaxonomia 19831255258
- 2Cigliano M.M. Braun H. Eades D.C. Otte D. Orthoptera Species File. Taxonomic Database of the World’s Grasshoppers, Locusts, Katydids, Crickets, and Related Insects 2026 Available online: https://orthoptera.speciesfile.org/(accessed on 28 January 2026)
- 3Li H.C. Xia K.L. Fauna Sinica, Insecta, Vol. 43, Orthoptera, Acridoidea, Catantopidae Science Press Beijing, China 2006
- 4You Q.J. Li T.S. Zhang Y.Q. Lin R.Z. Economic Insect Iconography of Guangxi, Phytophage Insects Guangxi Press of Science & Technology Nanning, China 1990
- 5Brunner von Wattenwyl C. Révision du système des Orthoptères et déscription des espèces rapportées par M. Leonardo Fea de Birmanie Ann. Mus. Civ. Stor. Nat. Genova 1893131230
- 6Storozhenko S.Y. A new species of the genus Mesambria Stal, 1878 with notes on the tribe Mesambriini (Orthoptera: Acrididae, Catantopinae)Zootaxa 20184418556510.11646/zootaxa.4418.1.230313601 · doi ↗ · pubmed ↗
- 7Mao B.Y. Ren G.D. Ou X.H. Fauna, Distribution Pattern and Adaptability on Acridoidea from Yunnan China Forestry Publishing House Beijing, China 2011
- 8Zheng Z.M. Lin L.L. Deng W.A. Shi F.M. Two new species of Catantopidae (Orthoptera: Acridoidea) from China Entomotaxonomia 201537177181