Nature-Derived Ferulic Acid Hybrids with Enhanced Antifungal and Antivirulence Activity Against Candida albicans
Dylan Lambert, Celia Lemaire, Louis Camaioni, Muriel Billamboz, Samir Jawhara

TL;DR
Scientists designed new antifungal compounds by modifying ferulic acid, which showed strong activity against Candida albicans without harming human cells.
Contribution
A novel hybridization strategy using ferulic acid to create antifungal molecules with enhanced potency and anti-virulence effects.
Findings
Hybrid molecules like ATF19, ATF20, and MB22 showed antifungal activity with MIC values below 50 µM.
Active compounds caused mitochondrial depolarization, inhibited hyphal growth, and suppressed biofilm formation in C. albicans.
Lead compounds were non-toxic to human cells and improved survival in a C. elegans infection model.
Abstract
The high incidence of Candida albicans infections and the limited efficacy of current antifungal therapies highlight the need for new antifungal agents. In this study, we present a bio-based hybridization strategy aimed at enhancing the antifungal activity of natural product scaffolds, with a particular focus on trans-ferulic acid. A library of twenty-nine hybrid molecules was rationally generated by grafting naturally occurring lipophilic moieties onto either the phenolic or carboxylic acid functions of ferulic acid. The antifungal activity of these molecules was then assessed against C. albicans. While the parent natural compounds exhibited weak activity (MIC > 500 µM), several hybrid derivatives (ATF19, ATF20, and MB22) demonstrated enhanced potency, with MIC values of <50 µM. Esters of the carboxylic acid or phenol group were essential for activity, with the most potent effects…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
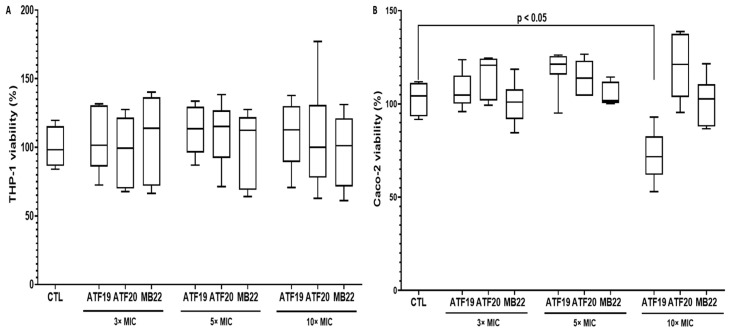 Figure 1
Figure 1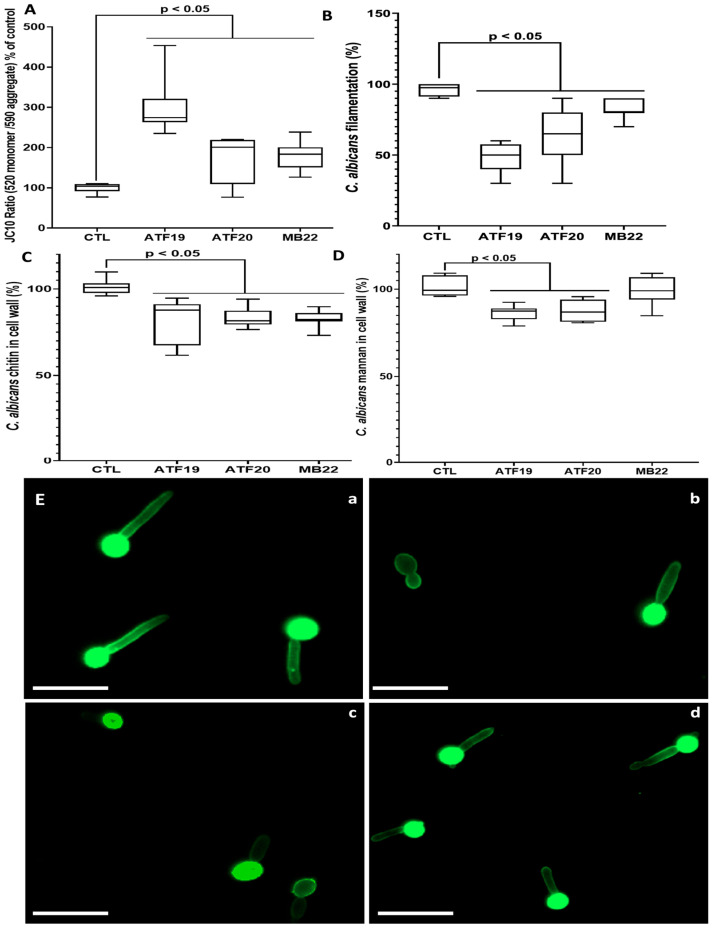 Figure 2
Figure 2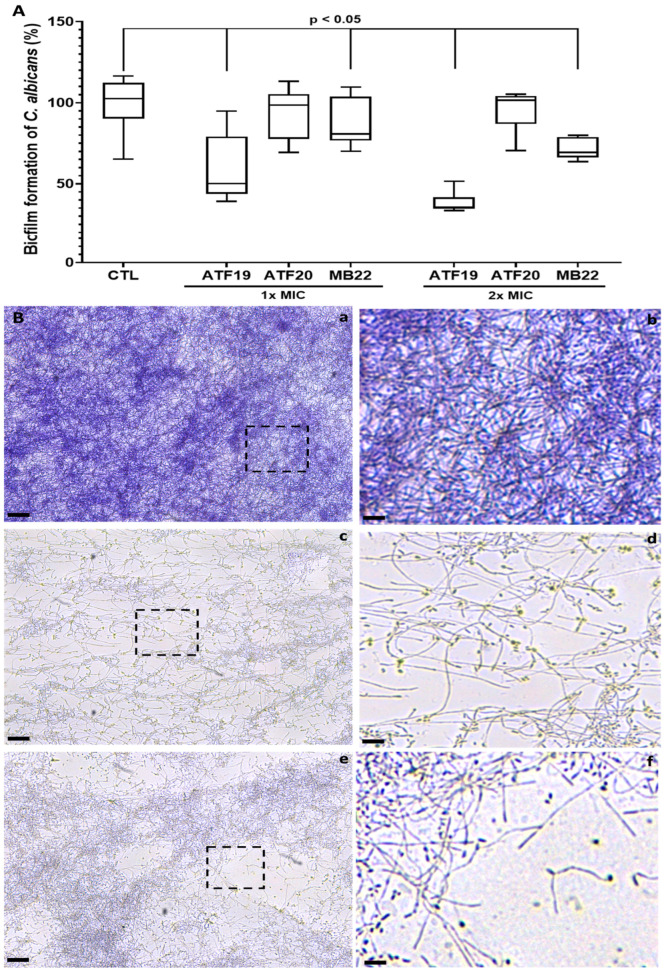 Figure 3
Figure 3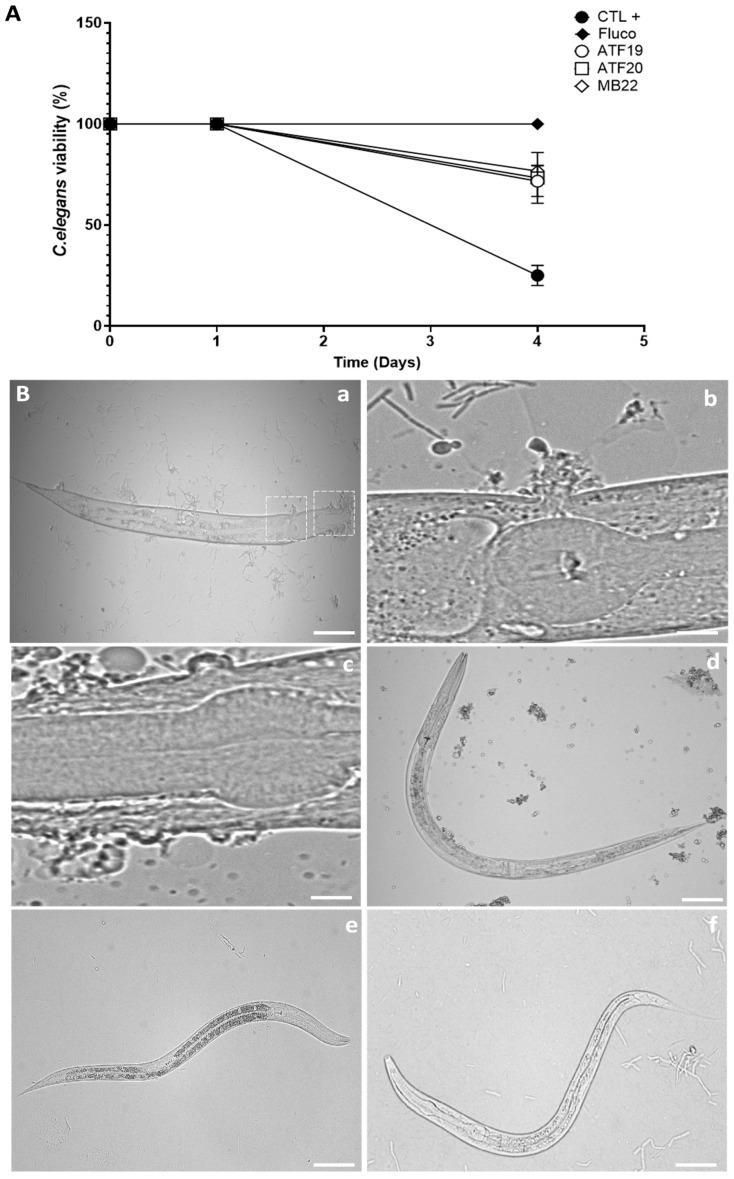 Figure 4
Figure 4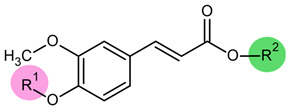 Figure 5
Figure 5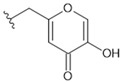 Figure 6
Figure 6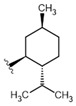 Figure 7
Figure 7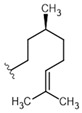 Figure 8
Figure 8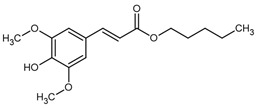 Figure 9
Figure 9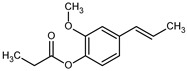 Figure 10
Figure 10- —CNRS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Essential Oils and Antimicrobial Activity · Plant-derived Lignans Synthesis and Bioactivity
1. Introduction
The increasing prevalence of drug-resistant Candida albicans strains highlights the urgent need to expand and strengthen the current therapeutic arsenal with new antifungal agents [1,2,3]. In this context, natural or bio-based compounds are particularly attractive candidates due to their unique combination of biological efficacy and environmental compatibility [4,5]. Unlike purely synthetic molecules, bio-based compounds are generally derived from renewable resources, rapidly biodegrade, and pose a minimal ecological burden. Their sustainable production and reduced persistence in the environment make them particularly advantageous for long-term use, as they mitigate the risks of chemical accumulation and environmental toxicity. Furthermore, their mechanisms of action often reduce the likelihood of resistance development in pathogens, enhancing their appeal as antifungal agents. Phenolic compounds, which are widely studied as secondary metabolites naturally abundant in numerous plant species, are among this diverse class [6]. Beyond their longstanding industrial applications in flavors, spices, cosmetics, and food additives, phenolic compounds are increasingly recognized for their broad antimicrobial properties [6].
Trans-ferulic acid emerges as a highly promising bioactive molecule within this chemical space. It consists of a phenolic aromatic ring that is substituted with a methoxy group in position 3 and is linked to an acrylic acid moiety in position 1. This compound is ubiquitous in the plant kingdom, particularly in the seeds and leaves of fruits, vegetables, and cereals, where it contributes to cell wall integrity by forming covalent bonds between polysaccharides and lignin [7,8]. Its wide availability, low cost, and natural origin make it an attractive and environmentally sustainable resource for further drug development.
Trans-ferulic acid exhibits broad-spectrum antimicrobial activity against both Gram-positive bacteria, such as Bacillus subtilis, Listeria monocytogenes, and Staphylococcus aureus, and Gram-negative bacteria, including Escherichia coli, Pseudomonas aeruginosa, and Salmonella paratyphi, with minimum inhibitory concentrations ranging from tens to hundreds of milligrams per liter [9]. Its moderate hydrophobicity (logP = 1.36) enables it to penetrate microbial membranes, inducing changes in membrane hydrophilicity, surface charge, and cytoplasmic permeability and, ultimately, disrupting cellular homeostasis [9].
Based on this rationale, the present study aimed to design and evaluate hybrid bio-based molecules derived from trans-ferulic acid as potential antifungal candidates against C. albicans. The effects of these molecules on Candida viability, cell wall integrity, filamentation, biofilm formation, and activity against clinical isolates resistant to conventional antifungal drugs were assessed. Additionally, their efficacy was tested using an in vivo Caenorhabditis elegans infection model to provide an integrated evaluation of antifungal potential and host–pathogen interactions. By developing ferulic acid-derived hybrids, this study seeks to advance the creation of sustainable, environmentally friendly, and highly effective antifungal agents that address the urgent challenge of multidrug-resistant C. albicans infections while minimizing ecological impact.
2. Results
Hybrid bio-based molecules derived from trans-ferulic acid were obtained as potential antifungal agents against C. albicans by chemically modifying the acid to introduce lipophilic moieties on either the phenolic group (sub-family F1–F8 and MB22) or on the carboxylic acid function (sub-family F9-ATF20). The lipophilic fragments were selected from naturally occurring compounds and consisted mainly of short- to medium-chain fatty acids. Additional natural molecules, including citronellol, (-)-menthol, and kojic acid, were also incorporated. Similarly, sinapic acid and isoeugenol were derivatized to evaluate the performance of ferulic acid-based analogs. The antifungal activity of all the compounds was evaluated by determining their minimum inhibitory concentrations (MICs) against a susceptible strain of C. albicans. All synthesized molecules and their respective MIC values are summarized in Table 1. In total, twenty-nine compounds were investigated. As shown in Table 1, none of the natural platform molecules (ferulic acid, citronellol, kojic acid, (-)-menthol, sinapic acid, and isoeugenol) exhibited significant antifungal activity, with MIC values of 500 µM or greater. By contrast, coupling these natural scaffolds either together or with fatty acids resulted in the identification of several compounds with markedly improved activity, with MIC values of ≤50 µM. For comparison, the reference antifungal drug fluconazole exhibited an MIC of 0.5 µg/mL under the same conditions. As fluconazole is a fully synthetic molecule, it was used only as a reference control and was not included among the bio-based hybrid molecules evaluated in Table 1.
The derivatization of the phenolic moiety with C2–C9 fatty acids (see Table 1, entries 2–9) provided valuable information. Most of these compounds exhibited improved activity compared with ferulic acid itself, with the exception of the butanoic acid derivative F4. These results suggest that the free phenolic group is not essential for antifungal activity. Furthermore, overall lipophilicity does not appear to be a determining factor, as no clear correlation was observed between activity and the length of the linear alkyl chain. From this series, MB22, bearing a propylate moiety, showed the highest potential with MIC = 10 µM against C. albicans.
By contrast, esterification of the carboxylic acid function of ferulic acid revealed a clear structure–activity relationship. Optimal antifungal activity was observed for derivatives bearing C6–C8 linear aliphatic chains (Table 1, entries 15–17). This trend was further supported by the incorporation of citronellol, which has a branched unsaturated C8 chain. Compound ATF20, resulting from the coupling of ferulic acid and citronellol, was one of the most active molecules identified. Conversely, the introduction of (-)-menthol, a cyclic analog of citronellol, led to a pronounced decrease in activity (Table 1, entry 20; compound F19), indicating a preference for linear lipophilic substituents. This observation is consistent with the weak activity of the kojic acid–ferulic acid hybrid F18.
Compound ATF19, obtained by esterification of sinapic acid with pentan-1-ol, is the direct analog of compound F13 (Table 1, entry 28). ATF19 exhibited a better MIC value, indicating that the additional methoxy group on the aromatic ring of sinapic acid could help improve the antifungal activity. Similarly, compound E2, derived from isoeugenol, can be considered an analog of F2 lacking the carboxylic acid function (Table 1, entry 29). The MIC values of these two compounds were comparable (47 µM vs. 56 µM), suggesting that the carboxylic acid moiety is not critical for activity.
These results demonstrate that chemical coupling of natural product scaffolds is an effective strategy to enhance antifungal activity against C. albicans. Based on their potency and structural features, compounds MB22, ATF19, and ATF20, derived from ferulic acid, sinapic acid, fatty acids, and citronellol, were selected for further investigation.
2.1. Cytotoxicity Assessment of Compounds in Macrophage and Intestinal Epithelial Cell Models
In order to evaluate the potential cytotoxic effects of the compounds ATF19, ATF20, and MB22 in vitro, differentiated THP-1 macrophages were exposed to increasing concentrations corresponding to 3×, 5×, and 10× the MIC. After 24 h of exposure, none of the tested compounds induced detectable cytotoxicity in THP-1 macrophages. Cell viability remained close to 100% across all concentrations tested, indicating a favorable safety profile in this immune cell model (Figure 1A).
The compounds’ cytotoxic potential was also assessed in human intestinal epithelial Caco-2 cells using the same experimental design. Overall, compounds ATF19, ATF20, and MB22 did not significantly affect Caco-2 cell viability at concentrations up to five times the MIC, with survival rates remaining near baseline levels after 24 h of treatment. However, exposure to compound ATF19 at 10× MIC resulted in a significant reduction in cell viability, with only around 75% of cells remaining viable after 24 h. This suggests that compound ATF19 may exert cytotoxic effects on intestinal epithelial cells at supra-therapeutic concentrations (Figure 1B).
2.2. Effects of Compounds on C. albicans Cellular Integrity
Several parameters were examined to determine whether compounds ATF19, ATF20, and MB22 affect the structural integrity of C. albicans. First, mitochondrial membrane potential was assessed using the JC-10 assay following treatment with each compound at 1× MIC. All treatments resulted in a marked and statistically significant increase in the JC-10 monomer-to-aggregate fluorescence ratio after 24 h, indicating mitochondrial depolarization. The observed increase ranged from approximately 200% to 300% relative to untreated controls. Of the tested compounds, ATF19 had the most pronounced effect, with the JC-10 ratio reaching nearly 300% of control values (Figure 2A). In addition to mitochondrial depolarization, we evaluated the production of ROS in C. albicans following treatment with the selected compounds. The results showed that treatment with compound ATF19 did not induce a significant increase in ROS production after 6 h of incubation. In contrast, compounds ATF20 and MB22 led to a noticeable increase in ROS levels under the same conditions (in Supporting Information).
Given the importance of filamentation as a major virulence trait of C. albicans, the impact of the compounds on hyphal development was next evaluated. Treatment with compounds ATF19, ATF20, or MB22 at 1× MIC significantly reduced filamentation after 2 h of incubation. Quantitative analysis revealed a 20–50% decrease in the proportion of filamentous cells compared to the untreated condition. Consistent with mitochondrial data, ATF19 was found to be the most inhibitory, reducing filamentation by around 50% (Figure 2B).
Further investigations of alterations in fungal cell wall composition involved quantifying chitin and mannan levels following exposure to compounds. Treatment with compounds ATF19, ATF20, and MB22 at 1× MIC resulted in a significant reduction in chitin content of between 10% and 20%, compared to untreated C. albicans cells (see Figure 2C). Concurrently, a notable decrease in mannan levels (approximately 15%) was observed following treatment with compounds ATF19 and ATF20. However, MB22 treatment did not result in a statistically significant change in mannan content relative to controls (Figure 2D).
To visually corroborate these quantitative findings, fluorescence and light microscopy analyses were performed after treatment at 1× MIC and staining with GNL. Untreated C. albicans cells exhibited extensive hyphal elongation and strong green fluorescence, reflecting abundant mannan deposition within the cell wall (Figure 2E, panel a). In contrast, compound-treated cells displayed shorter filaments than controls (Figure 2E, panels b–d). Furthermore, treatment with compounds ATF19 and ATF20 resulted in reduced GNL fluorescence intensity, consistent with the observed decrease in mannan content (Figure 2E, panels b and c).
2.3. Impact of Compounds on C. albicans Biofilm Formation
The effects of the compounds on biofilm formation, which is another key virulence factor in C. albicans, were then examined. Yeast cells were treated with compounds ATF19, ATF20, and MB22 at 1× MIC, as well as with compounds ATF19 and MB22 at 2× MIC. Treatment with compounds ATF19 and MB22 at 1× MIC significantly impaired biofilm formation after 24 h, resulting in reductions of approximately 50% and 15%, respectively, compared to untreated controls. Increasing the concentration to 2× MIC further enhanced this inhibitory effect, reducing biofilm biomass by approximately 65% and 30% for compounds ATF19 and MB22, respectively. Overall, compound ATF19 consistently exhibited the strongest antibiofilm activity. Treatment with compound ATF20 at 1× MIC showed a tendency towards reducing biofilms, although this effect was not statistically significant (Figure 3A). Microscopic visualization following crystal violet staining corroborated these quantitative results. Untreated C. albicans formed dense and homogeneous biofilms that covered the entire surface of the well (Figure 3B, panels a and b). In contrast, treatment with compounds ATF19 or MB22 at 2× MIC led to a substantial disruption of biofilm architecture, characterized by reduced staining intensity and the presence of large voids within the biofilm matrix. These images clearly demonstrate the more pronounced antibiofilm effect of compound ATF19 relative to MB22 (Figure 3B, panels c–f).
2.4. Effects of Compounds on C. elegans Survival Following C. albicans Infection
To evaluate the protective effects of the compounds in the context of a host–pathogen interaction, their efficacy was assessed using a C. elegans model of C. albicans infection. Treatment of nematodes with fluconazole at 1× MIC protected the nematodes, resulting in 100% survival at day 4. Infected nematodes were treated with the compounds ATF19, ATF20, or MB22 at a concentration of 1× MIC, and their survival was monitored over time. Treatment with any of the three compounds resulted in a substantial improvement in host survival. Around 75% of the treated nematodes remained alive four days after infection, compared to a survival rate of around 25% in the infected, untreated control group (see Figure 4A). Microscopic examination at the end of the experiment provided further evidence of the protective effects of the compounds. Untreated infected nematodes did not survive and exhibited multiple cuticular perforations, which are consistent with the invasive hyphal penetration and dissemination of C. albicans hyphae (Figure 4B, panels a–c). By contrast, nematodes infected with C. albicans and treated with ATF19, ATF20, or MB22 survived the infection, showing no visible signs of tissue perforation or structural damage (Figure 4B, panels d–f).
3. Discussion
The increasing incidence of C. albicans infections, together with the emergence of resistance to existing antifungal agents and the frequent toxicity issues associated with them, highlights the urgent need for novel, safe, and effective antifungal strategies. Against this backdrop, the present study explores a rational, bio-inspired approach based on the chemical coupling of natural product scaffolds to generate hybrid molecules with enhanced antifungal activity and reduced host toxicity. Our results demonstrate that the strategic derivatization of trans-ferulic acid with naturally occurring lipophilic moieties yields compounds with markedly improved antifungal potency, significant anti-virulence properties, and favorable safety profiles.
A key finding of this study is that individual natural platform molecules (ferulic acid, citronellol, kojic acid, (-)-menthol, sinapic acid, and isoeugenol) exhibited little to no antifungal activity when tested alone. This finding is consistent with previous reports describing the modest antimicrobial effects of phenolic acids and terpenoids, which are often limited by poor membrane penetration or insufficient intracellular targeting [10,11]. It is important to note that natural compounds such as ferulic acid generally display moderate antifungal activity compared to standard agents, such as fluconazole or amphotericin B, and are primarily investigated for their complementary or anti-virulence properties [9].
However, when these scaffolds were chemically coupled either to each other or to fatty acid chains, a clear increase in antifungal potency was observed, with several derivatives displaying MIC values ≤ 50 µM. These results strongly support the concept that hybridization of natural products can unlock synergistic or emergent bioactivities that cannot be achieved with the parent compounds alone [12,13].
Systematic structure–activity relationship analysis revealed that the site of derivatization on ferulic acid plays a critical role in determining its antifungal efficacy [12,13]. Modification of the phenolic hydroxyl group with short- to medium-chain fatty acids generally resulted in better activity, with MB22, the propyl derivative, being the most potent antifungal. Moreover, the lack of correlation between alkyl chain length and activity in this sub-series suggests that simple increases in lipophilicity are insufficient to drive potency, underscoring the importance of more subtle structural features [14].
In contrast, esterification of the carboxylic acid function of ferulic acid yielded a pronounced and well-defined activity profile. Derivatives bearing C6–C8 linear aliphatic chains exhibited optimal antifungal activity, indicating the existence of a favorable balance between hydrophobicity, molecular flexibility, and biological accessibility [15,16]. This optimal chain length likely facilitates efficient interaction with fungal membranes while avoiding excessive hydrophobicity that could impair solubility or selectivity. The high activity of the citronellol-ferulic acid hybrid (ATF20), which incorporates a branched unsaturated C8 chain, further supports this hypothesis and suggests that limited branching and unsaturation can be accommodated without loss of efficacy.
Conversely, the sharp decrease in activity observed upon the incorporation of (-)-menthol, a bulky cyclic analog of citronellol, highlights the importance of molecular shape and conformational flexibility [17,18]. Similarly, the poor performance of the kojic acid–ferulic acid hybrid suggests that excessive polarity or rigid aromatic frameworks are detrimental to antifungal activity [19,20]. Together, these observations suggest that linear or mildly branched lipophilic moieties are preferred, probably because they facilitate productive interactions with fungal lipid bilayers or membrane-associated targets.
Comparative analyses using sinapic acid and isoeugenol derivatives provided additional mechanistic insight. The different MIC values observed for sinapic acid and ferulic acid analogs differing only by an extra methoxy group indicate that increased aromatic substitution could help enhance antifungal potency [21,22]. ATF19 displayed the same activity as F6, which displayed the same overall lipophilicity, suggesting the importance of this balance. Similarly, the comparable activity of isoeugenol-derived compounds lacking a carboxylic acid function suggests that this moiety is not critical for antifungal action. Taken together, these findings argue against a single, highly specific enzymatic target, instead supporting a mechanism involving membrane perturbation and downstream cellular dysfunction.
Consistent with this hypothesis, mechanistic investigations revealed that the most active compounds induce significant changes to key cellular processes in C. albicans. All of the tested lead compounds triggered significant mitochondrial membrane depolarization, indicating the early disruption of fungal energy homeostasis [23,24]. Mitochondrial dysfunction is increasingly recognized as a potent antifungal mechanism, as it can simultaneously impair metabolism, stress responses, and morphogenetic programs. Consistent with this, the compounds also significantly inhibited the yeast-to-hypha transition, a central virulence trait of C. albicans that is required for tissue invasion and immune evasion [25,26]. In addition, ROS production assays showed that ATF19 did not significantly increase ROS levels after 6 h, whereas ATF20 and MB22 induced a marked increase under the same conditions. These results suggest that mitochondrial depolarization induced by ATF19 may occur through a mechanism independent of early ROS overproduction, whereas ATF20 and MB22 appear to induce oxidative stress in C. albicans. This indicates that the relationship between mitochondrial dysfunction and ROS generation may vary depending on the compound and its mode of action, as previously reported in fungal systems, where ROS production can differ in timing and magnitude depending on the stressor [27].
Furthermore, treatment with the lead compounds resulted in measurable reductions in cell wall chitin and mannan content, key structural and immunologically relevant components of the fungal cell wall [28]. Alterations in cell wall architecture not only compromise fungal integrity and sensitize cells to environmental stress and host immune defenses [29,30]. Microscopic analyses corroborated these biochemical findings, revealing shortened hyphae and reduced mannan deposition following exposure to the compounds. Importantly, these effects were observed at concentrations corresponding to the MIC, indicating that anti-virulence activity is closely linked to antifungal efficacy rather than being a secondary consequence of growth inhibition.
The compounds’ ability to interfere with biofilm formation further reinforces their therapeutic potential. Biofilms present a major clinical challenge due to their intrinsic resistance to antifungal drugs and the host immune response. Compound ATF19 exhibits particularly strong antibiofilm activity, suggesting that these hybrid molecules may be effective against both planktonic cells and structured fungal communities [31,32,33]. This property is particularly important for treating device-associated and mucosal candidiasis.
Equally important, the lead compounds exhibited a favorable safety profile in vitro. No cytotoxicity was detected in macrophages or intestinal epithelial cells at concentrations up to five times the MIC, and only limited toxicity was observed at supra-therapeutic concentrations for one compound [34,35]. These results indicate a promising therapeutic window and suggest that selective targeting of fungal cells can be achieved without compromising host cell viability.
Finally, the protective effects observed in the C. elegans infection model provide compelling in vivo support for the antifungal and anti-virulence activities of these compounds [20,36,37]. Treatment significantly improved host survival and prevented hyphal-mediated tissue damage, demonstrating that the compounds retain efficacy in a complex host–pathogen context. The concordance between in vitro mechanistic data and in vivo outcomes strongly supports the translational relevance of this approach.
4. Materials and Methods
4.1. C. albicans Strain and Culture Conditions
C. albicans reference strain SC5314 (wild type; ATCC MYA-2876) was used throughout this study [38]. Yeast cells were routinely maintained on Sabouraud Dextrose Agar (SDA) plates and incubated at 37 °C for 24 h. For experimental assays, colonies were harvested and resuspended in liquid Sabouraud medium, followed by incubation at 37 °C under shaking conditions. Actively growing C. albicans cultures were subsequently used for antifungal susceptibility and phenotypic analyses.
4.2. Chemical Compounds and Cell Culture Models
The synthetic compounds designated ATF19, ATF20, and MB22 were synthesized and provided by JUNIA-HEI (Lille, France). All procedures and descriptions were added as Supporting Information. Compounds were chemically characterized prior to biological evaluation. These compounds were obtained through single-step syntheses followed by purification, in contrast to the multi-step procedures typically required for commercially available antifungal agents. The stability of selected compounds was evaluated in pure water (pH 6.5) at a concentration of 1 g/L. After one week at room temperature, no degradation of these compounds was observed under these conditions. Their antifungal activities were assessed using both in vitro and in vivo experimental models involving C. albicans.
The human monocytic THP-1 cell line (ATCC TIB-202) was cultured in RPMI-1640 medium (Fisher Scientific, Illkirch, France) supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Quentin Fallavier, France) and 1% penicillin–streptomycin (Fisher Scientific) [20,39]. Human intestinal epithelial Caco-2 cells were maintained in DMEM (Fisher Scientific) supplemented with 20% FBS and 1% penicillin–streptomycin. All cell cultures were incubated at 37 °C in a humidified atmosphere containing 5% CO_2_.
4.3. Antifungal Activity and Determination of Minimum Inhibitory Concentration (MIC)
The effects of compounds ATF19, ATF20, and MB22 on C. albicans viability and growth were evaluated using decreasing concentrations. Minimum inhibitory concentrations (MICs) were determined using an Alamar Blue–based assay. Briefly, 5 × 10^3^ yeast cells were dispensed into each well of a transparent 96-well plate (Greiner Bio-One, 655101, Paris, France) containing 90 µL of RPMI medium. Alamar Blue reagent (10 µL; Thermo Fisher Scientific, Illkirch-Graffenstaden, France) was added, followed by the compounds at concentrations ranging from 5 × 10^−3^ M to 5 × 10^−6^ M [20].
MIC values were defined as the lowest compound concentration resulting in ≥95% inhibition of fungal growth. Optical density measurements were recorded at 600 nm using a FLUOstar microplate reader (BMG Labtech, Champigny-sur-Marne, France) at baseline (T_0_) and after 24 h of incubation (T_24_).
4.4. Cytotoxicity Assessment in Macrophage and Intestinal Epithelial Cell Models
THP-1 monocytes were differentiated into macrophages by treatment with phorbol-12-myristate-13-acetate (PMA; 200 ng/mL; Sigma-Aldrich, St. Quentin Fallavier, France) for 72 h at 37 °C in 5% CO_2_. Differentiated THP-1 macrophages and Caco-2 cells were seeded at a density of 1 × 10^5^ cells/mL in transparent 96-well plates and exposed to compounds ATF19, ATF20, or MB22 at concentrations corresponding to 3×, 5×, and 10× the MIC. Cells were incubated for 24 h at 37 °C in 5% CO_2_ [37].
Cell viability was evaluated using the MTT assay [20,40]. Following incubation, 10 µL of MTT reagent (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Bio-Techne, France) was added to each well, and the plates were incubated for an additional 4 h. Formazan crystals were solubilized using 100 µL of MTT detergent solution, and absorbance was measured at 570 nm to assess cellular metabolic activity.
4.5. Assessment of Mitochondrial Membrane Potential in C. albicans
To evaluate mitochondrial function, C. albicans cells (1 × 10^5^ cells/mL) were cultured in RPMI medium and treated with compounds ATF19, ATF20, or MB22 at 1× MIC for 24 h at 37 °C in transparent 96-well plates. Mitochondrial membrane potential was assessed using the JC-10 Mitochondrial Membrane Potential Assay Kit (Abcam, Cambridge, United Kingdom), according to the manufacturer’s instructions [41].
This assay is based on the potential-dependent accumulation of JC-10 dye within mitochondria, where intact mitochondria promote dye aggregation, while depolarized mitochondria favor the monomeric form. Fluorescence signals were measured at 490–525 nm (monomeric form) and 540–590 nm (aggregated form). The ratio of monomeric to aggregated fluorescence was calculated to quantify mitochondrial membrane integrity.
4.6. Effect of Compounds on C. albicans Filamentation
Yeast cells were suspended at a concentration of 1 × 10^4^ cells/mL in RPMI medium supplemented with 10% FBS and incubated in transparent 96-well plates at 37 °C [42]. Cells were treated with compounds ATF19, ATF20, or MB22 at 1× MIC and incubated for 2 h to induce filamentation. Following incubation, treated and untreated C. albicans cells were examined using light microscopy. The proportion of filamentous (hyphal) forms was quantified by counting cells in three randomly selected microscopic fields per well.
4.7. Quantification of Cell Wall Mannans
The C. albicans cell wall is composed primarily of chitin, β-glucans, and mannans. Alterations in mannan content were assessed as an indicator of cell wall integrity. C. albicans cells (1 × 10^5^ cells/mL) were treated with compounds ATF19, ATF20, or MB22 at 1× MIC in 200 µL RPMI medium and incubated for 24 h in black 96-well plates. Cells were then collected by centrifugation and stained with Galanthus nivalis lectin (GNL; Vector Laboratories, Burlingame, CA, USA) for 1 h [42].
After washing, 100 µL of PBS was added to each well, and fluorescence was measured at 450 nm to quantify mannan levels. GNL-stained cells were also visualized by fluorescence microscopy using a Zeiss AxioImager system at the Lille Cell Imaging Platform (BiCels).
4.8. Biofilm Formation Assay
C. albicans cells (5 × 10^3^ cells/mL) were cultured in RPMI medium supplemented with 20% FBS at 37 °C for 6 h to allow initial adhesion. Cells were subsequently treated with compounds ATF19, ATF20, or MB22 at 1× MIC and incubated for an additional 24 h in transparent 96-well plates. Wells were gently washed three times with PBS to remove non-adherent cells [40]. Biofilms were stained with crystal violet for 30 min (Honeywell Fluka, Sevrey, France), followed by ethanol destaining to remove excess dye. Biofilm biomass was quantified by measuring absorbance at 550 nm. Representative biofilm images were acquired using a Leica DMI8 microscope at the Lille imaging facility.
4.9. C. elegans Infection Model
C. albicans cells were cultured in liquid Sabouraud medium at 37 °C for 24 h. The wild-type N2 strain of C. elegans was maintained on nematode growth medium seeded with Escherichia coli OP50 at 20 °C. Worm populations were synchronized prior to infection [20,42].
C. albicans cells were plated on Brain Heart Infusion (BHI) agar supplemented with amikacin (45 µg/mL; Mylan, Paris, France) for 30 min. Synchronized nematodes were washed extensively with M9 buffer containing 90 µg/mL amikacin to remove E. coli and promote ingestion of C. albicans. Worms were then exposed to C. albicans on BHI plates for 6 h at 20 °C.
Following infection, nematodes were collected, counted, and distributed (10–20 worms per well) into transparent 12-well plates containing M9 buffer supplemented with 5% BHI and treated with compounds ATF19, ATF20, or MB22 at 1× MIC. Worm survival was monitored daily for 5 days post-infection. At the end of the experiment, representative images were acquired using a Leica DMI8 microscope at the Lille Cell Imaging Platform.
4.10. Statistical Analysis
All statistical analyses were performed using GraphPad Prism software (GraphPad Software, version 11.0.0, La Jolla, CA, USA). Differences between experimental groups were evaluated using the non-parametric Mann–Whitney–Wilcoxon test. Statistical significance was defined as p < 0.05, with additional thresholds at p < 0.01 and p < 0.001.
5. Conclusions
In conclusion, this study demonstrates that the chemical hybridization of natural product scaffolds is an effective strategy for generating antifungal agents with enhanced potency, antivirulence activity, and low host toxicity. Among the compounds investigated, the ferulic acid–citronellol hybrid ATF20, the sinapic derivative ATF19, and the propylate MB22 emerge as promising leads. Their multifaceted mode of action, which combines growth inhibition, virulence suppression, and host protection, makes them attractive candidates for further preclinical development and mechanistic exploration.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jawhara S. How Gut Bacterial Dysbiosis Can Promote Candida albicans Overgrowth during Colonic Inflammation Microorganisms 202210101410.3390/microorganisms 1005101435630457 PMC 9147621 · doi ↗ · pubmed ↗
- 2Poulain D. Sendid B. Standaert-Vitse A. Fradin C. Jouault T. Jawhara S. Colombel J.F. Yeasts: Neglected pathogens Dig. Dis.20092710411010.1159/00026812920203505 · doi ↗ · pubmed ↗
- 3Jawhara S. How Fungal Glycans Modulate Platelet Activation via Toll-Like Receptors Contributing to the Escape of Candida albicans from the Immune Response Antibiotics 2020938510.3390/antibiotics 907038532645848 PMC 7399910 · doi ↗ · pubmed ↗
- 4Li R. Yang X. Dan W. Dai J. Natural product-derived antifungals against Candida albicans: Chemical diversity and mechanisms of action Bioorg. Med. Chem.202613211843510.1016/j.bmc.2025.11843541082851 · doi ↗ · pubmed ↗
- 5Dantas T.D.S. Machado J.C.B. Ferreira M.R.A. Soares L.A.L. Bioactive Plant Compounds as Alternatives Against Antifungal Resistance in the Candida Strains Pharmaceutics 20251768710.3390/pharmaceutics 1706068740574000 PMC 12196126 · doi ↗ · pubmed ↗
- 6Mekinic I.G. Skroza D. Ljubenkov I. Katalinic V. Simat V. Antioxidant and Antimicrobial Potential of Phenolic Metabolites from Traditionally Used Mediterranean Herbs and Spices Foods 2019857910.3390/foods 811057931731762 PMC 6915508 · doi ↗ · pubmed ↗
- 7Graf E. Antioxidant potential of ferulic acid Free Radic. Biol. Med.19921343544810.1016/0891-5849(92)90184-I 1398220 · doi ↗ · pubmed ↗
- 8Iiyama K. Lam T.B.-T. Stone B.A. Covalent cross-links in the cell wall Plant Physiol.199410431510.1104/pp.104.2.31512232082 PMC 159201 · doi ↗ · pubmed ↗