Study on the Role and Mechanism of TOPORS in Regulating Aortic Dissection by Mediating SUMOylation
Yuan Hu, Luxi Yang, Wenjun Zhou, Hao Chen, Yuanmin Li, Bing Song, Cuntao Yu

TL;DR
This study explores how TOPORS regulates aortic dissection through SUMOylation, showing that TOPORS promotes inflammation and cell death, worsening the condition.
Contribution
The study reveals a novel role of TOPORS in aortic dissection via SUMOylation of p53, linking it to inflammation and cell apoptosis.
Findings
TOPORS inhibition reduces aortic dilation and elastic fiber degradation in AD models.
TOPORS suppression decreases inflammatory cytokines and promotes PI3K/AKT phosphorylation.
TOPORS-mediated SUMOylation of p53 contributes to vascular smooth muscle cell apoptosis and AD progression.
Abstract
Aortic dissection (AD) is a fatal acute cardiovascular emergency. SUMOylation participates in cell proliferation, apoptosis, and inflammation, but its role in AD, especially via TOPORS, remains unclear. This study investigates how TOPORS regulates AD pathogenesis through SUMOylation. AD and normal aortic samples were collected to detect TOPORS expression. AD mouse and VSMCs models were constructed to assess TOPORS depletion and overexpression effects on AD progression. In AD aortic tissues, TOPORS expression was upregulated, while tripartite motif containing 27 (TRIM27) and Sentrin-specific protease 6 (SENP6) expression showed no significant change. In vivo and in vitro experiments demonstrated that inhibition of TOPORS alleviated aortic dilation and elastic fiber degradation. TOPORS knockout suppressed the secretion of inflammatory cytokines (TNF-α, IL-1β, IL-6, and IFN-α), promoted…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
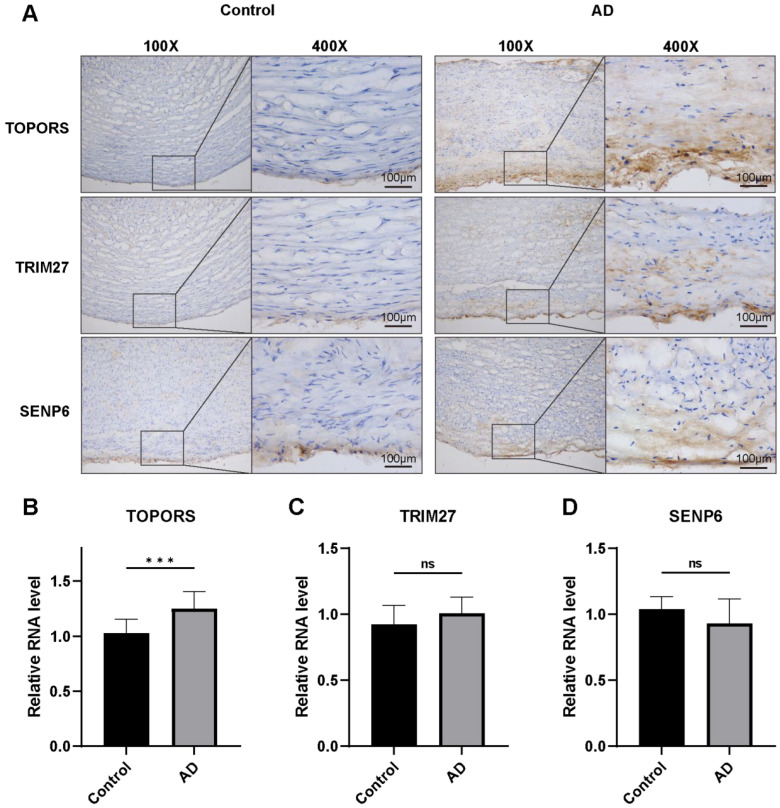 Figure 1
Figure 1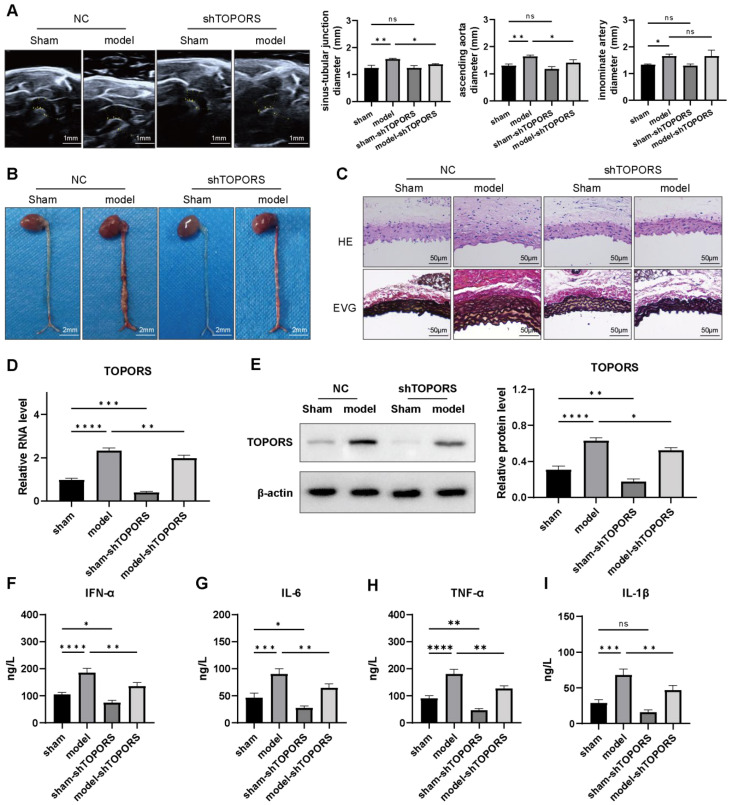 Figure 2
Figure 2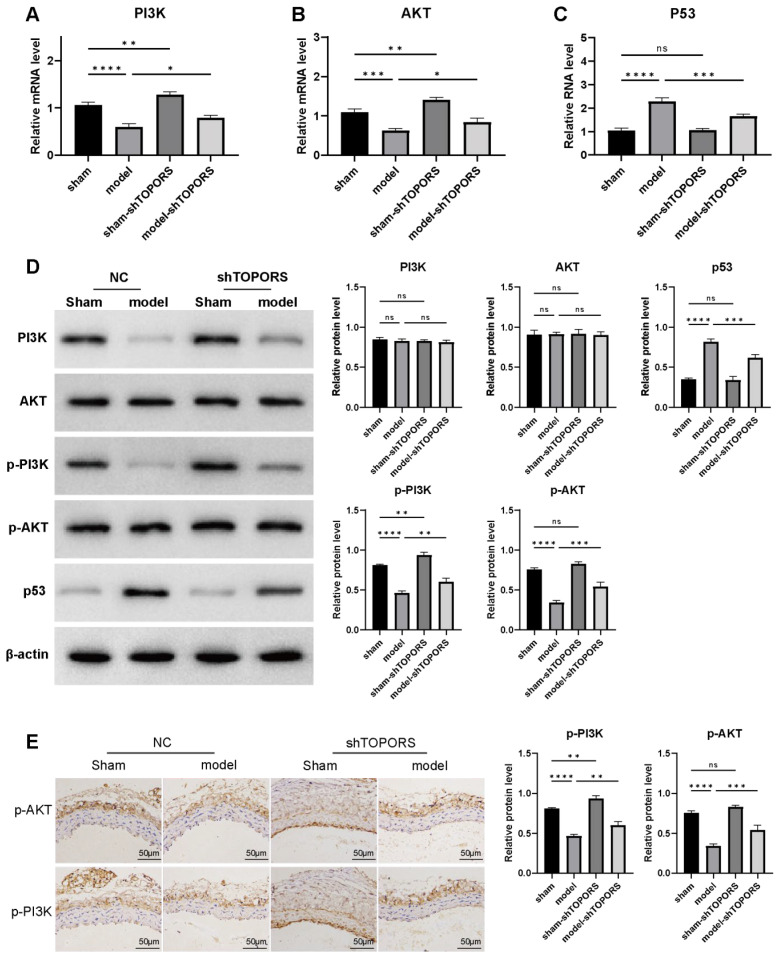 Figure 3
Figure 3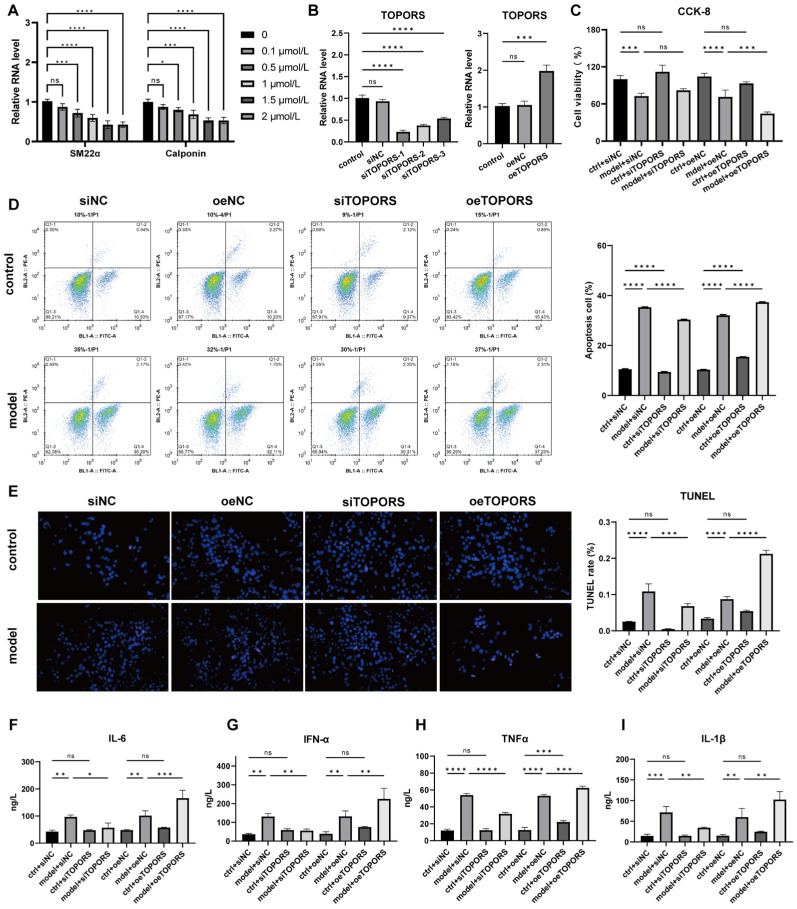 Figure 4
Figure 4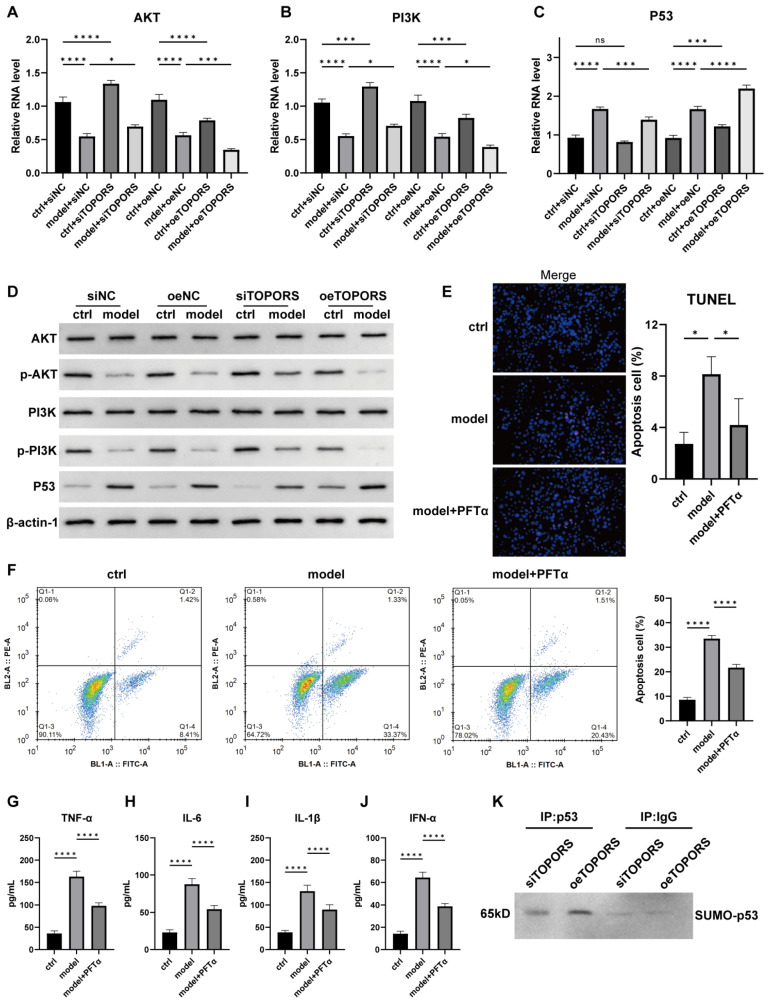 Figure 5
Figure 5- —Natural Science Foundation of Gansu Province, China
- —The First Hospital of Lanzhou University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsConnective tissue disorders research · Aortic Disease and Treatment Approaches · Ubiquitin and proteasome pathways
1. Introduction
Aortic dissection (AD), a life-threatening acute cardiovascular disorder, is characterized by blood infiltration into the aortic media following an intimal tear, leading to separation of the true and false lumens. Its abrupt onset and rapid progression contribute to a 30% 24 h mortality rate without timely intervention, with 1-week mortality exceeding 50%, highlighting the urgency to unravel its underlying pathogenesis for targeted therapy development [1].
In recent years, significant progress has been made in identifying genetic and post-translational modification (PTM) drivers of AD. Genetic studies have implicated genes such as CCDC80, ACKR1, and Ncf1 in AD pathogenesis: CCDC80 maintains vascular smooth muscle cell (VSMC) contractile phenotype to protect against AD rupture [2]; ACKR1-expressing endothelial cells promote AD by modulating macrophage behavior [3]; and Ncf1 knockout in VSMCs exacerbates angiotensin II-induced AD via STING pathway activation [4]. Additionally, lactylation, a novel PTM, has been linked to AD progression: mitochondrial ATP synthase subunit alpha (ATP5F1A) lactylation impairs mitochondrial function [5], while histone H4K16 lactylation drives metabolic remodeling in VSMCs to promote aortic aneurysm and dissection [6]. These findings have advanced risk stratification and therapeutic target identification, but critical gaps remain in understanding the complex regulatory networks governing AD pathogenesis.
SUMOylation is a dynamic and reversible post-translational modification mediated by small ubiquitin-like modifiers (SUMOs). It orchestrates core cellular processes by modulating target protein activity, subcellular localization, and protein–protein interactions, including transcriptional regulation, DNA repair, cell proliferation, apoptosis, and inflammation [7,8,9,10,11]. Its molecular cascade involves SUMO precursor maturation, activation by E1 enzymes, conjugation via the sole E2 enzyme, ligation to substrates by E3 ligases, and deSUMOylation to ensure reversibility. Emerging evidence links dysregulated SUMOylation to cardiovascular pathologies [12,13]. For example, SUMOylation of transcription factor HEY1 and fibroblast growth factor receptor 1 (FGFR1) modulates angiogenesis by balancing of endothelial angiogenic signals [14,15]; SUMOylation of serum response factor (SRF) drives vascular smooth muscle cell (VSMC) phenotypic switching, promoting vascular remodeling and neointimal hyperplasia [16]. However, research on SUMOylation in AD remains extremely limited. To date, only TRIM39, a SUMO E3 ligas, has been reported to regulate AD progression through SUMOylation of estrogen receptor alpha (ESR1) [17]. This scarcity of data indicates that the SUMOylation regulatory network in AD pathogenesis is largely uncharacterized.
The p53 and PI3K-AKT pathways are well-characterized regulators of VSMC survival, proliferation, and phenotypic transition—key events in AD progression. Notably, both pathways are tightly modulated by SUMOylation: UBC9-mediated p53 SUMOylation drives cellular senescence and exacerbates tissue injury [18]; SUMOylation of the class IA PI3K/AKT pathway affects cell-cycle dynamics and anti-apoptotic signaling [19]. Given the critical roles of p53 and PI3K-AKT in AD, their SUMOylation status is likely a key determinant of AD pathogenesis, yet the upstream regulators of this process in the aortic wall remain unknown.
TOPORS (topoisomerase I-binding RING finger protein) is a dual-function enzyme with both SUMO E3 ligase and ubiquitin E3 ligase activity, belonging to the SUMO-targeted ubiquitin ligase (STUbL) family. Emerging studies have established its role in regulating p53 stability and transcriptional activity through SUMOylation [20,21], with implications in tumorigenesis and neurodegenerative diseases. However, its functional relevance in cardiovascular pathologies, particularly aortic wall homeostasis, remains entirely unexplored. This represents a critical knowledge gap given the established role of SUMOylation in vascular smooth muscle cell (VSMC) phenotypic switching and extracellular matrix remodeling—core processes in aortic dissection (AD) pathogenesis. Against this backdrop, the present study focuses on TOPORS-mediated SUMOylation, aiming to systematically dissect its molecular mechanisms in AD pathogenesis, unravel the SUMOylation regulatory network in AD, and provide a theoretical basis for developing novel AD therapeutic targets.
2. Materials and Methods
2.1. Clinical Samples
The processing of human samples strictly adhered to the experimental protocol approved by the Ethics Committee of the First Hospital of Lanzhou University (Ethics Approval No.: LDYYLL2025-33). All participants provided written informed consent, and the entire study complied with the ethical requirements of the Declaration of Helsinki. Intraoperatively resected ascending aortic wall tissues were collected from 10 patients diagnosed with aortic dissection (AD) (5 males and 5 females). For the control group, normal ascending aortic tissues were obtained from 10 control patients without aortic dissection (5 males and 5 females), including those who underwent concurrent heart transplantation or aortic valve replacement with confirmed normal aortic diameter. Within 30 min post-explantation, all tissue samples were rinsed with pre-chilled PBS to eliminate residual blood, promptly snap-frozen in liquid nitrogen, and then stored at −80 °C for subsequent experimental procedures.
2.2. Mouse AD Model
Three-week-old male C57BL/6J mice, weighing 10 to 15 g, were acquired from SPF (Beijing) Biotechnology Co., Ltd. (Beijing, China). The experiment utilized a total of 40 mice. All mice were kept in a specific pathogen-free (SPF) barrier facility with a 12 h light/dark cycle, temperature ranging from 22 to 24 °C, and humidity between 50% and 70%. Animal experiments strictly adhered to the guidelines of the Animal Ethics Committee of Hebei Kangtai Medical Laboratory Service Co., Ltd. (Approval No.: MDL2024-07-03-01, Langfang, China) and complied with the Guide for the Care and Use of Laboratory Animals by the National Institutes of Health. Before surgery, the mice were randomly divided into four groups (10 mice in each group): sham operation (sham), AD model (model), sham-shTOPORS, and model-shTOPORS. The mouse AD model was induced by β-aminopropionitrile (BAPN): Alzet osmotic pumps (Alzet 2004, Cupertino, CA, USA) containing BAPN (1 g/kg/24 h, Sigma, Burlington, MA, USA) were implanted in the mice for continuous administration over 28 days. To ensure consistency and minimize variability, surgical procedures and drug administrations were carried out by the same experienced personnel. A small animal echocardiograph (D6LAB, VINNO, Suzhou, China) was employed to measure the diameters of the thoracic aortic root, ascending aorta, aortic arch, descending aorta, and abdominal aorta. At the experimental endpoint, mice were euthanized under deep anesthesia; aortic tissues were dissected, fixed in 4% paraformaldehyde for 24 h, and embedded in paraffin for 5 μm sectioning. All surgical interventions were performed under deep anesthesia induced by intraperitoneal injection of sodium pentobarbital (1% w/v in sterile saline, 50 mg/kg body weight). Anesthesia depth was confirmed by the absence of corneal reflex and a negative response to the toe pinch test.
2.3. Cell Culture and In Vitro Model
Human WSMCs were purchased from Wuhan Procell Life Science & Technology Co., Ltd. (Wuhan, China) and cultured in Smooth Muscle Cell Medium (Sciencell, San Diego, CA, USA) supplemented with 1% penicillin-streptomycin mixture. Cells were maintained in a humidified incubator at 37 °C with 5% CO_2_. To ensure cell purity and authenticity, routine mycoplasma detection and short tandem repeat (STR) profiling were performed, and no mycoplasma contamination was observed in any cell cultures.
For experimental grouping (n = 3), VSMCs were divided into eight groups: control group transfected with negative control siRNA (ctrl + siNC), AD model group transfected with negative control siRNA (model + siNC), control group transfected with negative control overexpression vector (ctrl + oeNC), AD model group transfected with negative control overexpression vector (model + oeNC), control group transfected with TOPORS siRNA (ctrl + siTOPORS), AD model group transfected with TOPORS siRNA (model + siTOPORS), control group transfected with TOPORS overexpression vector (ctrl + oeTOPORS), and AD model group transfected with TOPORS overexpression vector (model + oeTOPORS).
To establish the AD cell model, VSMCs were treated with angiotensin II (Ang II). To determine the optimal Ang II concentration for inducing VSMC phenotypic changes associated with AD, cells were exposed to different concentrations of Ang II (0, 0.5, 1.0, 1.5, and 2.0 μmol/L) for 48 h, followed by detection of calponin and SM22α expression levels via qRT-PCR.
For genetic manipulation, the TOPORS overexpression vector was constructed by synthesizing mouse TOPORS cDNA (Gene ID: 10210) and subcloning it into the pLVX plasmid vector (General Biosystems, Anhui, China). Small interfering RNAs (siRNAs) targeting TOPORS (sequences: 5′-CAUUGUGAUUCUAGUACAATT-3′; 5′-CGAUUUCGCUACCGUACAATT-3′; 5′-CUGUCUAGUAACAGAUCAATT-3′) and negative control siRNA (sequence: 5′-UUCUCCGAACGUGUCACGUTT-3′) were designed and synthesized by RiboBio Co., Ltd. (Guangzhou, China). Transfections were performed using Lipofectamine 2000 reagent (L3000015, Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions.
Furthermore, to investigate the regulatory role of the p53 signaling pathway in AD progression, VSMCs were treated with the p53 inhibitor PFTα at a final concentration of 10 μM.
2.4. Immunohistochemistry (IHC)
Aortic tissues from humans and mice, stored at −80 °C, were embedded in optimal cutting temperature (OCT) compound (C0171A, Beyotime, Shanghai, China) to maintain morphology. Using a cryostat microtome (CM1950, Leica, Walldorf, Germany), they were sectioned into 4 μm slices, mounted on poly-L-lysine-coated slides, and air-dried. The sections were fixed with 4% paraformaldehyde for 10 min, rinsed with PBS, and blocked with 0.3% Triton X-100 (containing 5% BSA) for 1 h at room temperature. Primary antibodies (Affinity, Cincinnati, OH, USA) were incubated overnight (12–16 h) at 4 °C: SENP6 (AF0277, 1:500), TOPORS (DF2475, 1:500), TRIM27 (DF12782, 1:500), AKT (AF0836, 1:500), p-AKT (AF0016, 1:500), p-PI3K (AF3242, 1:500), PI3K (AF6241, 1:500), and p53 (AF0879, 1:500). These primary antibodies are suitable for both human and mouse tissues. After washing with PBS, the sections were incubated with HRP-conjugated goat anti-rabbit secondary antibody (S0001, 1:3000, Affinity) in 5% BSA for 30 min at room temperature. The sections were stained with DAB (C1005, Beyotime, Shanghai, China) for 5–8 min, counterstained with hematoxylin for 30 s, dehydrated in graded ethanol, cleared in xylene, and mounted with neutral balsam. Images were captured using a light microscope (DM3000, Leica, Wetzlar, Germany) and analyzed using Image-Pro Plus 6.0; mean optical density (IOD/Area) was quantified from 5 fields per group.
2.5. Hematoxylin-Eosin (HE) Staining
Paraffin sections of mouse aortic tissues were dewaxed in xylene, then rehydrated through graded ethanol (100%, 95%, 80%, 70%, 2 min each) to distilled water. They were stained with hematoxylin solution from an HE Stain Kit (G1120, Solarbio, Beijing, China) for 10 min at room temperature, rinsed with tap water to remove excess stain, differentiated with kit-provided differentiating solution for 60 s, then rinsed with tap water for 15 min to blue nuclei. They were then stained with eosin solution from the same kit for 2 min, dehydrated through graded ethanol (70%, 80%, 95%, 100%, 2 min each), cleared in xylene (twice, 5 min each), mounted with neutral balsam, air-dried, and observed under a light microscope.
2.6. Elastica Van Gieson (EVG) Staining
Dewaxed sections were stained with Verhoeff staining solution (DC0057, Leagene, Beijing, China) for 30 min, rinsed with distilled water, differentiated with differentiating solution until elastic fibers turned black and background gray, then rinsed under running water for 5 min. They were deiodinated with 95% ethanol twice and counterstained with Orange G for 1 min, with the excess stain blotted. They were dehydrated in absolute ethanol twice (30 s each), cleared in xylene twice (5 min each), mounted with neutral balsam, and examined microscopically after drying.
2.7. Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from aortic tissue samples and VSMCs using TRIpure Reagent (RN0102, Aidlab Biotechnologies Co., Ltd., Beijing, China). RNA concentration and purity were determined with a UNano-1000 spectrophotometer (YoMim, Hangzhou, China), and RNA integrity was verified via 1% agarose gel electrophoresis. RNA was reverse-transcribed into cDNA using ExonScript RT SuperMix (A502-02, EXONGEN, Chengdu, China). qRT-PCR amplification was performed with SYBR Select Master Mix (4472920, Invitrogen, Carlsbad, CA, USA) on a Q2000B qPCR instrument (LongGene, Hangzhou, China). β-actin served as the internal reference, and the relative expression levels of TOPORS, SENP6, TRIM27, PI3K, AKT, and p53 were calculated using the comparative cycle threshold (Ct) method. The gene primer information is listed in Table 1.
2.8. Western Blotting
Aortic tissue samples and VSMCs were lysed with RIPA lysis buffer (P0013C, Beyotime, Shanghai, China) on ice for 30 min. After lysis, the samples were centrifuged at 12,000× g and 4 °C for 15 min to collect supernatants, and the protein concentration was determined using a BCA kit (P0012, Beyotime, Shanghai, China). For protein denaturation, 50 μg of protein was mixed with loading buffer and heated at 95 °C for 5 min. The denatured protein was then loaded onto 10% SDS-PAGE gels, and electrophoresis was performed at 80 V until the samples entered the separating gel, after which the voltage was adjusted to 120 V. Following electrophoresis, proteins were transferred to PVDF membranes (Millipore, Boston, MA, USA) at a constant current of 300 mA in an ice bath. The membranes were blocked with 5% non-fat milk in TBST for 1 h, then incubated with primary antibodies at 4 °C overnight, followed by three washes with TBST (10 min each). Subsequently, the membranes were incubated with secondary antibodies for 1 h and washed again. Protein bands were detected using an ECL system (ChemiScope6100, CLINX, Shanghai, China), and grayscale analysis was performed with ImageJ software (v1.8.0, NIH, Bethesda, MD, USA). The primary antibodies used were SENP6 (AF0277, 1:500), TOPORS (DF2475, 1:500), TRIM27 (DF12782, 1:500), AKT (AF0836, 1:500), p-AKT (AF0016, 1:500), p-PI3K (AF3242, 1:500), PI3K (AF6241, 1:500), and p53 (AF0879, 1:500); the secondary antibody was goat anti-rabbit IgG-HRP (S0001, 1:3000), all obtained from Affinity Biosciences.
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of inflammatory factors, including IFN-α, IL-6, TNF-α, and IL-1β, were measured using ELISA. Specifically, the following ELISA kits were employed: Mouse TNF-α Elisa Kit (H052-1-2), Mouse IFN-α Elisa Kit (H023-1-1), Mouse IL-6 Elisa Kit (H007-1-2), and Mouse IL-1β Elisa Kit (H002-1-2), all purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). All experimental procedures were performed strictly according to the manufacturer’s instructions to ensure the accuracy and reproducibility of the assay results.
2.10. Cell Proliferation Assay
Cell proliferation was evaluated using the Cell Counting Kit-8 (CCK-8) assay and 5-ethynyl-2′-deoxyuridine (EdU) incorporation assay. For the CCK-8 assay, VSMCs were seeded in 96-well plates at a density of 1 × 10^4^ cells/well and cultured for 12 h. Afterward, 10 µL of CCK-8 reagent (Beyotime, Shanghai, China) was added to each well, followed by incubation for 1 h. Absorbance at 450 nm was measured using a microplate reader (Model 550, BIORAD, Hercules, CA, USA), where absorbance values correlated positively with cell proliferative activity.
For the EdU assay, VSMCs were seeded in 24-well plates at 5.0 × 10^4^ cells/well, incubated with 10 µmol/L EdU solution (Beyotime, Shanghai, China) at 37 °C for 2 h, then fixed with 4% paraformaldehyde (PFA) at 4 °C for 15 min. Subsequently, 100 µL of Click reaction solution was added, and nuclei were stained with Hoechst 33,342. Fluorescent signals were visualized under a fluorescence microscope to assess cell proliferation.
2.11. Co-Immunoprecipitation
VSMCs were pretreated with 20 mM NEM (N-Ethylmaleimide) in medium for 20 min at 37 °C to block deSUMOylation, then lysed on ice for 30 min in RIPA buffer containing 1% SDS and 20 mM NEM. After centrifugation (14,000 rpm, 15 min, 4 °C), 500 µg supernatant protein was incubated with 2 µg anti-SUMO1 antibody (CST, Cat#4930S) at 4 °C overnight with rotation. Pre-washed Protein A/G beads (30 µL) were added for 3 h at 4 °C, then pelleted (3000 g, 5 min, 4 °C). Beads were washed 4× with NEM-containing RIPA buffer, boiled with 2× SDS loading buffer for 8 min, and subjected to SDS-PAGE. Proteins were transferred to PVDF membrane, blocked, and incubated with anti-p53 antibody (AF0879, 1:500) followed by HRP-conjugated secondary antibody (S0001, 1:3000). Signals were detected via ECL chemiluminescence.
2.12. Cell Apoptosis Assay
Cell apoptosis was evaluated using TUNEL staining and Annexin V/PI flow cytometry. For TUNEL staining, cells were washed twice with pre-cooled PBS to remove residual medium, fixed with 4% PFA for 15 min, and permeabilized with 0.25% Triton X-100 for 10 min. Samples were then incubated with TUNEL reagent (In Situ Cell Death Detection Kit, Beyotime, Shanghai, China) at 37 °C for 50 min, followed by nuclear staining with Hoechst 33342 for 10 min. Fluorescent signals were observed under a Leica fluorescence microscope (Wetzlar, Germany), and the apoptosis rate was calculated as the ratio of TUNEL-positive apoptotic cells to total DAPI-labeled cells.
For the Annexin V/PI assay, cells were resuspended in Binding Buffer at 1 × 10^6^ cells/mL; 100 µL cell suspension was mixed with 5 µL Annexin V/FITC in 5 mL flow tubes, incubated at room temperature for 5 min in the dark, then stained with 10 µL of 20 µg/mL propidium iodide (PI) solution. After adding 400 µL PBS, samples were immediately analyzed using a flow cytometer (BeamCyte-1026, BENM DIAG, Changzhou, China) with the Annexin V/PI Apoptosis Detection Kit (V13242, Invitrogen, Carlsbad, CA, USA).
2.13. Statistical Analysis
GraphPad Prism 9 (San Diego, CA, USA) was utilized for conducting statistical analyses. Student’s t-test was employed to analyze comparisons between two groups, and one-way ANOVA followed by Tukey’s post hoc test was used for multiple groups. Statistical significance was set at p < 0.05. Unless otherwise stated, data are shown as mean ± standard deviation (SD).
3. Results
3.1. TOPORS Expression Was Significantly Increased in Human AD
Human aortic dissection tissues and normal aortic tissue samples were carefully collected and subsequently divided into two distinct groups for comparative analysis: the control group, which consisted of normal aortic tissues, and the aortic dissection (AD) group, which included tissues affected by the dissection. To investigate the protein expression levels of SENP6, TOPORS, and TRIM27, immunohistochemistry (IHC) was employed as a reliable technique. Concurrently, qRT-PCR was utilized to measure the mRNA transcription levels of these same genes, providing a comprehensive assessment of both protein and genetic expression.
Upon conducting statistical analysis, it was observed that when compared to the control group, the protein and mRNA expression levels of TOPORS in the AD group exhibited a significant increase, indicating a potential role of this gene in the pathogenesis of aortic dissection (Figure 1A,B). In contrast, for TRIM27 and SENP6, although there were changes in their expression levels, these alterations did not reach statistical significance, suggesting that their involvement in aortic dissection may be less pronounced or require further investigation.
3.2. Inhibition of TOPORS Ameliorates AD in Mice
To thoroughly investigate the pivotal role of TOPORS in the pathogenesis of AD, we established an AD mouse model by inducing the condition through the administration BAPN. Subsequently, we evaluated the effects of TOPORS knockdown on this model. All mice in the sham and sham-shTOPORS groups survived, with no aortic dissection found in post-mortem examination. In the BAPN-induced model group, two mice died from ruptured aortic dissection. Of the eight surviving mice, seven developed aortic dissection, and five of them had rupture. In the model-shTOPORS group, there was no mortality during the experiment. Post-mortem analysis showed six mice developed aortic dissection and three of them had rupture.
Utilizing ultrasound imaging to measure aortic dilation on the 14th day post-model induction, we observed that the aortic diameter in the model group was significantly larger compared to the sham control group. However, this pronounced aortic dilation was markedly reduced in the model group treated with shRNA targeting TOPORS (model + shTOPORS group), as depicted in Figure 2A. Gross anatomical examinations further corroborated these findings. The model group exhibited notable thoracic aortic aneurysmal enlargement, which was filled with erythrocytes, indicating severe structural damage. In stark contrast, the inhibition of TOPORS led to a substantial alleviation of this aortic structural deterioration, as evidenced in Figure 2B.
To delve deeper into the histopathological changes, we conducted HE and EVG staining. These analyses revealed that the AD model mice displayed a disorganized aortic architecture characterized by severe degradation of medial elastic fibers. Conversely, the knockdown of TOPORS preserved the integrity of the elastic fiber layers and maintained the closed circular structure of the thoracic aorta, as illustrated in Figure 2C. Molecular validation through qRT-PCR and Western blotting assays confirmed that both the mRNA and protein levels of TOPORS were significantly elevated in the model group. However, these levels were markedly reduced following the silencing of TOPORS, as shown in Figure 2D,E. Given the critical role of inflammation in the progression of AD, we performed cytokine profiling using ELISA. The results indicated that the model mice exhibited elevated levels of pro-inflammatory cytokines, including *IFN-*α, IL-6, TNF-α, IL-1β. Importantly, the inhibition of TOPORS significantly reversed these elevated cytokine levels, as demonstrated in Figure 2F–I. Collectively, these comprehensive data suggest that the inhibition of TOPORS effectively mitigates the formation of AD by restraining aortic dilation, preserving the structural integrity of the vasculature, and reducing the inflammatory responses associated with AD progression.
3.3. Effects of TOPORS Inhibition on PI3K/AKT and p53 Signaling Pathways
qRT-PCR and Western blot analyses were performed to investigate the regulatory role of TOPORS in signaling pathway dysregulation during AD pathogenesis. Compared with the sham group, the AD model mice exhibited significant upregulation of both mRNA and protein expression levels of p53. For the PI3K/AKT pathway, mRNA expression of PI3K and AKT was markedly decreased, whereas their protein levels remained unchanged (Figure 3A–D). Notably, the phosphorylated active forms (p-PI3K and p-AKT) showed significant downregulation (Figure 3D,E). Furthermore, targeted inhibition of TOPORS gene expression in the aortic dissection model partially reversed these pathological molecular alterations: p53 upregulation was attenuated, and the reduction in p-PI3K/p-AKT levels was alleviated. These findings collectively suggest that TOPORS may contribute to the initiation and progression of aortic dissection by modulating the PI3K/AKT signaling pathway (via regulation of kinase phosphorylation) and the p53 pathway.
3.4. TOPORS Regulates Aortic Dissection-like Phenotypes in VSMCs
To investigate the role of TOPORS in AD pathogenesis, human VSMCs were treated with Ang-II to establish an in vitro AD cell model. qRT-PCR analysis demonstrated that Ang-II dose-dependently downregulated the mRNA expression of VSMC-specific contractile genes, including calponin and SM22α, confirming the phenotypic switch of VSMCs induced by Ang-II stimulation (Figure 4A). Subsequently, TOPORS knockout (siTOPORS) and overexpression (oeTOPORS) VSMC lines were constructed to evaluate the impact of TOPORS on cell function (Figure 4B). CCK-8 assays revealed that, compared to the control group, cell viability was significantly reduced in the Ang-II-induced model group. Notably, TOPORS inhibition partially rescued cell viability, whereas TOPORS overexpression further suppressed viability (Figure 4C). Flow cytometry and TUNEL staining consistently showed that Ang-II treatment markedly increased apoptotic rates compared to controls. This pro-apoptotic effect was reversed by TOPORS inhibition but exacerbated by TOPORS overexpression (Figure 4D,E). Additionally, ELISA results indicated that Ang-II stimulation significantly elevated the secretion of pro-inflammatory cytokines, including TNF-α, IL-1β, IL-6, and *IFN-*α (Figure 4F–I). TOPORS inhibition reduced the levels of these cytokines, while TOPORS overexpression further promoted their release. Collectively, these data suggest that TOPORS may exacerbate AD-like phenotypes in VSMCs by suppressing viability, promoting apoptosis, and enhancing inflammatory responses, whereas its inhibition alleviates these pathological processes.
3.5. Regulation of PI3K/AKT and p53 Signaling Pathways by TOPORS in VSMCs
qRT-PCR and Western blot analyses revealed that, compared with the sham control group, the mRNA and protein expression levels of p53 were significantly upregulated in the AD cell model group. Although the mRNA expression levels of PI3K and AKT were significantly decreased (p < 0.01), their protein expression levels did not change significantly, while their phosphorylation levels (p-PI3K and p-AKT) were significantly reduced (Figure 5A–D). Inhibition of TOPORS gene expression alleviated these changes induced by the AD model, whereas overexpression of TOPORS exacerbated these gene alterations. These results suggest that TOPORS may influence the progression of aortic dissection by regulating the PI3K/AKT and p53 signaling pathways. Subsequent treatment of VSMC models with the p53 inhibitor PFTα, followed by flow cytometry and TUNEL staining, demonstrated that PFTα could mitigate cell apoptosis induced by the AD model (Figure 5E,F) and reduce the upregulation of inflammatory factors such as TNF-α, IL-1β, IL-6, and IFN-α (Figure 5G–J). Co-immunoprecipitation experiments further confirmed that the level of SUMOylation of p53 was significantly higher in the oe-TOPORS VSMC model group than in the siTOPORS group (Figure 5K).
4. Discussion
Aortic dissection (AD) remains a critical challenge in clinical and basic research due to its complex pathological mechanisms and limited therapeutic options. In recent years, SUMOylation, a key post-translational modification, has been increasingly recognized for its roles in cardiovascular diseases [14,16], yet its specific regulatory network in AD pathogenesis requires further exploration. SENP6, TRIM27, and TOPORS are critical regulators of SUMOylation: TRIM27 and TOPORS function as E3 ligases (rate-limiting enzymes in SUMOylation), while SENP6 acts as a SUMO protease that specifically cleaves polySUMO2/3 chains to regulate DNA repair, apoptosis, and senescence [22]. In this study, clinical sample analysis first revealed that TOPORS was significantly upregulated in aortic tissues of AD patients, whereas SENP6 and TRIM27 expression remained unchanged. BAPN-induced animal models further demonstrated that TOPORS expression correlated with medial aortic injury, inflammatory responses, and VSMC dysfunction. Notably, TOPORS inhibition in BAPN-induced AD mice significantly reduced aortic dilation, elastic fiber fragmentation, and serum inflammatory cytokine levels, suggesting TOPORS as a potential driver of AD progression.
TOPORS was initially identified as a cellular binding partner of DNA topoisomerase I and p53. Subsequent studies showed that TOPORS enhances SUMO-1 conjugation to p53 in vitro and in vivo, with TOPORS-induced p53 SUMOylation associated with increased endogenous p53 protein levels [23,24,25]. In AD pathogenesis, massive VSMC apoptosis critically weakens aortic wall integrity. As a classic pro-apoptotic transcription factor, p53 is significantly upregulated in AD patients [26]. Consistent with this, our study found elevated p53 mRNA/protein expression and apoptosis in AD mouse models and VSMC models, which were reversed by TOPORS silencing. These results indicate that aberrant TOPORS overexpression promotes VSMC apoptosis via enhanced p53 SUMOylation during AD progression.
Inflammation is another key driver of AD. During AD progression, immune cell infiltration (e.g., macrophages, neutrophils) increases significantly [27], leading to excessive secretion of pro-inflammatory cytokines (IL-1β, TNF-α, IL-6) that activate VSMC apoptotic pathways, upregulate matrix metalloproteinases (MMPs), and exacerbate aortic wall damage [28]. Our study demonstrated that treating VSMC models with the p53 inhibitor PFTα significantly reduced secretion of IL-1β, TNF-α, IL-6, and *IFN-*α, thereby alleviating AD pathogenesis. Combining previous findings with our results, we hypothesize that TOPORS modulates p53 stability and transcriptional activity via SUMOylation, thereby regulating inflammatory responses in AD.
The PI3K/AKT signaling pathway is a central regulator of cell survival; its inactivation directly impairs VSMC proliferation and migration [29,30]. Clinical studies have shown reduced phosphorylated AKT (p-AKT) levels in AD aortic walls, accompanied by dephosphorylation of the pro-apoptotic protein Bad and activation of Caspase-3/9, leading to increased VSMC apoptosis [31]. In our experiments, TOPORS silencing promoted PI3K and AKT phosphorylation, whereas TOPORS overexpression decreased their phosphorylation. These data suggest that TOPORS exacerbates aortic injury not only by mediating p53 SUMOylation but also by inhibiting PI3K/AKT pathway activation.
Although TOPORS shows potential as a driver of AD progression, several key challenges need to be tackled before these findings can be applied clinically. The results come from preclinical models and small-scale clinical sample analyses; large, multi-center clinical trials are necessary to confirm the link between TOPORS expression levels and AD severity, along with the safety and effectiveness of therapies targeting TOPORS. Moreover, the intricate interactions among TOPORS, p53, and the PI3K/AKT pathway imply that treatments solely targeting TOPORS might have restricted efficacy, emphasizing the demand for combination therapies that concurrently regulate multiple disease-related pathways. Concerning patient classification, our data indicate that non-syndromic AD patients, especially those with heightened TOPORS expression and p53 overactivation in aortic tissues, are most apt to benefit from TOPORS suppression. Additionally, THE research offers a genetic screening tool for higher dissection risk.
5. Conclusions
In conclusion, this study is the first to demonstrate that TOPORS plays a critical role in the pathological progression of aortic dissection (AD) by regulating PI3K/AKT, the p53 signaling pathway and inflammatory responses through SUMOylation. These findings not only expand the current understanding of AD’s molecular mechanisms but also identify TOPORS as a novel candidate target for AD therapy, potentially facilitating a paradigm shift in clinical strategy. Future research should address existing technical limitations to accelerate the clinical translation of TOPORS-related mechanisms, thereby providing theoretical support for reducing AD-associated mortality and disability rates.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yin Z.Q. Han H. Yan X.C. Zheng Q.J. Research Progress on the Pathogenesis of Aortic Dissection Curr. Probl. Cardiol.20234810124910.1016/j.cpcardiol.2022.10124935568084 · doi ↗ · pubmed ↗
- 2Xiao Q. Li Y. Cai B. Huang X. Fang L. Liang F. Chen L. Xu K. Zhang W. Wang X. CCDC 80 Protects against Aortic Dissection and Rupture by Maintaining the Contractile Smooth Muscle Cell Phenotype Adv. Sci.202512 e 250210810.1002/advs.202502108 PMC 1224501240278823 · doi ↗ · pubmed ↗
- 3Wang Y.Y. Jia X. Zhang Y.F. Zhang B. Zhou Y.Z. Li X.R. Zhu X.Y. Xia J.Q. Ren J. Zou C. ACKR 1hi E Cs Promote Aortic Dissection Through Adjusting Macrophage Behavior Circ. Res.202513621122810.1161/CIRCRESAHA.124.32545839692014 · doi ↗ · pubmed ↗
- 4Liu H. Yang P. Chen S. Wang S. Jiang L. Xiao X. Le S. Chen S. Chen X. Ye P. Ncf 1 knockout in smooth muscle cells exacerbates angiotensin II-induced aortic aneurysm and dissection by activating the STING pathway Cardiovasc. Res.20241201081109610.1093/cvr/cvae 08138639325 PMC 11288755 · doi ↗ · pubmed ↗
- 5Yu T. Li X. Wang C. Yang Y. Fu X. Li T. Wang W. Liu X. Jiang X. Wei D. Lactylation of Mitochondrial Adenosine Triphosphate Synthase Subunit Alpha Regulates Vascular Remodeling and Progression of Aortic Dissection Research 20258079910.34133/research.079940800583 PMC 12342782 · doi ↗ · pubmed ↗
- 6Liu L. Zhang J. Dong Z. Cui Y. Zou X. Lai H. Gu J. Weng X. Jin X. Qiu T. Histone Lactylation-Mediated Metabolic Remodeling in Vascular Smooth Muscle Cells Aggravates Aortic Aneurysm and Dissection by Promoting Lactate Accumulation Circulation 202615318920910.1161/CIRCULATIONAHA.125.07257641487086 PMC 12952488 · doi ↗ · pubmed ↗
- 7Hertz E.P.T. Mailand N. The essential functions of protein SUM Oylation in cell proliferation Nat. Struct. Mol. Biol.2023301252125310.1038/s 41594-023-01046-z 37474740 · doi ↗ · pubmed ↗
- 8Liu J.C.Y. Ackermann L. Hoffmann S. Gál Z. Hendriks I.A. Jain C. Morlot L. Tatham M.H. Mc Lelland G.L. Hay R.T. Concerted SUMO-targeted ubiquitin ligase activities of TOPORS and RNF 4 are essential for stress management and cell proliferation Nat. Struct. Mol. Biol.2024311355136710.1038/s 41594-024-01294-738649616 PMC 11402782 · doi ↗ · pubmed ↗