Preliminary Investigation into the Predation of Pomacea canaliculata by Aquatica leii Larvae
Jiangtao Luo, Chunlin An, Yingjun Wu, Huachao Xu

TL;DR
This study shows that Aquatica leii firefly larvae, especially in their 4th-instar stage, can effectively prey on the invasive Pomacea canaliculata snail, offering a potential eco-friendly control method.
Contribution
The study identifies 4th-instar Aquatica leii larvae and their midgut extracts as highly effective biocontrol agents against Pomacea canaliculata.
Findings
4th-instar Aquatica leii larvae had the shortest lethal time (7.37 min) and highest consumption (1.23 g) of Pomacea canaliculata.
Midgut extracts caused 96.7% mortality in Pomacea canaliculata, indicating potent bioactive compounds.
Larvae accepted Pomacea canaliculata as prey, showing preference over native snails in controlled experiments.
Abstract
The search for environmentally friendly ways to control the invasive Pomacea canaliculata has led scientists to look at its natural predators. This study focuses on the larvae of Aquatica leii, a freshwater firefly native to China. We wanted to see if these underwater larvae could effectively prey on the destructive Pomacea canaliculata. In our lab tests, not only did the firefly larvae successfully attack and eat the Pomacea canaliculata, but we also found that one particular growth stage—the 4th-instar larvae—was the most effective hunter, killing Pomacea canaliculata the fastest and consuming the most. A particularly notable finding was that liquid extracted specifically from the larval midgut exhibited strong lethal activity against P. canaliculata under experimental conditions, causing over 96% mortality. This suggests that the larvae may use potent injected toxins to subdue their…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
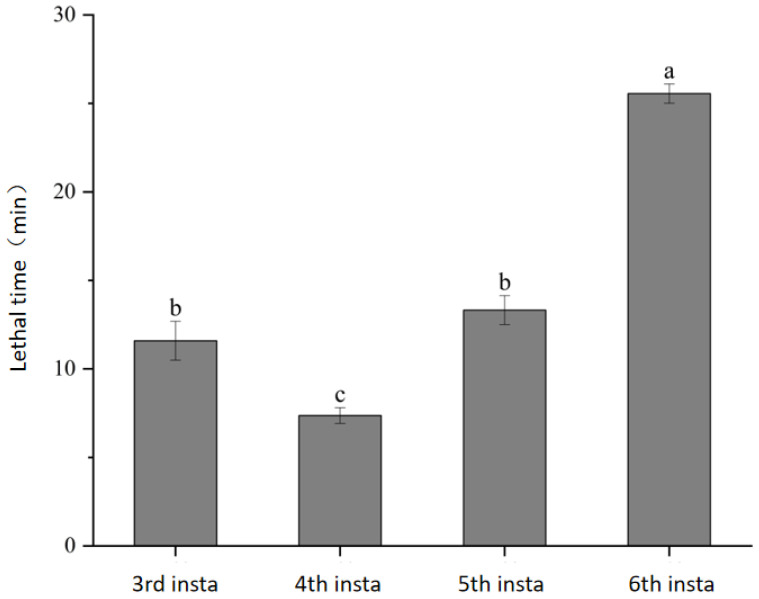 Figure 1
Figure 1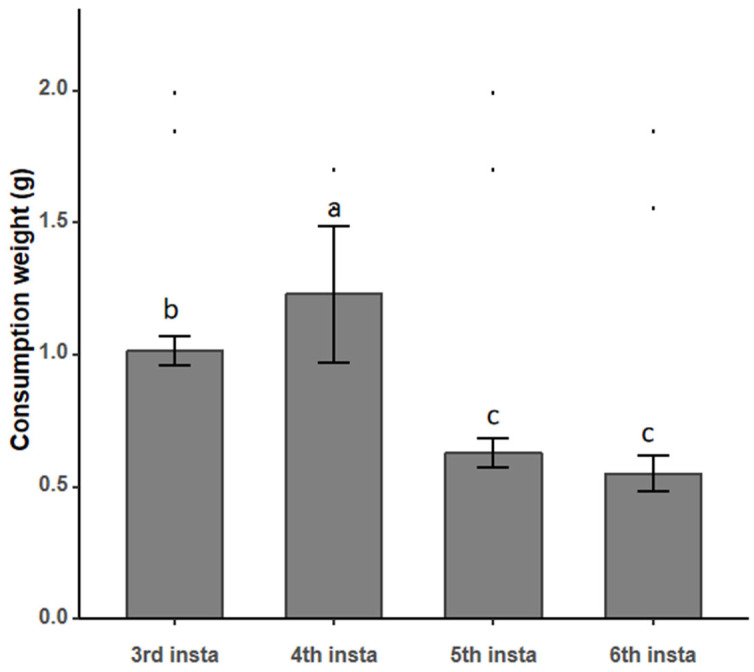 Figure 2
Figure 2- —National Key R&D Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMollusks and Parasites Studies · Forensic Entomology and Diptera Studies · Invertebrate Taxonomy and Ecology
1. Introduction
Pomacea canaliculata (Gastropoda: Mesogastropoda: Ampullariidae) [1] is a widespread South American freshwater mollusk whose range is rapidly expanding and is now becoming a common component of the fauna of many countries in Southeast Asia. It is listed among China’s first cohort of invasive alien species. Listed among China’s first cohort of invasive alien species, it has rapidly spread since its introduction, colonizing paddy fields, ditches, and wetland ecosystems across multiple southern provinces [2]. This snail exhibits a broad diet and high reproductive capacity, causing severe damage to economically important aquatic crops such as rice, water bamboo, and lotus root. This leads to significant yield losses, quality degradation, and substantial annual economic damage [3]. Statistics indicate that the infestation area of P. canaliculata in China had reached 1.7011 million hectares by 2020 [4].
Current control strategies for P. canaliculata are generally categorized into physical and chemical methods. Chemical control relies heavily on molluscicides such as niclosamide ethanolamine salt (NES) and metaldehyde (MD) [5]. However, their long-term and extensive use raises concerns regarding pesticide residues, water pollution, and potential threats to aquaculture and ecological security. Physical methods, such as manual collection of snails and egg masses, are notably inefficient [6]. Consequently, developing environmentally friendly and sustainable biological control technologies has become an urgent need for managing P. canaliculata.
Some practical attempts have been made at biological control using aquatic animals like ducks and soft-shelled turtles to consume P. canaliculata [7,8]. However, large-scale application is often limited by farming conditions, control precision, and scope of applicability. Finding a natural enemy organism that is adapted to aquatic environments, highly specific, and offers efficient control presents a new direction for managing this invasive snail.
Fireflies (Insecta: Coleoptera: Lampyridae) [9,10,11], represent an important group of resource insects. The larvae of many terrestrial species feed on mollusks such as snails and slugs, demonstrating considerable potential for the biological control of agricultural and forestry pests [12,13]. Aquatica leii is an aquatic firefly species endemic to China, belonging to the family Lampyridae [14]. Its entire larval stage occurs in clean aquatic environments such as streams and paddy fields, where it preys on small snails and bivalves [15]. Therefore, this study considers it to possess the biological basis for acting as a potential natural enemy against P. canaliculata.
To date, research on the biological characteristics and ecological functions of A. leii, both domestically and internationally, remains in its early stages [16,17,18]. There is a lack of systematic reports on its predatory capability against P. canaliculata, its feeding preferences, or its control efficacy. Therefore, this study employs A. leii larvae as test subjects. By measuring parameters such as lethal time and consumption rate per unit time against P. canaliculata, we preliminarily assess the potential of A. leii for application in the biological control of P. canaliculata. The aim is to provide a scientific basis for the eco-friendly management of P. canaliculata and the utilization of aquatic fireflies as a biological resource.
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Test Insects
The larvae of A. leii used in this experiment were all purchased in 2024 from firefly breeding bases in Jiangxi and Fujian provinces, China. To ensure physiological uniformity, only larvae originating from the same captive-reared population (maintained under standardized conditions for multiple generations) were used. All larvae were laboratory-reared and supplied with detailed rearing history, including diet (freshwater snails) and environmental conditions (28 ± 1 °C, 14 L:10 D photoperiod). A. leii undergoes complete metamorphosis, with a life cycle consisting of egg, larval (typically 6 instars), pupal, and adult stages. The larval stage lasts approximately 6–8 months under natural conditions, during which larvae molt 5–6 times before pupation [15,16]. Instar determination was based on head capsule width measurement and molting records, following the criteria established by Fu et al. [16]. Only larvae of the same instar and similar body size were selected for experiments.
2.1.2. Prey
P. canaliculata exhibits an extremely high reproductive rate, with juvenile snails reaching sexual maturity within just three months of development [19]. Consequently, the period before sexual maturity represents the optimal window for its control. Therefore, in this experiment, P. canaliculata individuals weighing 0.5–1 g (shell height approximately 10–15 mm) were used as the target prey, while Cipangopaludina chinensis (Gastropoda: Architaenioglossa: Viviparidae), a native sympatric snail species, served as the reference prey (weighing 0.8–1.2 g, shell height approximately 15–20 mm). All snails were collected from multiple irrigation ditches and ponds within a 5 km radius of Hangzhou Xiaoshan International Airport (30.23° N, 120.26° E), Zhejiang Province, China. To ensure genetic diversity and representativeness, snails were pooled from at least five distinct collection sites. Immediately after collection, snails were subjected to a 7-day quarantine period in the laboratory under controlled conditions (28 ± 1 °C, aerated dechlorinated water, natural light cycle). During quarantine, snails were visually inspected for physical damage, abnormal behavior, and the presence of parasites or egg masses. Only apparently healthy, active snails with intact shells were used. To acclimate snails to laboratory conditions and minimize stress, they were held in aquaria with dechlorinated water and fed fresh lettuce ad libitum for at least 3 days prior to experiments.
C. chinensis was chosen as the reference prey because it is a native sympatric snail species commonly found in the same habitats as P. canaliculata in southern China, and it represents a natural prey item for A. leii larvae [18]. This allows assessment of prey choice in a context relevant to the firefly’s natural foraging behavior.
2.2. Experimental Methods
2.2.1. Larval Pre-Treatment
Prior to experiments, all healthy and active A. leii larvae (3rd to 6th instar) were subjected to a standardized 7-day starvation period in a constant temperature incubator (Ningbo Jiangnan Instrument Factory, RXZ-328A, Ningbo, China) under a fixed photoperiod (L:D = 14 h:10 h) to normalize their hunger level.
2.2.2. Feeding Preference Assay
Randomly selected, pre-treated larvae (3rd to 6th instar, equal numbers per instar) were used as test insects and divided into 4 groups by instar, with 15 individuals per group. The assay was conducted in 15 cm diameter Petri dishes (Nest Biotechnology, 150101, Wuxi, China) lined with moist filter paper. The central area was temporarily divided into three equal-sized (sector-shaped) compartments by removable partitions. A single starved larva was placed in the central compartment. After a 5 min acclimation period, the partitions were removed, and one live P. canaliculata and one live C. chinensis of similar weight (approx. 1.0 g) were simultaneously placed in the two opposing (sector-shaped) compartments. A total of 36 independent trials were conducted using new larvae and snail individuals for each trial. Larvae were randomly selected from the 3rd to 6th instar groups (15 individuals per instar) to ensure representation across all instars. Larval behavior was recorded via video during a 2 h observation period. Recorded metrics included the first attack target (defined as contact with the mouthparts and attempted predation) and the final feeding target (sustained feeding for over 5 min). This experiment aimed to determine the fundamental feeding preference of A. leii larvae.
2.2.3. Determination of Lethal Time of Pomacea canaliculata by Aquatica leii Larvae at Different Instars
For each larval instar, 15 pre-treated larvae were randomly divided into three subgroups (n = 5 per subgroup), which served as three biological replicates. Thus, “three replicate treatment groups” refers to these three parallel subgroups run simultaneously. The experiment was performed once for each instar, with each larva tested individually. Mortality of P. canaliculata was confirmed when the snail showed no movement, failed to respond to gentle probing of the foot with a blunt needle, and retracted deeply into the shell for more than 5 min. In cases of uncertainty, observation was extended for an additional 10 min. Control groups (snails without larvae) were maintained under identical conditions to monitor background mortality.
2.2.4. Weekly Consumption Amount of Pomacea canaliculata by Aquatica leii Larvae at Different Instars
For each larval instar, 15 pre-treated larvae were randomly divided into three subgroups (n = 5 per subgroup) as three biological replicates. This experiment was conducted independently from the lethal time assay, using a new set of larvae and snail tissue. Each larva was provided with 5 g of fresh P. canaliculata tissue (dissected from live snails, excluding the shell). To correct for autogenic changes in tissue weight (e.g., water loss), three control containers without larvae were set up in parallel. The amount consumed per larva over 7 days was calculated as: (initial tissue weight—remaining tissue weight)—(mean weight loss in control containers). Tissue weight was measured after gently blotting with absorbent paper.
2.2.5. Determination of Lethal Efficiency of Aquatica leii Digestive Tract Fluids against Pomacea canaliculata
Sixty healthy 4th-instar A. leii larvae were immobilized by brief exposure to −20 °C for 1 min in a cryogenic freezer (Haier, DW-40L508, Qingdao, China). Dissection was performed on a wax plate under a stereomicroscope (Nanjing Jiangnan Novel Optics, JSZ6, Nanjing, China) in chilled physiological saline. The entire digestive tract was excised and divided into four anatomical regions based on external features: buccal region (mouthparts and pharynx), foregut (esophagus and crop), midgut (anterior and posterior sections, distinguished by darker pigmentation and presence of food bolus), and hindgut (intestine and rectum). Each section was weighed and homogenized manually in ice-cold physiological saline at a ratio of 1:1 (w/v). The homogenate was centrifuged at 4 °C, 6000 rpm for 15 min (Eppendorf, 5424R, Hamburg, Germany), and the supernatant was collected. The final concentration of the supernatant corresponded to approximately 0.5 g of tissue per mL (original tissue diluted 1:1 with saline).
Prior to the main experiment, a dose-finding pre-test was conducted using midgut extract at doses of 5, 10, and 20 µL per snail (n = 5 per dose). The 10 µL dose was selected because it induced clear mortality without causing immediate death from injection trauma, allowing assessment of extract-specific effects.
For the bioassay, 10 µL of each supernatant was injected into the foot muscle of individual P. canaliculata (0.5–1.0 g) using a 50 µL Hamilton syringe (Hamilton, 80965, Reno, NV, USA) fitted with a 30-gauge (30 G) needle (KDL, 30 G, Shanghai, China). Four treatment groups (buccal, foregut, midgut, hindgut extracts) and three control groups were included: (1) physiological saline (vehicle control), (2) sterile water (to control for osmotic effects), and (3) sham injection (needle insertion without fluid injection) to account for physical injury. Each group comprised 30 snails, divided into three replicates of 10 snails each. Mortality was recorded at 1, 3, 6, and 12 h post-injection; snails were considered dead if they showed no movement and failed to respond to gentle prodding.
2.2.6. Data Analysis
Data were analyzed using SPSS Statistics version 26.0 (IBM Corp., Armonk, NY, USA). For the feeding preference assay, a chi-square goodness-of-fit test [20] was used to compare observed frequencies of first attack and final feeding choices against an expected 1:1 distribution. For lethal time and consumption amount, data were first tested for normality (Shapiro–Wilk test) and homogeneity of variances (Levene’s test). As all data met these assumptions (p > 0.05), one-way analysis of variance (ANOVA) was performed to compare means among instars, followed by Tukey’s honestly significant difference (HSD) post hoc test for multiple comparisons. For the extract injection experiment, mortality rates among treatment groups were compared using Fisher’s exact test (due to small expected frequencies in some cells). A significance level of significance level (α) of 0.05 was used for all tests.
3. Results
3.1. Feeding Preference
To determine whether the larvae of A. leii actively select P. canaliculata as prey and to evaluate their choice preference in the presence of different prey items—thereby assessing their targeted predation potential against P. canaliculata in natural environments—a two-choice experiment was conducted, with the results summarized in Table 1.
The chi-square goodness-of-fit test results indicated that while the larvae of A. leii showed a tendency to preferentially select C. chinensis as the initial attack target, this difference was not statistically significant (χ^2^ = 2.78, p > 0.05). Similarly, no significant difference was observed in the proportion of larvae that ultimately fed on either snail species (χ^2^ = 1.00, p > 0.05). These results suggest that A. leii larvae are capable of and willing to accept P. canaliculata as a food source. However, it should be noted that the total number of replicates (36 across all instars combined) may limit the statistical power of the chi-square test to detect small but biologically meaningful preferences. Furthermore, prey choice experiments were conducted only at a 1:1 ratio, which does not capture potential density-dependent shifts in feeding decisions that may occur in natural habitats where prey abundance varies. Future studies should incorporate multiple prey density ratios to better understand the foraging behavior of A. leii under ecologically relevant conditions.
3.2. Lethal Time
This study found that the first and second instar larvae of A. leii were incapable of preying on intact P canaliculata, only consuming processed snail tissue. This observation is consistent with findings reported by Guo Zhaoxiang in a study on Pyrocoelia pectoralis [21]. To accurately assess the actual predatory effect of each larval instar, these two early instars were excluded from the experimental design. To facilitate observation and eliminate interference from factors such as natural mortality of P. canaliculata, individual snails were housed separately in transparent rearing containers measuring 4.0 cm × 4.0 cm × 2.5 cm. Under constant temperature conditions of 28 °C, the lethal time exerted by third, fourth, fifth, and sixth instar larvae on P. canaliculata was determined (Figure 1).
The data indicate that the lethal time required by sixth-instar larvae to kill P. canaliculata was significantly longer than that of all other instars, with a mean of 25.89 min, suggesting the slowest predatory process against P. canaliculata at this stage. In contrast, fourth-instar larvae exhibited the shortest mean lethal time of only 7.37 min, which differed significantly from the third-, fifth-, and sixth-instar groups, indicating that fourth-instar larvae likely possess the strongest lethal capacity against P. canaliculata.
The lethal times of third- and fifth-instar larvae were relatively similar, at 11.26 min and 12.99 min respectively, with no statistically significant difference between them. However, both were significantly longer than that of the fourth-instar group and shorter than that of the sixth-instar group. These results demonstrate that lethal time does not simply decrease or increase progressively with larval instar.
3.3. Measurement of Consumption Amount
Prey consumption is a key metric for evaluating the predatory efficacy and energy acquisition strategy of a natural enemy. To systematically analyze the nutritional niche and utilization efficiency of A. leii larvae at different instars toward their prey, this study, following the recording of lethal time, measured the actual amount of P. canaliculata tissue consumed per unit time by larvae of each instar (with three replicates per instar) (Figure 2).
As shown in Figure 2, the average consumption amounts of larvae at different instars exhibited significant inter-group differences. Among them, the fourth-instar larvae showed the highest consumption, with an average of 1.23 g, whereas the sixth-instar larvae showed the lowest consumption, averaging only 0.55 g. Overall, larvae at the third and fourth instars had relatively higher consumption levels, while consumption tended to decrease as larval development progressed beyond these stages.
3.4. Lethal Efficiency of Aquatica leii Digestive Tract Fluids Against Pomacea canaliculata
This study has confirmed that A. leii larvae are capable of preying upon P. canaliculata and accept it as a food source. The feeding process of lampyrid larvae is not merely a matter of physical ingestion. Studies have shown that they primarily inject digestive tract fluids rich in various enzymes (such as proteases and lipases) into their prey [13] to facilitate external pre-digestion, thereby liquefying and absorbing the prey’s tissues [18]. Consequently, digestive tract fluid is a key physiological factor enabling firefly larvae to kill and digest mollusk prey [13]. Therefore, this study investigated the direct effect of in vitro digestive tract fluids from A. leii on P. canaliculata (Table 2) to further elucidate its potential as a biocontrol agent.
The data presented in Table 2 clearly demonstrate that the midgut extract exhibited exceptionally strong lethal activity, causing the death of 27, 30, and 30 P. canaliculata individuals across the three replicate groups, respectively, resulting in an overall mortality rate as high as 96.7%. In contrast, the lethal effects observed in the other treatment groups were minimal.
4. Discussion
The strong reproductive capacity of P. canaliculata and the lack of effective natural enemies to control it make the management of this invasive species particularly challenging [22]. This study found that the larvae of the aquatic firefly A. leii possess clear predatory capability and lethal potential against P. canaliculata. The results indicate that although A. leii larvae show a preference for the native snail Cipangopaludina chinensis under free-choice conditions, they also exhibit a positive willingness to feed on P. canaliculata. This finding holds significant ecological importance: in habitats where P. canaliculata has invaded extensively and displaced native snail species, A. leii can utilize the invader as an effective alternative prey resource. This allows the firefly population to persist and continue exerting pest control functions. This ability reduces the dependency of its control efficacy on specific native prey, thereby enhancing its environmental adaptability and stability as a biological control agent.
It is important to note that in this part of the experiment, larvae were provisioned with dissected snail tissue rather than live, intact prey. Therefore, the measured consumption amount reflects ingestion rate under conditions of easy access to food, not the full predation capacity (which includes search, attack, and handling of live snails). This limitation should be considered when interpreting the ecological relevance of these consumption values.
The study further reveals that A. leii larvae are not only behaviorally receptive to P. canaliculata as food but also capable of killing it. Predatory efficacy varied significantly among larval instars, with the 4th-instar larvae exhibiting optimal performance, characterized by the shortest mean lethal time, the highest consumption per unit time, and the most prominent snail control efficiency. In contrast, consumption decreased and lethal time increased in later instars (5th–6th), which may be related to their physiological shift towards the prepupal stage and associated changes in energy allocation strategies—a finding consistent with observations in other Coleoptera species [21,23]. This instar-specific pattern provides clear guidance for practical application: when implementing control through mass rearing and release or field population augmentation, priority should be given to utilizing active 3rd and 4th instar larvae to achieve the best cost–benefit ratio and immediate pest suppression effect. Simultaneously, it highlights the need to consider the generation cycle and instar structure in natural population management to ensure sustained control pressure.
This study found that the lethal effect of A. leii larvae on P. canaliculata is not solely due to physical consumption but is significantly mediated by the injection of digestive tract fluid, with midgut secretions playing a decisive role. The lethal effects of extracts from different parts of A. leii varied markedly. No mortality of P. canaliculata was observed in the physiological saline control group or the foregut extract treatment group during the observation period, indicating that mere mechanical stimulation or foregut secretions lack significant lethal activity. While mouthpart and hindgut extracts caused mortality in a few individuals, their mortality rates were only 3.3% and 16.7%, respectively, indicating limited efficacy. In stark contrast, the midgut extract achieved a mortality rate as high as 96.7% against P. canaliculata, far exceeding that of extracts from other digestive tract sections. This suggests that the midgut likely contains specific enzymes or toxins [18] capable of efficiently disrupting the tissue and physiological functions of P. canaliculata. This data further confirms that the digestive fluid or related enzyme systems within the midgut of A. leii larvae are the key physiological factors responsible for the death of P. canaliculata. It is worth noting that all extracts were standardized to the same concentration (0.5 g tissue equivalent per mL) prior to injection, ensuring that the observed differences in lethal activity reflect intrinsic properties of each digestive tract region rather than concentration variations. The 10 µL dose was selected based on a pre-experiment showing that this volume induced clear mortality without causing immediate death from injection trauma. Future studies should explore dose–response relationships to further characterize the potency of midgut-derived compounds.
Although our experimental design included multiple controls (physiological saline, sterile water, and sham injection) to account for physical injury and solvent effects, we acknowledge that an additional control using gut contents from a non-molluscivorous aquatic insect would further strengthen the specificity of the observed activity. Future studies incorporating such controls, along with investigations into the temporal dynamics of toxin secretion and transport within the digestive tract, will help clarify the mechanisms underlying the potent lethal effect of A. leii midgut secretions.
5. Conclusions
This study provides the first systematic evidence that larvae of the aquatic firefly Aquatica leii have significant potential as a biocontrol agent against the invasive snail Pomacea canaliculata. The main findings and their implications are summarized as follows:
A. leii larvae readily accept P. canaliculata as prey, with the 4th instar exhibiting optimal predatory performance—characterized by the shortest mean lethal time (7.37 min) and the highest weekly consumption (1.23 g). This identifies the 4th instar as the key functional stage for snail suppression and provides practical guidance for mass rearing and release strategies in biocontrol programs.
Midgut crude extract from 4th-instar larvae induced 96.7% mortality in P. canaliculata within 12 h, far exceeding the effects of extracts from other digestive tract regions. This suggests that predation by A. leii may involve not only physical consumption but also the action of specific digestive enzymes or toxins, a hypothesis that requires further biochemical validation. Secreted by the midgut, which rapidly incapacitates prey through an “injection–external digestion” mode [18]. This discovery not only explains the high lethal efficiency observed but also points toward the development of novel biogenic molluscicides.
As a native aquatic natural enemy insect [24], A. leii possesses clear potential for integration into green control systems for P. canaliculata. The dual effectiveness of its predatory behavior and physiological lethal mechanism offers a new pathway to replace or supplement existing chemical methods. Future research should focus on field validation of control efficacy, ecological safety assessment, and isolation and characterization of the midgut active compounds to advance this resource insect from experimental promise to practical application.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gattamraju A. Accorsi A. The Golden Apple Snail Pomacea canaliculata: From Zygotes to Stable Mutant Lines J. Vis. Exp.2025 e 6926710.3791/6926741525236 · doi ↗ · pubmed ↗
- 2Dai L. Yang H. Li Z. Liu Y. Chen S. Zhong J. Liu W. Wang H. Integrated control of Pomacea canaliculata in different habitats Chin. J. Anim. Infect. Dis.202510.19958/j.cnki.cn 31-2031/s.20250307.001 · doi ↗
- 3Carlsson N. Kestrup A. Martensson M. Nyström P. Lethal and non-lethal effects of multiple indigenous predators on the invasive golden apple snail Freshw. Biol.2004491269127910.1111/j.1365-2427.2004.01269.x · doi ↗
- 4Zhuo F. Wang H. Liu W. The Occurrence and Control of Pomacea canaliculata in Chinese Farmland from 2011 to 2020 China Plant Prot.2022425255
- 5Wang Z. Yang H. Wan Z. Li B. Effects of different lethal concentrations of niclosamide ethanolamine salt and metaldehyde on the gutmicrobiota structure of Pomacea canaliculata J. Biosaf.202534145154
- 6Wang Z.G. Tan J.C. Tan L. Liu J. Zhong L. Control the egg hatchling process of Pomacea canaliculata (Lamarck) by water spraying and submersion Acta Ecol. Sin.20123218418810.1016/j.chnaes.2012.04.008 · doi ↗
- 7Uehara H. Yusa Y. Effects of predation by the carrion crow Corvus corone (Passeriformes: Corvidae) on field populations of the apple snail Pomacea canaliculata (Architaenioglossa: Ampullariidae)Appl. Entomol. Zool.20266116316910.1007/s 13355-026-00955-x · doi ↗
- 8Uehara H. Murakami H. Yusa Y. Predation by the carrion crow Corvus corone (Passeriformes: Corvidae) on the apple snail Pomacea canaliculata (Architaenioglossa: Ampullariidae) in different locations in Japan Appl. Entomol. Zool.20215638539210.1007/s 13355-021-00747-5 · doi ↗