Genome-Wide Identification and Expression Analysis of NHX Gene Family in Ziziphus jujuba var. spinosa Under Salt and Drought Stress
Lulu Li, Xiaojun Ma, Xinhong Wang, Congcong Liu, Xiaohan Tang, Dali Geng, Xuexiang Li, Aiqin Ding, Jing Shu

TL;DR
This study identifies and analyzes NHX genes in sour jujube, revealing their role in salt stress tolerance and providing insights for future research.
Contribution
The first genome-wide analysis of the NHX gene family in Ziziphus jujuba var. spinosa under abiotic stress.
Findings
Eight NHX genes were identified in sour jujube, distributed across six chromosomes and classified into three subfamilies.
Most ZjNHX genes are up-regulated by salt stress, with ZjNHX1 showing a 40-fold increase in root expression within 3 hours.
Promoter regions of ZjNHX genes contain cis-regulatory elements linked to stress adaptation and phytohormone signaling.
Abstract
Background/Objectives: Ziziphus jujuba var. spinosa (sour jujube) is a traditional medicinal plant with remarkable tolerance to abiotic stresses, particularly salinity. However, the regulatory mechanisms underlying its salt stress tolerance remain unclear. NHX genes play a crucial role in plant adaptation to salt stress by mediating Na+/K+ transport to maintain intracellular ion homeostasis and pH balance. Although the NHX gene family has been characterized in many plant species, its functional characteristics in sour jujube have not yet been systematically investigated. Methods: In this study, using Arabidopsis thaliana as a reference, we identified NHX genes in sour jujube through genome-wide analysis and molecular approaches, and systematically analyzed their phylogenetic relationships, chromosomal locations, conserved motifs, gene structures, cis-regulatory elements in promoter…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
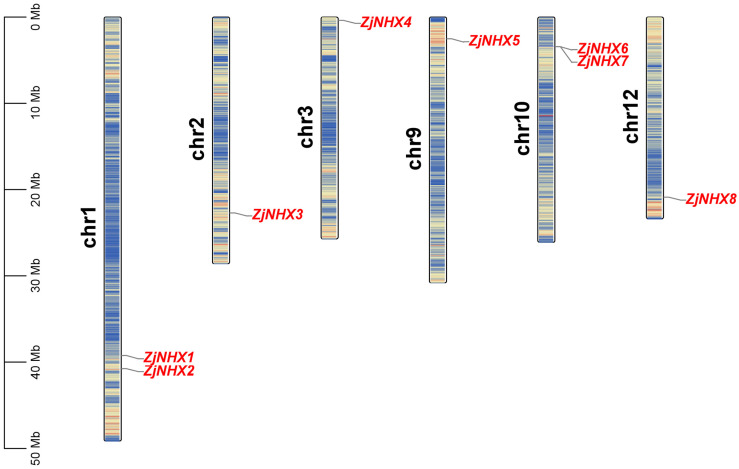 Figure 1
Figure 1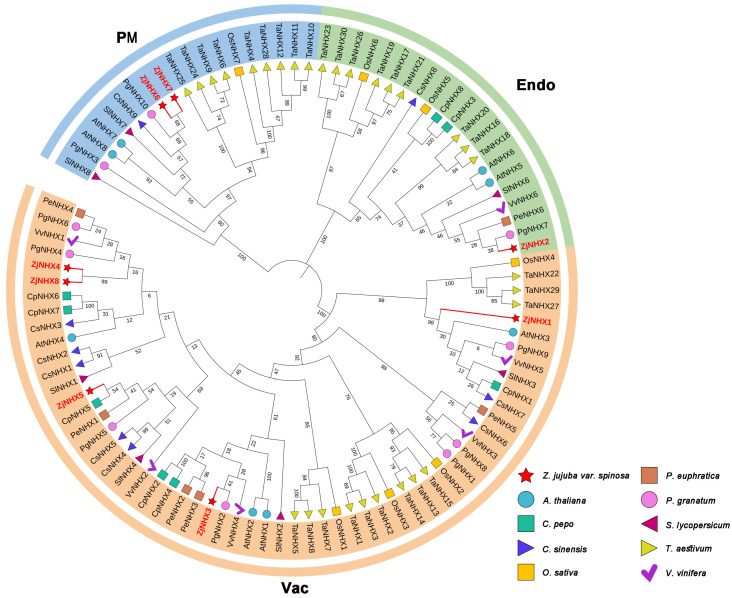 Figure 2
Figure 2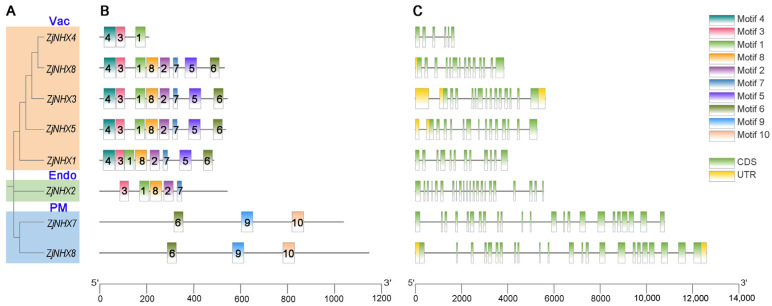 Figure 3
Figure 3 Figure 4
Figure 4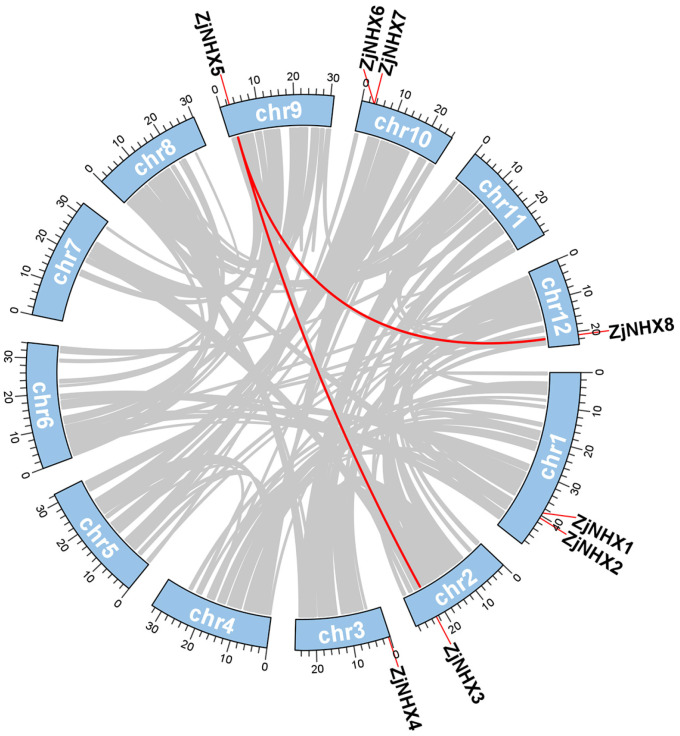 Figure 5
Figure 5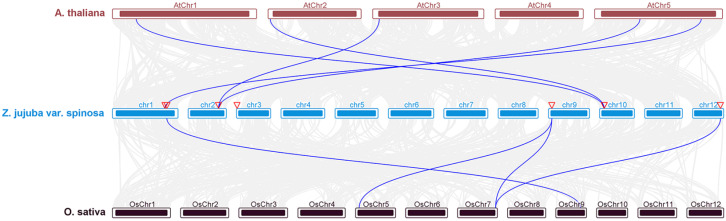 Figure 6
Figure 6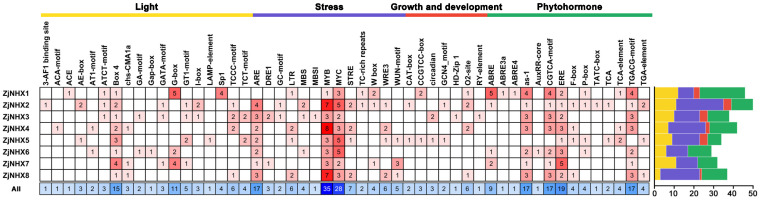 Figure 7
Figure 7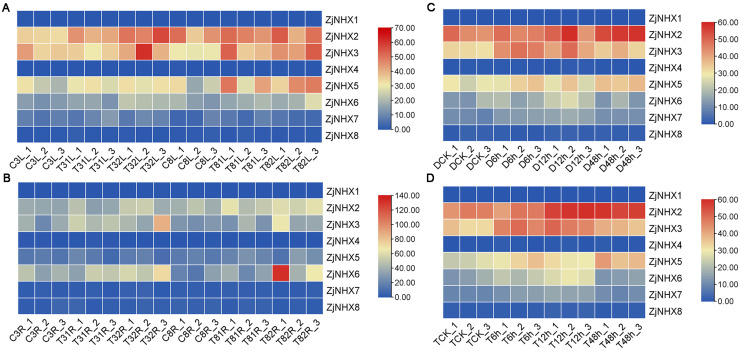 Figure 8
Figure 8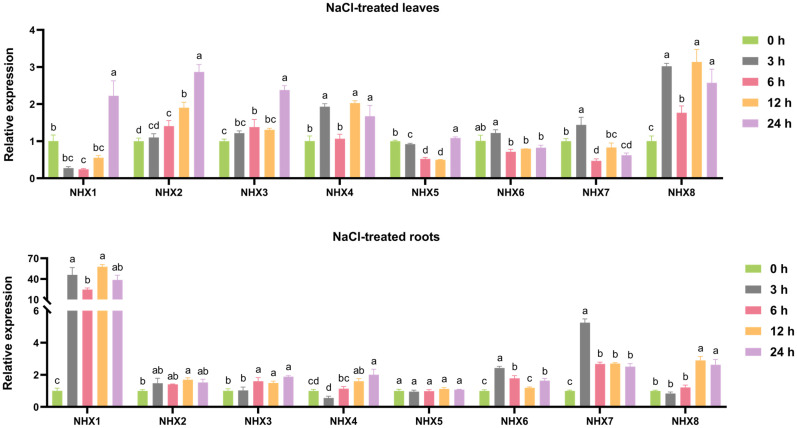 Figure 9
Figure 9- —the Key R&D Program of Shandong Province, China
- —Shandong Agriculture and Engineering University Start-Up Fund for Talented Scholars
- —Shandong Agriculture Research System
- —the Natural Science Foundation of Shandong Province Youth Fund Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsZiziphus Jujuba Studies and Applications · Plant Molecular Biology Research · Postharvest Quality and Shelf Life Management
1. Introduction
Soil salinization is one of the major abiotic stress factors restricting global agricultural production and sustainable development of the ecological environment. According to statistics, salt-affected soils occur in more than 100 countries, and their worldwide extent is estimated at about 1 billion ha [1]. In terms of the area of salt-affected lands, the top-ranking countries were China with 211.74 Mha [2]. Saline soils contain excessive soluble salts, mainly sodium chloride (NaCl) and sodium sulphate (Na_2_SO_4_) or other neutral salts. These salts increase osmotic pressure, diminish water availability and inhibit plant growth. Particularly, high concentrations of Na^+^ disrupt ionic homeostasis in plants, leading to osmotic stress, ionic toxicity, and oxidative damage, ultimately resulting in impaired growth and development as well as reduced yield and quality [3].
Over the course of long-term evolution, certain plants have gradually adapted to saline–alkaline environments. Ziziphus jujuba Mill. var. spinosa (sour jujube) is one such species. In China, sour jujube is an economically important fruit tree that exhibits remarkable tolerance to saline–alkaline, nutrient-poor, and drought conditions, maintaining normal physiological metabolism even in soils with 5‰ salinity [4,5]. According to Shennong’s Classic of the Materia Medica, sour jujube is documented to “calm the five viscera, lighten the body, and prolong life”. Its fruit pulp is rich in vitamins, polysaccharides, and trace elements; the buds and leaves can be processed into substitute tea and health products; the seedlings, known for their high stress tolerance, are widely used as rootstock for cultivated jujube; and the seeds (Suanzaoren) are a well-known traditional Chinese medicine, extensively applied in clinical practice for their sedative and mind-calming effects [6]. This inherent salt–alkali tolerance of sour jujube has motivated us to further investigate the underlying adaptive mechanisms.
To cope with salt stress, plants have evolved a series of complex mechanisms at both the physiological and molecular levels. Among these, Na^+^/H^+^ antiporters (NHXs) are central to maintaining osmotic balance and ion homeostasis under salt stress, regulating cytosolic Sodium (Na^+^), Potassium (K^+^), and Hydrogen (H^+^) [7]. NHX (Na^+^/H^+^ antiporter) belongs to the CPA1 superfamily and is a membrane protein composed of approximately 550 amino acid residues, exhibiting a characteristic transporter architecture [8]. The encoded protein contains a highly conserved Na^+^/H^+^ exchanger domain and typically maintains 10–12 transmembrane domains [9]. Additionally, most NHX proteins include a highly conserved 14-residue motif within the fourth transmembrane segment, represented by the consensus sequence FF(I/L)(Y/F)LFLLPPI. This region is recognized as the binding site for amiloride and its derivatives [10]. Based on their sequence identity and localization, the plant NHX family can be grouped into three distinct functional classes, which are plasma membrane (PM), vacuolar (Vac), and endosomal-associated (Endo), among which vacuolar and endosomal-associated belong to the intracellular class [8,9,11]. Different subclasses of NHX proteins exhibit significant functional differentiation.
In Arabidopsis thaliana, eight NHX genes have been identified. Six of these function as intracellular transporters, further divided based on their subcellular localization into two groups, vacuolar and endosome-associated [11]. The vacuolar membrane type includes AtNHX1 to AtNHX4. Studies have shown that this type of gene utilizes the proton gradient generated by V-type H^+^-ATPase and H^+^-pyrophosphatase to compartmentalize excess Na^+^ from the cytoplasm into the vacuole, reducing Na^+^ toxicity and maintaining an appropriate K^+^/Na^+^ ratio in the cytoplasm, while supporting osmotic regulation through solute accumulation [11,12,13,14,15]. Recent studies have revealed that in addition to sequestering Na^+^ into vacuoles, NHX proteins may also enhance plant salt tolerance by modulating intracellular K^+^ homeostasis under saline conditions [16,17]. Furthermore, distinct functional differences exist among the vacuolar-localized isoforms AtNHX1 to AtNHX4, particularly in their selectivity for transporting Na^+^ versus K^+^ [18]. AtNHX5/6 belong to the endosomal type. Studies have shown that endosomal-localized NHX transporters play a critical role in maintaining intracellular ion homeostasis and pH, regulating protein processing and trafficking of cellular cargo, while also being essential for normal plant growth [11,19]. AtNHX7/SOS1 and AtNHX8 belong to the plasma membrane-localized proteins, which work synergistically with the CBL-CIPK signaling pathway to expel Na^+^ out of the cell [20,21,22]. However, this process of expelling Na^+^ out of the cell further exacerbates the imbalance of osmotic pressure and ion concentration across the membrane. Therefore, this mechanism only has transient salt resistance and cannot enable plants to have long-term salt tolerance under natural environmental conditions [23,24]. Furthermore, some researchers classify plasma membrane-localized proteins into the SOS1 gene family. Studies have found that the NHX and SOS1 gene families have very different evolutionary trajectories and suggest that these divergent evolutionary histories are related to the evolution of their function and cellular localization [25,26].
Multiple studies have demonstrated that Na^+^/H^+^ antiporters play a crucial role in plant salt tolerance and hold potential for genetic engineering applications. This potential is exemplified by the fact that overexpression of NHX genes, whether heterologous or homologous, can significantly enhance plant salt tolerance. For example, studies have found that when the vacuolar NHX antiporter gene VvNHX1 from grapevine was introduced into potato, transgenic plants exhibited enhanced growth rates, tolerance, and reduced sodium ion accumulation under salt stress [27]. Moreover, overexpression of the radish RsNHX1 gene enhanced salt tolerance in A. thaliana. Conversely, plants with the silence of endogenous RsNHX1 were more susceptible to the salt stress [28], which indicates that RsNHX1 acts as a positive regulator of salt stress resistance in radish. Moreover, transgenic tobacco plants overexpressing IlNHX from Iris lactea could compartmentalize more Na^+^ into vacuoles via increased V-ATPase activity. This mechanism maintained Na^+^/K^+^ homeostasis, improved photosynthetic capacity, and preserved plasma membrane integrity under salt stress [29]. In applied research, NHX genes have emerged as pivotal targets for developing stress-tolerant crops in cotton [30,31] and rice [32]. For example, the expression of HtNHX1 or HtNHX2 from Helianthus tuberosus improved the rice tolerance to salinity. Especially, expression of HtNHX2 increased rice grain yield, harvest index, and total nutrient uptake under K^+^-limited salt-stress or general nutrient deficiency conditions [32]. Specifically, overexpression of NHX genes through transgenic approaches significantly enhances salinity tolerance, enabling tomato plants to sustain growth, flowering, and fruit yield under high-saline conditions. Concurrently, vacuolar Na^+^ compartmentalization mediated by these genes minimizes sodium accumulation in fruits, thereby preserving fruit quality from salt-associated impairment [33,34,35]. Collectively, these studies affirm that NHX genes are important candidate genes for the genetic improvement of crop salt tolerance, holding significant application potential.
In recent years, with the rapid development of genome sequencing technologies, the NHX gene family has been identified and functionally characterized at the whole-genome level in various plants, including crops, horticultural plants, and salt-tolerant model plants [36,37,38,39,40]. These studies have elucidated distinctive structural features, conserved motifs, and evolutionary patterns within the NHX gene family. Furthermore, accumulating evidence indicates that NHX proteins contribute not only to salt stress tolerance but also play pivotal roles in plant growth and development. Vacuolar membrane-localized NHX proteins are critical for the active uptake of K^+^ into vacuoles, regulation of cellular turgor pressure, and normal stomatal function [16,41]. Research has demonstrated that Arabidopsis AtNHX5 and AtNHX6 participate in the establishment and maintenance of auxin concentrations during embryogenesis and root tissue development [42]. Additionally, these proteins function within the brassinosteroid signaling pathway [43]. Collectively, NHX genes are implicated throughout the entire plant growth cycle, with their expression being significantly induced by multiple stress conditions and plant hormones.
Although sour jujube exhibits considerable salt tolerance, the underlying molecular mechanisms remain largely unexplored. Given the central role of NHX transporters in plant salt tolerance, a systematic identification and characterization of the NHX gene family in this species is essential for understanding its adaptive strategies and harnessing its genetic potential in jujube breeding. As a stress-resilient wild relative, sour jujube likely harbors unique NHX variants with distinct evolutionary and functional features. This study utilized the whole-genome data of sour jujube to comprehensively identify all members of its NHX gene family through integrated bioinformatics approaches. Systematic analyses were performed on their phylogenetic relationships, gene structures, conserved motifs, chromosomal distributions, collinearity patterns, cis-regulatory elements, and expression profiles across diverse tissues and under abiotic stresses. This study aims to elucidate the evolutionary characteristics and potential functions of the NHX gene family in sour jujube, thereby establishing a robust foundation for investigating its regulatory roles in stress responses and guiding subsequent functional validation. Ultimately, these findings will provide critical genetic resources for breeding salt-tolerant jujube varieties.
2. Materials and Methods
2.1. Genome-Wide Identification of NHX Genes in Z. jujuba var. spinosa
The genome-wide annotation data for Z. jujuba var. spinosa were retrieved from the figshare repository (https://figshare.com/s/ad5d747ccc2ccbb2b65b, accessed on 1 November 2025) [44]. A total of eight A. thaliana NHX protein sequences were downloaded from The Arabidopsis Information Resource (TAIR) (https://www.arabidopsis.org/, accessed on 1 November 2025). To identify candidate genes in Z. jujuba var. spinosa, BLASTP searches were conducted using the AtNHX1-AtNHX8 sequences as queries, applying a strict E-value cutoff of 1 × 10^−5^ in TBtools software version 2.372 [45]. In parallel, the Hidden Markov Model (HMM) profile corresponding to the conserved NHX domain (PF00999) was acquired from the Pfam database (http://pfam.xfam.org/, accessed on 1 November 2025). This HMM profile was employed to validate candidate proteins after preliminary screening with a stringent E-value threshold of 1 × 10^−5^. Only those sequences identified by both BLASTP and HMM searches were ultimately designated as members of the ZjNHX gene family. Candidate sequences were submitted to the NCBI CDD and SMART databases to ensure all identified members contained the characteristic Na^+^/H^+^ exchanger (NHX) domain with default parameter settings.
2.2. Chromosome Localization and Protein Physicochemical Property Analysis
Based on the genomic annotation data of Z. jujuba var. spinosa, chromosomal localization of the identified ZjNHX genes was visualized using TBtools software v2.372 [45]. For protein characterization, the physicochemical parameters, including theoretical isoelectric point (pI), molecular weight (MW), and instability index, were computed with the ProtParam tool on the ExPASy server (https://web.expasy.org/protparam/, accessed on 6 November 2025) [46]. Transmembrane helix regions in the ZjNHX proteins were then identified using the TMHMM-2.0 program [47]. Further analysis involved predicting the secondary structure composition with SOPMA (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 6 November 2025) and constructing the tertiary structures via homology modeling on the SWISS-MODEL platform [48]. Finally, the subcellular localization of each ZjNHX protein was predicted using the Plant-mPLoc server (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/, accessed on 7 November 2025) [49].
2.3. Phylogenetic Tree Construction and Analysis
Multiple sequence alignment was performed using MUSCLE in MEGA v11 software under default parameters [50]. Subsequently, phylogenetic analysis was conducted based on NHX protein sequences from Z. jujuba var. spinosa, A. thaliana, Cucurbita pepo, Camellia sinensis, Oryza sativa, Populus euphratica, Punica granatum, Solanum lycopersicum, Triticum aestivum, and Vitis vinifera. The maximum likelihood (ML) analysis was performed using the JTT model with frequencies (+F) and incorporated a Gamma distribution with Invariant sites (G+I) to model rate heterogeneity across sites. The heuristic search for the ML tree employed the Nearest-Neighbor Interchange (NNI) method, with the initial tree generated automatically (default NJ/BioNJ). Nodal support was evaluated using the bootstrap method with 1000 replications. Finally, the phylogenetic tree was visualized and annotated using the iTOL online tool (https://itol.embl.de, accessed on 13 November 2025).
2.4. Conserved Motif and Gene Structure Analysis
The conserved motifs of all ZjNHX proteins were predicted using the MEME suite (https://meme-suite.org/meme/tools/meme, accessed on 5 November 2025), with the number of motifs expanded from the default (3) to 10 motifs [51]. Motif functional annotation was performed using the InterPro database (https://www.ebi.ac.uk/interpro/search/sequence/, accessed on 5 November 2025) [52]. To analyze the gene architecture, the exon–intron organization of each ZjNHX gene was determined by aligning its coding sequence (CDS) with the corresponding genomic DNA sequence using TBtools v2.372 [45]. Finally, multiple sequence alignment focusing on the conserved domains was carried out with DNAMAN v9 under default parameters.
2.5. Collinearity and Selection Pressure Analysis
For intra- and inter-species synteny analysis of ZjNHX gene family members in sour jujube, the MCScanX plugin in TBtools was employed [45] with an E-value ≤ 1 × 10^−5^, and Num of BlastHits ≥ 5. All coding sequences of syntenic gene pairs were aligned using MAFFT (https://www.ebi.ac.uk/jdispatcher/msa/mafft, accessed on 15 November 2025) with default parameters. To assess whether synonymous substitutions were saturated, substitution saturation tests were performed using DAMBE v7 with default parameters [53]. Ka, Ks, and Ka/Ks values for collinear gene pairs were calculated using TBtools. Divergence time (T) was estimated with the formula T = Ks/(2r) × 10^−6^ million years (Mya), where Ks denotes synonymous substitutions per synonymous site, and r represents the dicot-specific substitution rate of 1.5 × 10^−8^ synonymous substitutions per site per year [54].
2.6. Cis-Acting Element Analysis
To investigate potential transcriptional regulation, the 2 kb upstream sequences from the transcriptional start sites (TSS) of each ZjNHX gene were extracted from the Z. jujuba var. spinosa genome using TBtools v2.372; these were designated as the putative promoter regions. Putative cis-acting regulatory elements within these sequences were then identified through queries against the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 24 November 2025) [55]. The identified elements, which were categorized into stress-responsive, hormone-related, plant development, and light-responsive groups, were visualized using Graphpad Prism v9 software.
2.7. Expression Analysis Based on RNA-Seq Data
The RNA-seq data used in this study were obtained from two publicly accessible datasets in the NCBI Sequence Read Archive (SRA) under accession numbers SRP344288 (salt stress) [5] and SRP321496 (drought stress) [56]. The salt stress dataset comprised samples from roots and leaves of sour jujube treated with 0 (control), 100, and 200 mM NaCl for 3 and 8 days [5]. The drought stress dataset included leaf samples from diploid and tetraploid plants subjected to 0 (control), 6, 12, and 48 h of drought treatment [56]. Raw reads (SRA format) were first converted to FASTQ format via the SRA Toolkit v3.0.0. Following adapter trimming and quality filtering with Trimmomatic v0.39, read quality was assessed using FastQC v0.11.9. The resulting high-quality reads were then aligned to the sour jujube reference genome employing STAR v2.7.11b, which concurrently handled genome indexing and read mapping. The expression matrix of ZjNHX genes was extracted using the Kallisto Super Wrapper plugin in TBtools, and expression pattern heatmaps were subsequently generated with the same software.
2.8. Plant Materials and Treatments
Seeds of sour jujube were collected from Zanhuang County, Hebei Province, China. Plump and uniform healthy seeds were selected and surface-sterilized with a 6–14% sodium hypochlorite solution for 15 min, followed by six successive rinses with sterile distilled water. The sterilized seeds were soaked in water for two days at 30 °C in a constant-temperature incubator to promote germination. Subsequently, the seeds were sown in seedling trays filled with a mixture of vermiculite and nutrient soil at a volume ratio of 2:1. Seedlings were cultivated under conditions of 22–25 °C with a photoperiod of 16 h light/8 h dark. At 60 days after sowing, seedlings with uniform growth were transplanted into new plug trays. After a one-week acclimatization period, all seedlings were uniformly irrigated with a 150 mM NaCl solution for salt stress treatment. Leaf and root samples were collected at 0 (control), 3, 6, 12, and 24 h after irrigation. For each time point, three biological replicates were collected, with each replicate comprising pooled samples from five individual seedlings. All harvested samples were immediately frozen in liquid nitrogen and subsequently stored at –80°C until further analysis.
2.9. RNA Extraction and Gene Expression Analysis
Total RNA was isolated from each sample using an RNA extraction kit (Vazyme, Nanjing, China). The concentration and purity of the RNA were assessed with a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). First-strand cDNA synthesis was then carried out with the HiScript III RT SuperMix (+gDNA wiper) (Vazyme, Nanjing, China). For quantitative real-time PCR (qRT-PCR) analysis, the ubiquitin (UBQ) gene was selected as the endogenous control. Reactions were carried out on a QuantStudio 5 Real-Time PCR system (Thermo Fisher Scientific, Waltham, MA, USA) with ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China). All reactions were run in triplicate to ensure technical reproducibility. Gene relative expression levels were calculated using the 2^−ΔΔCt^ method [57]. Statistical analyses were conducted with SPSS v27.0. Differences among treatment groups were evaluated by one-way analysis of variance (ANOVA), followed by Tukey’s honestly significant difference (HSD) post-hoc test for multiple comparisons. A probability (p) value of less than 0.05 was regarded as indicating statistical significance. All graphical representations were generated using GraphPad Prism v9. Detailed information on the primer sequences employed in this study is provided in Supplementary Table S1.
3. Results
3.1. Identification and Physicochemical Property Analysis of the ZjNHX Gene Family
In the genome of Z. jujuba var. spinosa, this study identified eight members of the ZjNHX gene family (Supplementary Table S2) distributed across six chromosomes (Figure 1). Following their linear chromosomal positions, these genes were designated ZjNHX1 through ZjNHX8. Physicochemical characterization revealed significant variations in their protein properties. ZjNHX proteins ranged in length from 207 aa (ZjNHX4) to 1145 aa (ZjNHX6), with molecular weights spanning 22.47 kDa (ZjNHX4) to 126.83 kDa (ZjNHX6). Theoretical isoelectric points (pI) varied between 5.35 (ZjNHX2) and 8.75 (ZjNHX5), while instability indices ranged from 31.40 (ZjNHX3) to 42.52 (ZjNHX8). The grand average of hydropathy (GRAVY) values was uniformly positive (0.074–0.701), indicating the ZjNHX proteins were generally hydrophobic, which was consistent with the biochemical functions of NHXs as transmembrane Na^+^/H^+^ exchangers. Subcellular localization predictions indicated that ZjNHX1–ZjNHX5 and ZjNHX8 are likely vacuolar, whereas ZjNHX6 and ZjNHX7 are predicted to reside on the plasma membrane. Transmembrane helix predictions confirmed most proteins contain 9–12 TMs, consistent with their roles as membrane-bound transporters; however, ZjNHX4 uniquely has only 4 TMs (Table 1).
The secondary structures of the ZjNHX proteins demonstrated that α-helices and random coils dominate these proteins, with proportions ranging from 43.29% to 62.30%, 23.72% to 39.32%, respectively (Supplementary Table S3). Additionally, to further analyze their structural characteristics, we predicted and visualized the three-dimensional structures using SWISS-MODEL (Supplementary Figure S1). These models revealed conserved structural architectures, providing a structural basis for functional similarity.
3.2. Phylogenetic Analysis
To elucidate the evolutionary relationships of the ZjNHX proteins, a phylogenetic tree was constructed in this study based on 99 NHX protein sequences from Z. jujuba var. spinosa and nine other species (Figure 2, Supplementary Table S4). According to the phylogenetic tree, the 99 NHX proteins were clustered into three distinct clades, which were consistent with previous findings in Arabidopsis AtNHX proteins. Based on the established classification of AtNHX proteins, these three clades were named the vacuolar (Vac), endosomal (Endo), and plasma membrane (PM). Phylogenetic analysis revealed that the Vac clad contained the largest number of NHX proteins, followed by the Endo clad, while the PM clad comprised the fewest members. In Z. jujuba var. spinosa, the ZjNHX proteins were also classified into three groups. Specifically, ZjNHX6 and ZjNHX7 were located within the PM clade, ZjNHX2 was positioned in the Endo clade, and the remaining five ZjNHX proteins (ZjNHX1, ZjNHX3, ZjNHX4, ZjNHX5, and ZjNHX8) were all distributed in the Vac clade. It is noteworthy that within the Vac clade, the four ZjNHX proteins exhibit a closer evolutionary kinship with NHX homologs from dicotyledonous plants such as P. granatum, P. euphratica, V. vinifera, and C. pepo compared with those from monocotyledonous plants like rice and wheat. This pattern is similarly observed in both the Endo and PM clades, suggesting a shared evolutionary trajectory for NHX proteins among dicotyledonous plants.
3.3. Exon–Intron Analysis and Motif Configuration
To provide an in-depth exploration of the structural characteristics and evolutionary relationships of the ZjNHX genes in Z. jujuba var. spinosa, this study performed phylogenetic analysis, conserved motif analysis, and gene structure analysis of eight ZjNHX proteins. This combined figure showed characteristics of the NHX gene family (Figure 3). The results of the phylogenetic tree demonstrated that all members could be divided into three major clades, which was consistent with the aforementioned phylogenetic analysis. The conservative motif distribution of ZjNHX proteins was consistent with the phylogenetic tree. Hence, ZjNHX members within the same clade exhibited similar conserved motif compositions. Specifically, motifs 4, 3, and 1 were predominantly found in the Vac clade, whereas motifs 6, 9, and 10 were mainly present in the PM clade. Functional annotation based on the InterPro database revealed that motifs 1, 2, and 3 correspond to conserved regions within the Na^+^/H^+^ exchanger domain and are widely distributed in both the Vac and Endo clades. Interestingly, motif 3 contains the amiloride-binding site (FFI/LY/FLLPPI), a characteristic feature of vacuolar NHX proteins, which was absent in the PM clade (Supplementary Figure S2). Multiple sequence alignment further corroborated this result (Figure 4). Moreover, while this site is completely conserved in four Vac-clade proteins, it harbors a single amino acid substitution in ZjNHX3. Gene structure analysis further revealed that genes within the same group share similarities in their genetic architecture, specifically in terms of the number and length of exons and introns. The results show that the proteins in the Vac clade mostly possess exon numbers ranging from 12 to 14. In contrast, ZjNHX4 of this clade exhibits a markedly simplified configuration, containing only six exons, a trend that aligns with its reduced count of conserved motifs. ZjNHX2 in the Endo clad contains 22 exons. In the PM clade, both ZjNHX6 and ZjNHX7 contain 23 exons, representing the most complex structure. Collectively, this gene structure analysis corroborates the classification established by the phylogenetic analysis and demonstrates that members of the same evolutionary clade exhibit a high degree of structural conservation in both conserved motif composition and exon number.
3.4. Analysis of Duplication Events, Collinearity, and Selection Pressure
Gene duplication events serve as a primary driving force for genome evolution and gene family expansion. Accordingly, we analyzed the duplication events within the ZjNHX gene family in sour jujube. The results indicate that the evolution of the ZjNHX gene family was driven by various types of gene duplications (Supplementary Table S5). Specifically, ZjNHX1, ZjNHX2, and ZjNHX4 originated from dispersed duplications, and ZjNHX3, ZjNHX5, and ZjNHX8 are products of whole-genome duplication (WGD) or segmental duplication, while ZjNHX6 and ZjNHX7 arose from tandem duplications. Furthermore, we identified two syntenic gene pairs within the sour jujube genome: ZjNHX5-ZjNHX3 and ZjNHX5-ZjNHX8, localized to chromosomes 9, 2, and 12, respectively (Figure 5). This spatial arrangement provides additional support for their origin via segmental duplication events. To further investigate the evolutionary constraints acting on these genes, we first assessed substitution saturation for all syntenic gene pairs. The index of substitution saturation (Iss) was significantly lower than the critical value (Iss.c, p < 0.05) in all pairwise comparisons, indicating no significant saturation and thus supporting the reliability of the Ks values used for subsequent divergence time estimation (Supplementary Table S6). Based on this validation, we calculated the non-synonymous (Ka) to synonymous (Ks) substitution rate ratios for these homologous pairs. The resulting Ka/Ks ratios were significantly less than 1 for all pairs (Table 2), providing clear evidence of strong purifying selection during evolution. This mode of selection likely acts to preserve the critical functional structures of the NHX gene family. Additionally, divergence time estimation revealed that these homologous pairs were formed by segmental duplications at distinct time points. The divergence between ZjNHX5 and ZjNHX3 occurred approximately 55.411 million years ago (Mya), whereas the divergence between ZjNHX5 and ZjNHX8 took place earlier, around 59.228 Mya. To further elucidate the evolutionary dynamics of the NHX gene family in Z. jujuba var. spinosa, we performed interspecies synteny analysis between sour jujube and the model plants A. thaliana and O. sativa. The results (Figure 6, Supplementary Table S7) revealed five syntenic gene pairs between sour jujube and A. thaliana, and four pairs between sour jujube and O. sativa. The higher number of syntenic pairs with A. thaliana likely stems from their closer phylogenetic relationship, as both are dicotyledonous plants.
3.5. Cis-Element Analysis of ZjNHX Promoters
To further explore the transcriptional regulation mechanisms and potential biological functions of the ZjNHX genes in sour jujube, promoter sequences spanning 2000 bp upstream of the start codon ATG for each ZjNHX gene were extracted and analyzed using the PlantCARE database. The results revealed a total of 52 types of cis-acting regulatory elements (CAREs), which were classified into four functional categories: light response, stress response, phytohormone response, and growth/development regulation (Figure 7; Supplementary Table S8). Light-responsive elements were the most abundant, suggesting a potentially universal role for light signaling in regulating ZjNHX gene expression. Beyond their well-known function in photosynthesis, these elements also integrate light signals with ABA signaling networks to modulate stress adaptation. Among the stress-responsive elements, MYB and MYC binding sites were identified 35 and 31 times, respectively, and were distributed across all ZjNHX promoters. These elements function as core dehydration-responsive motifs and bHLH transcription factor binding sites, both of which are critical nodes in ABA-dependent and -independent stress signaling pathways. This enrichment highlights their crucial role in mediating responses to salt, drought and low temperature [13,58]. The anaerobic induction element (ARE) was present in all members except ZjNHX1, implying a potential involvement of most ZjNHX genes in adaptation to low-oxygen environments. Within the phytohormone-responsive category, the ethylene-responsive element (ERE) was widely distributed. Elements such as “as-1” (responsive to salicylic acid and auxin), CGTCA-motif, and TGACG-motif (both responsive to methyl jasmonate) were commonly found in all promoters except in ZjNHX5, suggesting that these genes may be broadly involved in multiple hormone signaling pathways. Furthermore, several ZjNHX promoters contained elements related to growth and development, including the O2-site, CCGTCC-box, and circadian elements, hinting at their possible roles in regulating physiological processes such as seed development. Collectively, the promoter regions of sour jujube ZjNHX genes are enriched with diverse cis-acting elements involved in stress responses and hormone regulation, suggesting that this gene family may play a significant role in integrating various environmental signals and mediating stress adaptation processes. While these findings are descriptive and based solely on computational prediction, they provide a useful foundation for generating hypotheses about the possible involvement of this gene family in integrating environmental signals and mediating stress adaptation. Experimental validation will be necessary to confirm the actual regulatory functions of these elements.
3.6. Expression Analysis of ZjNHXs Based on Transcriptomic Data Under Drought and Salt Stress
To elucidate the functional divergence of ZjNHX genes under abiotic stress, we analyzed RNA-seq data to characterize their expression profiles under salt and drought (Supplementary Tables S9–S11). Transcriptome data revealed distinct response patterns of different ZjNHX genes to salt stress (Figure 8A,B). Specifically, ZjNHX2, ZjNHX3, ZjNHX5, and ZjNHX6 were up-regulated in both roots and leaves after 3 and 8 days of exposure to 100 mM and 200 mM NaCl. Higher expression levels were observed under 200 mM NaCl compared to 100 mM at the same time points, indicating a concentration-dependent response to salt stress. In contrast, ZjNHX1, ZjNHX4, ZjNHX7, and ZjNHX8 did not show significant changes in expression under salt stress. Under drought stress, five ZjNHX genes (ZjNHX2, ZjNHX3, ZjNHX5, ZjNHX6, and ZjNHX7) exhibited up-regulation in diploid and autotetraploid sour jujube (Figure 8C,D). Among them, the relative expression of ZjNHX2 continuously increased with prolonged stress duration, peaking at 48 h. ZjNHX3, ZjNHX6, and ZjNHX7 showed an initial up-regulation followed by a decline. ZjNHX5 displayed a fluctuating expression pattern throughout the drought treatment. In contrast, ZjNHX1, ZjNHX4, and ZjNHX8 did not exhibit significant expression changes during the 48 h drought period. The distinct expression patterns of ZjNHX genes in sour jujube under salt and drought stress indicate their diverse functional roles.
3.7. Expression Analysis of ZjNHX Genes Under Salt Stress
Previous studies have confirmed that salt treatment can induce the expression of NHX genes, which play a crucial role in enhancing salt tolerance in various plants. Therefore, to further understand the potential functions of ZjNHX genes in response to salt stress, we analyzed their expression in roots and leaves within 24 h after treatment with 150 mM NaCl, using 0 h as the control (Figure 9). RT-qPCR analysis revealed that the relative expression levels of most ZjNHX genes were up-regulated under salt stress. Furthermore, these genes exhibited significant temporal and spatial specificity in roots and leaves. In the roots, the expression levels of ZjNHX1, ZjNHX6, and ZjNHX7 were rapidly up-regulated within 3 h of salt treatment. Notably, the differential expression of the ZjNHX1 gene was the most pronounced, with its expression level increasing approximately 40-fold compared to the control group within 3 h of salt stress, and it remained at a high level at 24 h. In contrast, ZjNHX4 and ZjNHX8 showed an upregulating trend within 12 h of salt treatment, while the expression of ZjNHX2, ZjNHX3, and ZjNHX5 was not significantly altered, maintaining stable levels throughout. In contrast, the relative expression level of ZjNHX1 in leaves initially decreased and then increased, peaking at 24 h with an approximately 2-fold increase. Interestingly, transcriptomic data indicated that ZjNHX1 exhibited almost no expression after 3 and 8 days of salt stress. It is therefore hypothesized that ZjNHX1 expression is significantly up-regulated in response to salt stress over a short period, particularly within one day, with a more pronounced response in roots compared to leaves. Meanwhile, the expression levels of the ZjNHX2 and ZjNHX3 genes in leaves showed a gradual increasing trend with prolonged salt stress duration, which is consistent with the transcriptomic data. In contrast, ZjNHX4, ZjNHX7, and ZjNHX8 exhibited a fluctuating expression pattern in leaves, characterized by an initial increase, followed by a decrease, and then another increase. These findings suggest that different ZjNHX gene members possess distinct response mechanisms to salt stress. The observed differential expression patterns may reflect their divergent functional roles in the salt stress response.
4. Discussion
The core function of the NHX gene family is to maintain cellular ion homeostasis via Na^+^/H^+^ antiport activity. Consequently, NHX proteins play pivotal roles in regulating intracellular pH, maintaining Na^+^/K^+^ balance, facilitating plant adaptation to osmotic stress, and enhancing salinity tolerance by promoting excess Na^+^ sequestration or efflux [9,18]. The number of the NHX gene family in most plants ranges from 7 to 35. For instance, eight AtNHX genes have been identified in A. thaliana, seven LjNHX genes in honeysuckle [59], ten in pomegranate [37], and thirty-five MsNHX genes in alfalfa [58]. In this study, eight NHX genes were identified from sour jujube. These variations in NHX gene numbers among plant species are likely attributable to gene duplication or loss events occurring in different NHX subfamilies during evolution.
Phylogenetic analysis classified the eight ZjNHX proteins into three clades: Vac (five members), Endo (one member), and PM (two members). The predominance of Vac-clade members is a feature shared with several other plants (e.g., A. thaliana [11], T. aestivum [60] and Pilocarpus microphyllus [26]) and is consistent with the hypothesis that vacuolar NHX isoforms contribute prominently to cellular ion homeostasis. In model species, phylogenetic classification generally correlates with subcellular localization. In the present study, however, an exception was observed: although ZjNHX2 clustered within the endosomal clade, in silico tools predicted its localization to the vacuole. Similar inconsistencies between phylogenetic placement and computational predictions have been documented in other plant species, including pomegranate [37], amaranth [61], and tomato [62]. These discrepancies likely reflect the inherent limitations of current prediction algorithms when applied to complex membrane proteins such as NHX transporters [63]. Therefore, the precise subcellular localization of ZjNHX2 in sour jujube requires further experimental validation.
The gene structures and motif compositions within the ZjNHX family are highly conserved among members of the same clade, supporting the phylogenetic classification. In sour jujube, exon numbers range from 6 to 14 in the Vac clade, 22 in the Endo clade, and 23 in the PM clade—a pattern broadly similar to that observed in poplar [64], tomato [62], and Arabidopsis [8], reflecting conserved evolutionary constraints across dicots. All identified ZjNHX proteins contain 9–12 predicted transmembrane domains, consistent with the typical architecture of Na^+^/H^+^ exchangers [65]. A notable structural feature is the amiloride-binding motif FF(I/L)(Y/F)LFLLPPI, historically considered characteristic of vacuolar NHX proteins [65,66]. In sour jujube, this motif is present in all five Vac-clade members and, interestingly, also in the single Endo-clade member (ZjNHX2). The occurrence of this site in endosomal isoforms has been documented in several other species [37,62,67] and may indicate partial functional conservation between Vac- and Endo-class transporters, though this remains speculative [18]. Within the Vac clade, ZjNHX3 harbors a single amino acid substitution within the motif (I→M; FFIYMLPPI). Similar conservative substitutions have been reported in other plants (e.g., Sorghum bicolor [68]), and given the hydrophobic nature of both residues, the overall protein structure and core transport function are likely preserved. Whether this substitution influences inhibitor sensitivity, cation selectivity, or regulatory properties is unknown and warrants experimental investigation. Importantly, this variant may represent a lineage-specific feature of sour jujube, but its adaptive significance cannot be inferred without functional assays. Taken together, the conserved yet subtly divergent structural features of ZjNHX genes—particularly the atypical amiloride-binding motif in ZjNHX3 and its presence in ZjNHX2—distinguish the sour jujube NHX family from those of model species. These observations generate specific hypotheses regarding isoform-specific transport properties and regulation, providing a focused entry point for mechanistic dissection in this stress-tolerant woody perennial.
Consistent with findings in other plant species [36], the ZjNHX genes in sour jujube exhibited distinct spatiotemporal expression patterns under salt stress. RT-qPCR analysis revealed that most ZjNHX members were up-regulated upon NaCl treatment, with particularly rapid and pronounced induction in roots. Notably, ZjNHX1 expression increased approximately 40-fold within 3 h and remained elevated at 24 h; ZjNHX7 and ZjNHX8 showed moderate up-regulation at early time points (5-fold and 2-fold, respectively). Similar early, strong root induction of vacuolar-type NHX genes has been documented in salt-tolerant accessions of Medicago truncatula [69], soybean [70], and rice [71]. Although such expression kinetics are often interpreted as evidence of vacuolar Na^+^ sequestration in roots—a well-characterized function of Vac-class NHX orthologs in model species [13,18]—the present data do not directly demonstrate this mechanism in sour jujube. The observed up-regulation is consistent with, but not proof of, a role in restricting Na^+^ translocation to shoots. Direct evidence, such as ion content analysis, subcellular localization, or transgenic manipulation, is required to establish whether the rapid induction of ZjNHX1, ZjNHX7, and ZjNHX8 indeed contributes to vacuolar compartmentalization and whole-plant salt tolerance. Nevertheless, the strong, tissue-specific responsiveness of these genes positions them as high-priority candidates for future functional studies aimed at dissecting the molecular basis of salt adaptation in sour jujube. Their expression profiles, together with the genomic resources provided here, offer a focused entry point for hypothesis-driven investigation in this stress-tolerant woody perennial.
Leaf-expressed ZjNHX genes also displayed salt-responsive expression patterns, with distinct temporal dynamics. ZjNHX1 showed transient upregulation in leaves within 24 h of stress onset (~2-fold), but transcriptome data indicated negligible expression at later time points (3 and 8 d). In contrast, ZjNHX2 and ZjNHX3 exhibited progressive, sustained upregulation under prolonged salt exposure, with higher transcript levels under 200 mM NaCl than under 100 mM NaCl. These divergent kinetics suggest that different ZjNHX members may contribute to salt responsiveness over different time scales. Comparable leaf expression patterns have been reported in sugar beet (BvNHX3–5) [72] and pomegranate (PgNHXs) [37], indicating that delayed but sustained upregulation of certain NHX isoforms in photosynthetic tissues is not unique to sour jujube. In model species, such expression changes have been associated with vacuolar Na^+^ compartmentalization, which is thought to alleviate Na^+^ toxicity in chloroplasts and contribute to osmotic adjustment [29,73]. However, whether the observed upregulation of ZjNHX2 and ZjNHX3 reflects analogous physiological roles in sour jujube remains correlative and requires direct experimental testing (e.g., ion localization, transgenic approaches). Taken together, the tissue- and time-dependent expression profiles of ZjNHX genes under salt stress are consistent with the hypothesis that sour jujube employs spatially and temporally coordinated transcriptional responses to salinity.
5. Conclusions
This study presents the first genome-wide identification and characterization of the NHX gene family in sour jujube. Eight ZjNHX genes were identified and classified into three subfamilies based on phylogeny, conserved motifs, and gene structures. Promoter analysis revealed an enrichment of stress- and hormone-responsive cis-regulatory elements, suggesting that these genes may be transcriptionally regulated under adverse conditions. Expression profiling by both transcriptome analysis and qRT-PCR demonstrated that several ZjNHX members are differentially regulated under salt stress in a tissue-specific and time-dependent manner, with ZjNHX1, ZjNHX2, and ZjNHX3 emerging as strong candidates for further functional studies. Nevertheless, the comprehensive data presented here provide a valuable genomic resource and a foundation for hypothesis-driven functional dissection of the NHX family in sour jujube and related woody perennials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Montanarella L. Pennock D. Mckenzie N. Alavipanah S.K. Alegre J. Alshankiti A. Arrouays D. Singh Aulakh M. Badraoui M. dos Santos Baptista Costa I. The Status of the World’s Soil Resources Technical Summary Food and Agriculture Organization of the United Nations Rome, Italy 2015
- 2Du L. Zhang Z. Chen Y. Wang Y. Zhou C. Yang H. Zhang W. Heterogeneous Impact of Soil Acidification on Crop Yield Reduction and Its Regulatory Variables: A Global Meta-Analysis Field Crops Res.202431910964310.1016/j.fcr.2024.109643 · doi ↗
- 3Munns R. Tester M. Mechanisms of Salinity Tolerance Annu. Rev. Plant Biol.20085965168110.1146/annurev.arplant.59.032607.09291118444910 · doi ↗ · pubmed ↗
- 4Wang Y. An Y. Yu J. Zhou Z. He S. Ru M. Cui B. Zhang Y. Han R. Liang Z. Different Responses of Photosystem II and Antioxidants to Drought Stress in Two Contrasting Populations of Sour Jujube from the Loess Plateau, China Ecol. Res.20163176177510.1007/s 11284-016-1384-5 · doi ↗
- 5He A. Ma Z. Li Y. Huang C. Yong J.W.H. Huang J. Spatiotemporal, Physiological and Transcriptomic Dynamics of Wild Jujube Seedlings under Saline Conditions Tree Physiol.20234383285010.1093/treephys/tpad 00136617163 · doi ↗ · pubmed ↗
- 6Gong L. Xie J. Luo Y. Qiu Z. Liu J. Mei N. Chen Z. Wang F. Huang Y. Guo J. Research Progress of Quality Control for the Seed of Ziziphus Jujuba Var. Spinosa (Bunge) Hu Ex HF Chow (Suan-Zao-Ren) and Its Proprietary Chinese Medicines J. Ethnopharmacol.202330711620410.1016/j.jep.2023.11620436720435 · doi ↗ · pubmed ↗
- 7Jia Q. Zheng C. Sun S. Amjad H. Liang K. Lin W. The Role of Plant Cation/Proton Antiporter Gene Family in Salt Tolerance Biol. Plant.20186261762910.1007/s 10535-018-0801-8 · doi ↗
- 8Brett C. Donowitz M. Rao R. Evolutionary Origins of Eukaryotic Sodium/Proton Exchangers Am. J. Physiol. Cell Physiol.2005288 C 223C 23910.1152/ajpcell.00360.200415643048 · doi ↗ · pubmed ↗