Lung Adenocarcinoma Promotes NETosis via the NPM1–TNFAIP6–CD44–SPP1 Axis
Renwang Liu, Zixuan Hu, Mingbiao Li, Shen Yang, Jianfang Wang, Zhanrui Zhang, Long Yang, Jun Chen

TL;DR
This study identifies a new pathway in lung adenocarcinoma that promotes cancer-linked NET formation, offering a potential target for treatment.
Contribution
The novel NPM1–TNFAIP6–CD44–SPP1 axis is identified as a key driver of NETosis in lung adenocarcinoma.
Findings
TNFAIP6 from lung adenocarcinoma induces NETosis by interacting with CD44 and increasing SPP1 levels.
NPM1 enhances TNFAIP6 transcription by binding to its promoter region.
The NPM1–TNFAIP6–CD44–SPP1 axis is critical for tumor-promoting NET formation.
Abstract
Neutrophil extracellular traps (NETs) promote cancer progression, but the regulatory mechanism underlying their formation remains unclear. This study shows that tumor necrosis factor alpha-inducible protein 6 (TNFAIP6) derived from lung adenocarcinoma induces NET formation (NETosis) in vitro and in vivo. TNFAIP6 interacts with CD44, leading to increased extracellular secreted phosphoprotein 1 (SPP1) levels and subsequent NETosis. Additionally, nucleophosmin 1 (NPM1) upregulates the transcriptional activation of TNFAIP6 by interacting with the −2000 to −1700 bp region of its promoter. Together, these results indicate that the NPM1–TNFAIP6–CD44–SPP1 axis is a critical regulator of NETosis in lung adenocarcinoma, highlighting this pathway as a potential therapeutic target for suppressing tumor progression. Background: While neutrophil extracellular traps (NETs) have been shown to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
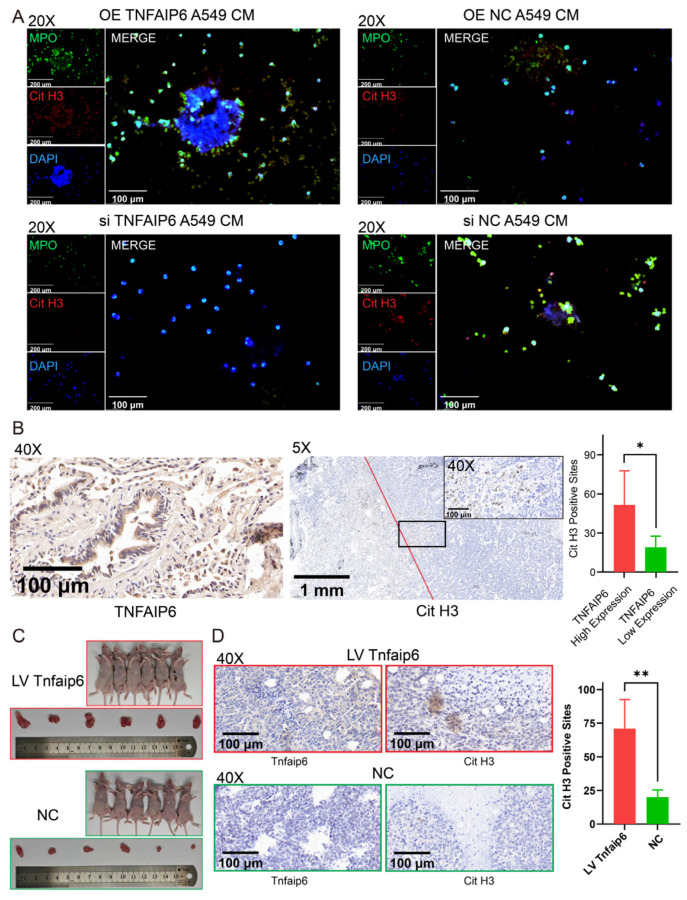 Figure 1
Figure 1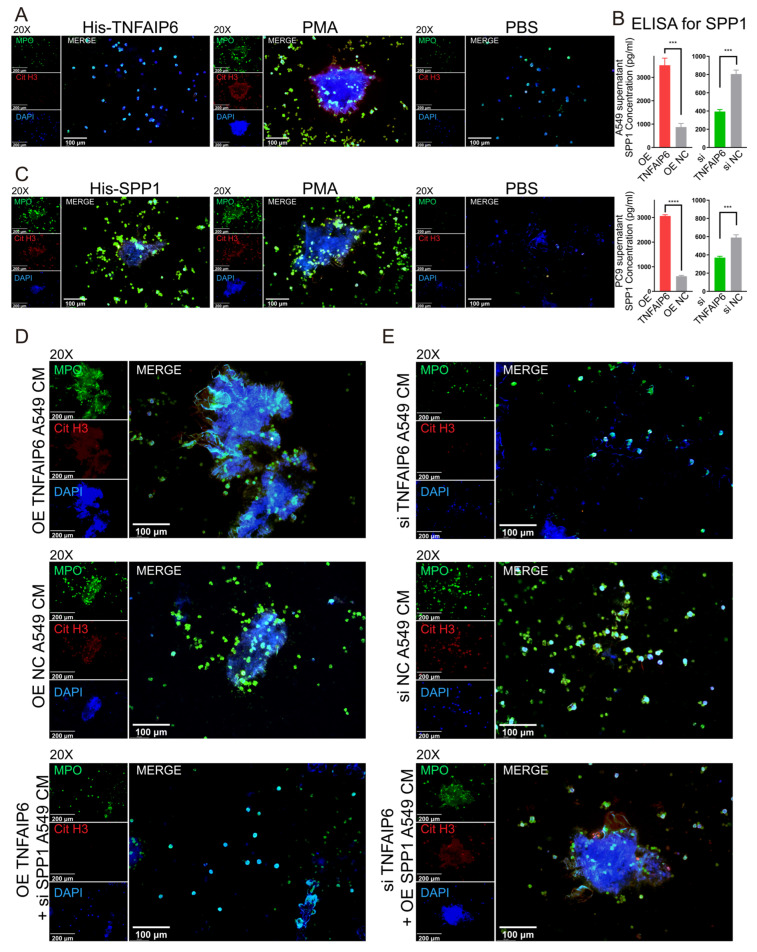 Figure 2
Figure 2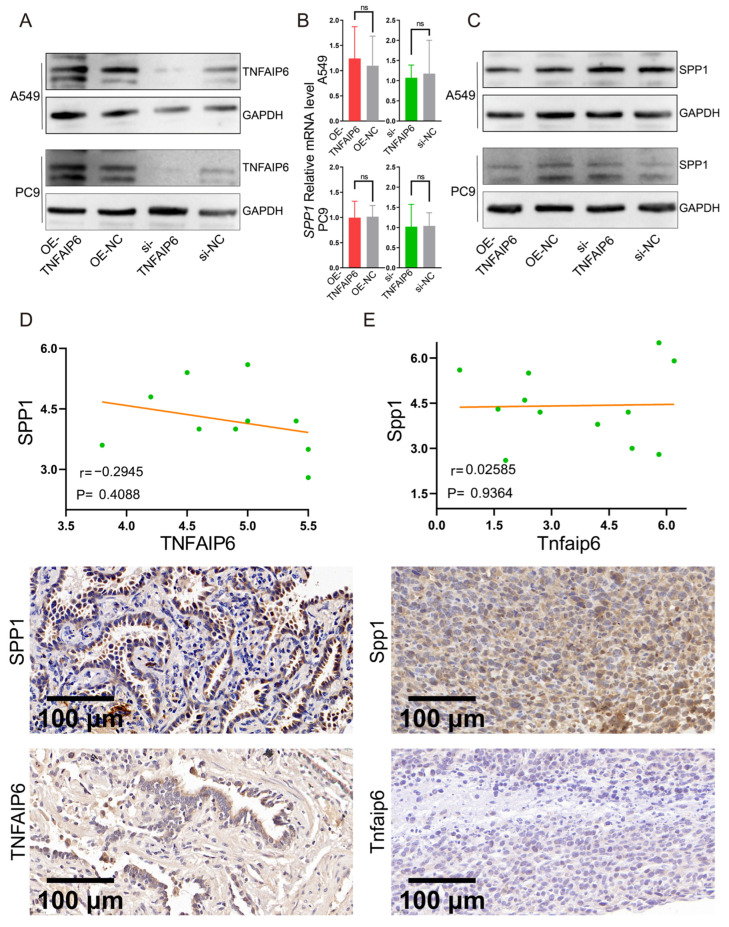 Figure 3
Figure 3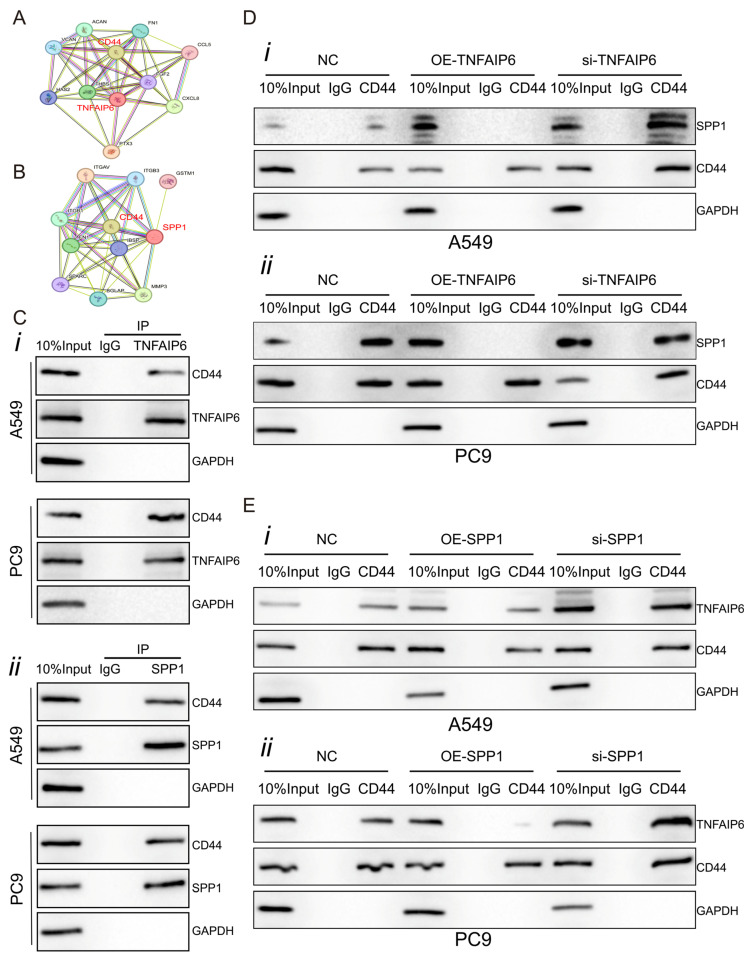 Figure 4
Figure 4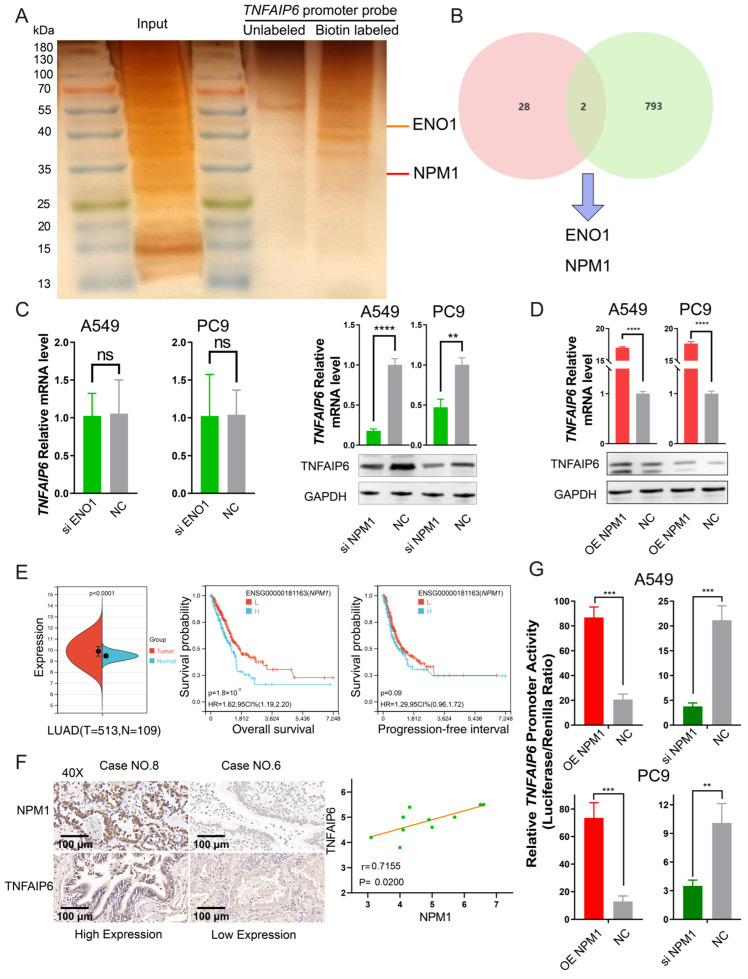 Figure 5
Figure 5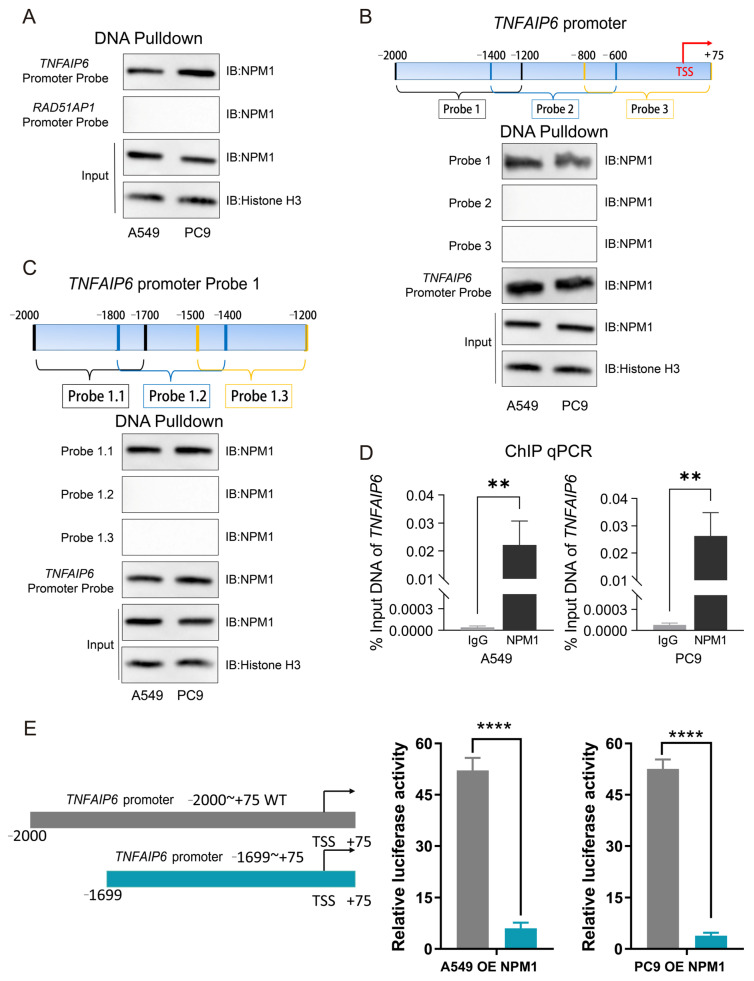 Figure 6
Figure 6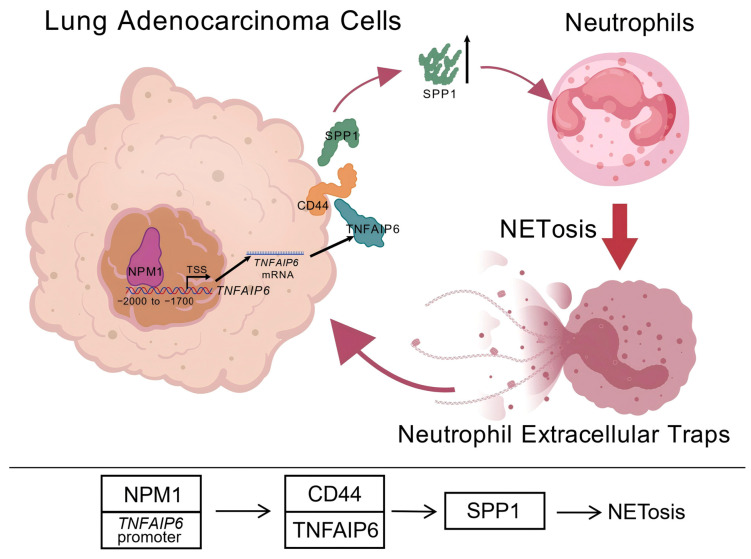 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Tianjin Key Medical Discipline Construction Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeutrophil, Myeloperoxidase and Oxidative Mechanisms · Immune cells in cancer · Cell Adhesion Molecules Research
1. Introduction
Lung adenocarcinoma has become the most prevalent histological subtype of lung cancer in recent decades [1,2]. Despite substantial advances in therapeutic strategies, many patients with lung adenocarcinoma still face unfavorable clinical outcomes due to disease progression and metastasis [3,4]. The tumor immune microenvironment (TIME) plays a critical role in this process and is extensively involved in key oncogenic events, including immune evasion, local recurrence, and distant metastasis [5,6,7].
Neutrophils represent a major cellular component of the TIME [8,9,10]. These cells can release neutrophil extracellular traps (NETs), which are composed of decondensed chromatin DNA, histones, and granule-derived proteins, through a specialized form of cell death known as neutrophil extracellular trap formation (NETosis) [11,12]. Accumulating evidence indicates that NETs within tumors, including lung adenocarcinoma, promote disease progression through multiple mechanisms [13,14,15]. NETs can ensnare circulating tumor cells to facilitate metastasis [16], release mediators that enhance angiogenesis [17], and suppress natural killer cell activity, thereby contributing to immune escape [18]. Consequently, inhibition of NET formation in tumors may mitigate their pro-tumorigenic effects and impede cancer progression.
Despite these observations, the mechanisms governing NETosis in the tumor context remain poorly defined. Previous work demonstrated that the expression of tumor necrosis factor alpha-inducible protein 6 (TNFAIP6) in lung adenocarcinoma is associated with neutrophil infiltration, polarization, and suppression of early apoptosis [19]. In addition, TNFAIP6 increases the proportion of neutrophils exhibiting “late-stage apoptosis,” as indicated by Annexin V and propidium iodide double positivity (Figure S1A) [19]. Because this phenotype may reflect necrosis, pyroptosis, or NETosis, neutrophils were treated with lung adenocarcinoma cell supernatants, and neutrophil-derived mediators were quantified by enzyme-linked immunosorbent assay (ELISA). TNFAIP6 did not significantly affect interleukin-18 (IL-18) or interleukin-1β (IL-1β) levels, but markedly increased myeloperoxidase (MPO) concentrations in neutrophil supernatants (Figure S1B–D). These findings suggest a potential role for TNFAIP6 in promoting NETosis rather than inflammatory cell death pathways.
Based on these preliminary observations, we hypothesized that TNFAIP6 expressed by lung adenocarcinoma cells plays a pivotal role in inducing NETosis within the TIME. Accordingly, we employed in vitro and in vivo approaches to systematically evaluate the effects of TNFAIP6 on NET formation, elucidate the underlying molecular mechanisms, and identify key functional regulatory sites.
2. Materials and Methods
2.1. Cell Culture, Transient Transfection, and Stable Transduction
Cell culture and genetic manipulations were performed as previously described [19,20,21]. The A549, H1975, BEAS-2B, Lewis lung carcinoma (LLC), and HEK293T cell lines were obtained from the American Type Culture Collection (ATCC). PC9 and H1299 cells were purchased from the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (Shanghai, China). All cell lines were maintained in RPMI 1640 medium (Gibco, Grand Island, NY, USA), except for HEK293T and LLC cells, which were cultured in DMEM (Gibco). All media were supplemented with 10% fetal bovine serum, 100 IU/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, Carlsbad, CA, USA).
Small interfering RNAs targeting TNFAIP6, secreted phosphoprotein 1 (SPP1), nucleophosmin 1 (NPM1), and ENO1, as well as a negative control siRNA, were purchased from RiboBio (Guangzhou, China). Overexpression plasmids for TNFAIP6, SPP1, and NPM1, together with lentiviral vectors encoding Tnfaip6 and corresponding control vectors, were constructed by GeneChem (Shanghai, China). To generate stable cell lines, LLC cells were transduced with LV-Tnfaip6 or LV-negative control according to the manufacturer’s instructions. Transient transfections were performed using Lipofectamine 2000 (Invitrogen) following the supplier’s protocol.
2.2. Neutrophil Isolation and Indirect Co-Culture
Neutrophil isolation and indirect co-culture assays were conducted as previously reported [19]. Briefly, neutrophils were isolated from the peripheral blood of healthy volunteers using Polymorphprep (Axis-Shield, Dundee, Scotland). Cell purity was assessed using a Fast Giemsa Stain Kit (Yeasen, Shanghai, China), prior to culture in RPMI 1640 medium (Gibco). Conditioned media from A549 and PC9 cells were collected 24 h after transfection and applied to freshly isolated neutrophils for 16 h.
2.3. Western Blotting and qPCR
Western blotting and quantitative PCR (qPCR) were performed as described previously [19]. The following primary antibodies were used: anti-TNFAIP6 (1:1000; Proteintech, Wuhan, China), anti-SPP1 (1:1000; Proteintech), anti-NPM1 (1:20,000; Proteintech), anti-CD44 (1:20,000; Proteintech), anti-histone H3 (1:2000; Proteintech), and anti-GAPDH (1:5000; Proteintech). All uncropped blots and molecular weight markers are shown in Supplementary Materials S1. For qPCR analysis, total RNA was extracted using TRIzol reagent (Invitrogen), and complementary DNA was synthesized with the PrimeScript RT Reagent Kit (TaKaRa, Beijing, China). Primer sequences were reported previously and are provided in Table S1 [19,22].
2.4. Immunofluorescence Assay
Immunofluorescence assays were performed as previously described [23]. Briefly, cells cultured on coverslips were fixed with 4% paraformaldehyde for 30 min, permeabilized with 0.5% Triton X-100 for 15 min, and blocked with 1% bovine serum albumin in phosphate-buffered saline for 1 h. Samples were incubated with primary antibodies overnight at 4 °C, followed by incubation with Alexa Fluor-conjugated secondary antibodies for 1 h at room temperature. Nuclei were counterstained with DAPI (Sigma-Aldrich, St. Louis, MO, USA) for 5 min, and images were acquired using a fluorescence microscope. Primary antibodies included anti-citrullinated histone H3 (Histone H3 [citrulline Arg17, citrulline Arg2, citrulline Arg8]; 1:200; NB100-57135, Novus, Chesterfield, MO, USA) and anti–myeloperoxidase (MPO; 5 μg/mL; AF3667, R&D Systems, Minneapolis, MN, USA).
2.5. Immunohistochemistry
Immunohistochemical staining was performed as described previously [19]. Briefly, tissue sections were deparaffinized, subjected to antigen retrieval in 5 mM Tris-HCl buffer using microwave heating for 10 min, and treated with 3% hydrogen peroxide to quench endogenous peroxidase activity. After serum blocking, the sections were incubated with primary antibodies overnight at 4 °C, followed by incubation with horseradish peroxidase-conjugated secondary antibodies for 30 min at room temperature. Signal development was achieved using diaminobenzidine, and sections were counterstained with hematoxylin.
The primary antibodies used included anti-citrullinated histone H3 (Histone H3 [citrulline Arg17, citrulline Arg2, citrulline Arg8]; 1:200; NB100-57135, Novus), anti-SPP1 (1:250; Proteintech), anti-TNFAIP6 (1:250; Proteintech), and anti-NPM1 (1:250; Proteintech). For citrullinated histone H3 (Cit H3) staining, positive sites were quantified by counting. For all other markers, staining was evaluated using a semiquantitative scoring system independently applied by two pathologists, based on staining intensity (0–3) and the proportion of positive cells (1–4), with the sum yielding a final score ranging from 1 to 7.
2.6. Syngeneic Tumor Model
To examine direct interactions between neutrophils and tumor cells in the absence of adaptive immune influences, syngeneic tumor models were established using C57BL/6 nude mice. LLC cells stably overexpressing Tnfaip6 were generated as described above. A total of 5 × 10^5^ LLC cells suspended in 50% Matrigel (Corning, Corning, NY, USA) were subcutaneously injected into the left lower abdomen of each mouse (n = 6 per group). Deoxyribonuclease I (DNase I, 0.1 U per mouse) was intraperitoneally injected on day 6 after inoculation and once daily for 3 consecutive days to verify the role of NETs in the influence of TNFAIP6 on tumor growth. Tumor length and width were measured every 3 days using a vernier caliper. The mice were euthanized 21 days after implantation, and tumors were excised for analysis. TNFAIP6 expression in tumor tissues was verified by immunohistochemistry (Figure S2D).
2.7. Enzyme-Linked Immunosorbent Assay
Following the collection of culture supernatants, the concentrations of SPP1, MPO, and IL-1β (MultiSciences, Hangzhou, China), IL-18 (Proteintech), as well as Cit H3 (Cayman, Ann Arbor, MI, USA), were measured using ELISA kits according to the manufacturers’ instructions. Standard curves were generated using a four-parameter logistic (4PL) regression model implemented in Python 3.13.2. Supernatants used for MPO measurement were diluted 100-fold, those for Cit H3 measurement were diluted 10-fold, and samples for all other analytes were analyzed without dilution.
2.8. Co-Immunoprecipitation
Co-immunoprecipitation (Co-IP) assays were performed as previously described [24]. To standardize total protein input across samples, cells were trypsinized and counted using a TC20 automated cell counter (Bio-Rad, Hercules, CA, USA), and the volume of RIPA lysis buffer was adjusted accordingly. Cell lysates were incubated with antibodies against SPP1 (22952-1-AP, Proteintech), TNFAIP6 (ab267469, Abcam, Cambridge, UK), CD44 (15675-1-AP, Proteintech), or normal IgG (Beyotime, Shanghai, China) at 4 °C for 24 h. Protein A/G agarose beads (Beyotime) were then added, and the samples were gently rotated for an additional 3 h at 4 °C. After centrifugation, the supernatant was discarded, and the beads were washed five times with lysis buffer. Immunoprecipitated proteins were eluted in SDS loading buffer, boiled at 100 °C for 10 min, and analyzed by Western blotting following centrifugation.
2.9. Recombinant Protein Preparation
All plasmids were constructed by GeneChem. His-tagged TNFAIP6 and SPP1 expression plasmids were generated for eukaryotic expression and transfected into HEK293T cells. Because TNFAIP6 and SPP1 are secreted proteins, culture supernatants were collected for protein purification. His-tagged fusion proteins were purified using a His-tag Protein Purification Kit (IDA–Ni agarose magnetic beads; Beyotime) according to the manufacturer’s instructions.
2.10. DNA Pulldown Assay
Biotin-labeled and unlabeled DNA probes were amplified by PCR using the primers listed in Table S1. PCR products were resolved by 1% agarose gel electrophoresis, excised under UV illumination, and purified using a gel DNA extraction kit (TaKaRa, Kusatsu, Japan).
For DNA pulldown assays, purified DNA probes were incubated with prewashed streptavidin magnetic beads (Solarbio, Beijing, China) for 30 min at room temperature. After two washes, bead–probe complexes were incubated with nuclear extracts at 4 °C for 4 h under gentle rotation. Following elution and centrifugation, supernatants were subjected to Western blot analysis.
2.11. Silver Staining and Liquid Chromatography–Mass Spectrometry
Proteins captured by TNFAIP6 promoter probe pulldown were visualized by silver staining using a fast silver stain kit (Beyotime) according to the manufacturer’s instructions. Nuclear extracts served as input controls, and pulldown products obtained with unlabeled probes were used as negative controls. Protein identification was performed by liquid chromatography–mass spectrometry (LC–MS) at Lumingbio (Shanghai, China). Briefly, protein samples were digested and desalted, separated using an EASY-nLC 1200 nano-HPLC system (Thermo Scientific, Waltham, MA, USA), and analyzed on a Fusion mass spectrometer (Thermo Scientific) in data-dependent acquisition mode. Data were processed using Proteome Discoverer software (version 2.5) for database searching. The raw data have been deposited in Figshare (https://doi.org/10.6084/m9.figshare.31033222).
2.12. Bioinformatics Analysis and Prediction
Differential gene expression and survival analyses were conducted using The Cancer Genome Atlas (TCGA) datasets with R software (version 3.6.4), as previously described [21]. Optimal gene expression cutoffs were determined using the MaxStat package, and differences in overall survival and the progression-free interval were analyzed using the survfit function. Lists of known transcription factors were obtained from the TRRUST database [25]. Protein–protein interaction predictions were performed using the STRING database [26]. Protein structures and predicted interaction interfaces for SPP1, TNFAIP6, and CD44 were generated using the AlphaFold Server [27].
2.13. Dual-Luciferase Reporter Assay
Wild-type and truncated fragments of the TNFAIP6 promoter were cloned into the GV238 firefly luciferase reporter vector by GeneChem. Plasmids were transfected into cells using Lipofectamine 3000 (Invitrogen). A Renilla luciferase-expressing plasmid (CV045) was co-transfected as an internal control for normalization. Forty-eight hours after transfection, luciferase activities were measured using a GloMax^®^ 20/20 luminometer (Promega, Madison, WI, USA) in accordance with the manufacturer’s instructions.
2.14. Chromatin Immunoprecipitation-qPCR
Chromatin immunoprecipitation (ChIP) assays were performed using the BeyoChIP™ ChIP Assay Kit (Beyotime) following the manufacturer’s protocol. Briefly, cells were cross-linked with formaldehyde for 10 min, lysed on ice, and sonicated to shear chromatin DNA. Immunoprecipitation was conducted using an anti-NPM1 antibody (10306-1-AP, Proteintech) or normal IgG as a control, followed by overnight incubation at 4 °C with rotation. Protein A/G magnetic beads conjugated with salmon sperm DNA (80 μL) were then added, and samples were rotated for an additional 1 h at 4 °C. Bead-bound complexes were sequentially washed five times with the provided buffers. After elution, cross-links were reversed by incubation with 5 M NaCl at 65 °C for 4 h. DNA was subsequently purified and analyzed by quantitative PCR. The primer sequences used for ChIP-qPCR are listed in Table S1.
2.15. Protein Degradation and mRNA Stability Assay
For protein degradation assays, cells were treated with cycloheximide (CHX; MedChemExpress, Monmouth Junction, NJ, USA) at a final concentration of 100 μg/mL to inhibit protein synthesis. The cells were harvested 0, 3, 6, and 9 h after CHX treatment, and cell lysates were analyzed using Western blotting to assess TNFAIP6 protein levels. Band intensities were quantified using ImageJ(version 1.53e) software. Protein half-life was calculated using the formula: K = [ln(C0) − ln(C9)]/(t9 − t0); t1/2 = 0.693/K.
For mRNA stability assays, actinomycin D (Psaitong, Zaozhuang, China), an inhibitor of RNA transcription, was added to the culture medium at a final concentration of 200 nM. Cells were collected 0, 3, 6, 9, and 12 h after treatment for RNA extraction. Tnfaip6 mRNA levels at each time point were quantified by qPCR. The mRNA half-life was calculated using the formula K = [ln(C0) − ln(C12)]/(t12 − t0); t1/2 = 0.693/K.
2.16. Statistical Analysis and Data Visualization
Statistical analyses were performed using R software (version 3.6.4) and GraphPad Prism (version 9.5), depending on the dataset. Student’s t tests were applied to data with equal variance, whereas Mann–Whitney U tests were used for data with unequal variance. Pearson correlation coefficients were used for correlation analyses, and survival differences were assessed using the log-rank test. Statistical significance was defined as p < 0.05. Protein–protein interactions were predicted using the STRING database [26]. The schematic diagram in this study was created using BioGDP.com [28].
3. Results
3.1. TNFAIP6 in Lung Adenocarcinoma Promotes NETosis in Neutrophils
To investigate the role of TNFAIP6 in NETosis, immunofluorescence staining was performed to detect NET-specific markers, including Cit H3 and MPO, in neutrophils cultured with lung adenocarcinoma-conditioned medium. Overexpression of TNFAIP6 in A549 and PC9 cells markedly enhanced NET formation in co-cultured neutrophils, whereas TNFAIP6 knockdown significantly suppressed NETosis (Figure 1A and Figure S2A). ELISAs also revealed that the overexpression of TNFAIP6 in A549 and PC9 cells significantly increased the Cit H3 concentration in co-cultured neutrophils, whereas TNFAIP6 knockdown decreased it and BEAS-2B had no such effect (Figure S2B).
In addition, ten freshly obtained lung adenocarcinoma specimens, collected previously [19], were subjected to immunohistochemical staining for Cit H3. The use of these specimens was approved by the Ethics Committee of Tianjin Medical University General Hospital. The results revealed a positive correlation between TNFAIP6 expression and NET formation in human lung adenocarcinoma tissues (Figure 1B). To minimize potential interspecies interactions and exclude the influence of adaptive immunity, a syngeneic tumor (SynT) model was established using the LLC cell line and C57BL/6 nude mice. In this in vivo model, Tnfaip6 expression significantly promoted tumor growth (Figure 1C) and increased NET formation within tumor tissues (Figure 1D). Meanwhile, DNase I treatment significantly abrogated its tumor-promoting effect (Figure S2C).
3.2. Secreted TNFAIP6 Promotes NET Formation via SPP1 Rather than Directly
TNFAIP6 is a secreted protein that has been reported to suppress immune responses in myocardial infarction and tissue injury repair [29,30] and, according to limited evidence, may also contribute to tumorigenesis and cancer progression [31,32]. Given its secretory nature and immunomodulatory functions, TNFAIP6 protein was ectopically expressed, purified under native conditions, and applied directly to neutrophils. This treatment did not induce NET formation, indicating that TNFAIP6 does not directly stimulate NETosis in neutrophils (Figure 2A).
Previous label-free quantitative mass spectrometry (LFQ-MS) analyses suggested that TNFAIP6 increases the concentration of secreted SPP1 in lung adenocarcinoma cell supernatants [19]. This observation was further validated by ELISA in both A549 and PC9 cells (Figure 2B; standard curves are shown in Figure S2E). Immunofluorescence staining demonstrated that recombinant His-tagged SPP1 directly induced NETosis in neutrophils (Figure 2C). Consistently, SPP1 knockdown in TNFAIP6-overexpressing A549 and PC9 cells abrogated the TNFAIP6-mediated enhancement of NET formation (Figure 2D and Figure S3A). Conversely, SPP1 overexpression rescued the inhibitory effects of TNFAIP6 silencing on NETosis in neutrophils co-cultured with A549 and PC9 cells (Figure 2E and Figure S3B).
3.3. TNFAIP6 Does Not Alter SPP1 Expression
A549 and PC9 cells were transiently transfected with TNFAIP6 overexpression or silencing constructs, and transfection efficiency was confirmed by Western blotting (Figure 3A). Quantitative PCR and Western blot analyses demonstrated that neither SPP1 mRNA nor protein expression was significantly affected by modulation of TNFAIP6 levels (Figure 3B,C). Consistently, SPP1 protein expression did not correlate with TNFAIP6 levels in human lung adenocarcinoma tissue samples (Figure 3D). Moreover, in the SynT model, Tnfaip6 overexpression did not result in increased SPP1 protein expression (Figure 3E).
3.4. TNFAIP6 Interacts with CD44 and Affects Extracellular SPP1 Availability
Based on predictions from the STRING database [26], both TNFAIP6 and SPP1 were predicted to interact with CD44 (Figure 4A,B). These predicted interactions were validated by co-immunoprecipitation followed by Western blot analysis (Figure 4C). Notably, overexpression of TNFAIP6 in A549 and PC9 cells markedly reduced SPP1–CD44 interactions, whereas TNFAIP6 silencing enhanced this interaction (Figure 4D). Consistent results were obtained when SPP1 expression was manipulated in lung adenocarcinoma cells, suggesting that TNFAIP6 and SPP1 mutually influence their association with CD44 (Figure 4E).
3.5. NPM1 Upregulates the Transcriptional Activation of TNFAIP6
Previous work demonstrated that TNFAIP6 expression is elevated at both the mRNA and protein levels in lung cancer tissues [19]. Consistent with these findings, TNFAIP6 expression was significantly increased at the mRNA and protein levels in A549 and PC9 cells compared with BEAS-2B cells (Figure S4A,B). No significant differences were observed in TNFAIP6 mRNA degradation rates (Figure S4C) or protein stability (Figure S4D) among A549, PC9, and BEAS-2B cells. Notably, dual-luciferase reporter assays demonstrated enhanced transcriptional activation of the TNFAIP6 promoter in lung adenocarcinoma cells, indicating transcriptional upregulation of TNFAIP6 (Figure S4E).
Given these observations, TNFAIP6 promoter probes were constructed and a DNA pulldown assay coupled with mass spectrometry was performed; silver staining of the pulldown products is shown in Figure 5A. NPM1 was identified as a candidate regulator binding to the TNFAIP6 promoter by intersecting the mass spectrometry results with known transcription factors curated in the TRRUST database [25] (Figure 5B). Quantitative PCR and Western blot analyses further confirmed that NPM1 upregulates TNFAIP6 expression at both the mRNA and protein levels (Figure 5C,D).
Consistent with these findings, bioinformatic analyses based on TCGA demonstrated that NPM1 is overexpressed in lung adenocarcinoma tissues and is associated with poorer patient prognosis (Figure 5E). At the protein level, NPM1 expression was positively correlated with TNFAIP6 expression in lung adenocarcinoma tissues (Figure 5F). Moreover, overexpression or silencing of NPM1 in A549 and PC9 cells significantly increased or decreased, respectively, the luciferase activity driven by the TNFAIP6 promoter (Figure 5G).
3.6. NPM1 Interacts with the −2000 to −1700 bp Region of the TNFAIP6 Promoter
The association of NPM1 with the TNFAIP6 promoter in A549 and PC9 cells was first confirmed by DNA pulldown followed by Western blot analysis (Figure 6A). To define the region of interaction, the TNFAIP6 promoter was divided into three fragments, revealing NPM1 association within the −2000 to −1200 bp region (Figure 6B). This region was further subdivided, and the NPM1 interaction was localized to the −2000 to −1700 bp segment (Figure 6C). Consistently, chromatin immunoprecipitation followed by quantitative PCR demonstrated significant enrichment of NPM1 at the −2000 to −1700 bp region of the TNFAIP6 promoter (Figure 6D). Dual-luciferase reporter assays further confirmed that NPM1-mediated transcriptional activation of TNFAIP6 depends on this −2000 to −1700 bp promoter region (Figure 6E).
4. Discussion
The present study demonstrates that lung adenocarcinoma cells directly promote NET formation by upregulating TNFAIP6 (Figure 1). Mechanistically, TNFAIP6 interacts with CD44 in lung adenocarcinoma cells, leading to increased extracellular levels of SPP1 (Figure 3 and Figure 4). SPP1 subsequently induces NET formation in neutrophils (Figure 2B–E), rather than TNFAIP6 directly triggering NETosis (Figure 2A). Upstream analyses further clarified the mechanism underlying TNFAIP6 overexpression in lung adenocarcinoma, attributing it to transcriptional activation (Figure S4). Specifically, NPM1 was shown to enhance TNFAIP6 transcription by interacting with the −2000 to −1700 bp region of its promoter (Figure 5 and Figure 6). A schematic summary of the NPM1–TNFAIP6–CD44–SPP1 axis uncovered in this study is presented in Figure 7.
NETs were first described in 2004 by Volker Brinkmann as neutrophil-derived reticular structures that trap and eliminate invading microorganisms [33]. Increasing evidence has since implicated NETs in cancer progression across multiple malignancies, including lung adenocarcinoma [34,35,36,37,38]. Despite these observations, the mechanisms governing NET formation within tumors remain incompletely understood. Tumor cells have been reported to directly induce NETosis in neutrophils. For example, melanoma and colon carcinoma cells secrete CXCR1 and CXCR2 agonists, particularly IL-8, to stimulate NET formation [18]. Consistent with these findings, our results suggested that lung adenocarcinoma cells directly induce NETosis within the TIME. Notably, this work provides evidence that this process is dependent on TNFAIP6 upregulation.
The role of TNFAIP6 within the TIME appears to be multifaceted. Previous work showed that TNFAIP6 upregulation in lung adenocarcinoma markedly inhibits early neutrophil apoptosis, as silencing TNFAIP6 in A549 cells increased the proportion of neutrophils in the Annexin V-positive, propidium iodide-negative quadrant [19]. In contrast, the current study demonstrates a NETosis-promoting role for TNFAIP6 in lung adenocarcinoma (Figure 1 and Figure 2). Rather than being contradictory, these findings underscore the complex regulatory functions of TNFAIP6 in tumor-associated neutrophils. TNFAIP6 may simultaneously promote neutrophil survival, either directly or through indirect mechanisms, while interacting with CD44 to enhance the extracellular availability of SPP1 and thereby induce NETosis. Together, these actions finely tune neutrophil responses and reshape the TIME. It should be noted that the present study focuses specifically on the role of TNFAIP6 in NETosis, and the broader regulatory network governing its functions in the tumor context warrants further investigation.
Meanwhile, the NETosis-promoting effects of TNFAIP6 were found to be dependent on extracellular SPP1 (Figure 2). Consistent with the report by Shen et al., SPP1 directly induced NET formation in neutrophils (Figure 2C) [39]. Notably, TNFAIP6 did not regulate SPP1 expression (Figure 3); instead, it increased the extracellular availability of SPP1 by interacting with CD44 in lung adenocarcinoma cells (Figure 4). These findings suggest a potential competitive relationship among TNFAIP6, CD44, and SPP1. Competitive protein–protein interactions are common in diverse biological processes [40], and the present results raise the possibility that such a mechanism underlies the TNFAIP6–CD44–SPP1 axis. However, whether TNFAIP6 mediates this effect through steric hindrance or induces conformational changes upon interactions remains unclear. Furthermore, it is unknown whether other proteins or molecules participate in this process, and whether the elevation in extracellular SPP1 results from enhanced secretion, membrane dissociation, or increased protein stability following TNFAIP6–CD44 interaction. These questions warrant further investigation.
With respect to upstream regulation, we found that NPM1 interacted with the −2000 to −1700 bp region of the TNFAIP6 promoter (Figure 6). The role of NPM1 as a transcriptional regulator in lung adenocarcinoma has rarely been reported [41]. Moreover, this interaction region is distal to the transcription start site (TSS) and lacks canonical promoter elements, such as TATA, GC, or CAAT boxes, making it difficult to classify NPM1 as a classical transcription factor in this context. Instead, NPM1 is more commonly recognized as a transcriptional co-regulator. For instance, in glioblastoma, NPM1 forms a complex with RUNX1 to bind upstream of a cis-regulatory element of FOSL2, thereby maintaining chromatin accessibility and promoting transcription [42]. Similarly, the present study identified NPM1 binding at a moderately distal site relative to the TSS, suggesting that TNFAIP6 transcription may be enhanced through an analogous co-regulatory mechanism.
In addition, the −2000 to −1700 bp region of the TNFAIP6 promoter contains guanine-rich sequences (e.g., 5′-TTGGGAGAGGGGAGAAAGAA-3′) that may form G-quadruplex structures upon DNA unwinding. As reviewed by Varshney et al., G-quadruplexes play critical roles in transcriptional regulation [43]. For example, their stable formation has been shown to repress c-MYC transcription [44]. Numerous factors modulate the dynamic equilibrium of G-quadruplex structures in eukaryotic gene regulation [45]. Thus, NPM1 may promote TNFAIP6 transcription by directly or indirectly influencing G-quadruplex formation within this promoter region.
The present study has several limitations. First, while our Co-IP data suggested an association between TNFAIP6 and CD44, these experiments did not establish direct competition or displacement of SPP1. Moreover, whether the TNFAIP6–CD44 interaction increases extracellular SPP1 via enhanced secretion or other mechanisms remains to be determined. Second, although NPM1 was identified as a transcriptional regulator of TNFAIP6, the present study did not systematically screen for other transcription factors or regulatory elements that may also contribute to TNFAIP6 expression in lung adenocarcinoma. It therefore remains unclear whether NPM1 acts independently or cooperatively with other factors to modulate TNFAIP6 transcription. Third, the human tissue analyses were based on a relatively small cohort (n = 10), which limits statistical power and the generalizability of conclusions regarding clinical relevance. Validation in larger, independent patient cohorts is warranted. Fourth, while this study provides mechanistic insights using in vitro and in vivo models, the heterogeneity of patient tumors and the tumor microenvironment may influence the extent to which the NPM1–TNFAIP6–CD44–SPP1 axis is activated. Further validation using patient-derived xenografts or additional orthotopic models would help to confirm the translational relevance of these findings. Addressing these questions in future studies will contribute to a more comprehensive understanding of how TNFAIP6-mediated NETosis regulates tumor progression.
5. Conclusions
This study suggests that NPM1 upregulates the transcriptional activation of TNFAIP6 in lung adenocarcinoma cells. TNFAIP6 then interacts with CD44 in these cells, leading to increased extracellular availability of SPP1 and subsequent induction of NETosis. Specifically, NPM1 interacts with the −2000 to −1700 bp region of the TNFAIP6 promoter relative to the TSS. Despite certain limitations, these findings establish a novel conceptual and mechanistic framework for the NPM1–TNFAIP6–CD44–SPP1 axis in promoting NET formation within the TIME. Therapeutic disruption of the interaction of NPM1 with the TNFAIP6 promoter, such as via small-molecule inhibitors, and/or interference with TNFAIP6–CD44 interactions via blocking antibodies, may attenuate tumor-induced NETosis and thereby suppress tumor progression. These findings may provide a conceptual basis for exploring whether targeting this axis could have therapeutic relevance, although further studies are needed to validate this possibility.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kratzer T.B. Bandi P. Freedman N.D. Smith R.A. Travis W.D. Jemal A. Siegel R.L. Lung cancer statistics, 2023 Cancer 20241301330134810.1002/cncr.3512838279776 · doi ↗ · pubmed ↗
- 2Siegel R.L. Kratzer T.B. Giaquinto A.N. Sung H. Jemal A. Cancer statistics, 2025 CA Cancer J. Clin.202575104510.3322/caac.2187139817679 PMC 11745215 · doi ↗ · pubmed ↗
- 3Cascone T. Awad M.M. Spicer J.D. He J. Lu S. Sepesi B. Tanaka F. Taube J.M. Cornelissen R. Havel L. Perioperative Nivolumab in Resectable Lung Cancer N. Engl. J. Med.20243901756176910.1056/NEJ Moa 231192638749033 · doi ↗ · pubmed ↗
- 4Wu Y.L. Dziadziuszko R. Ahn J.S. Barlesi F. Nishio M. Lee D.H. Lee J.S. Zhong W. Horinouchi H. Mao W. Alectinib in Resected ALK-Positive Non-Small-Cell Lung Cancer N. Engl. J. Med.20243901265127610.1056/NEJ Moa 231053238598794 · doi ↗ · pubmed ↗
- 5Hanahan D. Weinberg R.A. Hallmarks of cancer: The next generation Cell 201114464667410.1016/j.cell.2011.02.01321376230 · doi ↗ · pubmed ↗
- 6Wang X. Bai H. Zhang J. Wang Z. Duan J. Cai H. Cao Z. Lin Q. Ding X. Sun Y. Genetic Intratumor Heterogeneity Remodels the Immune Microenvironment and Induces Immune Evasion in Brain Metastasis of Lung Cancer J. Thorac. Oncol.20241925227210.1016/j.jtho.2023.09.27637717855 · doi ↗ · pubmed ↗
- 7Chen J. Huang Z. Chen Y. Tian H. Chai P. Shen Y. Yao Y. Xu S. Ge S. Jia R. Lactate and lactylation in cancer Signal Transduct. Target. Ther.2025103810.1038/s 41392-024-02082-x 39934144 PMC 11814237 · doi ↗ · pubmed ↗
- 8Mellman I. Chen D.S. Powles T. Turley S.J. The cancer-immunity cycle: Indication, genotype, and immunotype Immunity 2023562188220510.1016/j.immuni.2023.09.01137820582 · doi ↗ · pubmed ↗