Correlation Analysis of BLTP1 (KIAA1109) and KIF27 Gene Polymorphisms with Wool Traits in Subo Merino Sheep
Qingfa Yan, Sen Tang, Asma Anwar, Gvlnigar Amar, Yaqian Wang, Wenna Liu, Cuiling Wu, Xuefeng Fu

TL;DR
This study identifies gene mutations in Subo Merino sheep that affect wool traits like crimp number and staple length, offering potential markers for breeding.
Contribution
The study discovers specific SNPs in BLTP1 and KIF27 genes linked to wool traits in Subo Merino sheep, providing new molecular markers for breeding.
Findings
SNP1 in BLTP1 significantly affects wool crimp number and staple length in Subo Merino sheep.
SNP2 in KIF27 significantly influences live weight after shearing in the same breed.
Mutations at SNP1 and SNP2 alter protein structures, impacting α-helix and other structural elements.
Abstract
Background/Objectives: The Subo Merino sheep is a high-quality fine-wool breed developed through progressive hybridization, characterized by high wool yield and excellent wool quality. This study is designed to investigate the effects of two gene polymorphisms in Subo Merino sheep on wool traits, thereby providing critical theoretical and technical support for the breeding of high-quality fine-wool sheep. Methods: In this study, 944 one-year-old Subo Merino sheep were genotyped for coding regions of the BLTP1 and KIF27 genes using the Fluidigm BioMark™ HD system. Association between SNP loci and wool traits was analyzed via the least squares means method in SAS 9.4. Protein–protein interaction networks were constructed using the STRING database, and protein structures before and after mutation were predicted with SOPMA and SWISS-MODEL. Results: The results revealed that BLTP1 gene…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
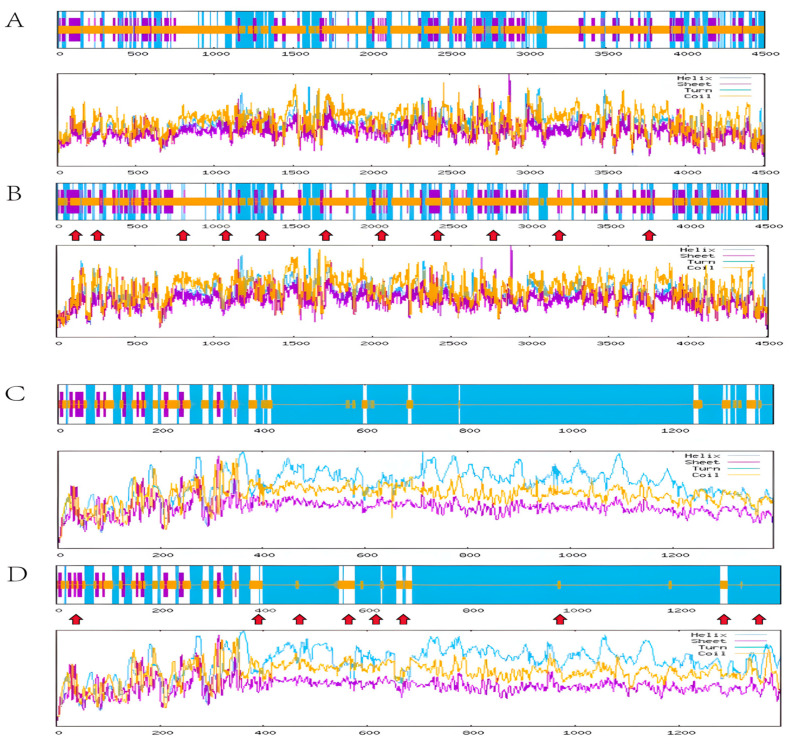 Figure 1
Figure 1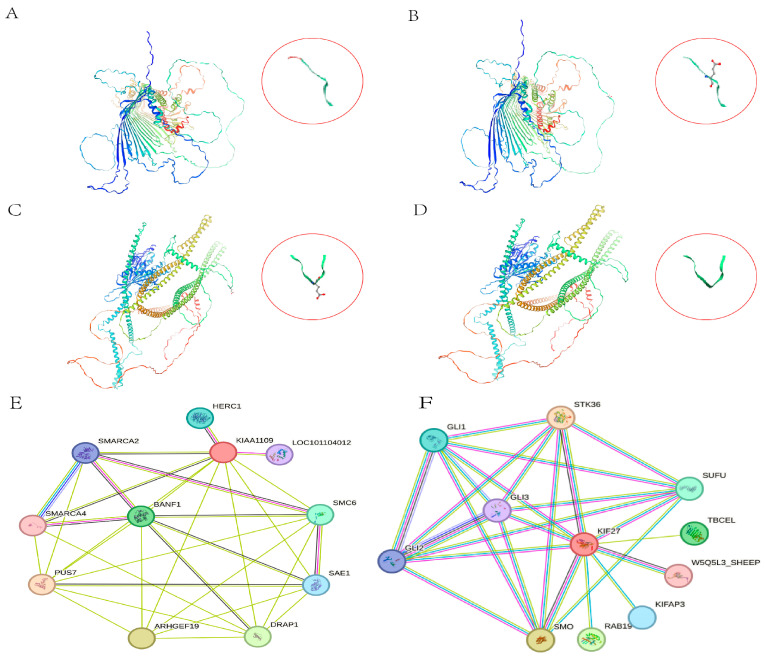 Figure 2
Figure 2- —earmarked fund for CARS
- —Natural Science Foundation of Xinjiang Uygur Autonomous Region
- —Xinjiang Uygur Autonomous Region “Tianshan Talents” training program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and phenotypic traits in livestock · Hair Growth and Disorders · Regulation of Appetite and Obesity
1. Introduction
China is one of the largest countries in sheep farming, but there remains a significant gap in the domestic processing demand for fine-wool [1]. The Chinese wool textile industry relies on imports for nearly half of its wool supply, with a particularly heavy dependence on imports for ultra-fine wool fibers measuring less than 19.5 microns in diameter. This underscores Chinese substantial market demand for high-quality wool, especially ultra-fine varieties [2].
Subo Merino Sheep are an ultra-fine-wool sheep breed developed in China in 2014 through strict phenotypic selection and genetic evaluation, after three breeding stages [3,4]. The wool diameter of Subo Merino Sheep is precisely controlled between 17.0~19.0 μm, with stable genetic performance, ensuring the continued transmission of excellent traits and reducing the gap between domestic and imported wool [5]. Exploring molecular markers related to wool traits in Subo Merino Sheep provides theoretical support for the utilization of this breed’s excellent genetic resources [6]. Wool traits are quantitative traits controlled by multiple minor-effect genes. Identifying genetic markers for these traits provides theoretical and technical support for molecular breeding in fine-wool sheep, ultimately enhancing wool production efficiency and quality—thereby boosting the self-sufficiency and market competitiveness of Chinese wool industry and delivering significant economic value.
Based on the results of the previous genome-wide association study conducted by the research team, it was suggested that the BLTP1 and KIF27 genes are associated with wool traits [7]. Related studies have shown that BLTP1 is an important non-vesicular lipid transport protein in multiple species [8]. In eukaryotic cells, the bridge lipid transport protein BLTP1 can promote the exchange of various lipids between the membranes of adjacent organelles through membrane contact sites, playing a crucial role in lipid metabolism, membrane transport, and cell signal transduction processes [9,10]. This mechanism is essential for the effective turnover and distribution of various intracellular lipids, such as phospholipids [8,9,10]. Studies on transmembrane proteins reveal that proteins in the TMEM170 family, acting as endoplasmic reticulum lipid flippases, can work synergistically with BLTP1 to form molecular bridges that mediate bulk lipid transport, ensuring the efficient flow of lipids between organelles [11]. Membrane contact sites are important locations for intracellular lipid metabolism and signal transduction, and BLTP1 regulates the function of these sites, making it a key component in maintaining material exchange and signal integration between organelles [9]. In Caenorhabditis elegans, the protein LPD-3 (the human homolog of BLTP1) constructs a molecular bridge between the endoplasmic reticulum and the plasma membrane, mediating lipid transport, and is crucial for the organism’s cold resistance [12]. In summary, BLTP1 has been confirmed in multiple species to be involved in lipid transport and metabolism processes, and its function is highly conserved in evolution. This key characteristic supports the hypothesis that BLTP1 may be an important candidate gene affecting wool traits by regulating lipid metabolism pathways.
KIF27 belongs to the kinesin-4 subfamily of motor proteins and is a key molecule in regulating cell mitosis and ciliogenesis. Recent studies have shown that KIF27 plays a central role in the formation and maintenance of cilia. Cilia play important roles in the physiological processes of mammalian nervous, respiratory, and reproductive systems [13,14]. Furthermore, KIF27 affects microtubule dynamics, suggesting its conserved role in microtubule organization within the kinesin-4 family [15]. Members of the kinesin protein family are generally involved in the intracellular transport of various “cargoes” along microtubules, including vesicles, protein complexes, and organelles [16]. In KIF27 and KIF7, changes in the chemical–mechanical coupling lead to impaired motility, and this unique motility property may correspond to their specific regulatory roles in the Hedgehog signaling pathway and ciliogenesis [15,17]. Moreover, abnormal expression and dysfunction of kinesin family genes are associated with various diseases [16]. In a study on the polymorphism of the KIF-I gene in cashmere goats, an SNP in exon 1 was found to be significantly related to fleece density [18]. In summary, functional variations in the kinesin family member KIF27 may influence the growth and development of wool traits from multiple dimensions.
Early studies have shown a significant genetic correlation between wool length and fineness in Subo Merino Sheep (r_A_ = 0.388), meaning that selecting for finer wool often leads to a reduction in wool length and clean fleece weight. This phenomenon has a stronger effect on live weight and clean fleece weight after shearing (r_A_ = 0.616), where selecting for finer wool leads to a decrease in clean fleece weight, indirectly resulting in a reduction in live weight after shearing [4]. This antagonistic relationship between traits restricts further improvement of the breed. Therefore, analyzing the polymorphism distribution of the BLTP1 and KIF27 genes in the Subo Merino Sheep population and evaluating their association with wool fineness, length, crimp, and other phenotypes will not only help elucidate the molecular mechanisms of ultra-fine-wool formation but also provide a theoretical basis for designing high-precision molecular markers. This will be of significant importance for early selection and accelerating genetic progress in fine-wool sheep breeding.
2. Materials and Methods
2.1. Sample Collection
This study used 944 one-year-old female Subo Merino sheep, this variety is seasonally propagated. The animals were sourced from two locations: 473 from the Gongnaisi Breeding Farm in Xinjiang Yili Prefecture (n = 473) and 471 from the Baicheng Breeding Farm in Aksu Prefecture, Xinjiang (n = 471). At both farms, the sheep were managed under a similar dietary regimen involving a combination of grazing and supplementary feeding. In May, blood samples (5 mL) were collected from the jugular vein of each sheep into anticoagulant tubes. The collected blood was mixed and stored at −20 °C for subsequent genomic DNA extraction. Additionally, wool samples were collected from each sheep at a point 10 cm above the posterior edge of the left scapula along the midline. The following wool traits were directly measured: greasy fleece weight (GFW), live weight before shearing (LWBS), and live weight after shearing (LWAS).
The environmental conditions during the measurement were: temperature (20 ± 2 °C) and humidity (65% ± 4%). The following wool traits were measured using the Optical Fiber Diameter Analyzer OFDA2000 (OFDA-2000BT, BSC Electronics, Perth, Australia): mean fiber diameter (MFD), staple length (SL), hair length (HL), Fineness Count (FC), coefficient of variation in fiber diameter (CVFD), fiber diameter standard deviation (FDSD), and crimp and crimp number (CN).
2.2. SNP Genotyping and Statistical Analysis
This study utilized the Fluidigm BioMark™ HD system (BioMark™ HD, San Francisco, CA, USA) to perform SNP genotyping for the BLTP1 and KIF27 genes in 944 individual sheep. The SNP genotype frequencies, allele frequencies, observed heterozygosity (Ho), effective allele number (Ne), expected heterozygosity (He), polymorphism information content (PIC), and Hardy–Weinberg equilibrium (HWpval) were calculated using Popgene 1.32 software.
The correlation between different SNP genotypes and wool traits was analyzed using SAS 9.4 software. The results are presented as least squares means ± standard errors. The linear model used was:
In the formula, Yick denotes the phenotypic value of an individual fine-wool sheep; μ denotes the population mean; Gi denotes the genotype SNP effect; Fc denotes the farm effect; and eick denotes random error.
2.3. Protein Structure Prediction and Network Interaction Analysis
Based on the sequence information of the target genes BLTP1 (ENSOART00000000336.1) and KIF27 (ENSOART00000009775.1) from the Ensembl database, the gene sequences were translated into amino acid sequences using the NOVOPRO website. The secondary protein structure was predicted using SOPMA “https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html” (accessed on 12 May 2025) and the tertiary protein structure was predicted using SWISS-MODEL “https://swissmodel.expasy.org/” (accessed on 14 May 2025). Based on protein interaction data from the STRING database, interaction networks were constructed with BLTP1 and KIF27 as the core nodes.
3. Results
3.1. Descriptive Statistics of Wool Traits
Descriptive statistical analysis of 11 wool traits (MFD, FDSD, CVFD, SL, FC, Crimp, SL, CN, GFW, LWBS, and LWAS) in Subo Merino Sheep was performed using SAS 9.4 software, and outliers were removed (Table 1). The data are presented as mean ± standard deviation. The analysis results show that the trait data align with objective facts, with the small standard deviation indicating that the variable distribution is relatively concentrated around the mean.
3.2. SNP Mutation Information Statistics
SNP genotyping of the two mutation sites, SNP1 in the BLTP1 gene and SNP2 in the KIF27 gene, was successfully performed on 944 Subo Merino Sheep using Fluidigm technology. After aligning with the reference gene sequence, SNP1 was identified as a missense mutation in exon 7 of the BLTP1 gene, resulting in the nucleotide change c.812 (C > T) and causing the amino acid change p.Pro271Leu. SNP2 was identified as a missense mutation in exon 18 of the KIF27 gene, resulting in the nucleotide change c.3896 (T > C) and causing the amino acid change p.Met1299Thr (Table 2).
3.3. SNP Genetic Polymorphism Analysis
This genotype frequencies, allele frequencies, observed heterozygosity (Ho), expected heterozygosity (He), and polymorphism information content (PIC) of the BLTP1 SNP1 and KIF27 SNP2 loci in Subo Merino Sheep are shown in Table 3. For BLTP1 SNP1, the genotypes AA, GA, and GG were identified, with GA (0.471) being the dominant genotype. For KIF27 SNP2, the genotypes TT, TC, and CC were identified, with TC (0.469) being the dominant genotype. The PIC for SNP1 was 0.354 and for SNP2 was 0.374, both of which are considered moderate polymorphism. The He for both SNP1 and SNP2 was close to 0.5, indicating that the genetic diversity at these two loci is at a moderately high level, with SNP2 showing slightly higher diversity. Both SNP loci also conform to Hardy–Weinberg equilibrium (p > 0.05), indicating that the studied population is in random mating at these loci.
3.4. Correlation Analysis Between SNPs and Wool Traits
The correlation between the SNP1 locus in the BLTP1 gene and the SNP2 locus in the KIF27 gene with wool traits in Subo Merino Sheep was analyzed using SAS 9.4 software. The results showed that the BLTP1 gene SNP1 locus was significantly associated with CN (p < 0.05) and SL (p < 0.05), but not with other phenotypes such as FC, MFD, FDSD, and MSL (p > 0.05). The KIF27 gene SNP2 locus showed a significant association with LWAS (p < 0.05), but no significant association with other traits (p > 0.05) (Table 4).
3.5. Protein Structure Prediction Analysis
The secondary protein structure before and after mutation was predicted using the SOPMA website, while the tertiary structure was predicted using the SWISS-MODEL website. A comparison of the protein secondary structure of the C and T base sequences for BLTP1 gene SNP1 is shown in Table 5. The change in the base at the SNP1 locus led to an increase in the α-helix content from 23.13% before mutation to 23.73% after mutation, an increase in the extended chain from 10.91% to 11.38%, while the random coil decreased from 65.96% to 64.89%. For KIF27 gene SNP2, a comparison of the T and C base sequences of the protein secondary structure is shown in Table 5. After the mutation at SNP2, the α-helix content increased from 75.63% to 75.84%, while the extended chain decreased from 4.95% to 4.87%, and the random coil decreased from 19.43% to 19.28% after the mutation.
The missense mutations in genes cause changes in amino acid residues, so the protein secondary structure before and after mutation was predicted using the SOPMA v.2 website. A comparison of the protein secondary structure of the C and T base sequences at the BLTP1 gene SNP1 locus is shown in Figure 1. The SNP1 locus causes the amino acid change p.Pro271Leu, and the secondary structure changes in 11 main regions, with overall significant changes that are not concentrated in one area. The changes occur between amino acids 0–4000 (Figure 1B). For KIF27 gene SNP2, a comparison of the T and C base sequences of the protein secondary structure is shown in Figure 1. The SNP2 locus causes the amino acid change p.Met1299Thr, with changes occurring in 9 main regions. These changes are concentrated in the 0–1400 amino acid range (Figure 1D).
Through observation of the tertiary structures of BLTP1 (Figure 2A,B) and KIF27 (Figure 2C,D), it can be seen that the mutations at SNP1 and SNP2 have a minimal impact on the overall shape of the protein’s tertiary structure, with no significant structural changes. However, localized changes were observed at specific nodes. Protein interaction network analysis revealed that several key potential genes, such as SMARCA2, SMARCA4, and BANF1, play crucial roles in the BLTP1 network topology (Figure 2E). Additionally, KIF27 interacts with potential genes including STK36, GLI1, GLI2, GLI3, SUFU, and SMO (Figure 2F).
4. Discussion
Currently, numerous scholars are conducting extensive research on molecular markers for wool traits using methods such as candidate gene approaches, QTL mapping, and genome-wide association studies. These studies have accumulated valuable experimental data and theoretical insights for the molecular breeding of fine-wool sheep [19,20,21]. Wool traits (such as wool diameter, wool yield, and wool length) are quantitative traits, controlled by multiple genes with small effects. Therefore, the discovery of genetic markers for these traits requires further in-depth investigation. In this study, based on our previous research, we selected the BLTP1 and KIF27 genes and performed polymorphism identification in the Subo Merino Sheep population. The results successfully genotyped SNP1 (c.812C > T, p.Pro271Leu) in BLTP1 and SNP2 (c.3896T > C, p.Met1299Thr) in KIF27, and both SNP loci were in Hardy–Weinberg equilibrium (p > 0.05). Phenotypic association analysis indicated that SNP1 and SNP2 were significantly associated with key wool traits. Although this study did not systematically explore the entire genome, our findings still provide valuable clues for the functional study of these genes. More importantly, the results fill the research gap of BLTP1 and KIF27 genes in Subo Merino sheep, a unique, high-quality fine-wool breed, and provide a reference for the molecular marker-assisted breeding technology of fine-wool sheep.
This study found a significant association between the SNP1 mutation in the BLTP1 gene and SL as well as CN in Subo Merino Sheep (p < 0.05). Previous studies have shown that in Chinese Merino sheep, live weight after shearing is moderately positively genetically correlated with staple length, and CN is negatively genetically correlated with MFD [22,23,24]. However, in our study, no similar trends were observed between the genotypes of SNP1. Specifically, the GA genotype showed higher live weight before shearing compared to the GG and AA genotypes. Although this difference did not reach statistical significance, it provides a reference for multi-trait synergistic improvement. We are also focused on identifying molecular genetic markers that can simultaneously improve both wool traits and growth performance in sheep, to provide theoretical support for the breeding of dual-purpose sheep (both wool and meat).
BLTP1, a member of the bridge lipid transfer protein family, primarily functions in lipid transport, maintaining the dynamic balance of organelle membranes, and is implicated in the pathogenesis of certain diseases [8]. In mouse models, the deletion of BLTP1 affects the survival and normal development of the neuromuscular junction, indicating the gene’s importance in the development and functional maintenance of the nervous system [10]. Case studies have shown that variations in the splicing sites of the BLTP1 gene can lead to multiple congenital malformations in fetuses, highlighting BLTP1’s indispensable role in mammalian development. Dysfunction in BLTP1 could result in severe congenital defects [25].
Currently, there is limited research on the role of the BLTP1 gene in hair follicle growth and development, but lipid transport plays a crucial role in maintaining the homeostasis of hair follicle cells [26,27]. Studies in mice have shown that immature dermal adipocytes and mature adipocytes play different roles in hair follicle development and hair growth [28,29]. In our previous research using transcriptomics and proteomics, we found that lipid-related genes are associated with the cyclical nature of secondary hair follicles in the Jiangnan cashmere goat. Cell experiments revealed that FA2H promotes the proliferation of secondary hair follicle dermal papilla cells, with FA2H encoding a fatty acid 2-hydroxylase [30]. PLIN2, a gene that regulates lipid metabolism, when overexpressed, can modulate the proliferation and apoptosis of DPCs, affecting the hair follicle growth cycle and cashmere fiber production [31]. Moreover, protein interaction network analysis showed that BLTP1 directly interacts with SMARCA2, SMARCA4, and BANF1. Notably, SMARCA4 and SMARCA2 are key pathogenic genes in Coffin–Siris syndrome, while pathogenic variants in both alleles of BANF1 cause autosomal recessive Néstor–Guillermo progeroid syndrome, both of which lead to sparse hair growth [32,33]. Combining these findings with our results, a novel insight is proposed: BLTP1 may regulate the proliferation and apoptosis of hair follicle dermal papilla cells by participating in lipid transport, and further interact with SMARCA2, SMARCA4, and BANF1 to affect hair follicle development, thereby regulating wool staple length and crimp number. Although the mechanism by which the SNP1 mutation in BLTP1 affects staple length and crimp number remains unclear, the above results suggest that BLTP1 may be involved in the regulation of hair follicle development and hair growth. The specific regulatory mechanisms will be further explored in our future research.
In this study, we found that the SNP2 mutation in the KIF27 gene was significantly associated with LWAS in Subo Merino Sheep (p < 0.05). KIF27 belongs to the kinesin-4 superfamily, and its members play key roles in intracellular material transport, microtubule dynamics, and the maintenance of ciliogenesis functions [15,16,17]. Notably, in the protein interaction network, KIF27 interacts with genes annotated in the Sonic Hedgehog (Shh) signaling pathway, including STK36, GLI1, GLI2, GLI3, SUFU, and SMO. This suggests that KIF27 may play an important role in the Hedgehog (Hh) signaling pathway in mammals. The Hh pathway, particularly Sonic Hedgehog is a core signaling pathway that regulates hair follicle development. Research on Shh has shown that when the SMO gene is specifically deleted in cells, Shh no longer affects hair follicles, adipocytes, and endothelial cells. Instead, Shh directly acts on adipocyte precursors to induce dermal fat generation [34]. Early studies have also confirmed that Shh is essential for the morphogenesis of hair follicles in mouse embryos [35], and continues to play a role in the cyclical regeneration of hair follicles [36]. The regulation of Gli2 by SUFU and SPOP is crucial for the differentiation of cochlear hair cells in mammals, and SUFU is a negative regulator of Shh signaling, controlling the timing and progression of hair cell differentiation [37]. In hair follicles, the Hh signaling pathway is transmitted through primary cilia and primarily activates downstream Gli2 transcription factors to perform its functions; however, Gli3 also has a compensatory activation pathway that does not depend on cilia [38]. Additionally, hair follicle development requires the coordinated regulation of several signaling pathways, including Wnt signaling pathways (necessary for structural formation), Notch signaling pathways (regulating cell differentiation), and Bmp signaling pathways (a negative regulator) [39]. Therefore, as a potential component of the Hh pathway, KIF27 may influence this pathway and contribute to the fine-tuned regulation of hair follicle development. In studies on goats, circKIF27 was found to be widely expressed in the basal layer melanocytes of the skin, with differential expression in melanocytes of white and brown skin. circKIF27 can inhibit cell proliferation, reduce tyrosinase activity, and decrease melanogenesis by targeting the miR-129-5p/TGIF-2 pathway, thus participating in goat skin color formation [40]. In our study, the SNP2 mutation in KIF27 was significantly associated with live weight after shearing (LWAS) in Subo Merino Sheep (p < 0.05), but there was no significant effect on wool traits (p > 0.05). Although the specific mechanism by which the SNP2 mutation in KIF27 affects live weight after shearing remains unclear, these results suggest that KIF27 may participate in the regulation of hair follicle development and hair growth through the Shh signaling pathway. Further research is needed to better understand the precise regulatory role of KIF27 in hair follicle development and wool growth.
From a practical perspective, the results of this study enrich the genetic marker resources of Subo Merino sheep, help protect and utilize the excellent genetic resources of local fine-wool sheep, and promote the sustainable development of the fine-wool sheep industry. It should be noted that this study still has certain limitations. Only one SNP locus was identified in each of the BLTP1 and KIF27 genes, and the regulatory mechanisms of these SNPs on wool traits and growth performance have not been verified by cell and animal experiments. In future research, we will further expand the sample size, screen more polymorphic loci of the BLTP1 and KIF27 genes, and verify the functional roles of target SNPs through in vitro cell experiments and in vivo animal experiments to clarify their specific regulatory mechanisms.
5. Conclusions
In this study, we identified two missense mutations: SNP1 (c.812C > T, p.Pro271Leu) in the BLTP1 gene and SNP2 (c.3896T > C, p.Met1299Thr) in the KIF27 gene. Both mutation loci were consistent with the Hardy–Weinberg equilibrium law in the Subo Merino sheep population. Association analysis confirmed that SNP1 significantly affected SL and CN (p < 0.05), while SNP2 was significantly correlated with LWAS (p < 0.05). Combined with protein structure prediction and protein–protein interaction network analysis, it is speculated that BLTP1 may regulate hair follicle development through lipid transport and its interaction with SMARCA2, SMARCA4, and BANF1, while KIF27 may participate in the hair follicle regulation process through the Sonic Hedgehog signaling pathway. In summary, SNP1 and SNP2 can be used as potential molecular markers for wool traits in Subo Merino sheep, providing important references for fine-wool sheep breeding and helping to shorten the breeding cycle, reduce the cost of phenotypic selection, and further improve the efficiency and accuracy of breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zheng W.X. Xiao H.F. Zhang M. Development status, future trends and suggestions of China’s wool and cashmere sheep industry Chin. J. Anim. Sci.202359300315300–306+315(In Chinese)10.19556/j.0258-7033.20230306-04 · doi ↗
- 2Tian K.C. Current situation, development trend and countermeasures of wool sheep industry in China Feed. Anim. Husb.20191102635(In Chinese)
- 3Guan M.X. Wei C. Si B.X. Zhang X. Mao J. Wu W. Hanikizi T. Zhang Y. Tian K. Huang X. Estimation of genetic parameters for major economic traits in Subo Merino Sheep Chin. J. Anim. Sci.20225897101(In Chinese)
- 4Liu N. Tian K.C. Shi G.Q. Di J. Yang Y. He J. Liu J. Effects of different generations of grading hybridization in the core flock on wool production traits of Subo Merino Sheep Chin. J. Anim. Sci.201551610(In Chinese)
- 5Zeng W.D. Analysis of DNA Methylation Patterns in Skin Tissues at Different Stages of Hair Follicle Development in Subo Merino Sheep Doctoral Dissertation Xinjiang Agricultural University Xinjiang, China 2020(In Chinese)10.27431/d.cnki.gxnyu.2020.000166 · doi ↗
- 6Song W.J. Gu Y. Lu Q.W. Liu W. De D. Zheng J. Sideng D. Fu X. He J. Polymorphisms of KAP 15-1 and KAP 27-1 genes and their association with wool traits in Ordos Fine Wool Sheep Chin. J. Anim. Sci.202460203210(In Chinese)10.19556/j.0258-7033.20221015-03 · doi ↗
- 7Zhao B. Luo H. He J. Huang X. Chen S. Fu X. Zeng W. Tian Y. Liu S. Li C. Comprehensive transcriptome and methylome analysis delineates the biological basis of hair follicle development and wool-related traits in Merino sheep BMC Biol.20211919710.1186/s 12915-021-01127-934503498 PMC 8427949 · doi ↗ · pubmed ↗
- 8Liu Y. Ye Q. Ma D.K. Rothermel B.A. Lin W. Bltp 1 Is Required for Survival and Normal Development and Function of the Neuromuscular Junction in Mice J. Neurosci.202545 e 002925202510.1523/JNEUROSCI.0029-25.202540973493 PMC 12572931 · doi ↗ · pubmed ↗