MeNADP-ME3 Confers Salt and Drought Tolerance in Arabidopsis and Drives Functional Diversification of the NADP-ME Family in Cassava
Shuwen Wu, Zhanming Xia, Jiazheng Zhao, Changyi Wang, Yi Min, Dayong Wang

TL;DR
This study explores how a specific NADP-ME gene in cassava helps the plant tolerate drought and salt stress, offering insights into improving crop resilience.
Contribution
The study identifies MeNADP-ME3 as a novel gene conferring stress tolerance and driving functional diversification in cassava.
Findings
Arabidopsis overexpressing MeNADP-ME3 showed 30–120% higher antioxidant enzyme activities under stress.
MeNADP-ME3 is localized in chloroplasts and responds to abiotic stresses like drought and salt.
Cassava's NADP-ME family shows distinct expression patterns and evolutionary diversification.
Abstract
As a typical C3-C4 intermediate plant, cassava (Manihot esculenta Crantz) exhibits high photosynthetic efficiency and low photorespiration. NADP-malic enzyme (NADP-ME) is a key enzyme in the C4 photosynthetic pathway that provides elevated CO2 concentrations for Rubisco. However, research on NADP-ME in C3-C4 intermediate species remains limited. In this study, we identified four NADP-ME genes in the cassava genome, with segmental duplication serving as the primary driving force for gene evolution. Cis-acting element analysis indicated potential roles of MeNADP-ME genes in environmental adaptation, stress responses, and growth regulation. Expression profiling using bulk RNA sequencing and single-cell RNA sequencing revealed distinct expression patterns in different tissues and cell subsets. Comparative analysis with Arabidopsis (Arabidopsis thaliana) and maize (Zea mays) NADP-ME families…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
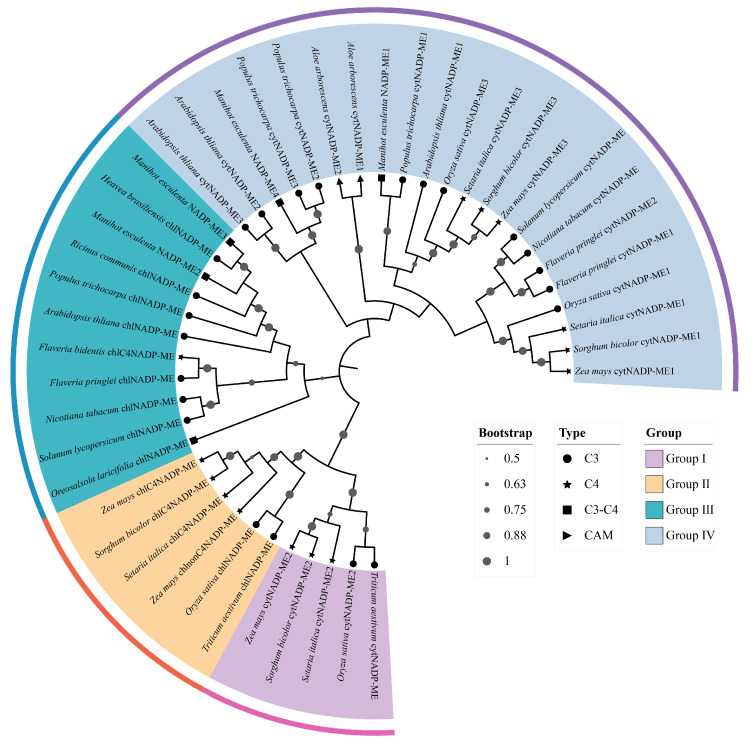 Figure 1
Figure 1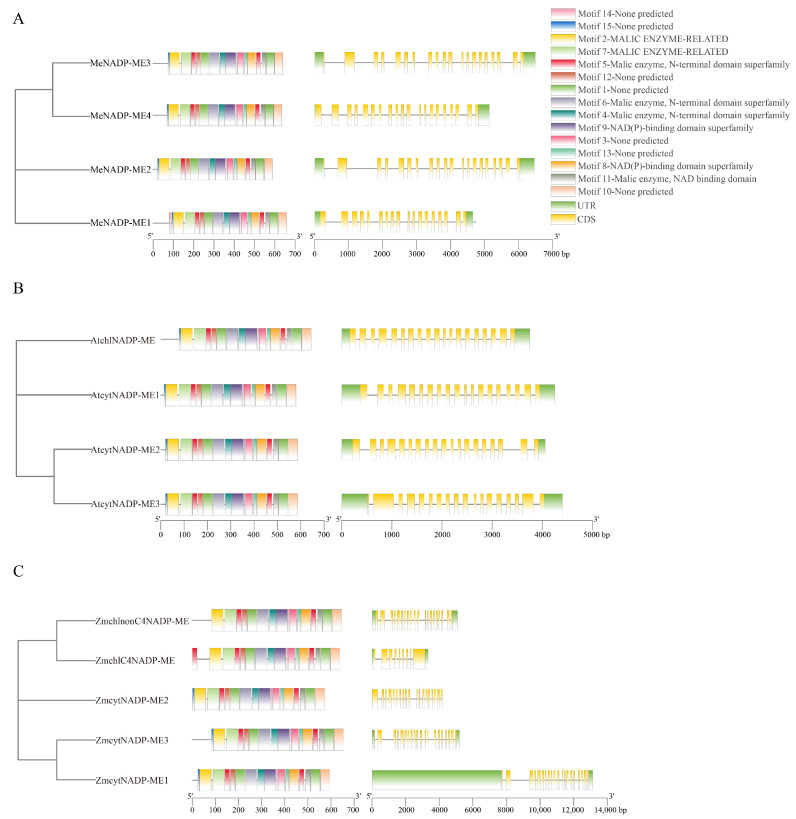 Figure 2
Figure 2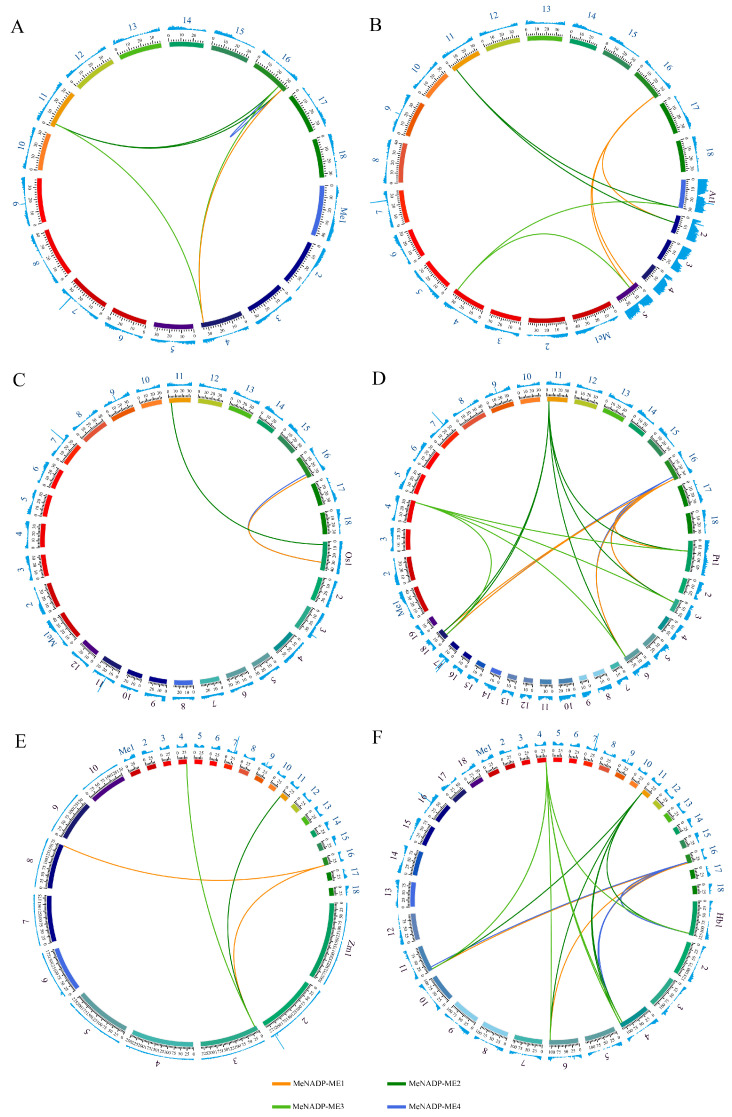 Figure 3
Figure 3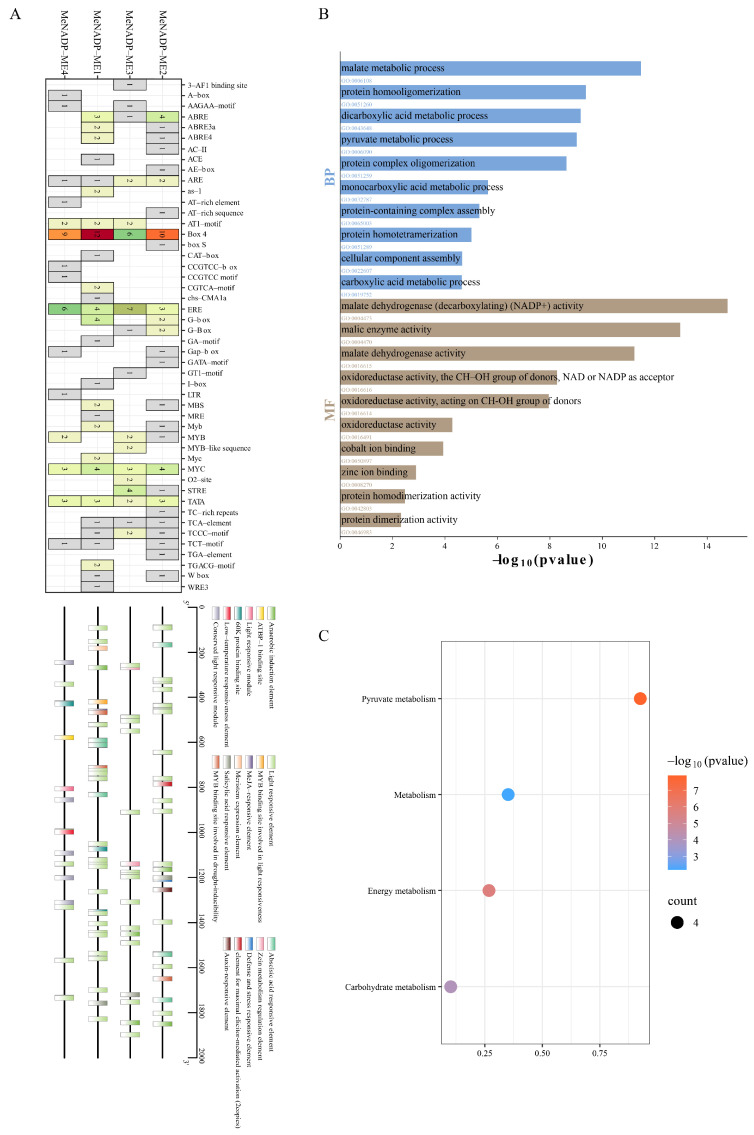 Figure 4
Figure 4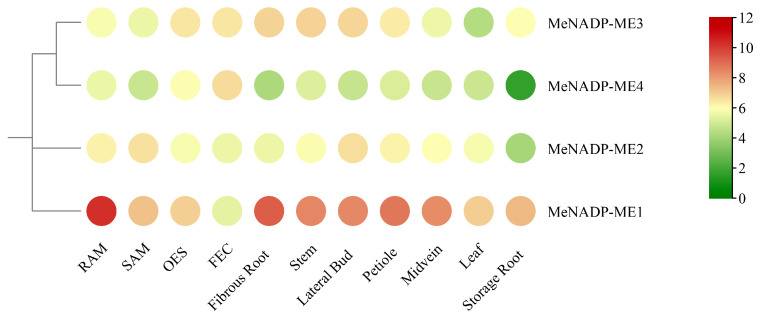 Figure 5
Figure 5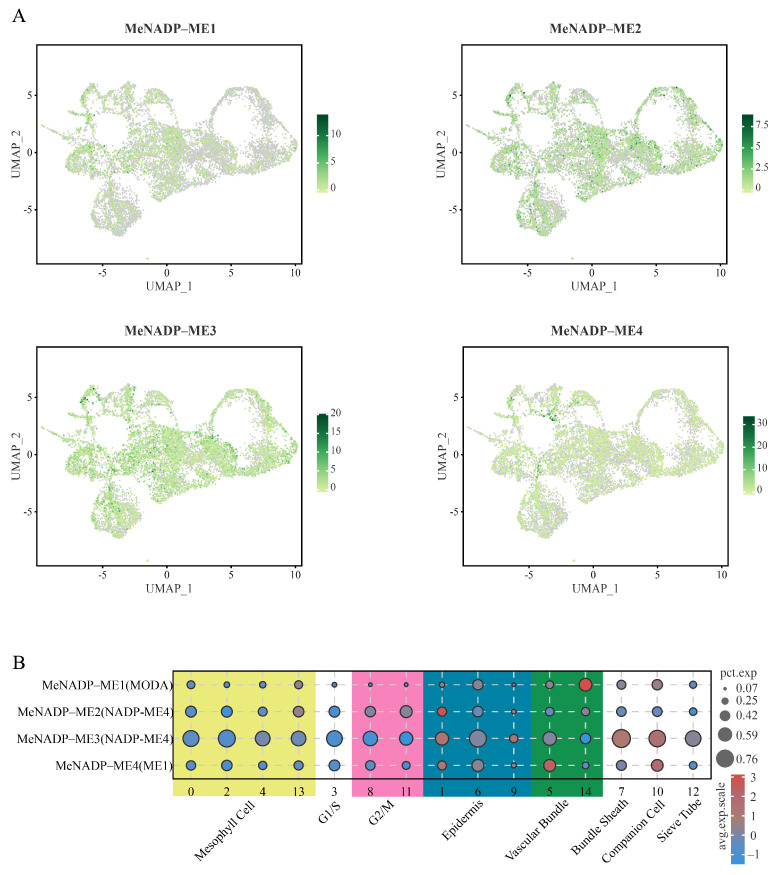 Figure 6
Figure 6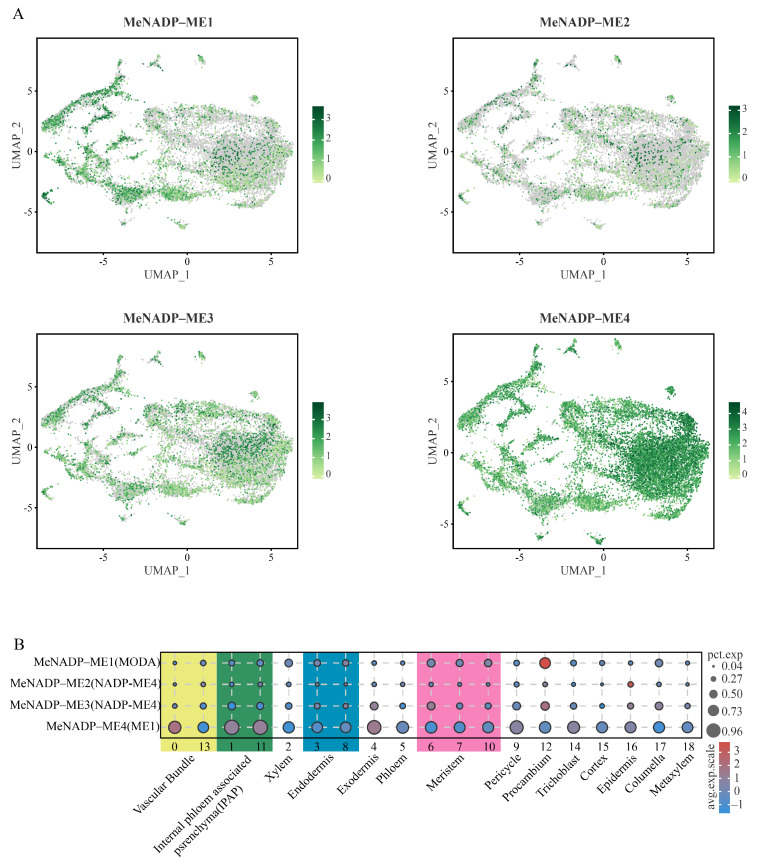 Figure 7
Figure 7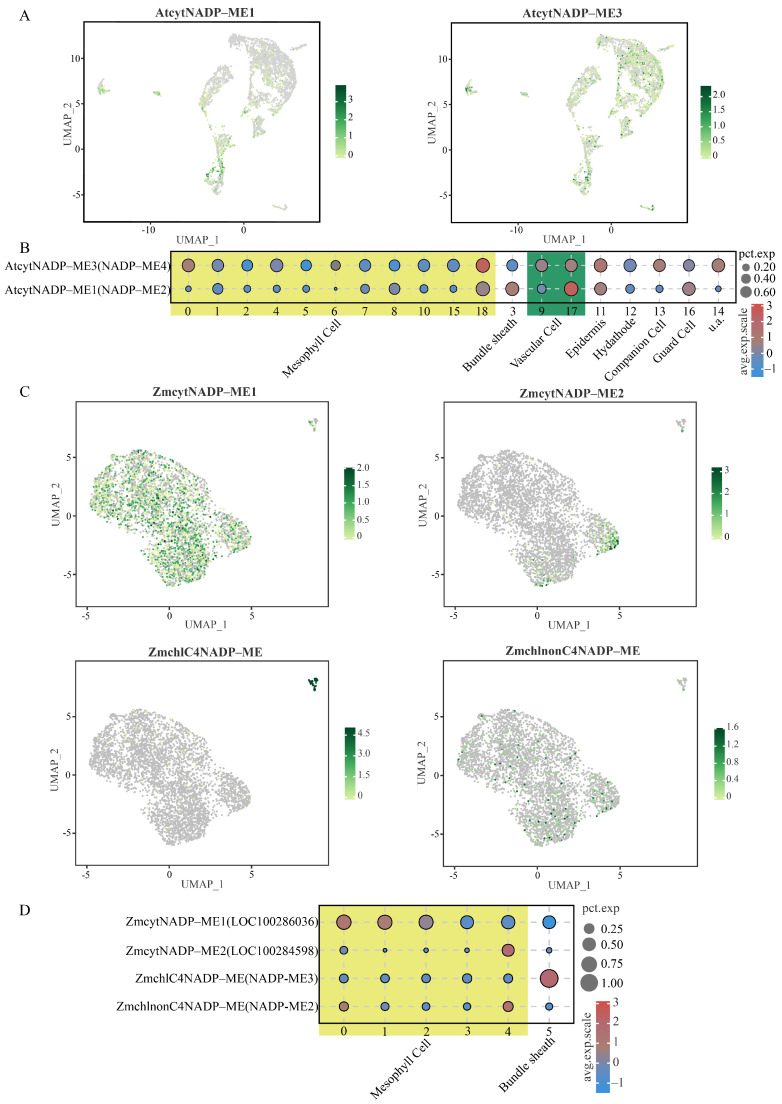 Figure 8
Figure 8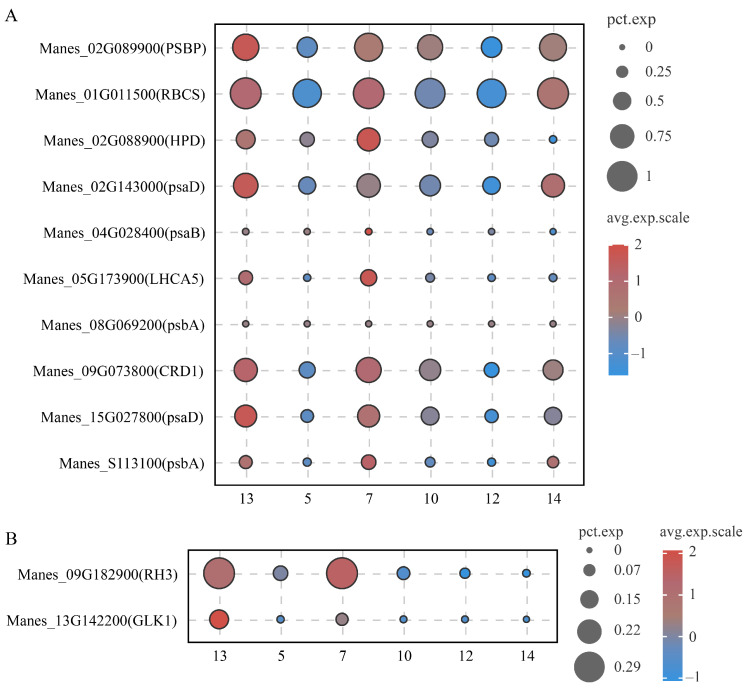 Figure 9
Figure 9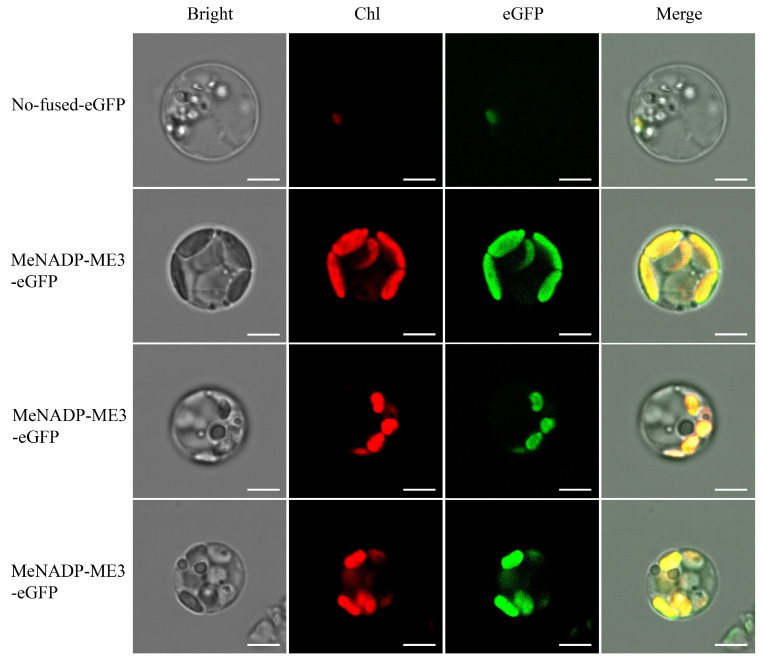 Figure 10
Figure 10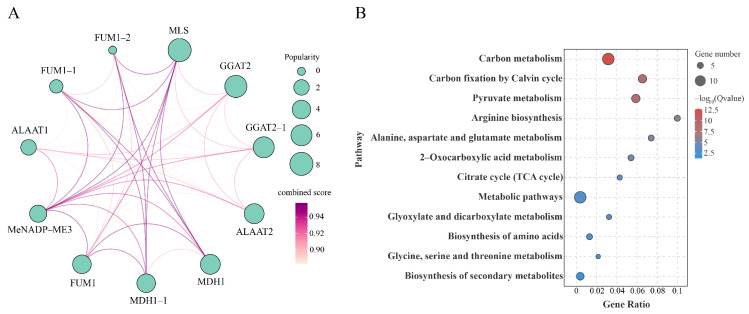 Figure 11
Figure 11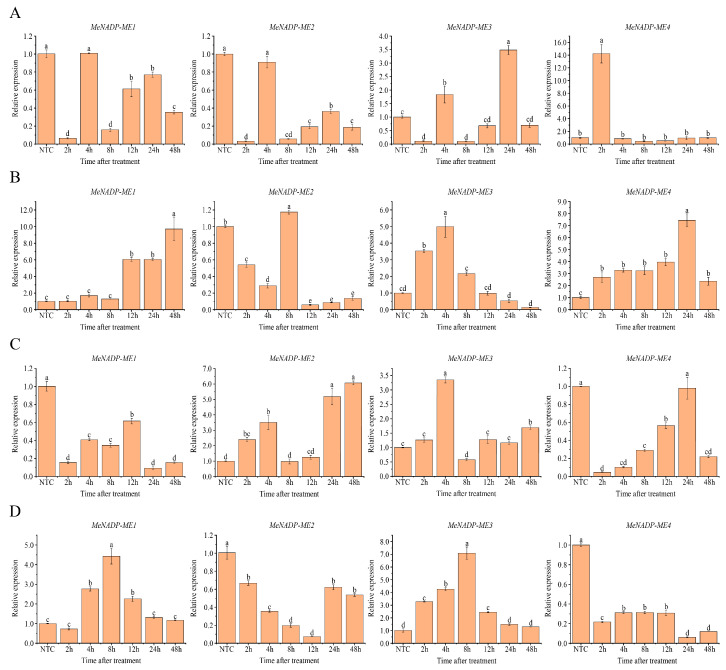 Figure 12
Figure 12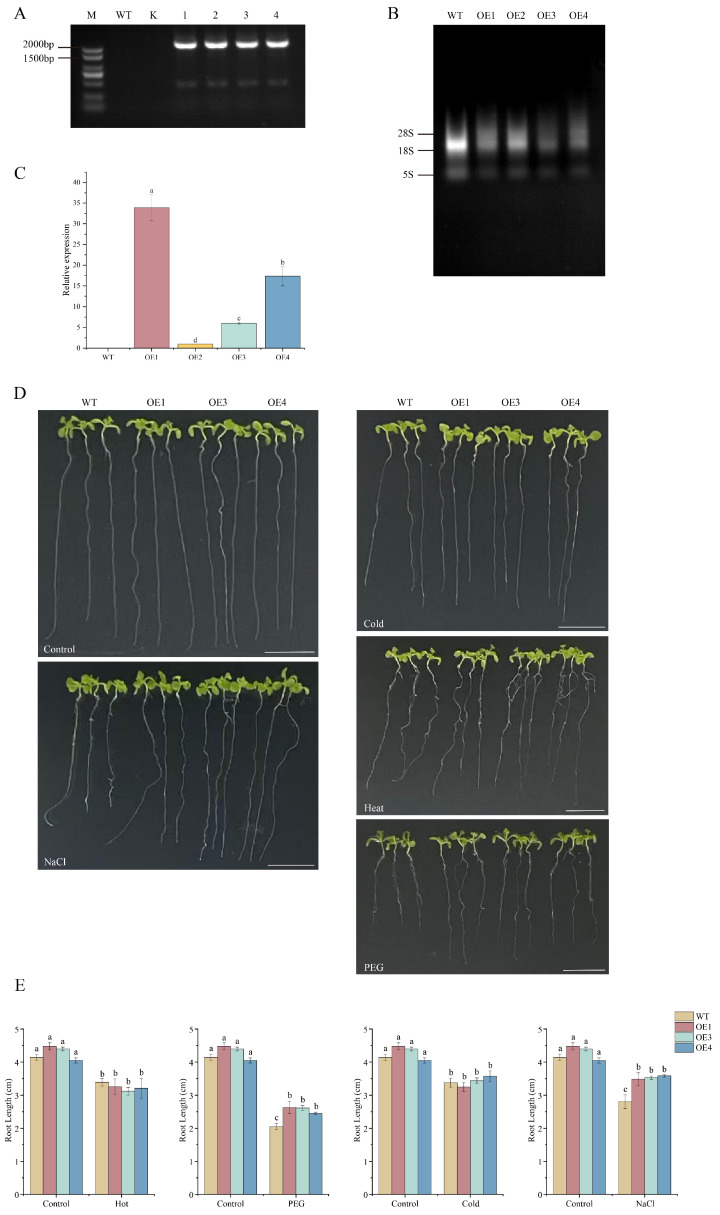 Figure 13
Figure 13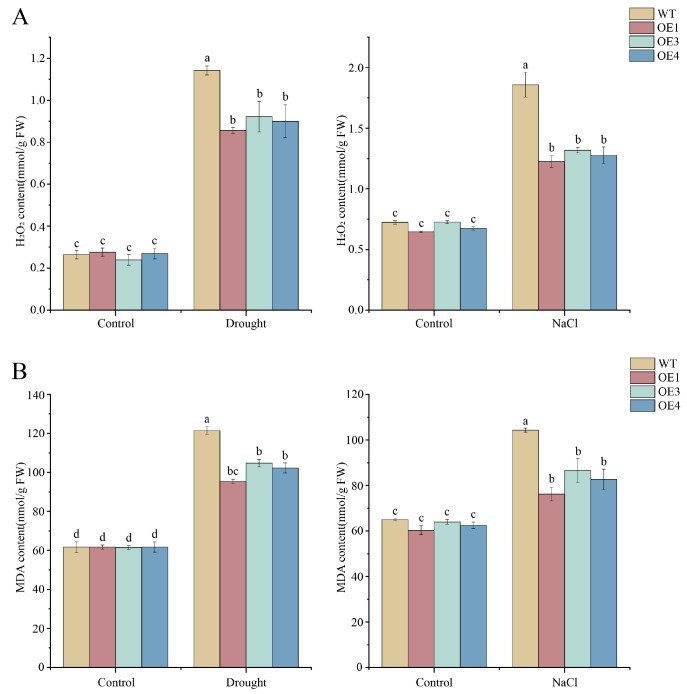 Figure 14
Figure 14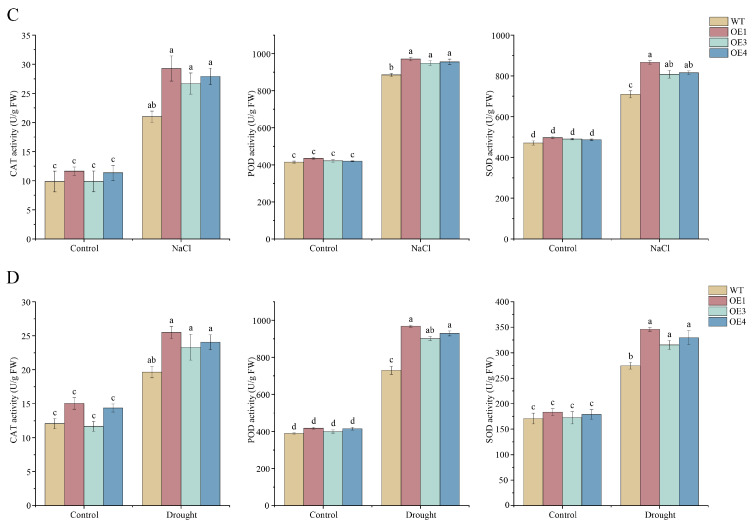 Figure 15
Figure 15Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCassava research and cyanide · Plant Micronutrient Interactions and Effects · Photosynthetic Processes and Mechanisms
1. Introduction
Cassava (Manihot esculenta Crantz) is recognized as a vital crop for addressing food security challenges and bioenergy development [1,2,3]. Compared to C3 and C4 plants, cassava features high photosynthetic efficiency and low photorespiration. Despite lacking Kranz anatomy, its phosphoenolpyruvate carboxylase (PEPC) activity exceeds that of C3 plants [4,5], supporting its classification as a C3-C4 intermediate species [6]. In C4 plants, PEPC initially fixes into C4 acids within mesophyll cells, which subsequently diffuse via plasmodesmata (PD) to bundle sheath cells for decarboxylation [7]. Subsequently, NADP-malic enzyme (NADP-ME; EC 1.1.1.40) catalyzes malate decarboxylation in bundle sheath cells, producing pyruvate, , and NADPH, thereby increasing the concentration available for ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) [8]. This unique concentrating mechanism (CCM) in C4 plants effectively suppresses photorespiration, enhances photosynthetic efficiency, and optimizes water and nitrogen resource utilization.
The NADP-ME gene family exhibits limited membership across plant species, functionally diverging into photosynthetic and non-photosynthetic isoforms [9]. Photosynthetic NADP-ME operates in bundle sheath chloroplasts of C4 plants and in the cytosol of CAM plants, supplying for carbon fixation [8]. Conversely, non-photosynthetic NADP-ME functions in plastids or the cytosol, participating in diverse physiological processes such as cytosolic pH homeostasis, stomatal regulation, providing NADPH for biosynthesis (e.g., of flavonoids and lignin) and antioxidant defense (via the ascorbate–glutathione cycle), alongside plant stress responses [10,11,12,13]. Structurally, plant NADP-ME enzymes universally feature two conserved domains (N-terminal domain and NAD(P)-binding domain) and typically assemble into tetrameric complexes [14,15].
Beyond its role in C4 photosynthetic carbon fixation, NADP-ME has emerged as a critical regulator of plant abiotic stress responses [16]. Studies in multiple plant species have established a conserved regulatory pattern of NADP-ME under osmotic stress. Under drought and salinity stress, elevated NADP-ME activity accompanied by increased chloroplastic NADP-ME transcripts and decreased cytosolic NADP-ME expression was observed in tobacco (Nicotiana tabacum) following PEG treatment or drought exposure [17]. Conversely, Potato virus Y (PVY) infection selectively enhanced cytosolic NADP-ME transcription in tobacco, suggesting distinct regulatory mechanisms for biotic versus abiotic stress responses [18]. This subcellular-specific regulation implies functionally distinct roles of NADP-ME isoforms in stress adaptation. Functional validation studies have demonstrated the causal role of plastidic NADP-ME in drought tolerance: constitutive expression of maize (Zea mays) ZmNADP-ME in tobacco chloroplasts conferred drought resistance by reducing stomatal conductance and improving water use efficiency [19]. Similarly, salt stress induces NADP-ME activity in rice (Oryza sativa), and transgenic Arabidopsis (Arabidopsis thaliana) plants overexpressing OsNADP-ME2, OsNADP-ME3, or OsNADP-ME4 exhibited enhanced osmotic regulation and improved salt tolerance [20,21,22]. Notably, overexpression of OsNADP-ME4 conferred tolerance to both drought and salt stresses [20], highlighting the dual function of specific NADP-ME isoforms. Heterologous expression experiments have further confirmed the conserved stress tolerance function across diverse plant species: overexpression of sweet sorghum (Sorghum bicolor) SbNADP-ME in Arabidopsis elevated salt tolerance by enhancing osmotic regulation, 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging capacity, and reducing membrane peroxidation [23], while SaNADP-ME4 overexpression from the halophyte Salsola laricifolia simultaneously enhanced drought and salt stress tolerance in Arabidopsis [24]. Beyond osmotic stress, NADP-ME also participates in heavy metal detoxification, as demonstrated by the observation that overexpression of Stylosanthes guianensis SgME1 increased aluminum tolerance in Arabidopsis by promoting malate synthesis and efflux [25].
In addition to osmotic and ionic stresses, NADP-ME is also involved in plant responses to temperature extremes, oxidative stress, and pathogen infection, primarily through modulating NADPH-dependent redox homeostasis. Low temperatures upregulated NADP-ME transcription in maize and sweet potato (Dioscorea esculenta) [26,27], suggesting its role in cold acclimation. UV radiation enhanced NADP-ME activity in Egeria densa, concomitant with increased activities of antioxidant enzymes including superoxide dismutase (SOD), peroxidase (POD), ascorbate peroxidase (APX), and catalase (CAT) [28], indicating a coordinated antioxidant defense response. The involvement of NADP-ME in plant immunity has also been established through genetic studies: Arabidopsis nadp-me2 mutants exhibited weakened apoplastic ROS bursts upon pathogen challenge and increased susceptibility to Colletotrichum higginsianum [29,30], while Magnaporthe oryzae subverts rice immunity by suppressing OsNADP-ME2 and OsNADP-ME3 activity, thereby reducing cytosolic NADPH and ROS production [31]. These diverse stress responses converge on a common mechanism: NADP-ME provides NADPH for ROS scavenging and biosynthetic processes essential for stress adaptation. Despite these advances in understanding NADP-ME functions in C3 and C4 model plants, plastidic NADP-ME from C3-C4 intermediate species such as cassava remains poorly characterized under abiotic stress. The metabolic and structural characteristics of cassava are distinct from those of C3 and C4 plants, and further research will enhance understanding of their functions and regulatory mechanisms in plant stress resistance, providing a theoretical basis for the breeding of stress-tolerant cassava varieties.
In this study, we conducted a comprehensive analysis of NADP-ME genes in cassava, encompassing their characteristic features, phylogenetic relationships, gene duplication, and expression profiles across various tissues and organs. Additionally, comparative expression profiling of NADP-ME genes was performed at single-cell resolution using single-cell RNA sequencing (scRNA-seq) in cassava, Arabidopsis, and maize, species that represent different photosynthetic types. The expression patterns of MeNADP-ME genes under diverse abiotic stresses were further analyzed via quantitative real-time PCR (qRT-PCR). Laser scanning confocal microscopy revealed subcellular localization of MeNADP-ME3 to chloroplasts within cassava leaf protoplasts. Concurrently, functional characterization of MeNADP-ME was conducted through overexpression in Arabidopsis, investigating its role under abiotic stress conditions. This study will provide a foundation for research on NADP-ME genes in C3-C4 intermediate species and facilitate the breeding of stress-resistant cassava cultivars.
2. Materials and Methods
2.1. Genome-Wide Identification and Characterization of NADP-ME Gene Family
Genomic sequence data and annotation files for cassava were retrieved from the NCBI Genome Database. Full-length protein sequences were extracted from the genomic dataset using the annotation files. The hidden Markov model (HMM) profiles corresponding to the NADP-ME family (accession IDs: PF00390, PF03949 [32]) were downloaded from the Pfam database (Pfam 38.0). Using the hmmsearch function in HMMER 3.0 (European Bioinformatics Institute, Cambridge, UK), these NADP-ME HMM profiles were employed to query the cassava full-length protein sequences, with proteins meeting an E-value threshold of < preliminarily retained. Concurrently, a BLASTp (https://blast.ncbi.nlm.nih.gov/Blast.cgi) search was performed against the cassava proteome using NADP-ME protein sequences from Arabidopsis as queries, selecting hits with E-value thresholds of < [33,34]. Final identification of cassava NADP-ME family members was achieved by integrating these results with conserved domain verification via the CDD database, retaining candidates exhibiting > sequence similarity to reference Arabidopsis NADP-ME protein sequences. Physicochemical properties of the identified cassava NADP-ME proteins were predicted using ExPASy (https://web.expasy.org/protparam/).
Positional information of MeNADP-ME was retrieved from genome annotation files, with gene structures analyzed based on exon-intron organization. Conserved motifs in NADP-ME proteins from cassava, Arabidopsis, and maize were identified using MEME and functionally annotated via the InterPro database [32]. The upstream 2000-bp regions of coding sequences (CDS) from each MeNADP-ME family member were extracted, and cis-regulatory elements were identified using the PlantCARE database [35]. Data were classified, deduplicated, and visualized via the ggplot2 (Springer-Verlag, New York, NY, USA, v3.5.1) package in R (R Foundation for Statistical Computing, Vienna, Austria, v4.2.2).
2.2. Phylogenetic and Evolutionary Analysis
NADP-ME protein sequences from Arabidopsis thaliana, Oryza sativa, Zea mays, Nicotiana tabacum, Flaveria pringlei, Populus trichocarpa, Sorghum bicolor, Aloe arborescens, Salsola laricifolia, Setaria italica, Triticum aestivum, Solanum lycopersicum, Ricinus communis, and Hevea brasiliensis were aligned with cassava NADP-ME protein sequences using MUSCLE in MEGA 7.0 (Institute for Genomics and Evolutionary Medicine, Temple University, Philadelphia, PA, USA). A phylogenetic tree was constructed via the Neighbor-Joining method (Bootstrap replicates: 1000; partial deletion) [36]. Groups were delineated based on phylogenetic clades and visualized using iTOL (https://itol.embl.de/).
Synteny gene pairs within cassava and across species were identified and visualized using the One-Step MCScanX-SuperFast module in TBtools (v2.331). Nonsynonymous (Ka) and synonymous (Ks) substitution ratios for MeNADP-ME were computed with the Simple Ka/Ks Calculator [37]. Integrated visualization of all results was performed with TBtools (The Xia Rui Research Group of South Agricultural University, Guangzhou, China, v2.331) [37].
2.3. Functional Annotation and Enrichment Analysis
For functional enrichment analysis, MeNADP-ME protein sequences were annotated through the eggNOG-mapper platform for Gene Ontology (GO) and KEGG pathway enrichment [38]. Results were collated with TBtools, then integrated and visualized using ggplot2 (v3.5.1). Protein–protein interaction network for MeNADP-ME3 was predicted using the STRING database. Generated interaction networks were visualized using the tidygraph package (Chinese Software Developer Network, Beijing, China, v1.3.1) in R (v4.2.2). MeNADP-ME3 and its interacting proteins were annotated for KEGG pathway enrichment through the eggNOG-mapper platform [38].
2.4. Transcriptome Data Analysis
RNA sequencing (RNA-seq) data from diverse cassava tissues were downloaded from the ENA database (Project: PRJNA324539) [39]. Following data preprocessing to generate raw count matrices, transcripts per million (TPM) values were calculated for MeNADP-ME genes. After log2 transformation, results were visualized using TBtools to generate gene expression heatmaps [37].
For single-cell RNA sequencing (scRNA-seq) analysis, cassava leaf and storage root data (Projects: PRJCA019992, PRJNA895163) were analyzed [40,41]. Gene expression UMAP projections and bubble plots for cassava NADP-ME genes were generated via the Omicsmart platform. Arabidopsis leaf scRNA-seq data (GEO database: GSE161482) were processed using the Seurat package (New York Genome Center, New York, NY, USA, v4.3.0). Data standardization employed the SCTransform function, and mitochondrial genes were regressed out. Dimensionality reduction utilized >50 principal components (PCA) with a clustering resolution of 0.8. UMAP visualization applied PCA = 10, neighbors = 30, and mindistance = 0.1 [42]. Maize leaf scRNA-seq data (GEO database: GSE157759) were processed using the Seurat package (thresholds: percent.pt < 4; percent.mt < 0.75; nFeature_RNA > 1800 & < 7000). Standardization (SCTransform) and clustering (PCA = 50, resolution = 0.2) were performed.
2.5. Plant Materials and Growth Conditions
The cassava cultivar SC8, developed by the Institute of Biology at Hainan University, was used in this study. Healthy tissue culture seedlings grown on Murashige and Skoog (MS) medium for two months were selected for experimental treatments. Stress treatments included cold stress (4 °C incubation), heat stress (42 °C incubation), salt stress (uniform foliar application of 300 mM NaCl solution), and drought stress (20% w/v PEG-6000 solution). All solutions were sterilized by autoclaving prior to aseptic application in a laminar flow hood. The seedlings were treated with spraying for 48 h. Leaf samples were collected at 2 h, 4 h, 8 h, 12 h, 24 h, and 48 h post-treatment (three biological replicates), flash-frozen in liquid nitrogen, and stored at −80 °C for subsequent RNA extraction.
Arabidopsis thaliana ecotype Col-0 was used as the wild-type (WT) control. Plants were cultivated under controlled conditions: 60% relative humidity, 20–22 °C constant temperature, 16 h light/8 h dark photoperiod, and 200 μmol· light intensity.
2.6. Molecular Cloning and Subcellular Localization
Total RNA was extracted from cassava leaves using the FastPure Plant Total RNA Isolation Kit (Vazyme Biomedical Technology, Nanjing, China; RC411). First-strand cDNA synthesis was performed with the HiScript III 1st Strand cDNA Synthesis Kit (Vazyme Biomedical Technology, Nanjing, China; R312). Primers for amplifying the MeNADP-ME3 gene sequence (terminator-free) were designed via SnapGene (GSL Biotech, San Diego, CA, USA, v6.0.2) software based on restriction sites within the CaMV 35S promoter-driven pCAMBIA1300-eGFP vector: 1300-eGFPMeNADP-ME3-F(Sal I): GCGTCGACATGATCTCCTTGAAA, 1300-eGFPMeNADP-ME3-F(Kpn I): GGGGTACCCCGATAGCTTCGGTA. The MeNADP-ME3 fragment was amplified from cDNA templates using PrimeSTAR Max DNA Polymerase (Takara Biomedical Technology, Beijing, China; R045A). The resulting amplicon and pCAMBIA1300-eGFP vector were digested with Sal I and Kpn I, purified, ligated, and transformed to construct the recombinant pCAMBIA1300-MeNADP-ME3-eGFP expression vector.
For subcellular localization analysis, approximately 0.2 g of leaf tissue from cassava tissue culture seedlings (cv. SC8) was used for protoplast isolation according to the Plant Protoplasts Isolation Kit (Beyotime Biomedical Technology, Shanghai, China; C0362S). The isolated protoplasts were transfected with concentrated plasmid DNA using the Plant Protoplasts Transfection Kit (Beyotime Biomedical Technology, Shanghai, China; C0563S). Post-transfection, samples were cultured overnight under low light at room temperature. Transfected protoplasts were imaged using 488 nm excitation on Olympus FV3000 (Olympus Corporation, Haikou, China) laser-scanning confocal microscope.
2.7. Gene Expression Analysis by qRT-PCR
Gene-specific primers for MeNADP-ME were designed using NCBI Primer-BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) (Table S1). β-tubulin and EF1α served as reference genes. The amplification protocol comprised 40 cycles of: 95 °C for 5 s, 58 °C for 15 s, and 72 °C for 15 s (Table S2 for reaction mixture details). Three technical replicates were performed per sample. Relative gene expression was calculated using the method [43]. Statistical analysis was conducted with SPSS (International Business Machines Corporation, Armonk, NY, USA, v23.0). Statistical significance was determined using Tukey’s test. Data visualization was performed using Origin (OriginLab, Northampton, MA, USA, 2018).
2.8. Generation and Functional Characterization of Transgenic Arabidopsis
The pCAMBIA1300-MeNADP-ME3-eGFP vector was introduced into wild-type Arabidopsis thaliana via the floral dip method [44]. Transgenic seeds were selected on half-strength Murashige and Skoog (1/2 MS) medium supplemented with 50 μg/mL hygromycin. Genomic DNA was extracted from leaves of putative transgenic seedlings using the Plant Genomic DNA Kit (Tiangen Biomedical Technology, Beijing, China; DP305). PCR verification was performed using vector-specific primers: 1300-F (CTCTAGATACATCACAATCA) and 1300-R (GCGGTTCACCAGGGTGTC). Transgene-positive plants were further screened to obtain homozygous lines (T3 generation). MeNADP-ME3 transcript levels in homozygous lines were quantified by qRT-PCR using Arabidopsis ACT2 and TUB2 as reference genes. Primer sequences are listed in Table S3.
For stress tolerance assays, transgenic Arabidopsis seeds were vernalized and grown on standard 1/2 MS medium for 5 days. For heat stress treatment, seedlings were exposed to 42 °C for 1 h, then incubated vertically at 21 °C for 7 days prior to observation and imaging. Cold stress was applied at 4 °C for 1 h. For NaCl or PEG-induced osmotic stress, seedlings were transferred to 1/2 MS medium supplemented with 100 mM NaCl or 7% PEG, respectively, and cultured vertically at 21 °C for 7 days before imaging. Root length measurements were performed in biological triplicates.
For physiological parameter determination, seven-day-old WT and transgenic Arabidopsis seedlings were transplanted into 10 cm × 10 cm pots containing a nutrient soil/vermiculite mixture (2:1, v/v). Plants were grown for 21 days under normal conditions in a substrate consisting of peat moss (Pindstrup, 0–6 mm; pH-balanced, organic matter-rich) and vermiculite (supplemented with trace elements including Fe and Mg) prior to treatment. For salt stress, plants were irrigated with 300 mM NaCl for 3 days, while drought stress was imposed via natural dehydration for 7 days. Pre- and post-stress, 0.1 g rosette leaf samples per plant were collected to quantify MDA content, levels, and activities of antioxidant enzymes (CAT, SOD, POD) using detection kits (Jiancheng Bioengineering Institute, Nanjing, China; A003, A064, A007, A001, A084). Statistical significance was determined using Tukey’s test. Data were analyzed statistically with SPSS (v23.0) and visualized using Origin (2018).
3. Results
3.1. Identification of Cassava NADP-ME Genes
A total of four MeNADP-ME genes were identified in the cassava genome through HMMER domain analysis and BLASTp searches. Table S4 summarizes the fundamental characteristics and physicochemical properties of these MeNADP-ME genes, including gene IDs, number of amino acids, molecular weight, theoretical isoelectric point, instability index, aliphatic index, and grand average of hydropathicity. Chromosomal localization analysis revealed that MeNADP-ME1 and MeNADP-ME4 reside on chromosome 16, while MeNADP-ME2 and MeNADP-ME3 are positioned on chromosomes 11 and 4, respectively (Table S5).
3.2. Phylogenetic Analysis of the NADP-ME Gene Family
Phylogenetic analysis of NADP-ME protein evolutionary relationships among cassava and other species was performed using the neighbor-joining method (Table S6). Based on branching patterns in the phylogenetic tree, plant NADP-MEs were classified into four subgroups [36]: Group I (cytosolic monocot types), Group II (plastidic monocot types), Group III (plastidic eudicot types), and Group IV (cytosolic types shared by monocots and eudicots) (Figure 1). Specifically, MeNADP-ME1 and MeNADP-ME4 formed a distinct clade within Group IV, whereas MeNADP-ME2 and MeNADP-ME3 clustered within Group III. The phylogenetic clustering patterns corresponded with evolutionary relationships and subcellular localization predictions, revealing higher homology between MeNADP-MEs and orthologs from rubber tree (Hevea brasiliensis) and castor bean (Ricinus communis), both members of the Euphorbiaceae family.
3.3. Gene Structure and Conserved Motif Analysis
Fifteen motifs were identified in the NADP-ME family of cassava, Arabidopsis, and maize (Table S7). Variations among the NADP-ME family members across the three species were principally localized to the N-terminal regions (Figure 2). Notably, MeNADP-ME1 possesses a unique N-terminal motif (Motif 14), ZmchlC4NADP-ME contains a distinctive N-terminal domain (Motif 5), while ZmchlnonC4NADP-ME lacked Motif 15 (a motif consistently conserved in other NADP-ME family members). The core functional domains, including the Malic enzyme N-terminal domain superfamily and NAD(P)-binding domain superfamily, were highly conserved across all family members. Analysis of the exon-intron structures of NADP-ME genes revealed that all members across the studied species contain introns, with the number of exons ranging from 12 to 15. ZmcytNADP-ME1 exhibited the longest gene length and a unique gene structure with extended intron regions compared to other members.
3.4. Intragenomic and Interspecific Synteny Analysis
Based on intraspecies synteny analysis of MeNADP-ME genes, six pairs of segmentally duplicated genes were identified (Figure 3A), including MeNADP-ME1 with MeNADP-ME2, MeNADP-ME3, and MeNADP-ME4; MeNADP-ME2 with MeNADP-ME3 and MeNADP-ME4; and MeNADP-ME3 with MeNADP-ME4. Gene and genome duplication represents a major evolutionary force driving biological diversity and complexity, and the results of intra-species synteny analysis indicate that segmental duplication serves as the primary driver of MeNADP-ME gene evolution. Ka/Ks ratios (nonsynonymous to synonymous substitution rates) were calculated for the identified duplicated gene pairs to infer selective pressures (Table S8). Analysis revealed that the duplicate gene pair MeNADP-ME3/MeNADP-ME4 (Ka/Ks > 1) arose under positive selection pressure, while other gene pairs exhibited Ka/Ks ratios less than 1, indicating purifying selection. To further elucidate the evolutionary relationships within the NADP-ME family, syntenic genes between cassava and plant species exhibiting different photosynthetic types were analyzed (Figure 3B–F, Table S9). The highest numbers of syntenic genes were detected between cassava and rubber tree (18 pairs) as well as the dicotyledonous plant Populus trichocarpa (18 pairs). Seven and three pairs of syntenic genes were detected with the model plants Arabidopsis and rice, respectively. Four pairs of syntenic genes were identified with the monocot C4 plant maize. These findings further elucidate the evolutionary origins and conservation patterns of MeNADP-ME genes across diverse plant lineages.
3.5. Cis-Regulatory Elements Analysis and GO/KEGG Enrichment Analysis
Seventeen distinct types of cis-acting regulatory elements were identified in the promoter regions of MeNADP-ME genes (Figure 4A, Table S10). Within the MeNADP-ME family, all sequences harbor a substantial quantity of light-responsive elements, particularly the Box 4 element, which is highly enriched in every gene (ranging from 6 to 13 instances per promoter), followed by G-box and GT1-motif elements. Regarding hormone responses, the MeNADP-ME family includes various hormone-responsive elements, such as methyl jasmonate (MeJA)-responsive CGTCA-motif and TGACG-motif, abscisic acid (ABA)-responsive ABRE elements, and ethylene-responsive ERE elements. The MeNADP-ME family also contains elements associated with diverse stress responses, including anaerobic induction (ARE), low-temperature responsiveness (LTR), drought inducibility (MBS), and defense/stress responsiveness (TC-rich repeats). Additionally, the presence of a CAT-box meristem expression element within MeNADP-ME1 suggests its potential involvement in actively proliferating meristematic tissues, linking development to stress responses. Furthermore, all MeNADP-ME family members contain MYB- and MYC-binding sites involved in transcriptional regulation. These findings demonstrate that the MeNADP-ME genes play important roles in plant adaptation to environmental shifts, stress responses, and the coordination of growth and development.
Based on screening and collation, the top ten most enriched GO terms and five enriched KEGG pathways were identified (Figure 4B,C). GO enrichment analysis revealed that the MeNADP-ME family is predominantly associated with malate metabolic process and pyruvate metabolic process under biological processes (BP) category. In the molecular function (MF) category, these proteins primarily function through protein homooligomerization, malic enzyme activity, and malate dehydrogenase (decarboxylating) (NADP+) activity. KEGG enrichment results indicated that the MeNADP-ME family is significantly enriched in core metabolic pathways related to photosynthetic carbon fixation, including pyruvate metabolism, carbohydrate metabolism, energy metabolism, and carbon metabolism. The high enrichment in these pathways underscores the central role of NADP-ME in primary metabolism and photosynthetic processes.
3.6. RNA-Seq Analysis of MeNADP-ME Genes
Through analysis of raw transcriptomic data downloaded from the ENA database (PRJNA324539) for 11 cassava tissues and organs, the expression patterns of MeNADP-ME genes across different tissues were characterized, and expression profiles were generated (Figure 5, Table S11). MeNADP-ME1 was highly expressed in all tissues except friable embryogenic callus (FEC), with the highest expression observed in the root apical meristem (RAM), followed by stems and fibrous roots—consistent with the predicted expression pattern based on prior cis-regulatory element analysis, particularly the presence of CAT-box element. MeNADP-ME2 exhibited low expression across most tissues and organs but showed elevated expression specifically in the shoot apical meristem (SAM) and lateral buds, suggesting its specialized role in shoot development. MeNADP-ME3 displayed relatively high and constitutive expression levels across multiple tissues and organs, especially in stems, lateral buds, and leaves, indicating its housekeeping function in cassava. MeNADP-ME4 was highly expressed in FEC and ovular embryogenic suspensor (OES) tissues, while expression remained low in other mature organs, suggesting its specific involvement in early embryonic development. These distinct and tissue-specific expression patterns suggest that MeNADP-ME genes have undergone functional divergence and play pivotal roles in cassava growth and development, warranting further investigation.
3.7. ScRNA-Seq Analysis of NADP-ME Genes in Cassava, Arabidopsis, and Maize
Based on single-cell RNA sequencing analyses of cassava leaves and tuberous roots (PRJCA019992, PRJNA895163), the expression patterns of MeNADP-ME genes were characterized at single-cell resolution (Figure 6 and Figure 7; Tables S12 and S13). In leaves (Figure 6), MeNADP-ME1 exhibited high expression restricted to vascular cells, albeit with relatively low cellular abundance. MeNADP-ME2 was exclusively highly expressed in a limited number of epidermal cells, showing extremely low cellular abundance across all cell types. MeNADP-ME3 showed the most prominent expression pattern, with elevated expression in bundle sheath cells and companion cells, and particularly high cellular abundance observed in bundle sheath cells, suggesting its critical role in vascular-associated metabolism. MeNADP-ME4 displayed moderate to high expression levels across multiple cell types including vascular cells, epidermal cells, and companion cells, though its cellular abundance remained relatively low in all these cell types. In tuberous roots (Figure 7), MeNADP-ME1 demonstrated prominent expression in numerous procambial cells. MeNADP-ME2 maintained high expression exclusively within sparse epidermal cells, consistent with its leaf expression pattern. In contrast, MeNADP-ME3 showed consistently low expression across diverse root cell types, markedly different from its high expression in leaves. Furthermore, MeNADP-ME4 exhibited the highest expression levels in vascular bundle cells, phloem parenchyma cells, and exodermis cells, accompanied by high cellular abundance within these populations, indicating its important role in root storage organ development and nutrient transport.
To investigate the expression patterns of NADP-ME genes across different photosynthetic plant types at single-cell resolution, we analyzed single-cell transcriptomic datasets of Arabidopsis and maize leaves (GSE161482, GSE157759; Figure 8, Tables S14 and S15). In Arabidopsis leaves (Figure 8A,B), expression was detected for only two NADP-ME genes: AtcytNADP-ME1 (corresponding to NADP-ME4 nomenclature) exhibited high expression in vascular cells, with moderate expression levels additionally observed in guard cells, epidermal cells, and bundle sheath cells. AtcytNADP-ME3 (corresponding to NADP-ME3 nomenclature) displayed a diffuse expression pattern across multiple cell types, showing higher expression in mesophyll cells and epidermal cells. In maize leaves (Figure 8C,D), four NADP-ME genes were expressed. Contrasting with the Arabidopsis expression patterns, ZmcytNADP-ME1 (LOC100286036) and ZmcytNADP-ME2 (LOC100284598) were highly expressed in mesophyll cells, although ZmcytNADP-ME2 expressing cells showed low cellular abundance. Notably, ZmchlC4NADP-ME (NADP-ME3) was highly expressed in bundle sheath cells but minimally expressed in mesophyll cells, paralleling the expression pattern of MeNADP-ME3. Furthermore, ZmchlnonC4NADP-ME (NADP-ME2) showed significant expression in mesophyll cells, albeit within a cellular population of low abundance. These comparative expression patterns across C3 (Arabidopsis), C3-C4 intermediate (cassava), and C4 (maize) species provide critical insights into the evolutionary trajectory of photosynthetic NADP-ME function.
MeNADP-ME3 exhibits an expression pattern at the single-cell level similar to that of ZmchlC4NADP-ME from the C4 plant maize, suggesting the potential existence of a limited C4 photosynthetic pathway in cassava. To further investigate this hypothesis, we analyzed the single-cell level expression profiles of photosynthesis-associated genes and PD-related genes in mesophyll cell subset 13 (adjacent to bundle sheath cells) and vascular tissue cell subsets (5, 7, 10, 12, 14) [40,45,46,47,48,49]. These findings (Figure 9) demonstrate that photosynthetic genes, including PHOSPHOENOLPYRUVATE CARBOXYLASE (PEPC, Manes_02G089900), RIBULOSE BISPHOSPHATE CARBOXYLASE SMALL CHAIN (RBCS, Manes_01G011500), HYDROXYPYRUVATE DECARBOXYLASE (HPD, Manes_02G088900), PLASMODESMATA-RELATED GENES (psaD, Manes_02G143000; psaB, Manes_04G028400), LIGHT-HARVESTING CHLOROPHYLL A/B BINDING PROTEIN 5 (LHCA5, Manes_05G173900), CHLOROPLAST RNA BINDING PROTEIN (CRD1, Manes_09G073800), and other photosynthesis-related genes (psaA, psaB, psbA), were highly co-expressed in mesophyll subset 13 and bundle sheath cells (subset 7), with substantial cellular abundance. Similarly, PD-related genes, including RH3 (Manes_09G182900) and GLK1 (Manes_13G142200), exhibited elevated expression in these cell subsets, particularly in mesophyll subset 13 (adjacent to bundle sheath) and bundle sheath cells (subset 7). This distinctive co-expression pattern of photosynthetic genes and PD-related genes in adjacent mesophyll and bundle sheath cells provides strong molecular evidence supporting the existence of a limited C4 photosynthetic mechanism in cassava, potentially facilitating intercellular transport and concentration. Transient expression of the CaMV 35S promoter-driven pCAMBIA1300-MeNADP-ME3-eGFP construct in cassava leaf protoplasts revealed chloroplast localization of MeNADP-ME3 via confocal microscopy (Figure 10). The green fluorescence signal of MeNADP-ME3-eGFP fusion protein co-localized with the red chlorophyll autofluorescence, as evidenced by the yellow merged signal in the overlay images, while the no-fused-eGFP control showed diffuse cell nucleus distribution. This chloroplast localization pattern corroborates the potential role of MeNADP-ME3 in photosynthetic carbon metabolism and supports its functional similarity to C4-type NADP-ME in cassava.
3.8. Protein Interaction Prediction and KEGG Enrichment Analysis for MeNADP-ME3
Based on protein–protein interaction network analysis of MeNADP-ME3 constructed using the STRING database (Figure 11A, Table S16), KEGG enrichment annotation revealed functional specializations of key interactors: Fumarate hydratase 1 and 2 (FUM1, FUM1-2) primarily engage in energy metabolism via the TCA cycle; Alanine aminotransferase 1 and 2 (ALAAT1, ALAAT2) predominantly participate in amino acid biosynthesis and interconversion; Malate dehydrogenase 1 and 1-1 (MDH1, MDH1-1) contribute to photosynthetic carbon assimilation and redox homeostasis regulation; Glutamate glyoxylate aminotransferase 2 and 2-1 (GGAT2, GGAT2-1) are mainly involved in photorespiration and amino acid metabolism; while Malate synthase (MLS) operates within the glyoxylate cycle (Figure 11B, Table S17). The interaction network showed strong connectivity between MeNADP-ME3 and its partners, with MLS exhibiting the highest interaction score, indicating a central metabolic hub. MeNADP-ME3 and its interacting partners were jointly enriched in carbon metabolism (highest significance), carbon fixation by Calvin cycle, pyruvate metabolism, and arginine biosynthesis pathways. Notably, the high enrichment in carbon metabolism and the strong interaction with MLS suggest that the low photorespiration characteristic of cassava may be attributed to the coordinated regulation between NADP-ME-mediated malate decarboxylation and the glyoxylate cycle, potentially providing an alternative pathway to bypass photorespiratory loss.
3.9. Expression Profiles of MeNADP-ME Under Different Abiotic Stresses
To investigate the expression patterns of MeNADP-ME in response to various abiotic stresses, qRT-PCR analysis was performed on leaves of cassava plantlets subjected to different stress treatments. Under 4 °C cold stress treatment (Figure 12A), MeNADP-ME1 expression was significantly downregulated at 2 h post-treatment. Although it recovered to baseline levels at 4 h, it was significantly downregulated again at 8 h and remained suppressed through 48 h. Similarly, MeNADP-ME2 expression decreased transiently at 2 h before returning to control levels, but exhibited significant downregulation at 8 h and remained low thereafter. MeNADP-ME3 expression was transiently suppressed at 2 h, followed by significant upregulation at 4 h, before rapidly declining. However, it showed significant upregulation again at 24 h post-treatment, reaching its peak expression. In contrast, MeNADP-ME4 expression was significantly and dramatically upregulated, peaking at 2 h, followed by a rapid decline to baseline levels by 8 h. Following 42 °C heat stress (Figure 12B), MeNADP-ME1 expression was significantly upregulated starting at 12 h and reached its peak at 48 h. MeNADP-ME2 expression initially declined transiently at 2 h, was significantly upregulated at 8 h, and then rapidly decreased to significantly lower levels at 24 h. MeNADP-ME3 expression was rapidly and significantly induced, peaking at 4 h, before gradually declining over time. MeNADP-ME4 expression was significantly upregulated after heat treatment, peaking at 24 h, displaying a trend similar to that of MeNADP-ME1. The distinct biphasic or sustained expression patterns of the four MeNADP-ME genes under cold and heat stresses suggest functional specialization: certain genes (MeNADP-ME3, MeNADP-ME4) may be associated with rapid cold/heat shock response, while others (MeNADP-ME1) may function in long-term temperature acclimation. This divergence reflects the differential regulatory strategies employed by plants to adapt and respond to thermal fluctuations and diverse environmental challenges.
Following salt stress treatment (300 mM NaCl) (Figure 12C), all MeNADP-MEs exhibited pronounced responses with distinct temporal dynamics. The expression levels of MeNADP-ME1 and MeNADP-ME4 were immediately and significantly downregulated at 2 h, though both showed gradual recovery trends over time. Notably, MeNADP-ME4 rebounded to baseline levels at 24 h post-treatment before rapidly declining again at 48 h. In contrast, MeNADP-ME2 and MeNADP-ME3 were significantly upregulated immediately after salt treatment, with MeNADP-ME3 reaching peak expression at 2 h, but both dropped sharply thereafter. MeNADP-ME2 subsequently exhibited a secondary upregulation, peaking at 48 h, suggesting a potential role in long-term salt adaptation. Under drought stress simulated by 20% PEG (Figure 12D), MeNADP-ME1 and MeNADP-ME3 were markedly induced at 2 h but declined significantly to basal levels after 8 h of treatment, indicating an early rapid response mechanism. Conversely, MeNADP-ME2 and MeNADP-ME4 displayed downregulation trends throughout the treatment period. Given that both salt and drought stresses cause osmotic imbalance and oxidative damage in plants, the divergent and temporally distinct response patterns of these four MeNADP-ME genes suggest stress-specific regulatory mechanisms and functional specialization underlying plant adaptation to distinct environmental challenges. The rapid induction of MeNADP-ME3 under both salt (early phase) and drought stresses further supports its critical role in osmotic stress responses.
3.10. Overexpression of MeNADP-ME3 Confers Enhanced Tolerance to Salt and Drought Stresses in Arabidopsis
Transgenic Arabidopsis plants overexpressing MeNADP-ME3 were generated via the floral dip method. Four independent transgenic lines (OE1-4) were obtained following screening on hygromycin-containing medium (Figure 13A). PCR amplification with vector-specific primers confirmed the presence of the transgene in all four lines, while no amplification was observed in wild-type (WT) or blank control (K). Total RNA integrity was verified by agarose gel electrophoresis (Figure 13B), showing clear ribosomal RNA bands. RT-qPCR analysis confirmed MeNADP-ME3 expression levels in transgenic lines (Figure 13C), with OE1 showing the highest expression, followed by OE3 and OE4. Three lines (OE1, OE3, and OE4) with the highest and most consistent relative expression were selected for subsequent functional experiments. As shown in Figure 13D,E, no significant difference in primary root length was observed between WT and transgenic seedlings under control conditions, confirming that MeNADP-ME3 overexpression does not affect normal development. Following exposure to various stresses, transgenic lines exhibited significantly longer primary roots than WT under PEG-mediated drought stress and NaCl salt stress, representing approximately 60% and 40% improvements, respectively. Conversely, temperature stresses (cold at 4 °C for 1 h/heat at 42 °C for 1 h) produced no distinct difference in root elongation between genotypes, with both showing similar root lengths. These findings indicate that MeNADP-ME3 overexpression specifically enhances tolerance to osmotic and ionic stressors but does not alter temperature stress responses in Arabidopsis, suggesting its primary function in maintaining cellular homeostasis under water-deficit conditions.
Following 21-day growth under soil culture conditions, stress tolerance-related parameters were measured in WT and transgenic plants subsequent to salt and drought stress treatments (Figure 14). Malondialdehyde (MDA) and contents are established indicators for evaluating membrane lipid peroxidation and oxidative stress status, respectively. Under control conditions, basal MDA levels and contents were comparable between WT and transgenic lines. After salt stress (300 mM NaCl for 3 days), both MDA and levels increased significantly in all plants; however, transgenic lines exhibited significantly lower accumulation compared to WT (Figure 14A,B). Specifically, WT plants showed elevated MDA content and levels under salt stress, while transgenic lines maintained significantly lower levels, representing approximately 27% and 32% reductions, respectively. Similar patterns were observed under drought stress (natural dehydration for 7 days), with transgenic lines showing approximately 20–25% lower MDA and accumulation compared to WT.
Activities of antioxidant enzymes (CAT, SOD, POD) constitute additional critical metrics in stress response studies, as they catalyze the detoxification of reactive oxygen species (ROS). As shown in Figure 14C,D, no significant differences in these enzyme activities were detected between WT and transgenic plants under control conditions. In contrast, under salt stress, all transgenic Arabidopsis lines displayed significantly higher antioxidant enzyme activities than WT: CAT activity increased by approximately 30%, SOD activity by approximately 75%, and POD activity by approximately 120%. Under drought stress, similar enhancement patterns were observed, with OE lines showing 20–25% higher CAT, 70–80% higher SOD, and 80–90% higher POD activities compared to WT. Collectively, these results demonstrate that overexpression of MeNADP-ME3 enhances salt and drought tolerance in transgenic Arabidopsis by augmenting antioxidant enzyme capacity, thereby reducing oxidative damage and maintaining cellular redox homeostasis under stress conditions.
4. Discussion
NADP-ME is ubiquitously distributed across diverse photosynthetic lineages, encompassing algae, mosses, ferns, and seed plants [50]. The majority of C4 species belong to monocotyledons, particularly within the Poaceae (4600 species) and Cyperaceae (1600 species). In contrast, only approximately 1600 C4 species have been identified in dicotyledons, distributed across 16 families, with nearly 75% concentrated within four dominant families: Amaranthaceae, Euphorbiaceae, Asteraceae, and Caryophyllaceae [51]. In this study, four MeNADP-ME genes were identified in the cassava genome through integrated HMM domain analysis and BLASTp homology searches. Evolutionary analyses revealed that although the cassava genome (640 Mb) is nearly twice the size of the rice genome (386 Mb), both possess an identical number of NADP-ME genes. This finding also supports the notion that expansion of the NADP-ME gene family did not result from whole-genome duplication events, but rather from lineage-specific expansion patterns following the divergence of monocotyledons and dicotyledons [36,52].
Phylogenetic analysis provides critical insights into evolutionary relationships among species, genes, or populations. In this study, phylogenetic analysis was conducted to investigate evolutionary relationships among NADP-ME proteins from cassava and diverse plant lineages with varying photosynthetic types. These NADP-MEs were categorized into four clades: Group I (cytosolic monocot types), Group II (plastidic monocot types), Group III (cytosolic eudicot types), and Group IV (cytosolic types shared by monocots and eudicots) [36]. Phylogenetic clustering aligned with evolutionary relationships and subcellular localization. Cassava displays closer phylogenetic affinity to its congeneric Euphorbiaceae species rubber tree and castor bean, as well as to Arabidopsis, Populus trichocarpa, and Flaveria bidentis, but exhibits distant relationships with rice, maize, and sweet sorghum. Notably, conserved motif analysis revealed interspecific divergence primarily within N-terminal regions across Arabidopsis, cassava, and maize NADP-ME families. MeNADP-ME1 possessed a unique N-terminal motif (Motif 14), whereas ZmchlC4NADP-ME featured a distinctive N-terminal domain (Motif 5), and ZmchlnonC4NADP-ME lacked the N-terminal motif (Motif 15). These structural divergences correlate with the expansion of the NADP-ME gene family during the evolutionary trajectory toward C4 photosynthesis in cassava and maize. Furthermore, six segmentally duplicated gene pairs were identified among MeNADP-ME genes, with MeNADP-ME3/MeNADP-ME4 (Ka/Ks ) originating under positive selection. Interspecies synteny analysis revealed that cassava shares fewer syntenic genes with monocots (rice, maize), whereas it exhibits greater synteny with eudicots (Arabidopsis, Populus trichocarpa). Notably, the highest number of syntenic genes was identified between cassava and its congeneric Euphorbiaceae species, the rubber tree. This finding further indicates the close evolutionary association of NADP-ME genes with species diversification and lineage divergence.
Cis-acting regulatory elements (CAREs) serve as specific functional modules on DNA sequences, which play critical roles in the dynamic regulation of gene expression [53,54]. Multiple CAREs associated with plant development and growth, hormone response, and stress response were identified in the upstream regions of cassava NADP-ME coding sequences. These types of elements are also commonly present within the upstream regions of NADP-ME genes in Arabidopsis and Brassica napus [33,55]. These findings indicate that NADP-ME genes exhibit broad responsiveness to external environmental stimuli and participate in mediating the regulation of gene expression and various physiological processes at the transcriptional level.
MeNADP-ME exhibits distinct expression patterns across different tissues and organs. Genes with high expression levels in specific tissues or organs are likely associated with particular physiological processes or functions crucial to cassava growth and development, potentially playing a vital role in these specific locations. The homologous genes AtNADP-ME2 and AtNADP-ME4 in Arabidopsis are involved in regulating plant organ development and growth and in responding to abiotic stress [23].
Single-cell RNA sequencing enables construction of high-resolution gene expression profiles at the cellular level, providing novel insights into key biological processes such as plant development and stress responses, and has emerged as a pivotal technique in modern plant research [56]. This study revealed that MeNADP-ME3 exhibits relatively high expression levels in the bundle sheath cells of cassava leaves, suggesting the potential existence of a limited C4 photosynthetic pathway in cassava leaves. This observation aligns with the tissue-specific high expression pattern of ZmchlC4NADP-ME in bundle sheath cells observed in the typical C4 plant maize [57]. In contrast, within the C3 model plant Arabidopsis, expression of AtcytNADP-ME1 is primarily localized to vascular cells, while AtcytNADP-ME3 is expressed at higher levels within mesophyll cells [42]. Cross-species comparisons demonstrate that the expression pattern of NADP-ME genes in bundle sheath cells indicates that MeNADP-ME3 likely fulfills a functional role analogous to its counterpart within the C4 photosynthetic pathway in cassava. Further single-cell clustering analysis revealed evolutionarily conserved expression patterns and functional adaptation of the MeNADP-ME gene family across distinct cell subsets. For instance, MeNADP-ME1 and MeNADP-ME4 are co-expressed at high levels in subsets of cells associated with substance transport in both cassava leaves and tuberous roots, suggesting potential synergistic involvement in metabolite translocation, whereas MeNADP-ME2 is only minimally enriched in epidermal cells. These tissue-specific expression patterns not only reflect functional divergence and coordination among members of the NADP-ME gene family but also provide transcriptional regulatory evidence supporting their classification within a gene family possessing a shared evolutionary origin.
Cassava is characterized as a C3-C4 intermediate species that, although lacking complete Kranz anatomy, displays characteristic C4 traits such as high photosynthetic efficiency and low photorespiration [58]. Significantly, the activity of phosphoenolpyruvate carboxylase (PEPC), a key enzyme in the C4 pathway, is markedly higher in cassava than in typical C3 plants [5]. Plasmodesmata (PD), which serve as nanoscale conduits for intercellular transport in plants, play a pivotal role in the concentration mechanism (CCM) of C4 plants [59]. Previous studies identified mesophyll cells possessing transport functions surrounding bundle sheath (BS) cells in cassava [40]. Our scRNA-seq analysis of mesophyll cell subset 13 adjacent to BS cells and vascular tissue cell subsets revealed a co-upregulation of photosynthesis-related genes and PD-associated genes in both mesophyll subset 13 and BS cells. Furthermore, subcellular localization analysis indicated that MeNADP-ME3 localizes to the chloroplasts. These findings collectively suggest the presence of a limited C4 photosynthetic pathway in cassava. Protein–protein interaction prediction for MeNADP-ME3 identified malate synthase (MLS), an enzyme involved in photorespiration, as its highest-scoring interactor. This interaction provides a plausible molecular basis underlying cassava’s characteristic low photorespiration phenotype through potential regulatory mechanisms. Consequently, cassava serves as an ideal natural model system for investigating the evolutionary mechanisms of the C4 photosynthetic pathway, thereby holding significant value for deciphering plant adaptive evolution and the genetic improvement in stress resistance in crops.
C4 plants generally exhibit greater thermotolerance and lower photorespiration rates compared to C3 plants. This advantage primarily arises from their CCM, which sustains photosynthetic activity under low atmospheric concentrations induced by high-temperature-triggered stomatal closure. This mechanism suppresses the oxygenase activity of Rubisco, thereby minimizing energy-wasting photorespiration. Investigation of the MeNADP-ME gene family in cassava revealed distinct expression patterns under abiotic stresses. Most members were highly responsive to thermal extremes: transcript levels were significantly downregulated following cold stress yet universally upregulated under heat stress. This differential responsiveness aligns with cassava’s intrinsic attributes of heat tolerance, high photosynthetic efficiency, and low photorespiration. During PEG-simulated drought stress, MeNADP-ME1 and MeNADP-ME3 exhibited significant upregulation—a pattern consistent with NADP-ME responses in tobacco and wheat (Triticum aestivum) leaves [17,60]. Similarly, salt stress induced marked upregulation of MeNADP-ME2 and MeNADP-ME3, corroborating cis-element predictions and echoing osmotic stress-responsive expression patterns observed in sweet sorghum [23]. The differential expression patterns of plant NADP-ME genes under environmental constraints reflect both their adaptive evolution during phylogeny and their functional specialization in mediating lineage-specific stress adaptation mechanisms.
Previous studies have validated the critical functional role of plastidic NADP-ME in plant stress responses [23,61]. In this study, we generated Arabidopsis transgenic lines overexpressing MeNADP-ME3 with chloroplast-directed localization. Root elongation assays under diverse stress treatments demonstrated that primary root lengths of transgenic lines were significantly greater than WT counterparts under drought and salt stress, indicating the gene’s potential to sustain root physiological activity for growth maintenance under adversity. When exposed to environmental stressors, ROS accumulation induces oxidative damage, subsequently compromising cellular integrity through destruction of membrane systems, DNA, proteins, and lipids [62]. Malondialdehyde (MDA) is the final product of membrane peroxidation and serves as an index to measure the oxidative damage of plant cell membranes [63]. (a predominant ROS in plants) acts both as a signaling molecule regulating physiological processes via aquaporin-mediated transmembrane transport and as an indicator of oxidative stress [64,65]. Transgenic lines exhibited significantly lower MDA content and accumulation relative to WT following drought and salt treatments, demonstrating the pivotal function of MeNADP-ME3 overexpression in mitigating oxidative injury.
Concurrently, activities of antioxidant enzymes (SOD, POD, CAT) were quantified. Superoxide dismutase (SOD) constitutes the primary defense by scavenging superoxide anions, while peroxidase (POD) and catalase (CAT) principally eliminate [66]. Transgenic lines displayed significantly enhanced SOD, POD, and CAT activities compared to WT under both stress conditions. MeNADP-ME3 may confer drought and salt tolerance in transgenic Arabidopsis through multiple mechanisms. It may indirectly upregulate antioxidant enzymes, thereby enhancing ROS scavenging and alleviating oxidative stress. Furthermore, it may promote osmotic adjustment and reduce membrane lipid peroxidation, as reflected by decreased MDA content. These combined effects ultimately improve the stress tolerance of transgenic lines.
5. Conclusions
This study reveals that the NADP-ME gene family in cassava is comparatively compact, with segmental duplication representing the primary evolutionary driving force. MeNADP-ME genes exhibit strikingly divergent expression profiles across different tissues and organs. Furthermore, their expression patterns within distinct cellular subpopulations in the single-cell RNA sequencing (scRNA-seq) demonstrate significant evolutionary conservation and functional adaptability. Subcellular localization confirmed MeNADP-ME3 targeting to chloroplasts in cassava leaves. ScRNA-seq analysis revealed its predominant expression in bundle sheath cells (BS), with an associated mesophyll subcluster 13 showing elevated transcription of photosynthesis-associated genes and plasmodesmata (PD)-related genes. These findings suggest the existence of a rudimentary limited C4 photosynthetic apparatus in cassava leaves. MeNADP-ME promoters harbor diverse cis-regulatory elements, indicating crucial roles in environmental adaptation, stress response, and developmental coordination. Differential expression analysis under environmental stresses identified MeNADP-ME3 as markedly responsive to multiple abiotic stressors, suggesting its functional significance in stress adaptation. Overexpression of MeNADP-ME3 in Arabidopsis was consequently performed for functional investigation. Under drought and salt stress, MeNADP-ME3 overexpression likely potentiates antioxidant enzyme activity (SOD, POD, CAT) through indirect transcriptional regulation, thereby enhancing ROS scavenging efficacy. Concurrently, it augments osmoregulatory capacity and reduces membrane peroxidation, ultimately conferring elevated drought and salinity tolerance in transgenic lines.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li S. Cui Y. Zhou Y. Luo Z. Liu J. Zhao M. The industrial applications of cassava: Current status, opportunities and prospects J. Sci. Food Agric.2017972282229010.1002/jsfa.828728233322 · doi ↗ · pubmed ↗
- 2Zhu F. Composition, structure, physicochemical properties, and modifications of cassava starch Carbohydr. Polym.201512245648010.1016/j.carbpol.2014.10.06325817690 · doi ↗ · pubmed ↗
- 3Amelework A.B. Bairu M.W. Advances in Genetic Analysis and Breeding of Cassava (Manihot esculenta Crantz): A Review Plants 202211161710.3390/plants 1112161735736768 PMC 9228751 · doi ↗ · pubmed ↗
- 4De Souza A.P. Massenburg L.N. Jaiswal D. Cheng S. Shekar R. Long S.P. Rooting for cassava: Insights into photosynthesis and associated physiology as a route to improve yield potential New Phytol.2017213506510.1111/nph.1425027778353 · doi ↗ · pubmed ↗
- 5El-Sharkawy M.A. Prospects of photosynthetic research for increasing agricultural productivity, with emphasis on the tropical C 4 Amaranthus and the cassava C 3-C 4 crops Photosynthetica 20165416118410.1007/s 11099-016-0204-z · doi ↗
- 6Wang H. Shen X. Lu C. Li K. Wang W. Molecular Functional Characterisation of Mechl PPDK Promoter in Transgenic Tobacco Trop. Plant Biol.20201328729510.1007/s 12042-020-09257-0 · doi ↗
- 7Danila F.R. Quick W.P. White R.G. Furbank R.T. von Caemmerer S. The Metabolite Pathway between Bundle Sheath and Mesophyll: Quantification of Plasmodesmata in Leaves of C 3 and C 4 Monocots Plant Cell 20162814611471 Erratum in Plant Cell 2018, 30, 51310.1105/tpc.16.0015527288224 PMC 4944413 · doi ↗ · pubmed ↗
- 8Alvarez C.E. Bovdilova A. Höppner A. Wolff C.C. Saigo M. Trajtenberg F. Zhang T. Buschiazzo A. Nagel-Steger L. Drincovich M.F. Molecular adaptations of NADP-malic enzyme for its function in C 4 photosynthesis in grasses Nat. Plants 2019575576510.1038/s 41477-019-0451-731235877 · doi ↗ · pubmed ↗