Epidemiological Insights into Carbapenem-Resistant Enterobacterales Throughout the COVID-19 Pandemic in Buenos Aires, Argentina
Francisco González-Espinosa, Francisco Magariños, Sofía Ciminello, Roque Figueroa-Espinosa, María Sol Haim, Tomas Poklepovich, Nicolas Potente, Cecilia Ormazabal, Gabriel Gutkind, Daniela Cejas, Marcela Radice

TL;DR
This study examines how antibiotic-resistant bacteria, specifically carbapenem-resistant Enterobacterales, evolved during the COVID-19 pandemic in Buenos Aires, Argentina.
Contribution
The study reveals a shift in resistance patterns and the spread of specific carbapenemase-producing Klebsiella pneumoniae clones during the pandemic.
Findings
CRE isolates showed a significant shift from multidrug-resistant to extensively drug-resistant phenotypes during the pandemic.
blaKPC-2 and blaNDM-5 were the most prevalent carbapenemase alleles in Klebsiella pneumoniae.
The CG258-tonB79 clone of Klebsiella pneumoniae became dominant in both hospitals by 2022.
Abstract
Background: Carbapenem-resistant Enterobacterales (CRE) are a global public health concern, with carbapenem-resistant Klebsiella pneumoniae (CR-Kp) recognised as the highest-priority pathogen. This study aimed to investigate the epidemiological features of CRE isolates throughout the COVID-19 pandemic in Buenos Aires, Argentina. Methods: A prospective study was conducted in two hospitals from 2019 to 2022, recovering all CRE from inpatients. Antimicrobial susceptibility was performed by automated and/or manual tests, according to CLSI. β-lactamases detection was performed using Multiplex PCR and MALDI-TOF MS. Kp typing was assessed by multiplex PCR and/or MLST based on WGS. Results: 22% (359/1594) were CRE, predominantly CR-Kp. Overall, high non-susceptibility (NS) rates were observed in both centres. NS remained largely stable in HA, except for a significant increase in colistin NS,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
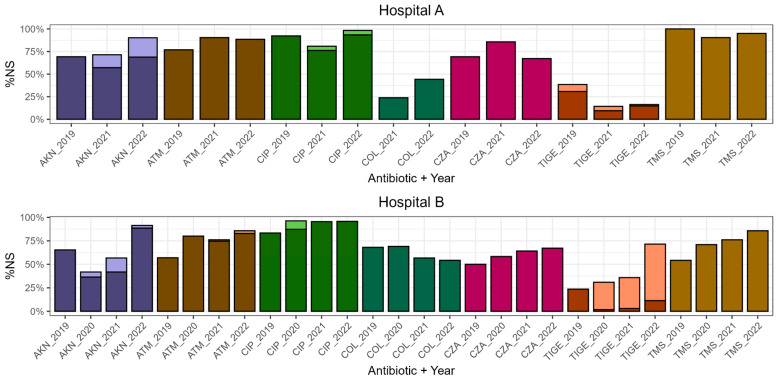 Figure 1
Figure 1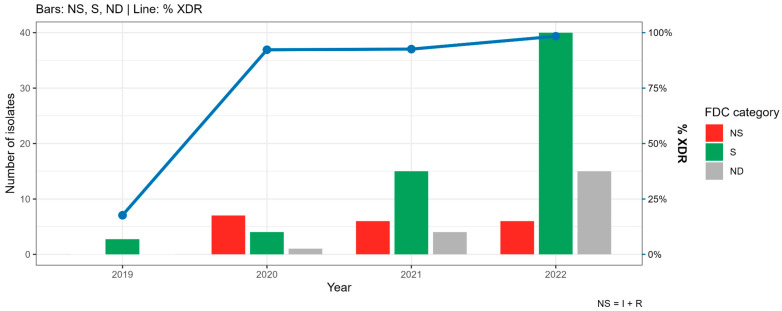 Figure 2
Figure 2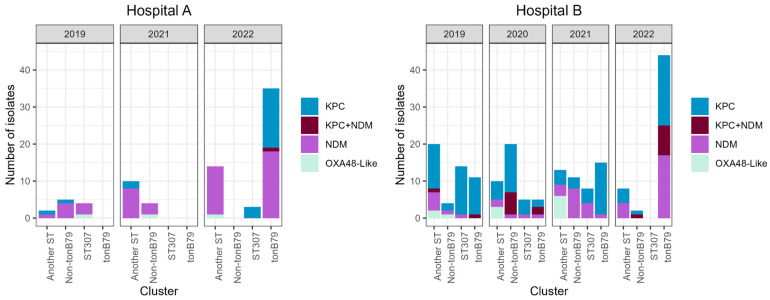 Figure 3
Figure 3| Gene | Primer | Sequence 5′-3′ | Product Size (bp) | Reference | Purpose |
|---|---|---|---|---|---|
|
| KPCm-F | CGTCTAGTTCTGCTGTCTTG | 798 | Poirel et al. (2011) [ | β-lactamase detection |
| KPCm-R | CTTGTCATCCTTGTTAGGCG | ||||
| KPC-F b | ATGTCACTGTATCGCCGTC | 890 | Bradford et al. (2004) [ | ||
| KPC-R b | TTTTCAGAGCCTTACTGCCC | ||||
|
| NDM-F a | GGTTTGGCGATCTGGTTTTC | 621 | Poirel et al. (2011) [ | |
| NDM-R a | CGGAATGGCTCATCACGATC | ||||
|
| OXAm-F | GCGTGGTTAAGGATGAACAC | 438 | Poirel et al. (2011) [ | |
| OXAm-R | CATCAAGTTCAACCCAACCG | ||||
|
| VIM-F | GATGGTGTTTGGTCGCATA | 390 | Poirel et al. (2011) [ | |
| VIM-R | CGAATGCGCAGCACCAG | ||||
|
| IMP-F | GGAATAGAGTGGCTTAAYTCTC | 232 | Poirel et al. (2011) [ | |
| IMP-R | GGTTTAAYAAAACAACCACC | ||||
|
| SPM-F | AAAATCTGGGTACGCAAACG | 271 | Poirel et al. (2011) [ | |
| SPM-R | ACATTATCCGCTGGAACAGG | ||||
|
| CIT-MF | TGGCCAGAACTGACAGGCAAA | 462 | Perez-Perez et al. (2002) [ | |
| CIT-MR | TTTCTCCTGAACGTGGCTGGC | ||||
|
| CTX-MU-1 | ATG TGC AGY ACC AGT AAR GT | 593 | Pagani et al. (2003) [ | |
| CTX-MU-2 | TGG GTR AAR TAR GTS ACC AGA | ||||
|
| PER2 PLUS | GTAGTATCAGCCCAATCCCC | 739 | Pasteran et al. (2006) [ | |
| PER 2 MINUS | CCAATAAAGGCCGTCCATCA | ||||
|
| pilV-F3 | CGATGGCGCTGGCGACGATTAT | 627 | Yu et al. (2018) [ | |
| pilV-R3 | CCCGATGGGCAAGAACATGCGT | ||||
|
| kphp-F3 | TGGCGGGTAATGCCCGATCAGT | 236 | Yu et al. (2018) [ | |
| kphp-R3 | AGGCCGCTTTCCATAAGCCGTT | ||||
|
| pilL-F2 | CGGTATTTGCTCTGCGTGATAG | 350 | Yu et al. (2018) [ | |
| pilL-R2 | TGGTTATACAGAACGGCATTGG | ||||
|
| Khe-F3 | CCGGAGCGTTTTTCAATCGGCG | 441 | Yu et al. (2018) [ | |
| Khe-R3 | CGCTTCGCCCCTCACCTGAAAT | ||||
|
| ST-307-F | AGGAAAGTCGCCGCAGGAGGAT | 628 | Lowe et al. (2011) [ | |
| ST-307-R | TGCTGCTGCCATAAAACGCACCT |
- —Universidad de Buenos Aires
- —Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
- —Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación
- —CONICET Scholarship
- —UBA Scholarship
- —Scientific and Technological Researcher Career (CIC), CONICET
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Nosocomial Infections in ICU
1. Introduction
Carbapenems are broad-spectrum β-lactam antibiotics widely used for the treatment of multidrug-resistant (MDR) Gram-negative infections. Representative agents include imipenem, meropenem, doripenem, and ertapenem, all of which are extensively used worldwide [1]. Their clinical use expanded considerably following the global dissemination of extended-spectrum β-lactamase (ESBL)-producing bacteria, which hydrolyse most β-lactam antibiotics except carbapenems. However, the increased use of these agents has been accompanied by a marked rise in bacterial isolates producing carbapenemases (β-lactamases capable of hydrolysing carbapenems). As a consequence, carbapenem-resistant Enterobacterales (CRE), defined by resistance to at least one carbapenem and/or by carbapenemase production, have emerged as a major global health threat [1,2]. CRE have been designated as critical priority pathogens by the World Health Organisation (WHO), representing the highest level of concern for public health. In the updated WHO bacterial priority pathogens list published in 2024, carbapenem-resistant Klebsiella pneumoniae was ranked as the highest-priority pathogen worldwide [3]. According to the national surveillance programme, in Argentina, 36% of K. pneumoniae isolates recovered from inpatients in 2024 (n: 6922) were resistant to carbapenems (https://antimicrobianos.com.ar/2025/10/tablero_ram/, accessed on 15 December 2025). In our region, the predominant mechanism of carbapenem resistance among CRE is the production of acquired carbapenemases, mainly Klebsiella pneumoniae carbapenemase (KPC) and New Delhi metallo-β-lactamase (NDM) [4]. According to the Ambler molecular classification of β-lactamases, the carbapenemases most frequently reported worldwide belong to classes A, B, and D. Class A (e.g., KPC) and class D (e.g., OXA-48-like) enzymes are serine carbapenemases, whereas class B enzymes, such as NDM, are metallo-β-lactamases (MBL) that require zinc ions for catalytic activity [5]. KPC hydrolyses cephalosporins, monobactams, and carbapenems and is not inhibited by clavulanate or tazobactam; however, it is effectively inhibited by newer β-lactamase inhibitors such as avibactam, relebactam, and vaborbactam [6]. KPC is the main worldwide distributed carbapenemase, and to date, there are 284 KPC variants described in the β-lactamase database [7,8]. However, KPC-2 and KPC-3 remain the most prevalent. The global expansion of KPC-2/-3 was mediated by the presence of their coding gene in the transposon Tn4401, which was located in the successful K. pneumoniae sequence type 258 (ST258). In Argentina, KPC-2-producing K. pneumoniae was established in 2010 and achieved epidemic status in many settings, associated with this pandemic clone. K. pneumoniae ST258 remained prevalent over the following five years, when a change in its epidemiology was described [9,10]. Its almost absolute prevalence was replaced by the emergence and dissemination of more virulent lineages such as ST25 and ST11, and the worrisome high-risk clone ST307. In this country, KPC-3-K. pneumoniae-ST307 was first detected in 2017 [10].
Meanwhile, NDM is capable of hydrolysing almost all β-lactams except aztreonam. More than ninety variants of NDM have been described, with NDM-1 followed by NDM-5 being the most prevalent worldwide in Enterobacterales [8]. NDM-producing CRE are commonly resistant to almost all antimicrobial agents because of the co-occurrence of additional resistance mechanisms [11]. blaNDM-1 was detected in different plasmid replicons (IncFII, IncL/M, IncN, IncR, IncHIB-M/FIB-M, IncX3), and was rarely found to be chromosomally integrated [12]. In Argentina, NDM-1 was first detected in Providencia rettgeri in 2014, and this marker remained sporadically reported in other Enterobacterales [13,14]. In 2018, the emergence and clonal expansion of K. pneumoniae ST307 isolates co-producing NDM-1 and KPC-3, reported in one hospital of Buenos Aires, alerted of a worrisome scenario, regarding the remarkable features of K. pneumoniae ST307 and its resistance profile to ceftazidime/avibactam [14]. On the other hand, NDM-5 was first described in 2021 in Escherichia coli clonal complex 354 recovered from an inpatient in another hospital in Buenos Aires [15].
OXA-48-type enzyme is a carbapenem-hydrolysing class D β-lactamase first described in 2001 in a K. pneumoniae isolate in Turkey [16]. Up to now, more than 80 OXA-48-like variants have been reported in clinical Enterobacterales isolates worldwide [8]. OXA-48 hydrolyses penicillins at a high level and carbapenems at a low level; this enzyme shows very weak activity against expanded-spectrum cephalosporins [17]. In Argentina, carbapenem-hydrolysing OXA-48-like enzymes are sporadically reported; OXA-438 was detected in 2014 in an E. coli isolate, and later, OXA-232 was detected in 2019 in an E. coli ST744 isolate that co-produced MCR-1 [18,19].
Oftentimes, CRE are frequently classified as difficult-to-treat (DTR) pathogens, as they exhibit resistance to first-line therapeutic options currently available in our country, including carbapenems, β-lactam/β-lactamase inhibitor combinations, and fluoroquinolones [20]. The co-production of carbapenemases, particularly KPC and NDM, constitutes an emerging and worrisome scenario, as it compromises the efficacy of novel β-lactam/β-lactamase inhibitor combinations and further narrows therapeutic alternatives [21]. In this context, cefiderocol, a siderophore cephalosporin active against carbapenem-resistant Gram-negative pathogens, has emerged as a valuable alternative for the treatment of severe infections with limited therapeutic options [22].
This study aimed to investigate and to comparatively analyse the epidemiological features of CRE isolates recovered throughout the COVID-19 pandemic in two hospitals of the Buenos Aires Metropolitan Area, Argentina.
2. Results
2.1. Antimicrobial Susceptibilities
During the prospective study, a total of 1594 Enterobacterales were isolated, with 359 (22%) resistant to at least one carbapenem (CRE) (95 CRE were recovered in HA and 264 CRE in HB). The proportion of CRE recovered in HA was: 13/252, 21/257 and 61/233, in 2019, 2021, and 2022, respectively. A significant increase in CRE was detected comparing pre (5% CRE) and post-pandemic (26% CRE) periods (p-value < 0.0001). Meanwhile, the proportion in HB was 72/128, 55/120, 67/284, and 70/320 in 2019, 2020, 2021 and 2022, respectively. Conversely, a significant decrease in CRE was observed in HB (p-value < 0.0001).
K. pneumoniae was the prevalent species of CRE (74%) in both hospitals in all periods, followed by Providencia stuartii (12%) and Escherichia coli (4%) (Table 1 and Table S1). Blood (42%) and urine (33%) were the most common sample types for CRE recovery across all periods in HA and HB. CRE were less frequently recovered from respiratory samples (9%) (Table S1).
Non-susceptibility (NS) rates to aztreonam, ceftazidime/avibactam (CZA), ciprofloxacin, amikacin, trimethoprim-sulfamethoxazole (TMS), tigecycline, and colistin among CRE isolates are presented in Figure 1.
In HA, the highest NS rates observed during the study period were 90% for aztreonam (2021), 86% for CZA (2021), 98% for ciprofloxacin (2022), 90% for amikacin (2022), 100% for TMS (2019), and 38% for tigecycline (2019), which remained stable across study periods (with no statistically significant differences). In contrast, colistin NS rates increased from 0% in 2019 to 24% in 2021 and 44% in 2022, showing a significant rise when comparing the pre- and post-COVID-19 pandemic periods (p-value = 0.003).
In HB, NS to aztreonam increased significantly from 57% in 2019 to 80% in 2020 (p-value = 0.008), a trend that persisted into the post-pandemic period. Significant increases between 2019 and 2022 were observed for CZA (50% to 67%, p-value = 0.0006), TMS (54% to 86%, p-value < 0.0001), and tigecycline (24% to 71%, p-value < 0.0001). For ciprofloxacin, NS rose significantly between 2019 and 2020 (83% to 96%, p-value = 0.002). Amikacin NS ranged from 42% in 2020 to 91% in 2022, representing a significant increase over time (p-value = 0.0002). In contrast, colistin NS rates ranged from 54% (2022) to 69% (2020), with no significant variation across study periods.
Given the resistance profiles, 82% of CRE were MDR during the COVID-19 pre-pandemic period, while the remaining 18% were XDR. Following the COVID-19 pandemic, XDR isolates increased significantly, reaching up to 88% (p-value < 0.0001). Additionally, in 2019, 56% of CRE were DTR, all of which were K. pneumoniae. DTR isolates increased significantly in 2022, up to 84% (p-value < 0.0001) (Table S1).
Regarding the evolution of antimicrobial resistance across the COVID-19 pandemic periods in the main pathogen, K. pneumoniae, a significant increase in NS isolates was observed in HA only for colistin (0% in 2019 vs. 46% in 2022, respectively, p-value = 0.0044). In comparison, in HB were detected a significant increase in NS isolates to amikacin (53% in 2019 vs. 91% in 2022, p-value < 0.0001), aztreonam (82% in 2019 vs. 100% in 2020–2022, p-value = 0.0008), CZA (27% in 2019 vs. 70–61% in 2020–2022, p-value = 0.0007), tigecycline (0% in 2019 vs. 17–69% in 2020–2022, p-value < 0.0001), and TMS (65% in 2019 vs. 87–94% in 2020–2022, p-value = 0.0003) (Table S1). With respect to cefiderocol activity, 76% of XDR MBL-producing K. pneumoniae were susceptible, while 15% and 9% were resistant and intermediate, respectively (Figure 2).
2.2. Detection of Resistance Markers
The double-disc synergy test was positive for KPC in 150/359 CRE; however, blaKPC was confirmed in 161/359 isolates. Among 161 positive blaKPC CRE, 20 co-harboured blaNDM. A total of 190 CRE displayed a positive double-disc synergy test for MBL, according to the detection of blaNDM, including those 20 co-producers (Table 1).
Twenty-eight CRE (negative for blaKPC and/or blaNDM) were positive for blaOXA-48-like; among these, 15/28 CRE were resistant to ertapenem but not to imipenem and/or meropenem. These CRE were positive for OXA-163 by LFIA (Table S1).
Moreover, blaCTX-M was the only ESBL coding gene detected by PCR among CRE (163/359), since blaPER-2 was not present in any of them. In turn, blaCMY was found in 48 CRE and blaCTX-M + blaCMY in 18. The co-carriage of blaKPC + blaCTX-M was detected in 54 CRE, blaKPC + blaCMY in 1, blaNDM + blaCTX-M in 75, blaNDM + blaCMY in 38, while blaKPC + blaNDM + blaCTX-M was detected in 7 CRE, and blaKPC + blaNDM + blaCTX-M + blaCMY in 3 (Table S1).
Among K. pneumoniae isolates, a high prevalence of blaKPC-2 with respect to blaKPC-3 was observed. Accordingly, specific peaks for KPC-2 and KPC-3 were detected by MALDI-TOF MS (87% corresponded to KPC-2 and 13% to KPC-3). blaNDM-5 was the most frequent variant, followed by blaNDM-1 (77% and 23%, respectively) (Table S1). Before the COVID-19 pandemic, KPC was the main carbapenemase in HB, while NDM was already the prevalent one in HA. In 2022, both enzymes showed similar prevalence (Figure 3).
2.3. Molecular Typing of K. pneumoniae Isolates
In HA, among the 77 K. pneumoniae isolates, 35 corresponded to the CG258-tonB79 cluster, 9 to the CG258-non-tonB79 cluster, 7 belonged to ST307, and 26 to other STs (Table S1). The emergence and widespread expansion of CG258-tonB79 was observed between 2019 and 2022; in parallel, an increase in blaKPC relative to blaNDM was detected. During 2022, the first KPC + NDM-producing isolate was recovered from this hospital (Figure 3).
In HB, among the 190 K. pneumoniae isolates, 75 corresponded to the CG258-tonB79 cluster, 37 to the CG258-non-tonB79 cluster, 27 belonged to ST307, and 51 to other STs (Table S1). The clusters CG258-tonB79 and ST307 were present in all periods except 2022, when ST307 was absent. Also, in 2022, CG258-tonB79 significantly increased its prevalence (fourfold) with respect to 2019 (p-value < 0.0001). CG258-non-tonB79 cluster displayed its greatest prevalence during 2020. Unlike HA, blaKPC was the prevalent carbapenemase persisting over the years in all clusters. Another difference with respect to HA is the presence of blaKPC + blaNDM co-harbouring isolates since 2019 in both CG258 clusters. In this hospital, during 2022, an increase in blaNDM detection was observed, associated with the expansion of CG258-tonB79 (Figure 3).
2.4. MLST of NDM-Producing K. pneumoniae Isolates
Out of 103 isolates subjected to WGS, all genome assemblies passed the quality control proposed by the pipeline, with no contamination markers detected. Coverage ranged from 28× to 809×, genome size from 5.3 Mb to 6 Mb and number of contigs from 35 to 206 (Table S1).
WGS confirmed ST258 as the unique ST in the prevalent CG258-tonB79 cluster and ST11 as the only lineage in the CG258-non-tonB79 cluster. In addition, WGS showed that isolates typed as ST307 by PCR actually corresponded to ST307 and also ST5994. Seventy-one out of 103 isolates corresponded to epidemic lineages: ST258 (n:41), ST11 (n:20), and ST307 (n:10). These results showed good correlation with PCR typing. The remaining 32 isolates corresponded to ST107 (n:6), ST1788 (n:4), ST1805 (n:3), ST207 (n:3), ST3430 (n:3), ST5994 (n:3), ST1229 (n:2), ST525 (n:2), ST15 (n:1), ST294 (n:1), ST45 (n:1), ST485 (n:1), ST6148 (n:1) and ST870 (n:1) (Table 2).
Among epidemic lineages, blaNDM-5 was confirmed as the prevalent allele being detected in 18, 16, and 9 isolates of ST258, ST11, and ST307, respectively. The second prevalent blaMBL was blaNDM-1, which was present in 15, 4, and 1 isolates of ST258, ST11, and ST307, respectively. Even blaNDM-5 was detected in 30 isolates belonging to different STs (Table 2).
3. Discussion
This study provides a dynamic view of the epidemiology, resistance profiles, and molecular characteristics of CRE recovered in two hospitals over four years spanning the COVID-19 pandemic. Across both hospitals and all study periods, K. pneumoniae was the predominant CRE in line with the well-established role of K. pneumoniae as the main driver of carbapenem resistance worldwide [3]. An increase in more extensive resistance phenotypes was observed; while MDR isolates predominated before the COVID-19 pandemic, the post-pandemic period was characterised by a striking increase in XDR isolates. Similarly, DTR phenotypes increased, particularly among K. pneumoniae, further limiting therapeutic options. This concomitant increase in XDR and DTR isolates indicates a progressive narrowing of therapeutic options, expanded to last-line agents. Overall, the year-by-year analysis revealed distinct resistance trajectories between hospitals. While HA showed relative stability in most NS rates, except for a marked increase in colistin resistance, HB exhibited a progressive and sustained rise in NS to multiple antimicrobial classes, suggesting heterogeneous local dynamics. The observed increase in XDR phenotypes may be related to the antimicrobial consumption. Consistent with the resistance patterns detected in CRE, penicillins and their derivatives were the most consumed antibiotic class from 2019 to 2022 in Argentina (46.5–51.6% annually). Although quinolones ranked third in 2020 and 2021 (10.9% and 11.3%, respectively), by 2022 they were surpassed by other β-lactams, including cephalosporins, monobactams, and carbapenems, while penicillins remained the most widely used class [23].
A significant rise in CRE was observed in HA when comparing the pre- and post-pandemic periods, consistent with reports from other regions describing the impact of the COVID-19 pandemic on antimicrobial resistance [24]. Notably, other regional studies have reported significant shifts in antimicrobial resistance profiles and antibiotic consumption during the COVID-19 pandemic compared with the pre-pandemic period [25,26,27]. The divergent resistance dynamics in HA with respect to HB underscore the importance of hospital-specific surveillance rather than relying on aggregated regional data.
NDM was detected across the different Enterobacterales species, with NDM-5 being the prevalent variant in K. pneumoniae. The second most common carbapenemase was KPC, mainly KPC-2, which was infrequent in species other than K. pneumoniae. This pattern is consistent with regional and global trends [4,24,28]. A temporal shift in carbapenemase prevalence was observed: while KPC predominated in HB before the pandemic and NDM was already prevalent in HA, both enzymes reached similar frequencies by 2022. This convergence suggests ongoing dissemination and replacement dynamics that may be driven by clonal expansion rather than sporadic horizontal gene transfer alone. The increase in NDM-producing isolates correlated with CZA resistance, and with aztreonam resistance when associated with CTX-M. Although CZA was introduced in Argentina as a targeted therapeutic option against KPC-producing Enterobacterales, its clinical implementation may have exerted selective pressure favouring the emergence and dissemination of NDM-producing CRE.
Double carbapenemase-producing isolates had been detected only once in our country before the studied period (2019), where KPC-3 + NDM-1-producing K. pneumoniae ST307 was detected in HB [14]. At the end of the COVID-19 pandemic, an increase in this resistance profile was observed in HB; however, it was due to the expansion of KPC-2 + NDM-5-producing K. pneumoniae ST258. All but one double carbapenemase producers were confined to CG258 (Table S1). The discrepancy between KPC detection based on phenotypic synergy tests with respect to LFIA and/or molecular methods could be explained by the presence of isolates co-harbouring blaKPC + blaNDM. In these co-producing isolates, inhibition of KPC by phenylboronic acid was not observed, likely due to the concurrent hydrolytic activity of NDM masking the KPC inhibition.
Since 2022, K. pneumoniae epidemiology has been driven by ST258, associated with the carriage of blaKPC-2 and/or blaNDM-5, in both HA and HB. Also, other epidemic/high-risk clones were confirmed, such as ST11 and ST307. As well as the ST15, ST45, and ST485, previously found in Argentina and others that have never been reported before in this country, were evidenced [10,29]. The detection of blaNDM-5 in a wide variety of STs highlights the successful dissemination of its genetic platform beyond classical high-risk clones [30].
Susceptibility testing of cefiderocol by broth microdilution requires iron-depleted Mueller–Hinton broth, whose multi-step preparation renders the method labour-intensive, difficult to standardise, and associated with reduced reproducibility, thereby limiting its routine implementation in clinical laboratories [31]. In addition, endpoint interpretation can be difficult and prone to error due to in vitro visual artefacts, as noted in the M100 manual of the Clinical and Laboratory Standards Institute (CLSI) [32], requiring further standardisation across successive M100 editions. Despite the marked increase in XDR K. pneumoniae isolates, cefiderocol retained activity against most XDR NDM-producing strains. However, resistant isolates were detected. Given that cefiderocol is not currently used in clinical practice in Argentina, this resistance may have arisen through indirect selective pressures. The high prevalence of NDM producers in our setting warrants particular attention, as NDM may act as a proxy for the emergence of cefiderocol resistance through the co-expression of additional resistance mechanisms. Moreover, overexpression of the blaNDM has been linked to the in vivo development of cefiderocol resistance during treatment [22,31]. These observations underscore the need for continuous surveillance of cefiderocol activity, especially in regions with endemic dissemination of NDM-producing Enterobacterales.
The limitations of this study include the following: Isolates from HA were unavailable during 2020, limiting direct comparison with the early COVID-19 pandemic period. Whole-genome sequencing was restricted to MBL-producing K. pneumoniae, while other isolates were characterised by PCR-based methods, and non-K. pneumoniae species were not molecularly typed. In addition, bioinformatic analyses did not detail outbreak or plasmid investigations. Despite these limitations, our results provide a valuable genomic baseline for future studies addressing the evolution and dissemination of high-risk clones in our region.
4. Materials and Methods
4.1. Bacterial Isolates, Antibiotic Susceptibility and Categorization
A prospective, observational, and multicentric study was carried out in two public hospitals (HA and HB) during May-September in 2019 (COVID-19 pre-pandemic), 2020–2021 (COVID-19 pandemic), and 2022 (COVID-19 post-pandemic). HA is a second-level hospital containing 200 beds, while HB is a third-level hospital containing 300 beds. Both hospitals experience substantial demand by socioeconomically vulnerable patients in densely populated peripheral areas of Buenos Aires metropolitan area, for whom the public healthcare system represents the main point of access to medical care.
In the hospitals, bacterial isolation from clinical specimens was carried out in accordance with the procedures outlined in the Manual of Clinical Microbiology of the American Society for Microbiology [33]. Antibiotic susceptibilities to aminopenicillins, cephalosporins, monobactams, carbapenems, ciprofloxacin, gentamicin, amikacin, tigecycline, and TMS were determined using automated methods (Phoenix, Bruker Daltonics, Bremen, Germany, in HA and Vitek2, bioMérieux, Craponne, France and BD in HB). CZA and colistin susceptibilities were determined by diffusion test and col-drop test, respectively. Interpretation was performed according to the Clinical and Laboratory Standards Institute (CLSI, https://clsi.org/) breakpoints applicable for each study year, except for ceftazidime–avibactam (CZA) and tigecycline, for which the European Committee on Antimicrobial Susceptibility Testing (EUCAST, https://www.eucast.org/) and U.S. Food and Drug Administration (FDA) criteria were applied [32,34,35,36]. All Enterobacterales isolates resistant to at least one carbapenem (meropenem, imipenem, and/or ertapenem) were delivered to IBaViM Institute in an encrypted way (to preserve the identity of the patients). HA did not conserve isolates to be delivered in 2020 due to health system overload. CRE identification was controlled at IBaViM by MALDI-TOF MS using a Microflex LT mass spectrometer (Bruker Daltonics, Bremen, Germany) with flexControl 3.4 software (Bruker Daltonics, Bremen, Germany), using the in situ extraction method, which consists of adding 1 μL of formic acid before sealing with the commercial hidroxicianocinamicacid (HCCA) matrix. All isolates were stored at −70 °C in brain heart infusion broth with 20% glycerol.
CRE isolates were classified as Multi-resistant (MDR), Extremely resistant (XDR), and Pan-resistant (PDR), according to Magiorakos et al., 2012 [37]. Intrinsic resistance was not addressed for MDR, XDR and PDR definitions. In accordance with the antimicrobial categories listed for Enterobacteriaceae, the MDR was assumed when an isolate was non-susceptible to at least 1 agent in ≥3 antimicrobial categories. XDR was assumed when the isolate was non-susceptible to at least 1 agent in all but 2 or fewer antimicrobial categories, and PDR was assumed for non-susceptibility to all agents in all antimicrobial categories for each isolate. Additionally, susceptibility to cefiderocol (FDC) was determined in 81 MBL-producing K. pneumoniae isolates, categorised as XDR according to Magiorakos et al.
In addition, the isolates were categorised as DTR pathogens, following the practical proposal of Kadri et al. [20] when resistance to all first-line agents was observed, such as resistance to all β-lactams, including carbapenems and β-lactamase inhibitor combinations, and fluoroquinolones.
4.2. Phenotypic Detection of Carbapenemases
The presence of carbapenemases was analysed by double disc inhibition test using both phenylboronic acid (PBA) (300 µg) and EDTA (1 µmol) for the detection of KPC and/or metallo-β-lactamase (MBL), as well as lateral flow immunoassay (LFIA) (RESIST-3 O.O.K K-SET, Britania, Argentina) to differentiate among KPC, OXA-48, and OXA-163. The direct detection of specific β-lactamases, such as KPC-2, KPC-3, KPC-8, and KPC-31, was carried out by analysing the protein spectrum obtained by MALDI-TOF MS, as described by Espinosa et al. in K. pneumoniae [38,39].
4.3. Genotypic Detection of β-lactamases Coding Genes
The carbapenemase-encoding genes blaKPC, blaOXA-48-like, blaNDM, blaIMP, and blaVIM were screened by multiplex PCR using total DNA as template obtained by the boiling method [40,41]. In addition, blaCTX-M, blaCMY, and blaPER-2 were screened by Multiplex-PCR using the primers CTX-MU-1, CTX-MU-2, PER-2-PLUS, PER-2-MINUS, CIT-MF and CIT-MR. In K. pneumoniae, larger amplicons of carbapenemase coding genes were obtained using the specific primers KPC-F and KPC-R for blaKPC, and NDM-F and NDM-R for blaNDM. (Table 3). Amplicons were sequenced at external facilities and analysed using the BLAST tool of the BLDB database (http://www.bldb.eu:4567/, accessed on 1 March 2025) [8].
4.4. Molecular Typing of K. pneumoniae Isolates
The two clusters of K. pneumoniae CG258: CG258-tonB79 (e.g., ST258, ST379, ST418, and ST512) and CG258-non-tonB79 (e.g., ST11, ST340, and ST437) were investigated by Multiplex-PCR proposed by Yu et al. (Table 3) [42]. In parallel, isolates belonging to ST307 were characterised by simplex PCR following Lowe et al. (Table 3) [43].
Additionally, Multi Locus Sequence Typing (MLST) was assessed in 103 NDM-producing K. pneumoniae genomes using the BIGSdb-Pasteur database (https://bigsdb.pasteur.fr/klebsiella/, accessed on 1 October 2025) [48,49]. Genomic DNA was extracted using a commercial kit (DNA Puriprep B-Kit, Inbio Highway, Argentina) and sequenced (WGS) by Illumina Miseq. Raw reads were processed through a standardised bioinformatics pipeline (NIHR Global Health Research Unit De novo assembly pipeline). Adapter sequences and low-quality bases were removed using Trimmomatic (v0.38), followed by read error correction with Lighter (v1.1.1). To normalise sequencing depth, reads were downsampled to 100× coverage using SeqTK (v1.3), and overlapping paired-end reads were merged with FLASH (v1.2.11). De novo genome assembly was performed using SPAdes (v3.12.0). Quality control of raw and processed data was assessed using FastQC (v0.11.8), MultiQC (v1.7), and Qualifyr (v1.4.4). Species identification was conducted with Bactinspector (v0.1.3), and potential sample contamination was evaluated using Confindr (v0.7.2) [50].
4.5. Statistical Analysis
Categorical variables, including non-susceptibility (NS) rates, resistance phenotypes (MDR/XDR/DTR), prevalence of resistance genes, and clonal distribution across hospitals and study periods, were compared using Fisher’s exact test. This test was selected due to our sample sizes. A p-value < 0.05 was considered statistically significant. The R software (version 4.5.1) has been used to analyse and graph the data [51].
5. Conclusions
This study evidences the increase in more extensive resistance phenotypes among CRE throughout the COVID-19 pandemic, driven by carbapenemase-producing K. pneumoniae, accompanied by a shift toward XDR and DTR phenotypes. Resistance dynamics differed between centres: while HA remained relatively stable—except for a rise in colistin resistance—HB showed a sustained increase in non-susceptibility across multiple antimicrobials. This trend may reflect local antimicrobial consumption patterns, which could contributed to the selection of more resistant isolates in Argentina during the study period.
NDM-5 and KPC-2 were the main carbapenemases identified, in line with regional and global trends. A temporal shift in carbapenemase prevalence was observed in each hospital, converging in similar frequencies of KPC and NDM by 2022 across both centres. This scenario was driven by the active dissemination of K. pneumoniae ST258.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Paudel R. Shrestha E. Chapagain B. Tiwari B.R. Carbapenemase Producing Gram Negative Bacteria: Review of Resistance and Detection Methods Diagn. Microbiol. Infect. Dis.202411011637010.1016/j.diagmicrobio.2024.11637038924837 · doi ↗ · pubmed ↗
- 2Alvisi G. Curtoni A. Fonnesu R. Piazza A. Signoretto C. Piccinini G. Sassera D. Gaibani P. Epidemiology and Genetic Traits of Carbapenemase-Producing Enterobacterales: A Global Threat to Human Health Antibiotics 20251414110.3390/antibiotics 1402014140001385 PMC 11852015 · doi ↗ · pubmed ↗
- 3World Health Organization WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance 2024 Available online: https://www.who.int/publications/i/item/9789240093461(accessed on 23 February 2026)
- 4Pillonetto M. Wink P.L. Melano R.G. Jiménez-Pearson M.A. Melgarejo Touchet N.L. Saavedra Rojas S.Y. Kulek D.N.O. Abreu A.L. Peral R.T. Miorando R. Carbapenemases Producing Gram-Negative Bacteria Surveillance in Latin America and the Caribbean: A Retrospective Observational Study from 2015 to 2020 Lancet Reg. Health—Am.20254910118510.1016/j.lana.2025.10118540697529 PMC 12281157 · doi ↗ · pubmed ↗
- 5Bush K. Jacoby G.A. Updated Functional Classification of β-Lactamases Antimicrob. Agents Chemother.20105496997610.1128/AAC.01009-0919995920 PMC 2825993 · doi ↗ · pubmed ↗
- 6Bush K. Bradford P.A. Epidemiology of b-Lactamase-Producing Pathogens Clin. Microbiol. Rev.202033 e 00047-1910.1128/CMR.00047-1932102899 PMC 7048014 · doi ↗ · pubmed ↗
- 7Castanheira M. Deshpande L.M. Mendes R.E. Canton R. Sader H.S. Jones R.N. Variations in the Occurrence of Resistance Phenotypes and Carbapenemase Genes Among Enterobacteriaceae Isolates in 20 Years of the SENTRY Antimicrobial Surveillance Program Open Forum Infect. Dis.20196 S 23S 3310.1093/ofid/ofy 34730895212 PMC 6419900 · doi ↗ · pubmed ↗
- 8Naas T. Oueslati S. Bonnin R.A. Dabos M.L. Zavala A. Dortet L. Retailleau P. Iorga B.I. Beta-Lactamase Database (BLDB)—Structure and Function J. Enzym. Inhib. Med. Chem.20173291791910.1080/14756366.2017.1344235 PMC 644532828719998 · doi ↗ · pubmed ↗