Genome-Wide Identification and Functional Analysis of the CNGC Gene Family in Suaeda glauca
Jun Wang, Chunxing Dong, Xiaoxue He, Dongpeng Zheng, Xingguang Chen, Jiahao Cai, Gang Wang, Boping Tang, Chunyin Zhang, Lulu Wang, Xiaoping Niu, Chunmei Lai, Yuan Qin, Yan Cheng

TL;DR
This study identifies and analyzes 44 CNGC genes in Suaeda glauca, revealing one gene that may reduce salt tolerance, offering insights for improving crops in saline soils.
Contribution
The study provides a genome-wide analysis of the CNGC gene family in Suaeda glauca and identifies a gene with a negative regulatory role in salt tolerance.
Findings
44 CNGC genes were identified in Suaeda glauca, showing structural and functional divergence.
Overexpression of SgCNGC13 in Arabidopsis increased salt sensitivity, indicating a negative regulatory role.
The study reveals evolutionary conservation and functional diversity of the CNGC gene family in S. glauca.
Abstract
Soil salinization is a growing problem that is harming agriculture and the environment. Suaeda glauca is a plant that grows in saline–alkali land, but the genetic regulatory mechanisms underlying its salt tolerance are not fully understood. This study explores a family of genes, Cyclic nucleotide-gated channel genes, that play key roles in helping plants cope with stress. A total of 44 members of this gene family were identified in Suaeda glauca, and their characteristics were analyzed. We found that when the expression of one gene was significantly increased in Suaeda glauca, the plant became more sensitive to salt, suggesting that this gene may negatively regulate salt tolerance, which provides a basis for understanding how the plant survives in high-salt environments. In the future, these findings may help scientists develop new crop varieties that are better suited to growing in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
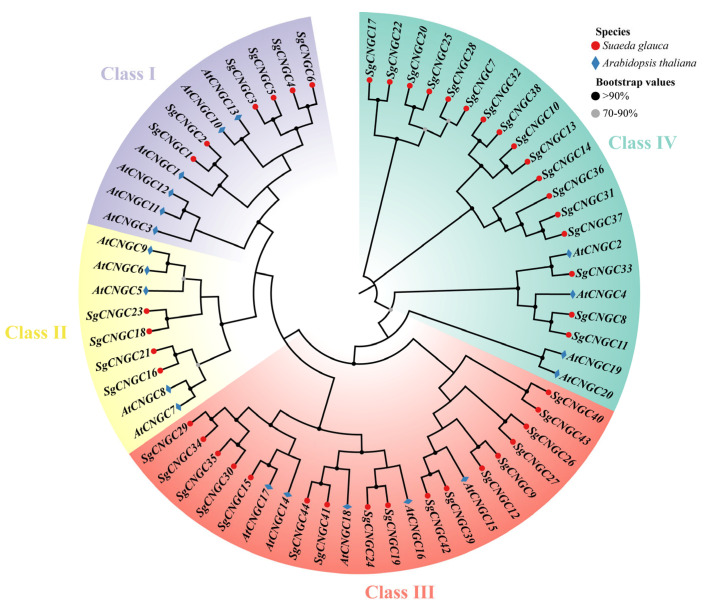 Figure 1
Figure 1 Figure 2
Figure 2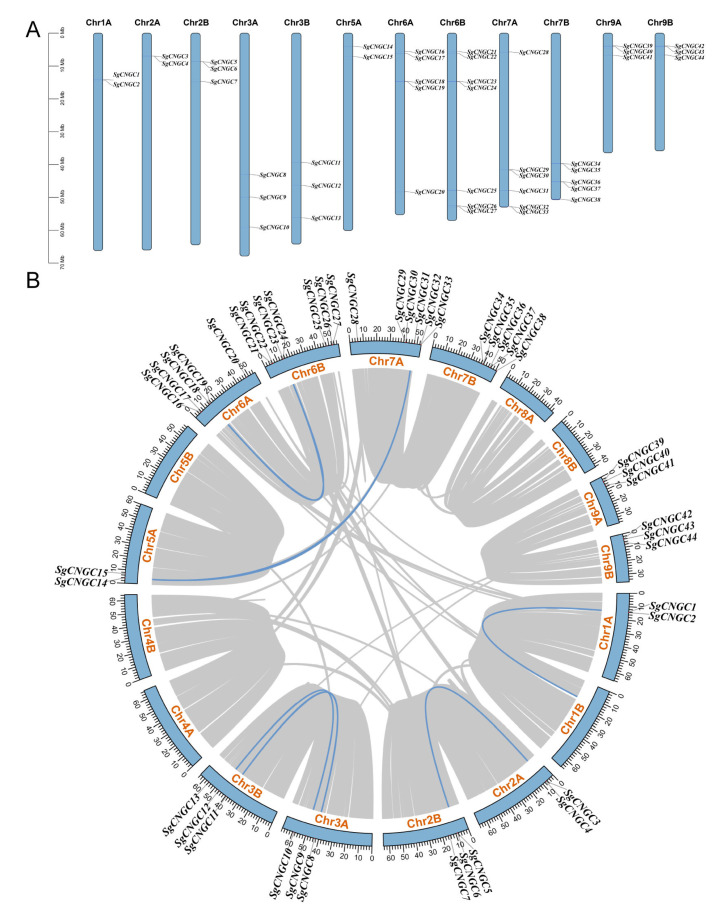 Figure 3
Figure 3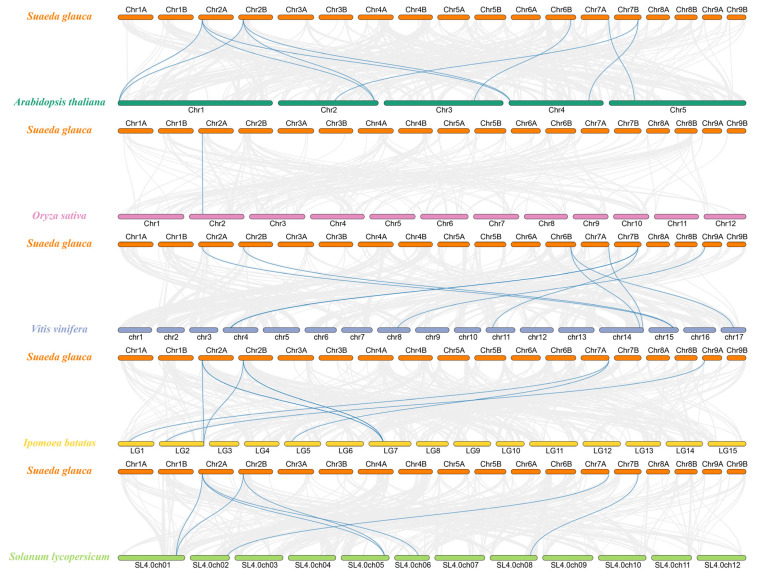 Figure 4
Figure 4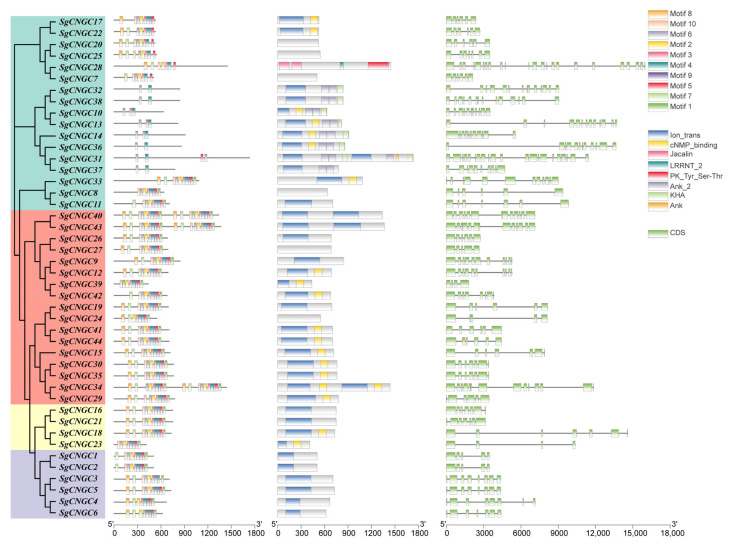 Figure 5
Figure 5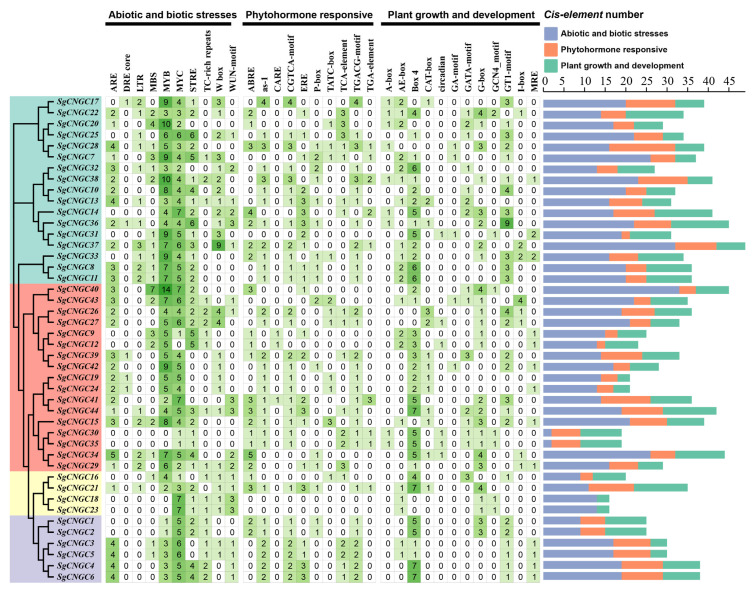 Figure 6
Figure 6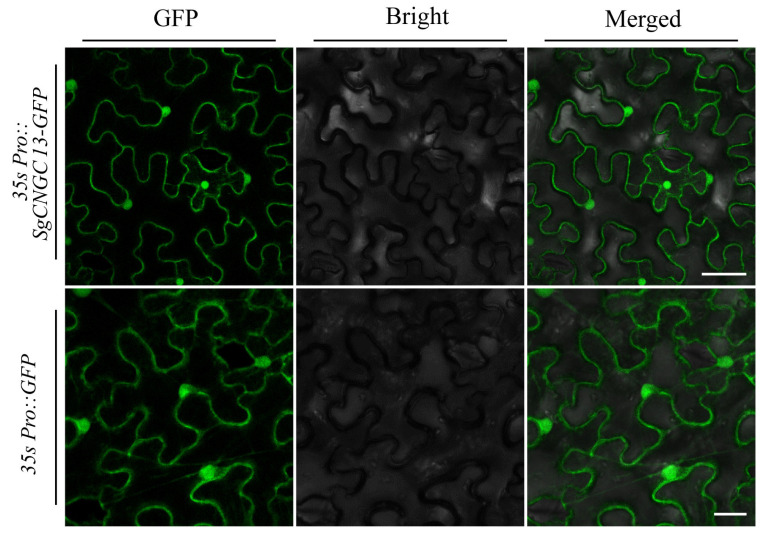 Figure 7
Figure 7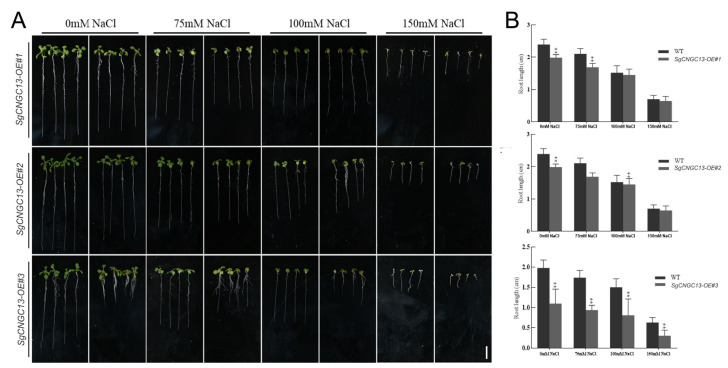 Figure 8
Figure 8- —Natural Science Foundation of Fujian Province
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Stress Responses and Tolerance · Plant and Biological Electrophysiology Studies
1. Introduction
The salinized land area in China accounts for 10.1% of the global total [1], making it the country with the largest area of soil salinization [2]. As potential reserve arable land resources, these lands hold significant development potential. Soil salinization is primarily caused by natural factors (e.g., evaporation exceeding precipitation in arid regions, seawater intrusion in coastal areas) and human activities (e.g., improper irrigation, excessive fertilizer application), often leading to soil compaction, decreased fertility, and ecological degradation. It has become a key constraint on the sustainable development of agriculture in China. Suaeda glauca (S. glauca) is an annual herb belonging to the family Chenopodiaceae and genus Suaeda. Morphologically, its succulent leaves and stems contribute to reduced transpiration and enhanced water storage capacity. Additionally, its cells exhibit strong osmotic potential, enabling tolerance to high-salt environments. It is widely distributed in coastal areas of China, playing an important role in the ecological restoration of saline–alkali land and the maintenance of biodiversity [3].
Cyclic nucleotide-gated channel (CNGC) genes are a type of calcium ion transport channels, generally composed of six transmembrane (TM) domains, calmodulin binding domains (CaMBDs), and cyclic nucleotide-binding domains (CNBDs). Among them, the CNBD domain is highly conserved and can specifically recognize and bind to cyclic nucleotide molecules [4]. Ca^2+^, as an important second messenger, is widely involved in regulating plant growth, development, and responses to abiotic stresses (such as osmotic and ionic stress) [5]. CNGCs are widely present in the plant kingdom (e.g., rapeseed, maize, tomato, potato) and function as calcium ion channels in plants, participating in signal transduction of various physiological processes by modulating calcium ion concentration. The number of CNGC genes and their distribution across clades vary among different plants, and their functions also differ. For example, Chen et al. identified 49 BnCNGC genes in Brassica napus, among which BnCNGC9 from Group IVa and BnCNGC27 and BnCNGC48 from Group IVb exhibited different expression patterns under various stress conditions [6].
In recent years, numerous studies have reported on gene families in Suaeda and the CNGC gene family in plants. Wang et al. [7] found that AGO proteins play an important role in salt stress response in Suaeda salsa. Furthermore, Baloch [8] and Zhang et al. [9] reported significant associations of this gene family with salt stress and drought tolerance in alfalfa and transgenic A. thaliana, respectively. However, there have been no reports on the genome-wide identification and systematic analysis of the CNGC gene family in S. glauca to date. This study utilized bioinformatics approaches to analyze the members, physicochemical properties, subcellular localization, and phylogenetic relationships of the CNGC gene family in S. glauca, aiming to provide a theoretical basis for further investigation into the functions of S. glauca gene families, screening of stress-resistant varieties, and promoting its application in saline–alkali land.
2. Materials and Methods
2.1. Experimental Materials
The S. glauca cultivar used in this study were provided by Yancheng Lvyuan Saline Soil Agriculture Technology Co., Ltd., Yancheng, Jiangsu, Southeast China (http://www.ychpz.com/, accessed on 15 September 2025). The cultivation of S. glauca followed that of Mohammadi et al. [10]. Nicotiana benthamiana (N. benthamiana) seeds were sourced from the laboratory seed bank and planted in a mixed substrate (vermiculite: nutrient soil = 1:1) at 28 °C. Arabidopsis thaliana Col-0 plants obtained from ABRC (Arabidopsis Biological Resources Center, https://abrc.osu.edu/, accessed on 15 September 2025) were used as the wild type and cultured on 1/2 MS medium at 25 °C.
2.2. Analysis of Protein Physicochemical Properties
Twenty CNGC protein sequences from Arabidopsis thaliana were retrieved from the Arabidopsis Information Resource (TAIR10) database (http://www.arabidopsis.org/, accessed on 20 September 2025). The genome of S. glauca was obtained from China National Center for Bioinformation (https://www.cncb.ac.cn/, accessed on 20 September 2025) under the accession number WGS038631 [11]. Using this information, BLASTp alignment was performed, combined with domain prediction using the SMART online tool (http://smart.embl-heidelberg.de/, accessed on 22 September 2025) [12]. Based on the presence of TM domains, CaMBD, and CNBD structures and manually removing redundant sequences, CNGC genes in S. glauca were identified. The online tool ExPASy ProtParam (https://web.expasy.org/protparam/, accessed on 22 September 2025) was used to analyze physicochemical properties such as the isoelectric point, number of amino acids, and molecular weight of the CNGC family members. The subcellular localization of SgCNGC proteins was predicted using the online software CELLO (https://cello.life.nctu.edu.tw/, accessed on 22 September 2025) [13].
2.3. Phylogenetic, Conserved Domain, and Sequence Alignment Analysis of Gene Families
Phylogenetic analysis of CNGC gene family members from Arabidopsis thaliana and S. glauca was performed using MEGA11.0 (https://www.megasoftware.net/, accessed on 22 September 2025). The maximum likelihood (ML) method [14] was employed with the JTT+G (Jones-Taylor-Thornton + Gamma distribution) model, and bootstrap support values were calculated from 1000 replicates. The Evolview online platform (https://www.evolgenius.info/evolview/, accessed on 25 September 2025) was used for further visualization and refinement of the phylogenetic tree [15]. The CNGC protein sequences from A. thaliana and S. glauca were imported into MEGA11.0 software for sequence alignment using the Muscle algorithm [16] to identify conserved domains, and the results were exported in FASTA format. Jalview 2 software was used for final visualization [17].
2.4. Chromosomal Localization and Collinearity Analysis of the Gene Family
TBtools software (v2.315) was used to analyze chromosomal localization of SgCNGC genes and to generate chromosomal distribution maps. Protein-coding genes for A. thaliana, Oryza sativa (O. sativa), Vitis vinifera (V. vinifera), Ipomoea batatas (I. batatas), and Solanum lycopersicum (S. lycopersicum) were downloaded from the public database EnsemblPlants (http://plants.ensembl.org/index.html, accessed on 25 September 2025). The multiple collinearity scan tool MCScanX was used to analyze the collinearity of CNGC genes between the S. glauca genome sequence and other species [18]. TBtools software was used for visualization, constructing intra-species collinearity circos plots and inter-species collinearity maps.
2.5. Analysis of Gene Structure and Cis-Acting Elements of Gene Family
The protein amino acid sequences of SgCNGC proteins were uploaded to MEME (https://meme-suite.org/, accessed on 26 September 2025) for motif analysis. The number of motifs was set to 10, with other parameters as default [19]. TBtools software was used for motif visualization. The upstream 2000 bp sequences of the transcription start site of SgCNGC genes extracted by TBtools were considered as the putative promoter region and were used to predict upstream cis-regulatory elements with the PlantCARE database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 26 September 2025) [20], and the results were visualized using TBtools software.
2.6. Protein Subcellular Localization
The cDNA samples of S. glauca were use as a template for amplifying the coding DNA sequence (CDS) sequence of SgCNGC13 [21]. The pCAMBIA2300-SgCNGC13-GFP fusion expression vector was constructed by homologous recombination. The resulting constructs were transiently expressed in 4-week-old N. benthamiana leaves via Agrobacterium-mediated transformation. After 24 h of transformation, the N. benthamiana leaf disks were observed under a Leica TCS SP8X confocal microscope [22].
2.7. Analysis of Transgenic Overexpression Lines
Base deletions and insertions were occurred in the sequence alignment of SgCNGC13, suggesting that its function might be altered. Therefore, it was selected for further functional verification. The CDS of SgCNGC13 was cloned into the pGWB505 binary vector using double digestion (XhoI and SalI) and the ligation cloning method. A. thaliana transformation was performed via the Agrobacterium-mediated floral dip method [23]. Transgenic seeds were screened on 1/2 MS medium (pH 5.8) containing 50 mg L^−1^ hygromycin. The T3 homozygous transgenic lines with high SgCNGC13 expression levels and wild-type A. thaliana seeds were sterilized and germinated on 1/2 MS plates supplemented with 0, 75, 100, or 150 mM NaCl, and cultured at 25 °C under a 16/8 h light/dark cycle. Root lengths were observed and measured after 2 weeks to evaluate their salt tolerance potential. For statistical analysis, root length measurements were collected from at least 15 seedlings per genotype per treatment, with three independent biological replicates. Data were expressed as mean ± standard deviation (SD). Statistical analyses were performed using GraphPad Prism software 10.1.2.
3. Results
3.1. Genome-Wide Identification and Physicochemical Property Analysis of CNGC Gene Family in S. glauca
44 non-redundant genes encoding CNGC proteins were identified from the S. glauca genome (Supplementary Table S1). Based on their chromosomal distribution [24] and the similarity to A. thaliana homologues, the CNCG genes in S. glauca were renamed SgCNGC1 to SgCNGC44. The encoded proteins of the SgCNGCs range in length from 411 to 1734 aa, with molecular weights from 47.60 to 194.87 kDa and aliphatic index from 80.72 to 107.66. Subcellular localization prediction indicated that the plasma membrane is the most consistently predicted location for most members.
3.2. Phylogenetic Analysis Showed a Close Genetic Relationship and Similarity Between S. glauca and A. thaliana CNGC Genes
The maximum likelihood phylogenetic tree clearly divided the 44 SgCNGC genes into four subgroups (Class I, II, III, IV), with a relatively even distribution of members across branches and no aggregation of identical protein genes (Figure 1). Bootstrap support values for most nodes were above 90%, with some between 70 and 90%, indicating a reliable topological structure of the phylogenetic tree. Phylogenetic analysis indicated a close genetic relationship and similarity between S. glauca and A. thaliana CNGC genes.
3.3. Analysis of Conserved Domains and Sequence Alignment of the S. glauca CNGC Gene Family
Sequence alignment results (Figure 2) show that all members of the S. glauca CNGC gene family possess CaMBD and CNBD domains at the carboxyl terminus (C-terminus) and transmembrane domains at the amino terminus (N-terminus), which are characteristic of typical plant CNGC gene families [25]. Motif analysis identified conserved motifs specific to the S. glauca CNGC genes in the CNBD, with glycine (G) and leucine (L) appearing most frequently as the most conserved residues. These residues are crucial in maintaining structural stability, cyclic nucleotide binding, and ion channel regulation in CNGC proteins. Notably, SgCNGC10, SgCNGC13, SgCNGC14, SgCNGC20, SgCNGC31, SgCNGC32, SgCNGC36, SgCNGC37, and SgCNGC38, located in subgroup IV, exhibited a high number of non-conserved residues in the CNBD domain. SgCNGC10 and SgCNGC13 contained unique insertion sequences, while SgCNGC13 also exhibited a deletion of only 48 amino acid residues. Overall, members of gene subgroups Class I and II showed extremely high sequence similarity in the CNBD region, with better conservation than Class III and Class IV. The conserved and variable patterns of S. glauca CNGC gene sequences were highly consistent with the classification in the phylogenetic tree (Figure 1).
3.4. Chromosomal Localization and Collinearity Comparative Analysis of the S. glauca CNGC Gene Family
The 44 SgCNGC genes were unevenly distributed across 12 chromosomes (Figure 3). Chromosomes 2B, 3A, 3B, 9A, and 9B each harbored three SgCNGC genes, whereas the number of genes on other chromosomes ranged from 2 to 7. Chromosome 6B contained the most genes, forming a distinct gene cluster. Based on conserved regions sequence comparisons, highly similar gene sequences and tandem duplications were observed on chromosomes 2A, 2B, 6A, 7A, and 7B. Although complete gene duplication was not observed on other chromosomes, the sequences were still highly similar. Intra-species collinearity analysis of the S. glauca CNGC gene family revealed large-scale gene duplication events, identifying six collinear gene pairs. Among them, SgCNGC23 & SgCNGC18, SgCNGC31 & SgCNGC14, SgCNGC4 & SgCNGC6, SgCNGC8 & SgCNGC11, and SgCNGC9 & SgCNGC12 belonged to the same subgroup, confirming closer evolutionary relationships among members of the same subgroup.
We compared S. glauca with both dicots and a monocot species to understand the conservation and divergence of CNGC genes during evolution. This multispecies comparison helps identify which lineages share close syntenic relationships with S. glauca. Collinearity analysis between S. glauca and five other species (A. thaliana, O. sativa, V. vinifera, I. batatas, and S. lycopersicum) yielded 10, 1, 8, 7, and 7 collinear gene pairs, respectively, with the highest number found with A. thaliana and the lowest in O. sativa (Figure 4). All dicot species formed collinear gene pairs with genes located on S. glauca chromosomes 2A, 2B, 7A, and 7B. These results indicate that A. thaliana is the most closely related model species to S. glauca among those analyzed.
3.5. Gene Structure Analysis of the S. glauca CNGC Gene Family
A total of 10 conserved protein motifs (Motif1–Motif10) were predicted among the 44 SgCNGC genes (Figure 5). Motif4 and Motif6 appeared most frequently. All SgCNGC genes contained Motif4, and except for SgCNGC4, SgCNGC6, SgCNGC24, and SgCNGC28, all gene possessed Motif6, indicating high conservation. Genes in subgroup Class IV contained fewer conserved motifs, suggesting that they may be more prone to mutation. Gene structure prediction identified eight conserved domains, with ion transport domains and CNBD domains being the most abundant. Additionally, CDS regions were detected in each gene. The number and length of exons and introns varied among subgroups, with members of the same subgroup showing similar exon–intron structures.
3.6. Cis-Acting Element Analysis Indicated That the SgCNGC Genes Might Be Involved in Drought and Salt Stress Response
Analysis of the upstream promoter region of the SgCNGC gene (Figure 6). revealed three major categories of cis-acting elements, related to abiotic stress (e.g., drought, salt, cold, heat) and biotic stress (e.g., pathogen infection) responses, phytohormone responses, and plant growth and development, corresponding to 10, 10, and 12 elements, respectively. These included elements responsive to low temperature, auxin, abscisic acid, gibberellin, light, etc. Members of the same subgroup had similar cis-acting elements. Among elements related to abiotic and biotic stress, MYC and MYB were the most common, followed by STRE and ARE. Feng et al. [26] previously identified a significant role of the CNGC gene family in regulating methyl jasmonate (MeJA), salt stress, and drought stress in tomato. Furthermore, MYB and MYC, in cooperation with other transcription factors, can participate in plant salt and drought stress responses [27], implying that SgCNGC genes may also be involved in drought and salt stress responses, enhancing salt and drought tolerance.
3.7. Subcellular Localization Analysis of SgCNGC13
Previous studies [14], have shown that plant CNGC genes are often localized to the cell membrane but also distributed in organelles such as the nucleus, cytoplasm, and chloroplasts. To further investigate the subcellular localization of SgCNGC genes, transient expression in N. benthamiana was performed to determine the localization of the SgCNGC13–green fluorescent protein (GFP) fusion construct, using an empty vector as a control. In the experimental group, green fluorescence signal was clearly localized to the cell periphery, highly consistent with the morphology of the cell membrane, and significant fluorescence signal was also observed in the nucleus. Overlay images of fluorescence and bright-field microscopy showed complete overlap of fluorescence signals at the cell membrane and nuclear positions with cellular structures (Figure 7).
3.8. Effect of SgCNGC13 Overexpression on Root Length in Transgenic A. thaliana
To investigate the salt tolerance mechanisms of the S. glauca CNGC gene family, this study compared the growth of SgCNGC13-overexpressing transgenic A. thaliana with wild-type A. thaliana under different salt concentrations. Increasing salt concentrations progressively inhibited shoot and root elongation. When overexpression lines were compared with wild-type plants, no significant difference in shoot growth rate was observed. Under normal, non-stress conditions (0 mM NaCl), the average root lengths of three overexpression lines were significantly shorter than those of the wild type. These results indicates that overexpression of SgCNGC13 affected normal plant growth and development, increasing root sensitivity. However, as salt concentration increased, the difference in root length between the two groups gradually decreased, suggesting that SgCNGC13 overexpression may negatively regulate the plant’s salt stress response (Figure 8).
4. Discussion
Globally, over 1.1 billion hectares of land are affected by salinization, with crop yield reductions reaching up to 30% in certain regions [28]. Approaches such as breeding salt-tolerant crops, developing saline agriculture, and exploring the economic value of halophytes are important strategic measures for achieving sustainable development and ensuring national food and ecological security. S. glauca possesses unique salt-tolerant properties and serves as a “pioneer” plant for ameliorating saline–alkali soil, playing a key role in the remediation of coastal and inland saline–alkali lands in China. Therefore, research on the response of S. glauca genes to abiotic stresses is crucial.
This study identified 44 SgCNGC gene family members in S. glauca using bioinformatics methods. Chromosomal localization revealed their presence on only 12 out of 18 chromosomes, with an uneven distribution. The gene number is quite different from that of A. thaliana (20 AtCNGC genes) but closest to that of apple (44 MdCNGC genes) [29] and Brassica napus (49 BnCNGC genes), suggesting diverse roles in stress response and similarity to apple and B. napus. Subcellular localization prediction placed SgCNGC proteins on the cell membrane, indicating that they function as ion channel proteins, similar to the localization pattern of A. thaliana CNGCs [30]. Concurrently, phylogenetic analysis showed similar classification and distribution of CNGC gene family members between S. glauca and A. thaliana, suggesting a close evolutionary relationship and homology between the two species. Further chromosomal and collinearity analysis detected tightly clustered gene distributions at chromosome ends, representing tandem duplications. This provides a driving force for gene family expansion, helping plants adaptively evolve (e.g., resisting pathogen infection, improving stress tolerance) [31], and is one of the major means of gene amplification (tandem duplication, segmental duplication, or whole-genome duplication). However, gene duplication can also result in the loss of less important duplicate genes following polyploidization [32]. Collinearity analysis results indicated close genetic relationships between S. glauca and A. thaliana, V. vinifera, I. batatas, and S. lycopersicum, consistent the phylogenetic tree analysis. Fewer collinear pairs with O. sativa suggest that S. glauca shares homology with these species, but due to the dicot–monocot division, its relationship with O. sativa is more distant than with other dicots [33]. These findings indicate gene differentiation during plant evolution, leading to variations in gene number and structural characteristics among species. Species with similar gene numbers and collinearity patterns may also share functional similarities.
Analysis of conserved domains and sequence alignment in the S. glauca CNGC gene family revealed that subgroups I, II, and III were more conserved than group IV. This finding is consistent with the conclusion of Maser et al. [34], which reported that the CNGC genes in subgroup IV exhibit a more distant evolutionary relationship. Insertions and deletions of non-conserved residues were observed in subgroup IV, especially in SgCNGC13. These structural differences may directly affect the ability of proteins to bind to cyclic nucleotides, interact with calmodulins, or regulate ion fluxes, potentially leading to alterations in the stress signaling pathway. Furthermore, the high conservation of Motif4 and Motif6, along with the universal presence of ion transport and CNBD domains, confirms their essential role in channel function. The coding region structure changes with gene differentiation. Genes within the same subgroup share certain similarities in exon–intron structure (number and length of introns and exons), which influences gene expression and protein function, further validating functional similarity within subgroups [35]. Analysis of cis-acting elements provides substantial evidence supporting the potential regulatory roles of the genes. Cis-acting elements in the S. glauca CNGC gene family participate in regulating functional domains related to plant responses to abiotic and biotic stresses, phytohormone responses, and growth and development. The widespread presence of MYB and MYC binding sites directly links this gene family to the regulatory transcription that controls salt and drought stress responses.
Among the identified S. glauca CNGC genes, SgCNGC13 exhibited lower conservation, possibly indicating mutation and loss of normal function. This study conducted subcellular localization of SgCNGC13 and performed overexpression analysis in A. thaliana, aiming to further explore its gene function and provide insights for modifying defective genes. SgCNGC13 was predicted to localize to the cell membrane but was found on both the cell membrane and nucleus, suggesting it primarily functions as a membrane ion channel protein while also potentially participating in nucleus-specific signaling pathways within the nucleus. Overexpression experiments suggested that SgCNGC13 may negatively regulate salt stress responses, providing a basis for further functional analysis.
In summary, this study provides a preliminary identification and analysis of the S. glauca CNGC gene family, laying a theoretical foundation for future functional investigations. It also provides valuable date for subsequent analysis of salt and drought tolerance mechanisms in S. glauca and for the screening of stress-resistant plants. However, the specific regulatory mechanisms underlying optimized salt and drought tolerance genes in S. glauca require further exploration. Future research should focus on elucidating these mechanisms to provide genetic resources and technical support for promoting saline-alkali land remediation and sustainable utilization.
5. Conclusions
Based on the whole-genome data of S. glauca, this study systematically identified 44 members of the SgCNGC gene family. Through multi-dimensional bioinformatics analysis, the structural characteristics, evolutionary relationships, and potential functional differentiation mechanisms of these members were characterized. The results indicate a close evolutionary relationship between S. glauca and A. thaliana. Furthermore, subgroups III and IV exhibited lower conservation and more distant relationships compared to other CNGC gene members, suggesting potential functional differences. These genes are likely involved in salt and drought stress responses. Overexpression of SgCNGC13 in A. thaliana resulted in enhanced sensitivity to salt stress, suggesting that SgCNGC13 gene may play a negative regulatory role in salt stress responses. This study provides a valuable resource for further exploration of S. glauca CNGC gene functions and stress resistance breeding. However, this study has certain limitations that cannot be ignored. At present, the identification and characterization of the SgCNGC gene largely rely on bioinformatics prediction, and the specific regulatory mechanism of the SgCNGC gene underlying their functions require further experimental validation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lei S. Jia X. Zhao C. Shao M. A review of saline-alkali soil improvements in China: Efforts and their impacts on soil properties Agric. Water Manag.202531710961710.1016/j.agwat.2025.109617 · doi ↗
- 2Hassani A. Azapagic A. Shokri N. Predicting long-term dynamics of soil salinity and sodicity on a global scale Proc. Natl. Acad. Sci. USA 2020117330173302710.1073/pnas.201377111733318212 PMC 7776813 · doi ↗ · pubmed ↗
- 3Wang S. Zhao Z. Ge S. Zhang K. Tian C. Mai W. The Effects of Suaeda salsa/Zea mays L. Intercropping on Plant Growth and Soil Chemical Characteristics in Saline Soil Agriculture 20221210710.3390/agriculture 12010107 · doi ↗
- 4Tan J. Xuan X. Su S. Guo H. Jiao Y. Genome-wide identification, characterization and expression analysis of CNGC Genes in moso bamboo (Phyllostachys edulis)BMC Plant Biol.20252583310.1186/s 12870-025-06808-z 40604466 PMC 12220554 · doi ↗ · pubmed ↗
- 5Köster P. De Falco T.A. Zipfel C. Ca 2+ signals in plant immunity EMBO J.202241 e 11074110.15252/embj.202211074135560235 PMC 9194748 · doi ↗ · pubmed ↗
- 6Chen S.Y. Ding Y.J. Sun J.M. Huang D.W. Yang N. Dai Y.H. Wan H.F. Qian W. Genome-wide identification of Bn CNGC and the gene expression analysis in Brassica napus challenged with Sclerotinia sclerotiorum and PEG-simulated drought Acta Agron. Sin.20224813571371(In Chinese)10.3724/SP.J.1006.2022.14091 · doi ↗
- 7Wang W. Fan S.L. Li J. Gao W. Jin Y.X. Zheng M.G. Zheng L. Isolation and expression analysis of argonaute genes from Suaeda salsa Adv. Mar. Sci.201937306314(In Chinese)
- 8Baloch A.A. Kakar K.U. Rais S. Nawaz Z. Almoneafy A.A. Raza A.M. Khan S. Ullah R. Genome-wide analysis of CNGC gene family in Brassica juncea (L.) Czern reveals key targets for stress resistance and crop improvement Plant Gene 20244110048710.1016/j.plgene.2024.100487 · doi ↗