Nitric Oxide Donor Spermine-NONOate Elicits Endogenous Dispersal-Associated Transcriptional Responses to Promote Biofilm Dispersal in Pseudomonas aeruginosa
Xavier Bertran Forga, Kathryn E. Fairfull-Smith, Jilong Qin, Makrina Totsika

TL;DR
This study shows that the NO donor Spermine-NONOate promotes biofilm dispersal in Pseudomonas aeruginosa by activating key dispersal-related genes.
Contribution
The study identifies specific transcriptional responses linked to NO-induced biofilm dispersal and validates known dispersal biomarkers.
Findings
SP-NONO activates canonical NO-regulated responses and disrupts energy metabolism pathways.
Shared transcriptional shifts in SP-NONO-treated and spontaneously dispersed biofilms include branched-chain amino acid catabolism genes.
Nine of 14 known dispersal biomarkers were confirmed in SP-NONO-treated biofilms.
Abstract
Background/Objectives: Bacterial biofilms are structured communities of sessile cells embedded in a self-produced extracellular matrix. Within biofilms, bacteria become highly tolerant toenvironmental stressors such as host immune responses and antimicrobial treatments. In response to specific cues, however, biofilm cells can revert to a planktonic free-swimming lifestyle through a process termed biofilm dispersal. When dispersed cells escape the biofilm matrix, they lose biofilm-associated antibiotic tolerance, a major barrier to treating medical biofilms. As such, dispersal-inducing compounds like nitric oxide (NO) are actively investigated as adjuvants to potentiate the biofilm-eradicating activity of existing antibiotics. We recently characterised the transcriptomic responses elicited during spontaneous biofilm dispersal in closed culture-grown Pseudomonas aeruginosa biofilms. Here,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
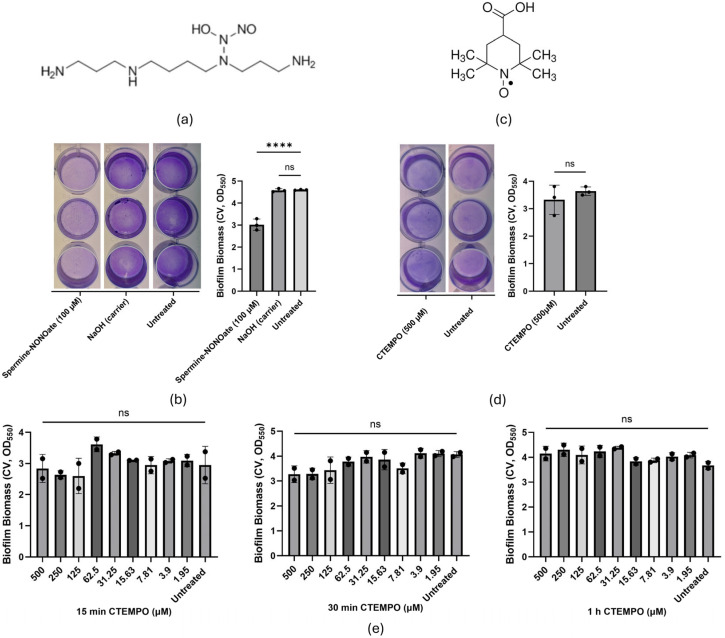 Figure 1
Figure 1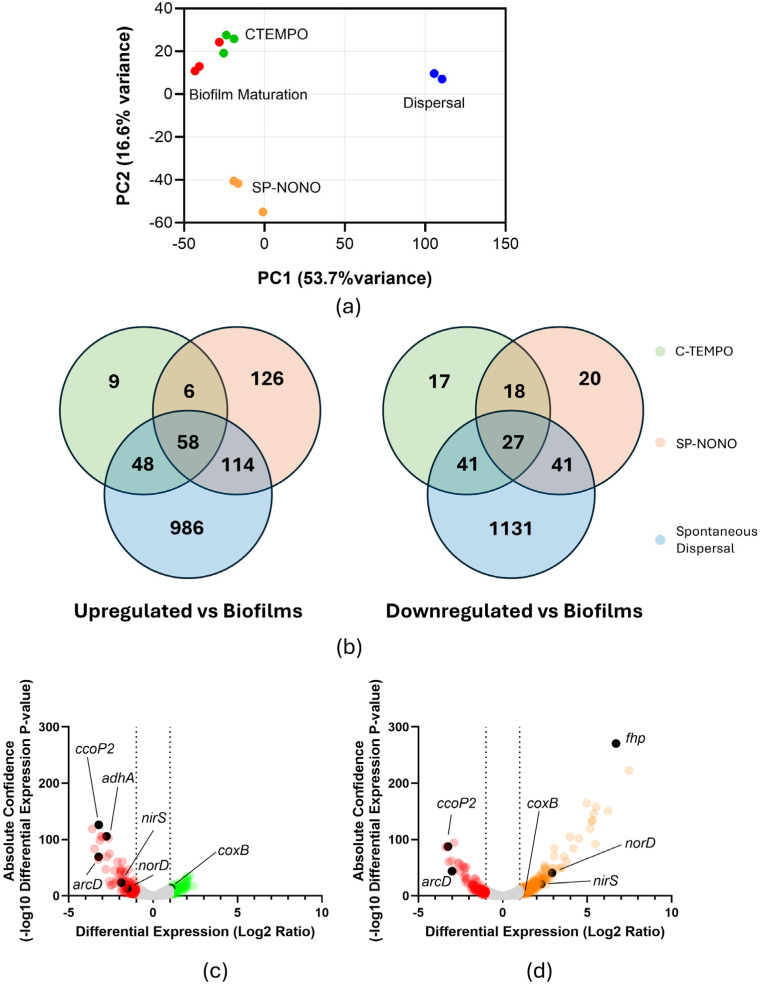 Figure 2
Figure 2- —Australian Research Council
- —Max Planck Queensland Centre on the Materials Science of Extracellular Matrices
- —Queensland University of Technology (Australia)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Vibrio bacteria research studies · Antimicrobial Peptides and Activities
1. Introduction
Bacterial biofilms are aggregates of sessile cells, typically attached to surfaces, and encased within an extracellular matrix [1]. During biofilm maturation, cells acquire an elevated tolerance to environmental stress, host immune defences and antibiotics [2]. As part of their life cycle, mature biofilms naturally undergo biofilm dispersal, activated by environmental or self-synthesised cues [3,4]. During dispersal, biofilm-residing cells regain planktonic attributes to resume a free-swimming lifestyle [5], while simultaneously losing their biofilm-associated antibiotic tolerance [6]. This makes dispersal-inducing compounds a promising strategy to combat chronic biofilm infections through the potentiation of antibiotics that are otherwise ineffective against biofilms.
The water-soluble radical gas nitric oxide (NO) has gained attention as one of the most promising dispersal agents, showing broad-spectrum effects on Gram-positive and Gram-negative bacteria [7,8]. In the opportunistic biofilm-forming bacterium Pseudomonas aeruginosa, the exogenous addition of NO-releasing compounds (NO donors) such as Spermine-NONOate (SP-NONO; Figure 1a) induced rapid reductions (~40–88%) in the biomass of surface-attached biofilms [9,10,11,12,13]. Furthermore, NO gas and NO donors have proven effective in the dispersal of highly recalcitrant P. aeruginosa isolates from cystic fibrosis patients in vitro, and reduced the bacterial load in their sputum when used adjunctively with antibiotics [8,12]. In P. aeruginosa, these responses are associated with the activation of pathways leading to the reduction of c-di-GMP, the intracellular second messenger controlling bacterial lifestyle, leading to increased motility through rhamnolipid production and reduced transcription of matrix exopolysaccharides [14,15,16,17]. Notably, dispersal occurs despite P. aeruginosa possessing distinct NO-responsive transcriptional regulators that tightly control the expression of detoxification mechanisms to prevent NO-derived toxicity. These NO-detoxification regulators are the dissimilatory nitrate respiration regulator DNR, and FhpR, a homolog of the E. coli transcriptional regulator NorR [10,18,19], which respectively regulate the expression of the NO reductase NorBC and the flavohaemoglobin Fhp [20,21].
Despite preclinical efficacy, the clinical application of NO remains challenging, as its small size, high diffusion capacity and pronounced chemical reactivity result in a short biological half-life and complicate controlled, localised delivery at the site of infection [22]. Nitroxides are considered NO analogues due to sharing key chemical features with NO while presenting significantly reduced reactivity [23]. Like NO, nitroxides possess a delocalised unpaired electron over the nitrogen and oxygen atoms. However, this group is sterically hindered by adjacent methyl groups that limit their reactivity (e.g., 4-carboxy-TEMPO; Figure 1b) [23]. Nitroxides were shown in some studies to induce slow dispersal of P. aeruginosa biofilms grown under continuous flow conditions (open culture systems) and potentiate the bactericidal activity of antibiotics, thus highlighting their utility as antibiofilm agents [24,25].
Recently, we reported the biofilm culture kinetics of P. aeruginosa PAO1 in closed culture systems, showing they recapitulated the main stages of the biofilm life cycle (attachment, maturation and spontaneous dispersal) described under continuous flow culture conditions [26]. Moreover, by characterising the transcriptomic profile of cells undergoing each stage, we temporally resolved canonical stage-specific transcriptional responses, such as the activation of surface-sensing pathways during attachment, polysaccharide production during biofilm maturation and quorum sensing signals during dispersal [26]. Notably, the upregulation of fourteen genes was associated with the onset of rapid loss of biofilm biomass, and this set was thus proposed to serve as specific biomarkers of biofilm dispersal [26]. Dispersal biomarkers included genes encoding transcriptional regulators amrZ (PA3385) and cdpR (PA2588); chemotaxis- and redox-associated genes cheR2 (PA0175), pqqA (PA1985) and PA0743; fimbrial- and cupin-mediated adhesion loci tadA (PA4302), rcpA (PA4304), rcpC (PA4305), flp (PA4306) and cupE1/E2 (PA4648 and PA4649); and uncharacterised genes PA0111, PA1353 and PA4523. Whether biofilm dispersal agents, like NO and nitroxides, activate transcriptional responses that govern spontaneous biofilm dispersal has not been investigated to date.
Here, we assessed the dispersal activity of NO donor Spermine-NONOate (SP-NONO) and nitroxide 4-carboxy-TEMPO (C-TEMPO) using microplate-grown P. aeruginosa biofilms. Subsequently, we performed RNA sequencing on biofilm cells treated with each compound and characterised their transcriptional profiles relative to spontaneous dispersal. Specifically, we report that >50% of genes differentially regulated under SP-NONO treatment were found similarly altered during spontaneous dispersal, and that SP-NONO, but not C-TEMPO, upregulated most transcriptional biomarkers associated with spontaneous dispersal in 12-hour-old biofilms. These findings suggest that NO prematurely activates spontaneous dispersal pathways to induce dispersal of P. aeruginosa biofilms. Collectively, our data indicate that these biomarkers robustly represent central pathways involved in the activation of dispersal responses in P. aeruginosa.
2. Results
2.1. The NO Donor SP-NONO, but Not the Nitroxide C-TEMPO, Rapidly Induced Biofilm Biomass Reduction of P. aeruginosa Biofilms in Closed Systems
To determine the treatment durations of SP-NONO and C-TEMPO required to induce biofilm dispersal, and to define appropriate time points for harvesting mRNA to assess gene expression changes, we performed biofilm dispersal assays using both compounds. In closed culture systems such as microtiter plates, P. aeruginosa PAO1 rapidly forms biofilms within 4 h of inoculation [13,26]. Accordingly, the dispersing activities of the C-TEMPO and SP-NONO were assessed at sub-inhibitory concentrations on P. aeruginosa biofilms grown for 4 h (Figure S1A). In this experimental model, dispersal is reported as a reduction in attached biofilm biomass, quantified by crystal violet staining at 550 nm of optical density, which we demonstrated to correlate with decreased biofilm-embedded cells after treatment with NO donors [13]. Consistent with our previous work [13], both untreated biofilms and mock-treated biofilms (NaOH vehicle control) showed high biomass (OD_550_~4.5), whereas biofilms treated with SP-NONO (100 µM, 15 min) exhibited significantly reduced biomass (OD_550_~3, Figure 1a). Based on these results, biofilm cells treated with SP-NONO (100 µM) for 15 min were used for downstream RNA-seq to characterise the transcriptional responses induced by this biofilm-dispersing NO donor.
To assess the dispersal activity of C-TEMPO in closed culture-grown P. aeruginosa PAO1 biofilms, we initially treated biofilms for 15 min (to match SP-NONO treatment) with a broad range of concentrations (500–1.95 µM; Figure 1e). No biomass changes were observed after 15 min of C-TEMPO treatment, or when treatment time was extended to 30 min or 1 h (OD_550_~3.5; Figure 1b,e). In contrast to previous reports using flow cell systems, these results indicate that C-TEMPO does not elicit detectable dispersal activity in closed biofilm cultures [24,27]. We reasoned that the divergent effects on biofilms displayed by two representatives of analogue compound families (NO donors and nitroxides) could be leveraged to identify common transcriptional variations induced by SP-NONO and nitroxides and, therefore, discriminate unique transcriptional responses by SP-NONO to refine candidate pathways associated with dispersal. Consequently, RNA-seq samples were collected from biofilm P. aeruginosa PAO1 cells treated with C-TEMPO (500 µM) for 30 min. This extended exposure reflects the distinct physicochemical properties of the compounds, accounting for the bulkier structure of C-TEMPO relative to NO (Figure 1a,c), which likely reduces the penetration kinetics of the compound. On the other hand, treatment durations were limited to 30 min, due to P. aeruginosa PAO1 biofilm culture kinetics rapidly progressing through stages in closed systems [26], suggesting that a longer treatment would risk confounding transcriptional differences arising from biofilm ageing.
2.2. SP-NONO and C-TEMPO Disrupt ANR-Regulated Energy Production Pathways
To compare the global transcriptional profiles between treated and untreated groups, RNA-seq data was plotted using ClustVis (Figure 2a). C-TEMPO-treated biofilms (Figure 2a—green) clustered closely with untreated biofilm-residing cells (Figure 2a—red) obtained from earlier work [26], indicating that C-TEMPO treatment induced minimal transcriptional changes in biofilms. In contrast, SP-NONO-treated biofilms (Figure 2a—orange) clustered separately from untreated biofilms, indicating that a significant transcriptional shift was induced. To quantify these responses, we used a negative binomial test (p-value ≤ 0.01; Log_2_ fold-change ≥ |1|) comparing NO-treated or C-TEMPO-treated samples to untreated, 4 h biofilm cells.
Treatment of biofilms with C-TEMPO altered the transcription of 224 genes (121 upregulated; 103 downregulated), with downregulated genes exhibiting markedly greater absolute fold changes compared to upregulated genes (Figure 2c), whereas SP-NONO treatment affected the transcription of 407 genes (304 upregulated; 103 downregulated) (Figure 2b,d). Despite that C-TEMPO treatment did not change biofilm biomass, unlike SP-NONO, we observed a substantial overlap in transcriptional responses under C-TEMPO and SP-NONO treatments, with 64/121 C-TEMPO-upregulated genes and 45/103 C-TEMPO-downregulated genes also similarly regulated by SP-NONO (Figure 2). The genes with the largest transcriptional differences shared between C-TEMPO and SP-NONO were involved in ANR-controlled oxidative phosphorylation and energy production (Table 1). In contrast, genes under regulation from dedicated NO sensors were uniquely upregulated by SP-NONO. These included genes encoding elements of the denitrification pathway (Table 1) or the aerobic NO detoxification flavohaemoprotein Fhp (104-fold, Table S1). Collectively, the observed differences in gene expression suggest that C-TEMPO overlaps with SP-NONO in disrupting O_2_-mediated transcriptional regulation, while being unable to induce canonical responses to NO.
2.3. SP-NONO Upregulates Metabolic Pathways of Spontaneous Dispersal
In a previous study, we identified the transcriptional profiles of P. aeruginosa in each stage of the biofilm life cycle, including attachment, biofilm maturation and, more importantly, spontaneous dispersal (Figure 2a—green) [26]. Considering that SP-NONO elicited significant reduction in biofilm biomass analogous to that observed during spontaneous dispersal (Figure 1c), we hypothesised that central responses involved in dispersal may be reflected in the transcriptional profiles of cells treated with SP-NONO. Indeed, we found 176/304 genes upregulated and 68/103 genes downregulated by SP-NONO that overlapped with transcriptional changes occurring during spontaneous dispersal (Figure 2b). To identify gene expression changes strictly associated with the dispersal phenotype, genes uniquely upregulated by SP-NONO and during spontaneous dispersal, but not by C-TEMPO, were compiled into Table 2 (a full list of genes was included in Table S2). Transcriptomic profiles associated with biofilm dispersal displayed a downregulation of genes predicted to be involved in the import of sulphur-containing metabolites such as sulphate (cysW, cysT and cysP) and taurine (PA3936–PA3938), based on KEGG pathway annotation [27]. cobP, cobU and cobV, involved in the cobalamin biosynthesis pathway, were similarly downregulated (Table 2) [28]. In contrast, pathways associated with energy generation were upregulated. These included genes involved in the catabolic degradation of amino acids such as lysine to glutarate (davD and davT) [29], or of valine, leucine and isoleucine into precursors of the TCA cycle (braC, bkdB, lpdV, PA3417 and ldh) (Table 2) [30,31], as well as genes involved in pyrroloquinoline quinone biosynthesis (pqqA, pqqD, pqqE and pqqF) (Table 2), a cofactor required for the periplasmic oxidation of ethanol [32]. Moreover, cells undergoing SP-NONO treatment or spontaneous dispersal promoted the upregulation of genes belonging to the Che2 chemotaxis system (PA0173–PA0179) (Table 2) [33], suggesting a role of this chemosensory pathway in biofilm dispersal.
2.4. SP-NONO Treatment of PAO1 Biofilms Upregulates Biomarkers of Spontaneous Dispersal
By analysing the temporal changes in global gene expression across the biofilm life cycle stages, we previously identified fourteen transcriptional biomarkers that are distinctly and reproducibly upregulated during spontaneous dispersal in 12-hour-old biofilms [26]. Importantly, the dispersal-inducing NO donor SP-NONO largely recapitulated this distinct transcriptional response of spontaneous dispersal, whereas C-TEMPO did not (Table 3). Of the fourteen dispersal biomarkers, SP-NONO significantly increased the transcription of nine. These included PA0111 (6.62-fold), cheR2 (3.41-fold), pqqA (2.52-fold), tadA (2.21-fold), rcpA (2.12-fold), rcpC (2.18-fold), flp (2.27-fold) and cupE1E2 (2.36- and 2.09-fold, respectively) (Table 3). In contrast, C-TEMPO only upregulated three biomarkers: PA0111 (5.15-fold), tadA (2.01-fold), and flp (2.20-fold) (Table 3). The observed transcriptional overlap suggests that SP-NONO signals the upregulation of pathways activated during spontaneous dispersal to prematurely elicit a reversion to the planktonic lifestyle. Furthermore, the upregulation of this set of genes during exogenously induced and spontaneous dispersal (reported as reduced surface-attached biofilm biomass) in closed culture systems strongly supports them as biomarkers representative of central dispersal pathways in P. aeruginosa.
3. Discussion
Biofilm dispersal agents have gained increasing attention as a therapeutic approach to enhancing antibiotic activity against clinical biofilms, which are the source of ~80% of chronic hospital infections. Biofilm infections show elevated incidence in prostheses and in-dwelling devices, leading to bacteraemia, ventilator-associated respiratory infections and catheter-associated urinary tract infections [34,35]. Among biofilm-forming bacterial pathogens, P. aeruginosa stands out due to causing ~7% of all healthcare-associated infections, with incidence rates as high as ~23% in ICU infections and in cystic fibrosis patients, causing highly recalcitrant endobronchiolitis, bronchiectasis, and pneumonia [36,37]. To fast-track screening of dispersal-inducing compounds, we previously identified a subset of P. aeruginosa genes that serve as transcriptional biomarkers of the onset of spontaneous dispersal [26]. Here, we demonstrate the robustness of these biomarkers using the biofilm dispersal NO donor SP-NONO, which effectively dispersed P. aeruginosa biofilms and elicited the upregulation of 9 of the 14 biomarkers. In contrast, the nitroxide C-TEMPO, which did not promote biofilm dispersal, only stimulated the upregulation of three dispersal biomarkers.
As a nitroxide, C-TEMPO possesses a significantly bulkier structure than NO and is sterically hindered by the four adjacent methyl groups. This implies significantly reduced reactivity and penetration relative to NO [23], which we addressed by testing a range of concentrations and treatment times. Nevertheless, no significant changes in attached biomass were observed in microplate-grown biofilms by any C-TEMPO treatment tested. Our data contrast with previous reports of C-TEMPO-mediated biofilm dispersal of P. aeruginosa in flow cells [24,25]. Importantly, P. aeruginosa biofilms were cultured for 48 h prior to being treated with C-TEMPO for a further 24 h, when an increased cell density was detected in the culture effluent [25]. Under flow culture conditions, the constant stream of media replenishes nutrients while removing metabolic by-products and quorum sensing molecules [38]. Therefore, biofilm maturation may span days, leading to the formation of niches within the multicellular community [25,39]. This is supported by reports indicating that ratio of exopolysaccharides to biofilm cell density is enhanced in Pseudomonas fluorescens under flow conditions relative to biofilms in closed systems [39]. Additionally, shear forces under flow conditions promote P. aeruginosa surface attachment and c-di-GMP biosynthesis, leading to increased biofilm formation [40]. Longer maturation times under conditions promoting the synthesis of matrix components likely promotes physiological variations relative to younger biofilms under closed culture conditions, which would lead to the development of nitroxide-responsive subpopulations.
Contradictory dispersal activity across different biofilm models, however, is not unique to nitroxides. Previously, we reported that the NO donor sodium nitroprusside, which has been widely documented to disperse P. aeruginosa biofilms in continuous flow cultures, unexpectedly increased biomass of P. aeruginosa biofilms cultured in microtiter plates [13]. While no studies have directly compared transcriptomic or metabolomic differences in biofilms of the same strain cultured under different culture platforms, it is likely that longer incubation times in continuous flow cultures account for biological differences in P. aeruginosa biofilms, thus affecting the dispersal activity of some drug candidates.
Here, we report for the first time the transcriptional response of P. aeruginosa to C-TEMPO treatment. This analysis revealed pronounced changes in genes whose transcription is modulated by the transcriptional regulator of anaerobiosis ANR and the redox-responsive two-component regulator RoxSR, which were also identified under SP-NONO treatment. These include genes required for survival under conditions of hypoxia [41], such as the NO-sensitive regulator encoded by dnr, the operon ccoP2-ccoO2 encoding the high-affinity cbb3-type cytochrome C oxidase, the aa3-type cytochrome encoded by coxAB, and cioAB, encoding a cyanide-insensitive terminal oxidase [42]. Additionally, the ANR-regulated operon, arcDABC, and adhA were among the most downregulated genes by C-TEMPO and SP-NONO. These genes respectively encode the arginine deiminase pathway for the degradation of arginine to ornithine and a NAD^+^-dependent alcohol dehydrogenase, which participate in processes necessary for ATP generation under anaerobiosis, and are defined as strongly regulated by ANR and DNR [41,43]. In contrast, our analysis revealed no observable overlap between the SP-NONO and C-TEMPO regarding NO-mediated transcriptional regulation. Neither genes involved in denitrification (including the NIR, NOR and NOS operons), nor the O_2_-dependent flavohaemoglobin Fhp were upregulated by C-TEMPO, whereas SP-NONO strongly stimulated their transcription. Hence, despite nitroxides being widely proposed to act as NO analogues, our data suggest that C-TEMPO would primarily interact with ANR-regulated pathways, and that these would not be directly involved in SP-NONO-mediated dispersal.
NO is a well-established biofilm dispersal agent, known to increase cell motility and stimulate the enzymatic hydrolysis of the cellular signalling molecule cyclic-di-GMP [14]. Here, we reported the overlapping transcriptomic responses of chemically and spontaneously dispersed P. aeruginosa PAO1 biofilms in closed cultures. In both datasets, genes involved in the catabolism of valine, leucine and isoleucine (braC, bkdB, lpdV, PA3417 and ldh) were upregulated, together with genes encoding enzymes mediating the degradation of the lysine metabolite δ-aminovalerate to glutarate (davD and davT) [44], and a δ-aminovalerate-ABC transporter (agtABC but not agtD) [45]. Additionally, spontaneously and SP-NONO-dispersed cells largely upregulated dctA encoding the C_4_-dicarboxylic acid transport for the primary import of succinate, malate and fumarate [46], and PA0752-PA0754 (tctABC), encoding a citrate and cis-aconitate import [47]. Metabolic pathways involved in amino acid catabolism have also been reported as upregulated in independent transcriptomic studies of dispersal using SP-NONO [9]. Namely, bkdA1, bkdA2, bkdB and lpdV were amongst the most upregulated genes in SP-NONO-dispersed cells relative to untreated biofilms [48], suggesting that the activation of amino acid catabolic pathways and transport systems for TCA cycle intermediates may be derived from a dispersal-induced metabolic shift aiding cell reversal to the planktonic lifestyle.
In closed systems, we report that treatment with the NO donor SP-NONO caused maturing P. aeruginosa biofilms to upregulate PA0111, cheR2, pqqA, tadA, rcpA, rcpC, flp, cupE1 and cupE2. These were proposed as transcriptional biomarkers of dispersal, as their transcription was largely increased in biofilms undergoing spontaneous dispersal [26]. Here, we demonstrate that, despite 4-hour-old and 8-hour-old biofilms displaying markedly distinct transcriptomic profiles, treatment with SP-NONO prematurely induced the discrete upregulation of most transcriptional biomarkers of dispersal. Therefore, we here report an overlap between two transcriptional signatures corresponding to distinct modes of dispersal induction at different biofilm ages, indicating that NO promotes the upregulation of a core set of transcriptional biomarkers associated with the onset of spontaneous biofilm dispersal.
4. Conclusions
Altogether, the findings presented in this work suggest that NO elicits a metabolic burst in biofilm cells, and that a disruption in biofilm maturation occurs through the upregulation of endogenous central pathways that govern the transition from the biofilm to the planktonic lifestyle. Importantly, these data establish a relevant transcriptional framework that can serve as a benchmark for the future screening and mechanistic evaluation of candidate biofilm-dispersing agents.
5. Materials and Methods
5.1. Strains, Media and Culture Conditions
Pseudomonas aeruginosa PAO1 cultures were routinely grown overnight in LB (lysogeny broth) media at 37 °C, 200 rpm before incubation in fresh M9 media (9 mM NaCl, 22 mM KH_2_PO_4_, 48 mM Na_2_HPO_4_, 19 mM NH_4_Cl, 2 mM MgSO_4_, 100 µM CaCl_2_, 0.4% glucose, pH 7.0) at 37 °C with shaking.
5.2. Chemical Preparation and Storage
Spermine-NONOate (SP-NONO, CAT#0634655-16, (Z)-1-[N-[3 aminopropyl]-N-[4-(3-aminopropylammonio)butyl]-amino]diazen-1-ium-1,2-diolate) (Cayman Chemical, Ann Arbor, MI, USA) was dissolved in 10 mM NaOH to a final concentration of 10 mM and stored at −20 °C. Stock solutions were used within 3 months. 4-carboxy-TEMPO (C-TEMPO, CAT# 23139, 4-carboxy-2,2,6,6-tetramethyl-1-piperidinyloxy) (Cayman Chemical, Ann Arbor, MI, USA) was dissolved in water to a 10 mM solution. Only freshly made solutions were used in dispersal assays.
5.3. Biofilm Dispersal Assays
Biofilm formation and dispersal assays were performed as previously described [13]. Briefly, 10^7^ colony-forming units (CFU)/mL bacterial suspensions were prepared in M9 media and inoculated into a 24-well plate (Nunc, ThermoFisher Scientific, Waltham, MA, USA). Biofilms were grown at 37 °C, 180 rpm. At 4 h post-inoculation, SP-NONO (100 µM; Cayman Chemical), NaOH (100 µM) or 4-carboxy-TEMPO (500 µM; C-TEMPO; Cayman Chemical) were added to a final concentration of 100 µM or 500 µM, respectively, and incubated for another 15 min or 30 min. Wells were stained with 0.1% (w/v) crystal violet in 6.25% (v/v) methanol for 20 min, washed twice with 1 mL phosphate-buffered saline (PBS, Gibco, Franklin, TN, USA) and finally solubilised with ethanol (absolute). Optical density at 550 nm (OD_550_) was measured with a SPECTROStar Nano microplate reader (BMG LabTech, Ortenberg, Germany). Micrographs of stained biofilm were taken as described above.
5.4. RNA Sequencing and Analysis
Biofilms were incubated in tissue culture flasks as previously described with some adjustments [26]. Briefly, tissue culture flasks (Nunc, ThermoFisher Scientific, Waltham, MA, USA) were seeded with 10^7^ CFU/mL cells in 50 mL of M9 medium, and biofilms were grown at 37 °C with shaking (70 rpm). At 4 h, biofilms were treated with either SP-NONO (100 µM) or C-TEMPO (500 µM) for 15 min or 30 min. After treatment, the liquid phase was discarded, and the remaining attached cells were gently washed with PBS. Surface-attached cells were resuspended in a 1:2 mixture of PBS and RNA-protect solution (QIAGEN, Cat# 76506, Venlo, The Netherlands) using a cell scraper. Resuspended cells (~10^8^) were pelleted at 5000 g (10 min, 25 °C). RNA extraction was performed with the RNeasy mini kit (QIAGEN, Cat# 74104, Venlo, The Netherlands) following the manufacturer’s protocol. Samples were treated with DNase and subjected to RNA sequencing using DNBSEQ PE100 (BGI Genomics, Shenzhen, China). Cleaned reads were mapped to P. aeruginosa PAO1 chromosome (AE004091.2) using Bowtie2 v 1.50.2. Differentially transcribed genes between groups (untreated biofilms, biofilms treated with SP-NONO and biofilms treated with 4-carboxy-TEMPO; see Table S1) were identified with DESeq2 on Geneious Prime (v2024.07, Dotmatics, Boston, MA, USA). Principal component analysis (PCA) was conducted exclusively as an exploratory visualisation to assess overall transcriptomic similarity within biological replicates and to examine separation between treatment groups. For this purpose, the reads per kilobase of transcript per million mapped reads (RPKM) of genes with >|2-fold| transcription and p-value < 0.01 were employed. RPKM data was transformed into PCA scores using ClustVis [49], and subsequently plotted (Graphpad Prism 10.4.1, La Jolla, CA, USA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Flemming H.C. Wingender J. The biofilm matrix Nat. Rev. Microbiol.2010862363310.1038/nrmicro 241520676145 · doi ↗ · pubmed ↗
- 2Ciofu O. Tolker-Nielsen T. Antibiotic tolerance and resistance in biofilms Biofilm Infections Bjarnsholt T. Jensen P.Ø. Moser C. Høiby N. Springer New York, NY, USA 201121522910.1007/978-1-4419-6084-9 · doi ↗
- 3An S. Wu J. Zhang L.H. Modulation of Pseudomonas aeruginosa biofilm dispersal by a cyclic-di-gmp phosphodiesterase with a putative hypoxia-sensing domain Appl. Environ. Microbiol.2010768160817310.1128/AEM.01233-1020971871 PMC 3008239 · doi ↗ · pubmed ↗
- 4Sauer K. Cullen M.C. Rickard A.H. Zeef L.A.H. Davies D.G. Gilbert P. Characterization of nutrient-induced dispersion in Pseudomonas aeruginosa PAO 1 biofilm J. Bacteriol.20041867312732610.1128/JB.186.21.7312-7326.200415489443 PMC 523207 · doi ↗ · pubmed ↗
- 5Barraud N. Kjelleberg S. Rice S.A. Dispersal from Microbial Biofilms Microbiol. Spectr.2015334336210.1128/microbiolspec.MB-0015-201427337281 · doi ↗ · pubmed ↗
- 6Chambers J.R. Cherny K.E. Sauer K. Susceptibility of Pseudomonas aeruginosa Dispersed Cells to Antimicrobial Agents Is Dependent on the Dispersion Cue and Class of the Antimicrobial Agent Used Antimicrob. Agents Chemother.201761 e 00846-1710.1128/AAC.00846-1728971863 PMC 5700346 · doi ↗ · pubmed ↗
- 7Barraud N. Storey M.V. Moore Z.P. Webb J.S. Rice S.A. Kjelleberg S. Nitric oxide-mediated dispersal in single- and multi-species biofilms of clinically and industrially relevant microorganisms Microb. Biotechnol.2009237037810.1111/j.1751-7915.2009.00098.x 21261931 PMC 3815757 · doi ↗ · pubmed ↗
- 8Howlin R.P. Cathie K. Hall-Stoodley L. Cornelius V. Duignan C. Allan R.N. Fernandez B.O. Barraud N. Bruce K.D. Jefferies J. Low-Dose Nitric Oxide as Targeted Anti-biofilm Adjunctive Therapy to Treat Chronic Pseudomonas aeruginosa Infection in Cystic Fibrosis Mol. Ther.2017252104211610.1016/j.ymthe.2017.06.02128750737 PMC 5589160 · doi ↗ · pubmed ↗