T-2 Toxin Neurotoxicity: Molecular Mechanisms and Emerging Chemoprotective Strategies
Chunyan Xu, Gea Oliveri Conti, Shusheng Tang, Jianzhong Shen, Chongshan Dai

TL;DR
This review explores how T-2 toxin causes brain damage and discusses new ways to protect against its harmful effects.
Contribution
The paper provides new insights into molecular mechanisms of T-2 toxin-induced neurotoxicity and novel chemoprotective strategies.
Findings
T-2 toxin causes neurotoxicity through oxidative stress, mitochondrial dysfunction, and neuroinflammation.
Several signaling pathways are involved in T-2 toxin-induced neurotoxicity.
Antioxidants and natural products can reduce the harmful effects of T-2 toxin.
Abstract
Mycotoxins can contaminate food and raw food materials and are a threat to animal and human health. T-2 toxin is the most toxic secondary metabolite mainly produced by Fusarium species among trichothecenes. T-2 toxin exposure can induce multiple toxic effects, including hepatotoxicity, nephrotoxicity, immunotoxicity, gastrointestinal toxicity, and reproductive toxicity. Recent studies have reported that T-2 toxin can cross the blood–brain barrier and trigger neurotoxicity. In this review, we summarized the neurotoxic effects caused by T-2 toxin exposure and the underlying molecular mechanisms. Additionally, effective neuroprotective agents, potential clinical applications, and future prospects are discussed. The current studies revealed that the molecular mechanisms of T-2 toxin-induced neurotoxicity involve oxidative stress, mitochondrial dysfunction, neuroinflammation, autophagy,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
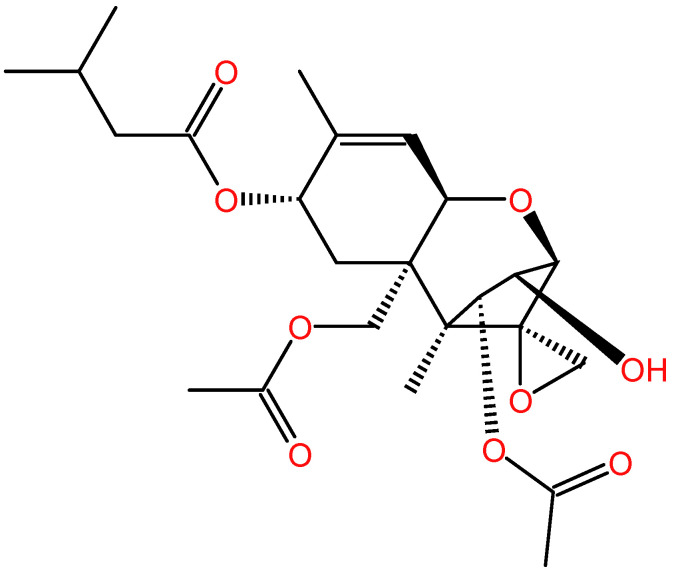 Figure 1
Figure 1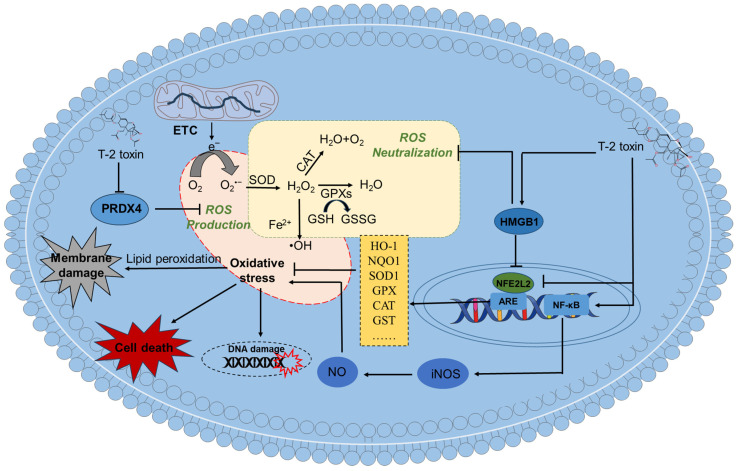 Figure 2
Figure 2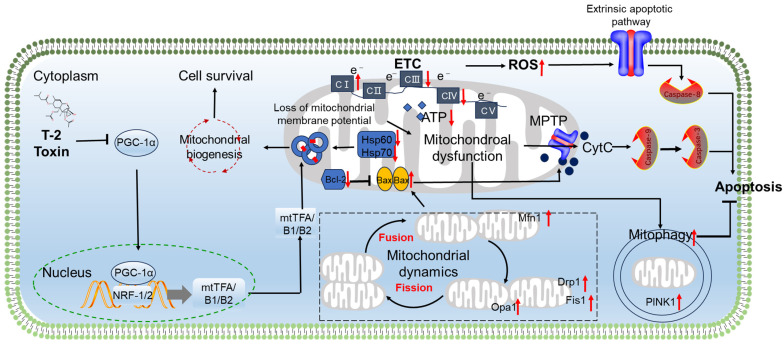 Figure 3
Figure 3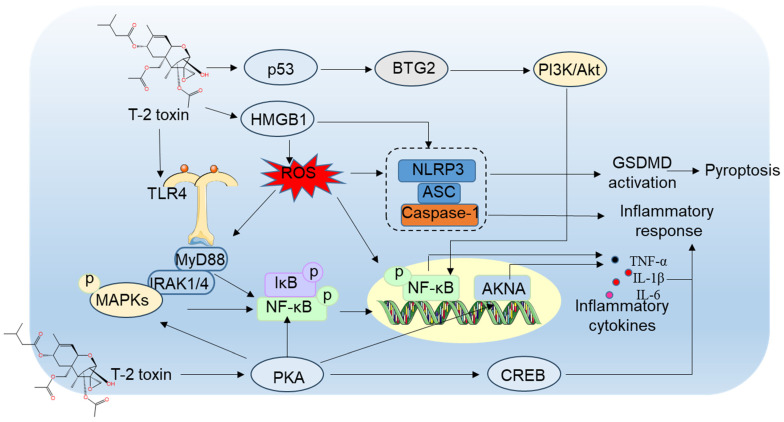 Figure 4
Figure 4 Figure 5
Figure 5- —Beijing Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycotoxins in Agriculture and Food · Plant and fungal interactions · Marine Toxins and Detection Methods
1. Introduction
Mycotoxins are structurally diverse secondary metabolites produced by filamentous fungi, such as Fusarium, Penicillium, and Aspergillus. Worryingly, mycotoxins commonly contaminate the human food chain across various grains, food, fruits, nuts, Chinese herbal medicines, drinking water, and feed, posing a threat to animal and human health [1,2,3,4,5,6]. Currently, due to climate change and the development of detection technology, the contamination rate of mycotoxins has increased to 60–80%, far exceeding 25% in the 1980s. This figure may be even higher in low-income countries [7,8,9]. In addition, the diversity of mycotoxins and the multifaceted toxic effects further exacerbate the difficulty of preventing and controlling fungal toxins.
To date, more than about 700 mycotoxins have been detected and identified in food and food raw materials [10,11]. Of these identified mycotoxins, many mycotoxins exhibit potent toxic effects to humans and animals even at low levels, and therefore pose a serious threat to public health and food security [5,12]. For example, aflatoxin B1 (AFB1), ochratoxin A (OTA), and fumonisin B1 (FB1) were classified as human Groups 1, 2B, and 2B carcinogens, respectively, by the WHO International Agency for Research on Cancer [13]. Epidemiological studies have shown a positive correlation between exposure to certain mycotoxins and the occurrence of infertility, cardiovascular disease and cancer [14,15,16,17].
T-2 toxin (Figure 1) is the most toxic secondary metabolite among trichothecenes, mainly produced by Fusarium species, which commonly occur typically in various food crops, such as barley, wheat, maize, rice, oat, and animal feed in the field or during storage [18]. Recent studies showed that T-2 toxin is also detected in drinking water in endemic areas of China (e.g., Qinghai and Sichuan Provinces) and in traditional Chinese medicines [19,20,21,22]. A recent epidemiological investigation showed that the contents of T-2 toxin in brick tea are positively related with the occurrence and development of Kashin–Beck disease in Tibet, China [23]. It poses a high risk in the field of public health and safety.
Over the past forty years, the toxicology files of T-2 toxin have been extensively studied. T-2 toxin exhibits a potent acute toxicity, and the lethal doses of 50% of T-2 toxin via the intravenous injection at a signal dose in pigs and rats are 1.21 and 0.9 mg/kg body weight, respectively [24,25,26]. The exposure of T-2 toxin and its metabolic derivatives (such as T-2 toxin and T-2-glucoside A) to rodents can induce hepatoxicity, nephrotoxicity, immunotoxicity, reproductive toxicity, cardiac toxicity, skin toxicity and gastrointestinal toxicity [27,28,29,30,31,32,33].
T-2 toxin can also cross the blood–brain barrier and enter brain tissues, then induce brain injury, finally culminating in neurotoxicity [34,35,36]. Animal studies found that T-2 toxin exposure can induce anorexia and decrease learning and memory [37,38]. Mechanistic investigations revealed that T-2 toxin-mediated neurotoxicity involves mitochondrial dysfunction, oxidative stress, neuroinflammation, autophagy, pyroptosis, ferroptosis and cell apoptosis [37,38,39]. Several signaling pathways, including NFE2L2, NRF-2, PGC-1, p53, BTG2, AKNA, MAPK, Akt, mTOR, HMGB1, CREB, and NF-κB, are known to be involved. In the present review, we utilized keywords such as “T-2 toxin” and “neurotoxicity” or “neurotoxic effects” to gather research on the neurotoxic effects of T-2 toxin and the underlying molecular mechanisms from databases such as Web of Science and PubMed, from 1980 up to December 2025. The collected information was then summarized and discussed. Additionally, we discuss effective neuroprotective agents and their clinical application. We hope this review can provide valuable insights into the underlying causes of T-2 toxin-induced neurotoxicity and effective detoxification strategies.
2. T-2 Toxin’s Metabolism and the Accumulation in Brain Tissues
Typically, ingestion serves as the main route of T-2 toxin poisoning. Once ingested, it is absorbed across the gastrointestinal tract and enters the bloodstream. In mammals, the biotransformation of T-2 toxin mainly occurs in liver tissue, and this process involves conjugation, deoxidation, and hydroxylation reactions [40]. Correspondingly, it is metabolized to T-2 triol, 3′-OH-HT-2, 3′-OH-T-2, HT-2, and neosolaniol (NEO) [41,42,43,44,45]. Early toxicological studies reported that the half-lives of T-2 and HT-2 toxins are approximately 21 and 73 min after intramuscular injection in dogs. In whole blood, HT-2 and T-2 toxins were both unstable and can be degraded by carboxyl-esterase enzymes in the red blood cells, and the corresponding stability half-lives were 0.84 and 6.9 h, respectively [46,47,48]. In human primary astrocytes, T-2 toxin can be quickly uptaken then subjected to metabolism, leading to HT-2 toxin [36].
Guo et al. found that, when rats were orally given T-2 toxin at 2 mg/kg body weight, low concentrations of T-2 toxin could be detected in the brain tissues on the 1st day, but it was undetectable on the 3rd and 7th days [49]. This may be relative to the stability of T-2 toxin. The C-3′ hydroxylation of T-2 and HT-2 toxins can be mainly catalyzed by intracellular cytochrome P450 enzymes (CYP450s), such as CYP1A1, CYP1A2, CYP2J2, CYP2E1, CYP3A4, CYP3A9, CYP3A11 and CYP3A13. In general, these CYP450s are mainly expressed in the liver tissues. Recent studies showed that they are also expressed in the brain tissues of both rodents and humans, although the levels are low [50,51,52,53,54]. This indicated that T-2 toxin can be catalyzed in brain tissues. Consistently, it was reported that a higher dose of T-2 toxin (i.e., at 100 ng/mL) can significantly upregulate the expression of CYP4501A1 and CYP4503A via the activation of aromatic hydrocarbon receptors (AhR) and then can conversely regulate the metabolism of T-2 toxin [55]. This information indicated that T-2 toxin can be metabolized in brain tissues. This can also explain the detected low level of T-2 toxin in the brain tissues. However, the precise mechanisms are still unclear and require further exploration.
Additionally, with the development of technology, the detection of T-2 toxin is becoming increasingly sensitive, and the minimum detection limit can reach 0.1 ng/mL in human urine samples [56,57]. A recent study investigated the levels of T-2 toxin, T-2 Toxin-3-Glucoside and their metabolites in human urine samples (a total of 300 samples) in South Italy using a high-resolution mass spectrometry method. It found that almost all the major T-2 metabolites can be detected, including 3′-OH-T-2, T-2 triol, HT-2, NEO, HT-2-3-GlcA, T-2-3-GlcA, and HT-2-4-GlcA. The levels of 3′-OH-T-2, T-2 triol, HT-2, NEO, and HT-2-3-GlcA are high, and they can be potential monitoring biomarkers. T-2 was quantified in 21% of samples at a mean concentration of 1.34 ng/mg creatinine [56].
Currently, toxicokinetic or biomonitoring studies of T-2 toxin and metabolites in humans are severely lacking, which significantly limits effective risk assessment. More research is still needed in the future.
3. An Overview of T-2 Toxin Exposure-Induced Neurotoxic Effects
In humans, exposure to T-2 toxin is primarily from the ingestion of contaminated cereals and grains or the food chain [58]. Given the ubiquitous occurrence of Fusarium species across agricultural landscapes, there is a possibility of persistent low-level exposure for the global population. A recent epidemiological investigation suggested a plausible connection between mycotoxin contamination and an elevated susceptibility to neurodegenerative conditions such as Alzheimer’s and Parkinson’s diseases [59]. Nevertheless, these analyses frequently encounter limitations due to confounding elements, including the co-occurrence of multiple mycotoxins and other environmental contaminants, which complicates the causation of T-2 toxin-caused toxic effects in humans.
The neurobehavioral effects of T-2 toxin exposure range from acute neurological deficits to chronic behavioral problems [26,37,38,49,60,61,62,63]. For example, in rodent studies, acute T-2 toxin exposure results in vomiting, rapid heart rate, diarrhea, a lack of coordination, muscle weakness, a loss of appetite, depression, and reduced movement [26,37,38,60,61,62,63,64]. Guo et al. found that, when rats were given T-2 toxin orally at a dose of 2 mg/kg body weight, it caused marked brain damage and abnormal neurological responses (e.g., fear) [49]. Similarly, Li et al. showed that T-2 toxin treatment via an intraperitoneal injection at 4 mg/kg body weight can significantly reduce spatial learning, memory, and movement abilities in a mouse model [37]. More recently, Chen et al. reported that T-2 toxin treatment at 1.5 mg/kg body weight every day for 14 days via oral administration can induce depressive-like behaviors such as feeling hopeless and losing interest in pleasurable activities, without showing anxiety [65]. In addition, an exposure to T-2 toxin caused obvious histopathological changes in the brain tissues of mice [49,64,66]. Maroli and his colleagues found that, when T-2 toxin was injected intravenously at 2, 4, or 6 μg/kg body weight, it led to gliosis with acute inflammatory infiltrates in the cerebral and hippocampal tissues. It also damaged glial cells and caused neuroepithelial cell apoptosis [66]. Similarly, Guo et al. found that, when rats were exposed to T-2 toxin orally at 2 mg/kg body weight, it caused bleeding and damage to the brain tissue [49]. Pei et al. showed that, when rats were administrated orally with T-2 toxin at 1–2 mg/kg every day for 28 days, serious hippocampal damage, with disordered cell arrangements and neuronal degeneration, was observed in the brain tissues [67]. Moreover, T-2 toxin can harm the vascular system (such as microvascular dilation and swelling) in brain tissues [49,68,69,70,71,72,73,74,75,76,77]. Table 1 summarizes the various T-2 toxin exposure-caused neurotoxic effects in vivo and in vitro.
4. Roles of Neurotransmitters in T-2 Toxin-Induced Neurotoxicity
An exposure to T-2 toxin can have a significant impact on central nervous system (CNS) neurotransmitter levels. For instance, Chi et al. reported that T-2 toxin significantly increased the levels of dopamine (DA) and norepinephrine in chicken brain tissue [78]. This suggests that changes in catecholamines may predominantly contribute to the neurotoxicity induced by T-2 toxin. On the contrary, Huang et al. found that a single intraperitoneal injection of T-2 toxin at a dose of 4 mg/kg body weight significantly decreased the levels of DA, 5-HT, and acetylcholine (Ach) in mouse brain tissue [74]. More recently, Chen et al. observed that an exposure to T-2 toxin significantly decreased the levels of DA by increasing the expression of the dopamine transporter in the nucleus accumbent of the male mouse brain, which then triggers depressive-like behavior [65]. Additionally, Wang et al. showed that the oral administration of T-2 toxin at doses of 0.1, 1, and 2.5 mg/kg body weight can dose-dependently perturb the levels of 5-HT, 5-hydroxyindole acetic acid (5-HIAA), norepinephrine, and DA in specific rat brain regions within 10 h. Notably, these authors reported a significant increase in 5-HT at 2 h after dosing in the nucleus raphe magnus and locus coeruleus across all treatment groups and an increase in DA at 6 h in the locus coeruleus and hypothalamic paraventricular nucleus [79]. This suggests that the effects of T-2 toxin on neurotransmitters in different brain regions may be diverse and varied.
It is known that cholinergic and glutamatergic receptors in the subgranular zone are important for maintaining the proper proliferation and differentiation of granule cell lineages in the hippocampal formation of the brain [80,81,82]. It was reported that T-2 toxin exposure in the pregnant mice significantly decreased the transcript levels of cholinergic and glutamate receptor subunits (i.e., cholinergic receptor nicotinic beta 2 subunit [chrnb2], cholinergic receptor nicotinic alpha 4 subunit [chrna4], and glutamate receptor 2 [gria2]) and glutamate transporters (i.e., solute carrier family 17 member 6 [Slc17a6]) in the dentate gyrus in the offspring, indicating decreased cholinergic signals on hilar GABAergic (which produce gamma-aminobutyric acid [GABA]) interneurons, innervating type-2 cells and decreasing glutamatergic (which produce glutamate) signals in type-1 and type-2 cells [81]. This information indicated that T-2 toxin exposure in pregnant mice may disturb the production of GABA and glutamate, thus affecting the development of offspring hippocampal tissue via disturbing glutamatergic and GABAergic pathways. However, the precise molecular mechanisms remain unclear, and further investigations are required.
Overall, this collective evidence indicates that the effects of T-2 toxin on neurotransmitters are complex and depend on various factors. Of note, the changes in several neurotransmitters including DA, 5-HT, and Ach are various, and this may be dependent on the dosage, animal species, exposure time, and brain region. It may also affect glutamatergic and GABAergic pathways to disturb the development of offspring hippocampal tissue. To date, these precise molecular mechanisms remain unclear, and further investigations are still required.
5. Molecular Mechanisms of T-2 Toxin-Induced Neurotoxicity
The fundamental molecular pathways associated with T-2 toxin-induced neurotoxicity have been comprehensively investigated, and key pathways include the overproduction of reactive oxygen species (ROS), oxidative stress, inflammatory responses, mitochondrial dysfunction, apoptosis and autophagy. The aforementioned cellular events are driven by various signaling mechanisms, such as nuclear factor erythroid 2-related factor 2 (NFE2L2), NRF-2, p53, PGC-1α, HIF-1, p53, MAPK, serine/threonine protein kinase/mammalian target of rapamycin (Akt/mTOR), HMGB1, CREB, and NF-κB. In the following section, we will discuss these elements in depth.
5.1. Role of Oxidative Stress
The excessive production of reactive oxygen species (ROS) can induce oxidative stress damage. In vitro and in vivo studies have shown that T-2 toxin can cause oxidative stress damage in nerve cells (including neuronal cells or glial cells) or brain tissues [12,49,67,70,83,84,85]. Zhang et al. found that exposing mouse N2a cells to 5–80 ng/mL of T-2 toxin for 24 h can significantly increase the levels of intracellular ROS and malondialdehyde (MDA), a lipid peroxidation biomarker [12,66]. T-2 toxin can also downregulate the activities of several intracellular antioxidant enzymes, such as SOD and CAT, and the levels of GSH [12]. Consistently, it was reported that exposing PC12 or microglial cells to T-2 toxin can significantly increase intracellular ROS and MDA levels and significantly decrease the activities of SOD and CAT and the levels of GSH [70,72]. Similarly, T-2 toxin treatment at 1.57 mg/kg body weight via subcutaneous administration significantly decreased the activities of SOD and CAT and levels of GPX in the brain tissues of mice at the 1st day after administration, and an opposite trend was found in the experiment at the 7th day [61]. Huang et al. showed that a single intraperitoneal injection of 4 mg/kg body weight of T-2 toxin significantly increased ROS and MDA levels and decreased the activities of CAT and GPX and GSH levels in mouse brain tissues [74]. These data suggest that an imbalance of the intracellular antioxidant system was involved in T-2 toxin exposure-induced neurotoxicity.
Similar to ROS, the production of reactive nitrogen species (RNS) also contributes to oxidative stress in brain tissues. RNS include nitric oxide (NO) and its derivatives, such as peroxynitrite (ONOO−), which can cause DNA damage [86]. Pei et al. found that treating PC12 cells with 3–12 ng/mL of T-2 toxin for 24 h can significantly upregulate the expression of NF-κB and iNOS proteins, ultimately promoting the production of NO [70]. This indicates that the activation of the NF-κB/iNOS/NO signaling pathway also partly contributes to T-2 toxin-induced oxidative stress and neurotoxicity. In addition, different models of oxidative stress have been studied to clarify the effects of oxidative stress on NF-κB-related activities [87].
Supplementation with N-acetylcysteine (NAC), an aminothiol and synthetic precursor of intracellular cysteine and GSH, significantly reduces ROS production, effectively alleviating T-2 toxin-induced lipid peroxidation, oxidative damage and cytotoxicity in N2a neuronal cells [12]. Moreover, a recent study revealed that exposing GH3 cells to 10 or 40 nM of T-2 toxin significantly decreased the expression of peroxiredoxin 4 (PRDX4) protein; furthermore, the overexpression of PRDX4 significantly promoted T-2 toxin-induced ROS production, mitochondrial dysfunction and cell apoptosis [88]. PRDX4 is the only secreted antioxidant enzyme in the peroxidase family capable of converting H_2_O_2_ into harmless O_2_ to mitigate oxidative stress. It also plays a crucial role in essential biological processes, including protein folding, DNA repair, inflammatory regulation and tumor development [89]. These findings suggest that T-2 toxin-induced oxidative stress may be partly due to its inhibition of PRDX4 enzyme expression.
Nuclear factor erythroid 2-related factor 2 (NFE2L2) is a crucial transcription factor in the antioxidant defense system [90,91]. It can transcriptionally regulate the expression of various antioxidant genes, including those encoding SOD, CAT, GPX, and heme oxygenase 1 (HO-1) [90]. Previous studies have demonstrated that NFE2L2 is essential for mitigating oxidative damage caused by various toxic compounds, such as cadmium, aflatoxin B1 and cisplatin [3,92,93]. Consistently, Zhang et al. found that exposing mouse neuronal N2a cells to 5–80 ng/mL of T-2 toxin for 24 h can dose-dependently inhibit the expression of NFE2L2 protein and its downstream protein HO-1 [12]. In mice treated with 5.94 mg/kg of T-2 toxin, the mRNA levels of NFE2L2 and the phase II detoxifying enzymes NQO-1, GCLC, GCLM, and HO-1 were significantly decreased on the 1st, 3rd, and 7th days after percutaneous treatment [61]. In another study, it was found that treating SH-SY5Y cells with 5 or 10 ng/mL of T-2 toxin for 6 h significantly upregulated the expression of NFE2L2 protein [83]. Huang et al. found that treating GH3 cells with T-2 toxin also significantly increased the mRNA expression of the NFE2L2 gene [94]. These findings indicate that the expression of NFE2L2 in response to T-2 toxin-induced oxidative stress damage in neuronal cells is context dependent. Consistently, the pharmacological inhibition or gene knockout of NFE2L2 was shown to exacerbate T-2 toxin-induced neuronal cell death, confirming the survival-promoting role of NFE2L2 in T-2 toxin-induced cytotoxicity in neuronal cells [12,94].
Pei et al. showed that silencing the HMGB1 gene significantly inhibits the production of ROS and MDA, increases the activity of SOD, and then alleviates mitochondrial dysfunction and caspase -9, -3-dependent apoptosis [70]. Furthermore, it was found that silencing the HMGB1 gene significantly reduces the expression of the NFE2L2 protein in T-2 toxin-treated PC12 cells [70]. This indicates that HMGB1 may be a key target for intervening in T-2 toxin-induced neurotoxicity.
In summary, these studies indicate that T-2 toxin exposure can cause oxidative stress damage in nerve cells by inducing ROS and RNS production and downregulating the intracellular antioxidant system, including a decrease in antioxidant enzyme activity and antioxidant content. T-2 toxin exposure can also affect the NFE2L2 pathway, thereby regulating the intracellular antioxidant reduction system. The activation of NFE2L2 can protect against T-2 toxin-induced oxidative stress damage and neurotoxicity. As shown in Figure 2, the current evidence suggests that T-2 toxin-induced oxidative stress in neuronal cells is mainly due to lipid peroxidation, disruption of the body’s antioxidant system, the inhibition of peroxidase expression and the dysregulation of the NFE2L2 pathway.
5.2. Role of Mitochondrial Dysfunction and Apoptosis in T-2 Toxin Neurotoxicity
Mitochondria are both generators and targets of ROS [95,96]. Excessive ROS production can disrupt mitochondrial function and lead to cell death. Several studies have shown that an exposure to T-2 toxin can cause mitochondrial dysfunction, resulting in neuronal cell apoptosis [12,67,70,71,94]. Noticeable mitochondrial damage, such as mitochondrial swelling, vacuole formation, and a loss of cristae, has been observed in neuronal cells or brain tissues exposed to T-2 toxin [62,97]. Bin-Umer et al. reported that several trichothecenes, including T-2 toxin, can directly hinder mitochondrial translation [98]. Wan et al. observed that 40 nM of T-2 toxin reduces the expression of heat shock proteins Hsp60 and Hsp70 in rat GH3 cells [99]. Hsp60, a mitochondrial-specific chaperone protein, coordinates the import and folding of cytoplasmic proteins within mitochondria [100,101]. Importantly, the inhibition of Hsp70 impairs mitochondrial proteostasis and function [102]. Thus, the suppression of Hsp60/Hsp70 induced by T-2 toxin likely explains its disruptive effects on mitochondrial genomics (transcription/replication) and the translational mechanism. Consistently, it was also reported that an exposure to T-2 toxin can significantly increase the expression of critical transcription factors and co-activators that control mitochondrial biogenesis, mt-DNA transcription, and replication in GH3 cells. These include mitochondrial transcription factor A (mtTFA), mtTFB1, mtTFB2, estrogen-related receptor alpha (Err-α), nuclear respiratory factors 1 and 2 (NRF-1 and NRF-2), peroxisome proliferator-activated receptor gamma co-activator-related protein 1 (PPRC1), peroxisome proliferative-activated receptor gamma co-activator 1 alpha (PGC-1α), and PGC-1β [94]. In another study, it was reported that treatment with 0.5 ng/mL of T-2 toxin for 72 h or 120 h significantly reduces the expression of PPRC1, PGC-1α, nuclear respiratory factor 1 (NRF-1), and mitochondrial transcription factor A (mtTFA) in murine embryonic stem cells [103]. These different changes may depend on the treatment dose and time of T-2 toxin and the cell type.
The mitochondrial oxidative phosphorylation (OXPHOS) system is the metabolic center of eukaryotic cells, driving the synthesis of ATP. It has been shown that an exposure to T-2 toxin can significantly increase the activity of mitochondrial complex I and the expression of most mitochondrial ETC core subunits (including NADH dehydrogenase iron–sulfur protein 1 [Ndufs1], Ndufs3, Ndufs4, Ndufs6, Ndufs7, Ndufs8, NADH dehydrogenase flavoprotein 1 [Ndufv1], Ndufv2, Ndufv3, NADH dehydrogenase alpha subcomplex assembly factor 1 [Ndufaf1], Ndufaf2, Ndufaf3, Ndufaf4, NADH dehydrogenase subunit 6, mitochondrial [ND6], succinate dehydrogenase flavoprotein subunit A [Sdha], Sdhb, cytochrome c-1, heme protein [Cyc1], rieske iron-sulfur polypeptide 1 [Uqcrfs1], cytochrome c oxidase subunit 1, mitochondrial [COX1], COX2, COX3, ATP synthase F0 subunit 6, mitochondrial [mt-Atp6] and mt-Atp8) in GH3 cells [94]. Furthermore, it has been demonstrated that this upregulation may be due to the activation of nuclear respiratory factor 2α (NRF-2α) [104]. Notably, knocking down NRF-2α significantly reduces the expression of Ndufs3, Ndufs7, Ndufaf1, Ndufaf2, Ndufaf4, mtTFA, mtTFB2, and Drp1 mRNAs in T-2 toxin-treated GH3 cells, promotes the production of ROS, eliminates the increase in ATP and mitochondrial complex I activity induced by T-2 toxin and inhibits the mitochondrial DNA copy number [104]. This indicates that the activities of ETCs, mitochondrial biosynthesis, and mitochondrial dynamics mediated by NRF-2α play a protective role.
Mitochondria maintain the dynamic balance of the mitochondrial network through continuous division and fusion, a process known as mitochondrial dynamics. This is an important basis for maintaining mitochondrial morphology, distribution, and quantity and ensuring cellular homeostasis. In addition, the body degrades dysfunctional mitochondria inside the cell through mitochondrial autophagy, maintaining mitochondrial homeostasis [105]. Guo et al. found that exposing GH3 cells to 40 nM of T-2 toxin for 24 h significantly increases the mRNA expression of Drp1, Fis1, Mfn1, and Opa1 genes, which are involved in mitochondrial dynamics [104]. Additionally, treatment with T-2 toxin can activate mitophagy, as evidenced by the increased expression of the mitophagy-specific proteins NIP-like protein X (NIX), PTEN-induced putative kinase protein 1 (PINK1), and E3 ubiquitin ligase Parkin in GH3 cells [94]. Furthermore, it was found that knocking down PINK1 significantly promotes the production of ROS, the increase in ATP, and cell apoptosis induced by T-2 toxin [104]. This indicates that PINK1-mediated mitophagy plays a protective role in T-2 toxin-induced oxidative stress damage and cell apoptosis. Additionally, Huang et al. demonstrated that knocking down the NFE2L2 gene significantly blocks the expression of PINK1 [94]. This indicates that T-2 toxin-induced PINK1-mediated mitophagy is partly dependent on the activation of the NFE2L2 pathway.
Wan et al. showed that an exposure to T-2 toxin significantly reduces the gene and protein expression related to glycolysis and the TCA cycle in GH3 cells. These include pyruvate kinase isozymes M1/M2 (PKM1/2), malate dehydrogenase (MDH), aconitate hydratase (Aco2), isocitrate dehydrogenase (NAD) subunit α (IDH), and ATP citrate lyase. This suggests that an exposure to T-2 toxin can damage mitochondrial energy metabolism.
Huang et al. reported that exposing GH3 cells (a rat pituitary cell line) to a high dose of T-2 toxin (i.e., 40 nM for 24 h) can decrease mitochondrial membrane potential, reduce the expression of the Bcl-2 protein and increase the expression of the Bax protein, ultimately resulting in cell apoptosis [94]. Similarly, Zhang et al. reported that exposing N2a neuronal cells to 5–80 ng/mL of T-2 toxin can cause a decrease in mitochondrial membrane potential and the expression of the Bcl-XL protein and an increase in the expression of Bax proteins and the activity of caspases-9 and -3, finally leading to cell apoptosis in a dose-dependent manner [12]. Previous studies have shown that the activation of p53 can activate Bax, which then makes the mitochondrial membrane permeable and triggers apoptosis [12,94]. It was also reported that an exposure to T-2 toxin can activate the expression of the p53 protein in neuronal cells [12]. This indicates that the activation of the p53 pathway also partly contributes to mitochondrial dysfunction caused by T-2 toxin.
Zhang et al. also found that treatment with T-2 toxin significantly increases the activity of caspase-8 and reduces the expression of the pro-BH3 interacting domain death agonist (BID) protein [12]. Similar findings were also observed in T-2 toxin-induced toxicity in GH3 cells. It is well known that caspase-8 is a biomarker of the extrinsic apoptotic pathway (i.e., the death receptor apoptosis pathway) [12]. Therefore, the activation of the extrinsic apoptotic pathway caused by T-2 toxin exposure may also contribute to its neurotoxicity.
Overall, T-2 toxin exposure has been shown to trigger mitochondrial impairment and apoptosis in neuronal cells. This mitochondrial dysfunction is linked to disruptions in key processes such as mitochondrial biosynthesis, dynamics, mitophagy and electron transport chains. The resulting dysregulation of mitochondrial function can activate both intrinsic and extrinsic apoptotic pathways, ultimately leading to cell death. As illustrated in Figure 3, these events involve multiple signaling pathways, including PGC1α, intrinsic apoptotic, extrinsic apoptotic, p53, NRF-2α, and PINK1 pathways.
5.3. Role of Inflammatory Responses and Cell Pyroptosis
T-2 toxin elicits inflammatory reactions in the central nervous system [106]. Nakajima et al. reported that maternal T-2 toxin exposure in mice markedly boosts astrocyte activation in cerebral cortex tissue, evidenced by elevated GFAP and STAT3 protein levels. This suggests that STAT3-mediated transcriptional GFAP upregulation in cerebellar astrocytes is essential in T-2 toxin-mediated neurotoxicity [107]. In murine models, T-2 toxin administration also significantly increased metallothionein (MT)-I/II-positive cells in brain regions like the dentate gyrus hilus, cerebral cortex, corpus callosum and cerebellum at doses of ≥3 mg/kg or 9 mg/kg body weight post-weaning, reflecting oxidative stress and inflammatory induction [107]. Previous investigations revealed that T-2 toxin concentrations from 0.625 ng/mL to 10 ng/mL provoke cytotoxicity, excessive ROS generation, oxidative stress, mitochondrial impairment, autophagy induction and mitochondrial apoptotic pathway activation in BV2 microglial cells, with effects depending on dose and duration. This underscores the vulnerability of microglia to T-2 toxin [72]. Similarly, Weidner et al. observed a substantial cellular toxicity and apoptosis in human primary astrocytes at low T-2 toxin exposures (at 1 nM to 10 μM) [36]. Microglial activation by T-2 toxin in mice leads to cognitive deficits, and suppressing this activation alleviates learning and memory dysfunction [37]. These results imply that astrocytes are the primary targets of T-2 toxin, and their activation is fundamental to neurotoxicity mechanisms. Li et al. established that microglia activation induced by T-2 toxin involves MAPK and NF-κB pathway stimulation in BV-2 cells [37]. NF-κB, a central inflammatory regulator, governs the transcription of pro-inflammatory genes like IL-1β, COX-2, iNOS, IL-6, and TNF-α [108]. In resting states, NF-κB is bound in the cytoplasm by IκB proteins. Upon stimulation by inflammatory cytokines, IκB degradation enables NF-κB nuclear translocation and gene transcription initiation. This mechanism is vital in microglia, the CNS’s primary immune cells, where NF-κB upregulation in pathological states can worsen disease via cellular death and inflammation amplification [109]. In another study, it was documented that T-2 toxin treatment at 5 ng/mL for 24 h elevates the phosphorylation of NF-κB and IκBα proteins, subsequently upregulating COX-2, CD11B, and mRNA levels for IL-1β, iNOS, IL-6, and TNF-α in BV2 cells [37].
Additionally, T-2 toxin exposure robustly activates the MAPK pathway in microglia. In treated BV2 cells, phosphorylated JNK and Erk proteins increase significantly, but not p-p38. The pharmacological inhibition of JNK and Erk pathways suppresses T-2 toxin-induced TNF-α, COX-2, IL-6, and IL-1β mRNA expression, indicating their contribution to neuroinflammatory responses. MAPK pathways serve as upstream regulators of NF-κB signaling [110], and T-2 toxin’s impact on JNK and Erk may influence NF-κB, though the molecular details require further exploration. T-2 toxin also upregulates BTG2 mRNA and protein expression in mouse hippocampus, cortex tissues, and BV2 cells [84]. Knocking down BTG2 attenuates T-2 toxin-induced neuroinflammation in BV2 cells, as seen by the reduced IL-1β, IL-6, and TNF-α expression, highlighting BTG2’s key role in microglial activation [84]. Moreover, inhibiting BTG2 expression reverses T-2 toxin-mediated PI3K/AKT and NF-κB pathway activation in microglia. PI3K/AKT pathway inhibition with LY294002 decreases p-IκB-α and p-NF-κB levels, lowering inflammatory markers IL-1β, IL-6, and TNF-α [84]. This evidence reveals BTG2’s involvement in microglial activation through PI3K/AKT/NF-κB signaling. BTG2 participates in diverse physiological and pathological functions, including cell differentiation, proliferation, apoptosis, and tumor suppression [111]. It is regulated by p53, suggesting that T-2 toxin-induced p53 activation might modulate inflammation via BTG2, but this complex mechanism needs additional study [111].
Liu et al. demonstrated that T-2 toxin at 5–80 nM over 1–12 h downregulated AT-hook transcription factor (AKNA) expression in GH3 cells dose- and time-dependently [112]. AKNA is crucial for immune response, inflammation, development, cancer, autoimmunity and neurogenesis [113]. As a master regulator of inflammation, AKNA silencing reduces T-2 toxin-induced inflammatory cytokines such as TNF-α, IL-1β, and IL-6, implying its role in neuroinflammatory outcomes [112,113,114]. T-2 toxin also increases phospho-PKA/CREB expression. Blocking PKA, NF-κB, p38, and Erk activities using gene silencing or inhibitors enhances AKNA expression in T-2 toxin-treated GH3 cells, indicating that PKA/CREB, NF-κB, and MAPK pathways negatively regulate AKNA to control inflammation [112].
Pyroptosis, a type of programmed cell death that generates intense inflammation, has been linked to inflammatory damage in the brain [115]. The assembly of inflammasomes elicits the activation of caspase proteins. These activated caspases then promote the formation of the N-terminal pore-forming gasdermin D (GSDMD) fragment (i.e., GSDMD-NT) [116]. Subsequently, IL-1β and IL-18 mature and are released through the ruptured cell membrane, leading to pyroptosis activation. Pei et al. discovered that, when mice were orally treated with T-2 toxin at a dose of 0.5–2 mg/kg body weight for 28 days, it significantly increased the expression of caspase-1, cleaved-IL-1β, IL-18, and GSDMD proteins in the brain tissues [117]. Most pyroptosis processes are initiated by the NLRP3 inflammasome complex [90]. The endogenous host sensor activated by pathogens initiates macrophage pyroptosis through the deubiquitylation of NLRP3, resulting in GSDMD cleavage and the leakage of cellular contents [90]. So, it follows that the NLRP3 inflammasome serves as an important mediator between extracellular stimulation and pyroptosis. In addition to the NLRP3 protein, the NLRP3 inflammasome also consists of the ASC recruiting domain and caspase-1 protein [118]. Similarly, in HT22 neuronal cells, an exposure to T-2 toxin can upregulate the expression of high-mobility group B1 protein (HMGB1), NLRP3, ASC, caspase-1 and GSDMD-NT proteins. The pharmacological inhibition of GSDMD activation by dimethyl fumarate or the silencing of HMGB1 gene expression by SiRNA can significantly ameliorate the T-2 toxin-induced activation of the NLRP3/caspase-1 inflammasome, then reduce cell pyroptosis [67]. This suggests that T-2 toxin exposure can activate cell pyroptosis in a GSDMD-dependent manner, which may also involve the activation of the NLRP3/caspase-1 inflammasome pathway and the HMGB1 pathway. NF-κB acts as a promoter molecule and translocates into the nucleus to elicit the activation of NLRP3 inflammasomes [67]. Currently, it remains unclear whether the activation of the NLRP3 inflammasome is due to the activation of NF-κB caused by T-2 toxin.
The available research data suggest that T-2 toxin exposure can induce inflammatory responses in nerve tissues by activating the MAPK, AKNA, PKA/CREB, GSDMD, HMGB1, NF-κB and NLRP3 pathways (Figure 4). These findings offer crucial information for the targeted intervention of T-2 toxin-induced neuroinflammation and neurodegenerative diseases. The activation of these signaling pathways plays a pivotal role in the development of neuroinflammation, and understanding these mechanisms can help in formulating strategies to mitigate the harmful effects of T-2 toxin on the nervous system. Glial cells, which are known to be involved in neuroinflammatory processes, may also be affected by the activation of these pathways during T-2 toxin exposure. Further research is needed to fully understand the complex interplay between these signaling pathways, glial cells, and the overall process of T-2 toxin-induced neuroinflammation.
5.4. Role of Autophagy
Autophagy, a cellular degradation process, plays a crucial role in maintaining neuronal health by removing damaged organelles (including mitochondria and endoplasmic reticulum) and protein aggregates caused by DNA damage, hypoxia, nutrient deprivation, and oxidative stress [119]. Its dysfunction is increasingly recognized as a significant factor in the pathogenesis of various neurotoxicity or neurological diseases. It has been reported that the activation of autophagy is involved in the maintenance of neuronal homeostasis, particularly in response to oxidative stress and mitochondrial dysfunction induced by drugs and environmental toxins (such as colistin, methylmercury, and bupivacaine) [120,121,122]. In general, the fate of the cell depends on the interplay between pro-apoptotic factors and autophagy, wherein the latter blocks the induction of apoptosis or acts to delay apoptotic cell death; conversely, the activation of pro-apoptotic caspases shuts off autophagy. Notably, the formation of autophagosomes was observed using transmission electron microscopic examination in neuronal cells exposed to low doses of T-2 toxin [94]. Sun et al. reported that T-2 toxin treatment at 2.5–5 ng/mL significantly upregulated the expression of Beclin1 and LC3II proteins, promoted the formation of autophagosomes and autolysosomes and upregulated autophagy flux in BV2 cells [72]. Furthermore, Guo et al. demonstrated that the T-2 toxin-induced formation of autophagosomes and autolysosomes is partly dependent on the activation of autophagy-related gene 5 (ATG5) [104]. The inhibition of autophagy by chloroquine, a specific autophagy degradation inhibitor, significantly exacerbated T-2 toxin-induced caspase-3-dependent cell apoptosis [72]. This indicates that autophagy activation by T-2 toxin plays a protective role in the CNS.
A recent study showed that T-2 exposure in rats significantly downregulated the expression of ATG5 and mammalian target of rapamycin (mTOR) mRNAs on the 1st day, but significantly upregulated the expression of ATG5 and mTOR mRNA in brain tissue on the 3rd day post-T-2 exposure; the autophagy gene LC3B was significantly upregulated [49]. It is known that mTOR controls protein synthesis by activating S6 kinase 1 (S6K1) and 4E binding protein 1 and regulates autophagy via regulating the expression of ATG5 [123]. Consistently, Wu et al. showed that rapamycin, a special mTOR inhibitor, can markedly reduce T-2 toxin-induced apoptosis [94]. Additionally, T-2 toxin treatment also activated mitophagy, which was evident by the increased expression of mitophagy-specific proteins NIX, PINK1 and Parkin mRNA and proteins in GH3 cells, and the inhibition of PINK1-mediated mitophagy also promoted T-2 toxin-induced cell apoptosis.
In short, the current evidence indicates that T-2 toxin exposure can promote the formation of autophagosomes and autolysosomes and upregulate autophagy flux in nerve cells. It also induces mitophagy. These processes involve the mTOR, ATG5, and PINK1 pathways. Autophagy activation plays a protective role in T-2 toxin-induced cytotoxicity and apoptosis in nerve cells.
5.5. The Induction of Cell Cycle Arrest and Cellular Senescence
In neuronal cells, an exposure to T-2 toxin can induce cell cycle arrest and senescence [71,97,124]. For example, Agrawal et al. reported that exposing IMR-32 cells to 40 ng/mL of T-2 toxin for 8–24 h resulted in sub-G1 arrest and significantly elevated the mRNA levels of CDK2, CDK6, cyclin A and p21 [71]. Consistently, Fatima et al. discovered that treating GH3 cells with 40 nM of T-2 toxin significantly promoted G1 phase arrest, upregulated the expression of p16 and p21 proteins and downregulated the levels of cyclin D1, CDK4 and p-RB. Moreover, this process might be partly regulated by the p53 and MAPK pathways [71,97].
T-2 toxin treatment was demonstrated to induce cell cycle arrest and senescence in SH-SY5Y neuronal cells via upregulating the expression of p16, p21, and p53 and enhancing the activity of SA-β-gal [125]. Inhibiting HIF-α expression significantly decreased the expression of p53, p21 and p16 proteins; reduced the increased SA-β-gal activity; and ultimately alleviated T-2 toxin-induced neuronal cell cycle arrest and senescence. This implies that T-2 exposure-induced HIF-α activation partly contributes to cell cycle arrest and senescence in neuronal cells [125]. Additionally, T-2 toxin exposure can alter the expression of Alzheimer’s disease-related proteins, such as Tau, phosphorylated Tau (p-Tau), and amyloid precursor protein, indicating a significant association between T-2 toxin exposure and the development of neurodegenerative diseases [125].
5.6. Imbalance of Gut Microbiota
Research has shown that short-chain fatty acids (SCFAs) impact the cellular system and interact with gut–brain signaling pathways. Specifically, they can regulate immune, endocrine, neural, and humoral pathways, reduce inflammatory factors, activate the vagus nerve, and protect neurons against damage induced by environmental toxins via the gut–brain axis [126]. For example, Wu et al. reported that a dietary intake of 1 mg/kg T-2 toxin increased the enrichment of g_norank_f_T34, Faecalibacterium, and f_Clostridium_methylpentosum_group, while significantly reducing the levels of acetic acid, propionic acid, butyric acid and total SCFAs in the intestinal tissues of weaned piglets [127]. Su et al. showed that T-2 toxin exposure significantly increased the abundance of Enterobacteriaceae and decreased the abundance of Lactobacillus in the intestinal tissues of mice [128]. It was demonstrated that the increased Shigella abundance in the intestinal tissues induced by T-2 toxin is positively associated with inflammatory markers (e.g., TNF-α and IL-6) in the hippocampal tissues. Reversing these changes in gut microbiota can significantly improve T-2 toxin-induced neuroinflammation and neuro-memory dysfunction [128], indicating that the T-2 toxin-induced disruption of gut microbiota is involved in regulating neuroinflammatory responses and neurobehavioral disorders. Huang et al. reported that T-2 toxin disturbed the composition of gut microbiota, particularly Faecalibaculum and Allobaculum, which were positively correlated with anorexia and the levels of serum cholecystokinin, glucagon-like peptide-1, 5-HT, IL-1β, IL-6 and TNF-α [38]. These data suggest that T-2 toxin-induced changes in gut microbiota may partly contribute to its induction of neuroinflammation and anorexia. However, to date, the precise molecular mechanisms remain unclear, and more experimental evidence and further studies are needed.
6. Chemo-Protective Agents for T-2 Toxin-Induced Neurotoxicity
It was reported that several antioxidants, natural products and specific small-molecule inhibitors can effectively mitigate T-2 toxin-mediated neurotoxicity by simultaneously targeting oxidative stress, inflammatory cascades, and programmed cell death pathways (as shown in Table 2).
6.1. Antioxidants
GSH, a crucial tripeptide that regulates cellular detoxification, antioxidant defense, thiol homeostasis, and proliferation, is biosynthesized in the cytosol under strict control. Experimental research has repeatedly shown that T-2 exposure significantly depletes neuronal GSH reserves at both the tissue and cellular levels [12,74,88]. Due to the limited membrane permeability of exogenous GSH, oral supplementation is ineffective for direct intracellular delivery. NAC primarily increases GSH levels by providing the amino acid L-cysteine, which is the rate-limiting substrate for de novo GSH synthesis [129,130]. Zhang et al. reported that NAC supplementation can notably increase intracellular GSH levels, thereby significantly reducing T-2 toxin-induced caspase-mediated neuronal cell apoptosis [12]. Sun et al. found that NAC supplementation can markedly lower intracellular ROS levels and alleviate T-2 toxin-induced cytotoxicity in BV-2 cells [72]. Moreover, NAC supplementation can also significantly relieve T-2 toxin-induced cell pyroptosis by inhibiting the NLRP3 pathways [131], which may be attributed to its radical scavenging ability. Studies in animal models have indicated that NAC supplementation can effectively improve the clinical symptoms or pathological processes of neurodegenerative diseases or neurotoxicity induced by environmental toxins through the targeted inhibition of oxidative stress or inflammatory responses in neuronal cells [132,133,134,135,136]. This is in line with recent clinical trials that have shown that oral NAC supplementation can notably reduce neurotoxicity caused by chemotherapeutic agents (e.g., paclitaxel or oxaliplatin) in cancer patients [137,138].
Vitamin E, also known as alpha-tocopherol, is a lipid-soluble vitamin that serves as a potent antioxidant against oxidative damage. It was reported that vitamin E supplementation at 100 mg/kg body weight can significantly inhibit the production of ROS and MDA, upregulate the activities of GPX and GSH levels, and reduce the levels of IL-1β and TNF-α mRNAs in order to markedly attenuate T-2 toxin-induced mouse brain damage [74]. Notably, in the brain tissues, the transport of BBB is regulated by several transporters, such as α-tocopherol (binding) transport protein, phospholipid transfer protein, and scavenger receptor class B type 1, whose kinetics are quite slow in mammals [139]. This is the key reason for the lower levels of vitamin E in brain tissues compared to other organs (such as the liver) during dietary supplementation [139]. This a key limitation for vitamin E application in clinical practice.
In addition, several antioxidants such as melatonin, vitamin C, silymarin, selenium, L-carnitine, alpha-tocopherol, and coenzyme Q10 (CoQ10) have been demonstrated to have protective effects against T-2 toxin-induced cytotoxicity and tissue damage via scavenging radicals [127,140,141,142,143,144,145]. Clinical trials have also confirmed that supplementation with these antioxidants can effectively improve cognitive impairment and anorexia caused by anti-cancer drugs [146,147,148]. Therefore, it is worthwhile to further explore whether these antioxidants can effectively improve T-2 toxin-induced neurobehavioral defects or neurotoxicity, and more clinical trial studies are required.
6.2. Small-Molecule Inhibitors
Several small-molecule inhibitors, including PDTC, SP600125, PD98059, dimethyl fumarate, VX-765, YC-1, and LY294002, can target the NF-κB, JNK, Erk, GSDMD, caspase-1, HIF-1, and PI3K/Akt pathways, respectively, to inhibit neuroinflammatory responses, mitochondrial dysfunction, cell apoptosis, and pyroptosis caused by T-2 toxin exposure in neuronal or glial cells [37]. For instance, VX-765 and dimethyl fumarate can significantly inhibit the expression of caspase-1 and GSDMD-NT proteins in HT22 cells, respectively, thereby attenuating T-2 toxin-induced neuroinflammatory responses and neuronal pyroptosis [67]. Li et al. demonstrated that PDTC, SP600125, and PD98059 can markedly inhibit the NF-κB, JNK, and Erk pathways, respectively, and subsequently reduce microglial activation, as evidenced by the decreased expression of TNF-α, IL-6, COX-2, and IL-1β mRNA in BV-2 cells [37]. Zhao et al. reported that YC-1 supplementation can effectively inhibit the upregulation of HIF-1α induced by T-2 toxin. It then significantly downregulates intracellular MDA and ROS levels, while upregulating the expression of APP and p-Tau proteins in BV-2 cells via activating the JNK pathway. Inhibiting JNK can mitigate the adverse effects of YC-1 treatment, suggesting that HIF-1α may have a dual role, and co-targeting HIF-1α and JNK could be a new strategy to improve T-2 toxin-induced neurotoxicity [149]. Li et al. found that LY294002’s targeted inhibition of PI3K/Akt in BV-2 cells, then significantly inhibited the activation of NF-κB and the expression of its downstream genes, such as IL-1β, IL-6, and TNF-α mRNAs [84]. Additionally, 4-phenylbutyrate (4-PBA), an ER stress inhibitor, can effectively attenuate T-2 toxin-induced cytotoxicity and apoptosis in porcine renal epithelial cells [150] and goat endometrium epithelial cells [151]. Several clinical trials have shown that 4-PBA supplementation has potential therapeutic effects on inflammatory or neurological disorder-related diseases [152,153,154]. This information indicated that 4-PBA may be a promising candidate drug against T-2 toxin-induced neurotoxicity in animals and humans. Recently, Chen et al. reported that the local administration of AHN 1-055 hydrochloride, a dopamine transporter inhibitor, can significantly alleviate T-2 toxin-induced depression-like symptoms in mice [65]. Similarly, knocking down the dopamine transporter can produce obvious antidepressant like effects in a mouse model [155].
6.3. Natural Products
Plant natural products such as flavonoids, polysaccharides and peptides can improve various neurodegenerative diseases (e.g., cerebral ischemia, stroke, and Alzheimer’s disease) or neurotoxicity induced by environmental toxins or drugs [90,156,157]. These agents achieve this neuroprotective effect by alleviating oxidative stress, regulating cytokine levels and autophagy flow, and inhibiting excitotoxicity.
It has been reported that certain natural products such as saffron, daucosterol, resveratrol, quercetin, and betulinic acid have strong antioxidant, anti-inflammatory, and immune-regulatory activities by targeting NFE2L2, PGC-1α, NF-κB, NLRP3, and MAPK pathways. They exhibit potential protective effects against neurotoxicity caused by several mycotoxins [128,158,159,160]. For example, Gu et al. reported that the intraperitoneal injection of daucosterol (a phytosterol glycoside found in many plants) at 30 mg/kg body weight can significantly reduce damage to the BBB, hippocampal neuronal degeneration, and neuroinflammatory response in the brain tissues of mice exposed to T-2 toxin by specifically activating the PGC-1α protein [85]. Huang et al. reported that supplementing betulinic acid at 0.25–1 mg/kg body weight per day for fourteen days can significantly improve cognitive function, antioxidant capacity, and brain neurotransmitter (i.e., Ach, 5-HT, and DA) levels and inhibit apoptosis and the secretion of pro-inflammatory cytokines in brain tissues. It offers a protective effect against brain damage caused by T-2 toxin in a mouse model by specifically regulating NFE2L2, MAPK, and NF-κB pathways [74]. Moreover, it was shown that betulinic acid supplementation can significantly reduce T-2 toxin-induced cytotoxicity, hepatotoxicity, nephrotoxicity, enterotoxicity, spleen toxicity, and lymphotoxicity in vitro and in vivo by regulating the aforementioned pathways [161,162,163,164,165,166,167]. This indicates that betulinic acid supplementation can not only alleviate T-2 toxin-induced neurotoxicity but also systematically improve the toxic effects of T-2 toxin on different organs. Su et al. reported that an oral supplementation of resveratrol at 100 mg/kg can effectively reduce T-2 toxin-induced cognitive deficits, neuronal damage, and disruption of the gut–brain axis. The potential mechanisms may involve the restoration of Lactobacillus abundance and gut barrier integrity, thereby reduce systemic inflammation and restoring hippocampal synaptic protein expression and function in mice through gut–brain axis modulation [128]. A clinical trial further confirmed that resveratrol supplementation (up to 1 g orally twice daily) for 52 weeks can significantly treat or prevent neurodegenerative disorders in patients with Alzheimer’s disease [168,169]. These findings highlight the therapeutic potential of these compounds in dealing with oxidative stress- and inflammation-related neurotoxicity, providing promising directions for future research and clinical applications to diminish and prevent T-2 toxin-induced neurological disorders.
6.4. Other Neuroprotective Agents
Minocycline, a broad-spectrum tetracycline antibiotic, was recently found to exert broad protective effects in a multitude of neurological disorders, including spinal cord injury, stroke, multiple sclerosis, amyotrophic lateral sclerosis (ALS), Huntington’s disease, epilepsy, and PD, via inhibiting neuroinflammation [170,171,172].
It was reported that minocycline administration at 50 mg/kg body weight markedly reduced T-2 toxin exposure-induced microglia activation, downregulated the expression of inflammatory markers, including TNF-α, IL-1β, IL-6, COX-2, CD11b mRNAs or proteins in the brain tissues of mice, and finally improved learning, memory impairment, and locomotor inhibition in mice [37]. These findings highlight the utility of minocycline as a neuroprotective agent against T-2 toxin-induced neurotoxicity.
7. Conclusions and Future Perspectives
Neurotoxicity is a significant adverse effect resulting from exposure to T-2 toxin. Animal experiments have revealed that a continuous exposure to T-2 toxin allows the toxin to penetrate the BBB, then accumulate and damage CNS tissues, finally inducing a decline in learning, memory, cognition, and motor function. T-2 toxin-induced neurotoxicity is related to multiple detrimental cellular mechanisms, including oxidative stress, mitochondrial dysfunction, neuroinflammation, autophagy, ferroptosis and apoptosis. These cellular processes are in turn driven by signaling pathways, such as NFE2L2, NRF-2, PGC-1, p53, BTG2, AKNA, MAPK, Akt, mTOR, HMGB1, HIF-1, CREB, and NF-κB. T-2 toxin exposure can also disrupt the synthesis and release of various neurotransmitters, including DA, GABA, glutamate, norepinephrine, 5-HT and Ach. Additionally, T-2 toxin exposure can also induce the disruption of gut microbiota, which contributed to the induction of neuroinflammatory responses and neurobehavioral disorders. Accumulating evidence demonstrates that several specific antioxidants (such as NAC, vitamin E, vitamin C, silymarin, selenium, L-carnitine, alpha-tocopherol, and coenzyme Q10), natural products (such as betulinic acid, resveratrol, and daucosterol), small-molecule inhibitors (such as DTC, SP600125, PD98059, dimethyl fumarate, VX-765, YC-1, LY294002, and AHN 1–055 hydrochloride), and FDA-approved drugs (i.e., minocycline) can effectively mitigate T-2 toxin-mediated neurotoxicity via the inhibition of oxidative stress, the inactivation of inflammatory cascades, the reduction of programmed cell death pathways, and the reconstruction of gut microbiota. Notably, human clinical trials have suggested that supplementation with minocycline and resveratrol can improve both neuropathological injury and abnormal changes in neurobehavior. Minocycline and resveratrol may be considered as the promising candidate therapeutic drugs for T-2 toxin exposure-induced neurotoxicity in animals and humans, albeit more animal model evaluations and clinical studies are still needed.
To develop effective policy and regulatory implications for improving or treating T-2 toxin-induced neurotoxicity in humans and animals, several directions for future research are warranted:
(i) It is crucial to investigate the relationships between T-2 toxin at the environmental exposure dose and neurological disorders in animals and humans, as well as the underlying molecular mechanisms. Currently, there is still a dearth of evidence on the long-term health effects of T-2 toxin exposure in humans, especially for the nervous system and its contribution to the occurrence and the development of neurodegenerative diseases. We need to establish dose correlations between chronic low-level T-2 toxin exposure via diet and clinically relevant neurological disorders, such as anorexia, depression, and neurodegenerative diseases. Additionally, new knowledge relating to the underlying molecular mechanisms of T-2 toxin-induced neurotoxicity remains unclear. For example, miRNAs have been proven to play critical roles in brain injury and the development of neurological disorders, but their role in T-2 toxin-induced neurotoxicity is poorly understood. The application of emerging technologies such as single-cell sequencing, CRISPR-Cas9 and multi-omics may also help elucidate the underlying mechanisms.
(ii) The risk assessment of T-2 toxin exposure in human samples is too limited, and further research should be conducted in the future, which is of great significance for public health safety and risk assessment.
(iii) A co-exposure risk assessment for T-2 toxin with other environmental toxins should be especially noted. It has been reported that the combination exposure between T-2 toxin and NIV, HT-2 toxin or DON showed a marked synergistic toxic effect [173,174,175]. Therefore, it is necessary to assess the neurotoxic risk caused by the combined exposure of T-2 toxin and other coexisting environmental toxins, such as heavy metals or other toxins.
(iv) The precise molecular mechanisms by which the above-mentioned neuroprotectants operate against T-2 toxin-induced neurotoxicity remain largely unknown, especially their effects on the changes in various neurotransmitters. Additionally, human clinical trials and animal experiments are still required to fully substantiate the neuroprotection conferred by these protective agents. A synergistic policy framework combining preventive agriculture, technological innovation, global standardization and consumer empowerment is critical to curb T-2 toxin exposure.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ayelign A. De Saeger S. Mycotoxins in Ethiopia: Current status, implications to food safety and mitigation strategies Food Control 202011310716310.1016/j.foodcont.2020.107163 · doi ↗
- 2Mukhtar K. Nabi B.G. Ansar S. Bhat Z.F. Aadil R.M. Mousavi Khaneghah A. Mycotoxins and consumers’ awareness: Recent progress and future challenges Toxicon 202323210722710.1016/j.toxicon.2023.10722737454753 · doi ↗ · pubmed ↗
- 3Dai C. Tian E. Hao Z. Tang S. Wang Z. Sharma G. Jiang H. Shen J. Aflatoxin B 1 Toxicity and Protective Effects of Curcumin: Molecular Mechanisms and Clinical Implications Antioxidants 202211203110.3390/antiox 1110203136290754 PMC 9598162 · doi ↗ · pubmed ↗
- 4Dai C. Sharma G. Liu G. Shen J. Shao B. Hao Z. Therapeutic detoxification of quercetin for aflatoxin B 1-related toxicity: Roles of oxidative stress, inflammation, and metabolic enzymes Environ. Pollut.202434512347410.1016/j.envpol.2024.12347438309422 · doi ↗ · pubmed ↗
- 5Dai C. Das Gupta S. Wang Z. Jiang H. Velkov T. Shen J. T-2 toxin and its cardiotoxicity: New insights on the molecular mechanisms and therapeutic implications Food Chem. Toxicol.202216711326210.1016/j.fct.2022.11326235792220 · doi ↗ · pubmed ↗
- 6Mhlongo T.N. Ogola H.J.O. Selvarajan R. Sibanda T. Kamika I. Tekere M. Occurrence and diversity of waterborne fungi and associated mycotoxins in treated drinking water distribution system in South Africa: Implications on water quality and public health Environ. Monit. Assess.202019251910.1007/s 10661-020-08477-x 32671553 · doi ↗ · pubmed ↗
- 7Eskola M. Kos G. Elliott C.T. HajšlováJ. Mayar S. Krska R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25Crit. Rev. Food Sci. Nutr.2020602773278910.1080/10408398.2019.165857031478403 · doi ↗ · pubmed ↗
- 8Xu H. Wang L. Sun J. Wang L. Guo H. Ye Y. Sun X. Microbial detoxification of mycotoxins in food and feed Crit. Rev. Food Sci. Nutr.2022624951496910.1080/10408398.2021.187973033663294 · doi ↗ · pubmed ↗