A Review of Gut Microbiota Dynamics: From Healthy Gestation to Gestational Diabetes in Human and Mouse Models
Dat Da Ly, Bryony A. McNeill, Kathryn Aston-Mourney, Leni R. Rivera

TL;DR
This review explores how gut microbiota changes during pregnancy and how these changes may contribute to gestational diabetes, highlighting differences from type 2 diabetes and the need for further research.
Contribution
The paper provides a comprehensive synthesis of gut microbiota dynamics during healthy and diabetic pregnancies, identifying pregnancy-specific microbial signatures.
Findings
Late pregnancy is marked by increased lactic acid-producing bacteria and reduced Firmicutes-to-Bacteroidetes ratios.
Gestational diabetes shows dysbiosis with elevated F/B ratios and reduced Bifidobacterium.
GDM butyrate-producer patterns differ from type 2 diabetes, indicating unique pregnancy-specific mechanisms.
Abstract
Over the past decades, gut microbiota has emerged as a critical modulator of human health and disease. Pregnancy involves substantial microbiota remodelling that influences offspring development, yet mechanisms linking maternal microbiota changes to gestational diabetes mellitus (GDM) remain unclear. The current literature lacks a comprehensive synthesis of pregnancy microbiota dynamics across healthy gestation to GDM, comparative human–murine analysis, and pregnancy-specific mechanistic frameworks distinct from type 2 diabetes models. This narrative review comprehensively synthesised evidence on gut microbiota composition in healthy pregnancy and GDM (2005–2025, NCBI PubMed) to identify convergent signatures and articulate pregnancy-specific mechanisms. Early pregnancy microbiota resembles non-pregnant individuals, whereas late pregnancy exhibits increased lactic acid-producing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
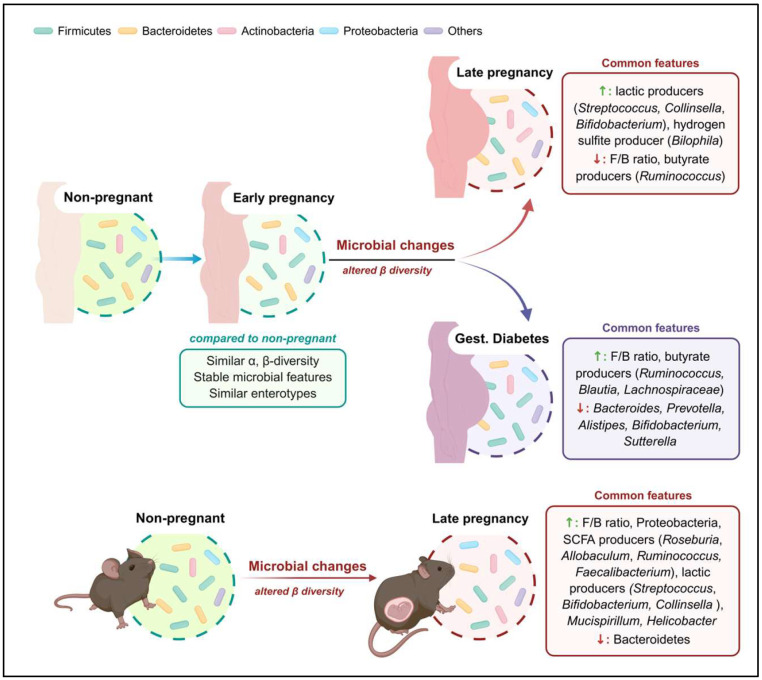 Figure 1
Figure 1| Year | Country | Sequencing |
| Gestational Week | Status | Microbial Composition |
|---|---|---|---|---|---|---|
| 2012 | Finland | 16S rRNA (V1-V2) | 91 | ~13.84 | 2 * | Relative abundance: F ~22–88%, B ~5–70%, A ~5%, P ~1% |
| 2016 | Australia | 16S rRNA (V6-V8) | 44 | 16 | 3 * | Relative abundance: F ~58–81% |
| 2017 | Finland | 16S rRNA | 60 | ~12.9 | 3 * | Relative abundance: F ~43.2%, B ~50.5%, A ~1.2%, P ~3.2% |
| 2019 | Israel | 16S rRNA (V4) | 35 | 12–14 | 1 * | Abundant genera: |
| 2020 | Finland | Metage- | 203 | ~13.9 | 3 * | Relative abundance: F ~37.9%, B ~56.9%, A ~1.7%, P ~2.5% |
| 2020 | China | 16S rRNA (V3-V4) | 103 | ~9 | 1 * | Relative abundance: F ~40%, B ~50%, A ~2%, P ~3% |
| 2020 | China | 16S rRNA (V4) | 98 | 10–15 | 1 * | Relative abundance: F ~45%, B ~47%, A ~2%, P ~5% |
| 2020 | China | 16S rRNA (V3-V4) | 45 | Trimester 1 | 1 * | Relative abundance: F ~47–56%, B ~36–45%, A ~1–3%, P ~2–4% |
| 2022 | Czechia | 16S rRNA (V3-V4) | 22 | Trimester 1 | 1 * | Abundant class: Clostridia, Bacilli, Negativicutes, Bacteroidia, γ-Proteo-bacteria |
| 2023 | Malaysia | 16S rRNA (V3-V4) | 12 | Trimester 1 | 1 * | Relative abundance: F ~47%, B ~43%, A ~2%, P ~7% |
| 2023 | China | Whole genome | 120 | ~10.2 | 1 * | Relative abundance: F ~40%, B ~55%, A ~2.3%, P ~2% |
| 2024 | New Zealand | 16S rRNA (V3-V4) | 27 | 12 | 1 * | Relative abundance: F ~52%, B ~39%, A ~2.3%, P ~3.1% |
| Year | Country | Sequencing |
| Gestational Week | Status | Microbial Alterations |
|---|---|---|---|---|---|---|
| 2008 | Finland | Targeted qPCR * | 36 | 30–35 | 1 * | |
| 2010 | Spain | Targeted qPCR * | 34 | 24 | 1 * | |
| 2012 | Finland | 16S rRNA (V1-V2) | 91 | 33.72 | 2 * | |
| 2013 | Norway | Sanger | 87 | 30–40 | 1 * | |
| 2014 | Finland | Metage- | 203 | ~35.2 | 3 * | |
| 2017 | China | Whole | 81 | 21–29 | 1 * | Abundant genera: |
| 2018 [ | Brazil | 16S rRNA (V4) | 42 | ~33.89 | 1 * | Abundant genera: |
| 2018 [ | Denmark | 16S rRNA (V1-V2) | 157 | ~28.4 | 1 * | Abundant genera: |
| 2019 [ | China | 16S rRNA (V3-V4) | 11 | ~32,7 | 1 * | Abundant genera: |
| 2019 [ | China | Whole | 26 | ~40.15 | 1 * | Abundant genera: |
| 2019 [ | China | 16S rRNA (V3-V4) | 16 | ~25.9 | 1 * | Abundant genera: |
| 2019 | Israel | 16S rRNA (V4) | 35 | 34–36 | 1 * | |
| 2020 | China | 16S rRNA (V3-V4) | 103 | ~24 | 1 * | |
| 2020 [ | China | 16S rRNA (V3-V4) | 31 | ~38.5 | 1 * | Abundant genera: |
| 2023 [ | China | 16S rRNA (V4) | 41 | 24–28 | 1 * | Abundant genera: |
| 2023 [ | China | 16S rRNA (V3-V4) | 9 | Full | 1 * | |
| 2023 | China | Whole genome | 120 | ~24 | 1 * | |
| 2023 [ | China | 16S rRNA (V4) | 38–42 | T3 | 1 * | |
| 2023 | Malaysia | 16S rRNA (V3-V4) | 12 | Trimester 3 | 1 * | |
| 2024 [ | China | 16S rRNA (V4) | 31 | 32–34 | 1 * | Abundant genera: |
| 2024 | Austria, | Varied | 2564 (before batch effect removal) | Trimester 3 | 2 * | |
| 2024 | New | 16S rRNA (V3-V4) | 27 | 24 | 1 * | |
| 2024 [ | China | 16S rRNA (V3-V4) | 30 | ~18 | 1 * |
| Year | Country | Sequencing | GDM | Gestational Week | Healthy Ctrl | Microbial Alterations |
|---|---|---|---|---|---|---|
| 2012 [ | Finland | 16S rRNA (V1-V2) | 15 | 13.84 | 76 | |
| 2017 [ | China | Whole genome | 43 | 21–29 | 81 | |
| 2017 [ | Finland | 16S rRNA | 15 | ~12.9 | 60 | |
| 2018 [ | China | 16S rRNA (V3-V4) | 26 | Full term | 23 | |
| 2018 [ | Brazil | 16S rRNA (V4) | 26 | ~33.89 | 42 | |
| 2018 [ | Denmark | 16S rRNA (V1-V2) | 50 | ~28.4 | 157 | |
| 2019 [ | China | 16S rRNA (V3-V4) | 11 | ~31.2 | 11 | |
| 2019 [ | China | Whole genome | 23 | ~39.1 | 26 | |
| 2019 [ | China | 16S rRNA (V3-V4) | 24 (successful control) | ~26 | 16 | |
| 12 (fail to control) | ~25.2 | 16 | ||||
| 2020 [ | Finland | Metage- | 53 | ~35.2 | 203 | |
| 2020 [ | China | 16S rRNA (V3-V4) | 31 | ~8 | 103 | |
| 31 | ~24 | 103 | ||||
| 2020 [ | China | 16S rRNA (V3-V4) | 30 | ~38.3 | 31 | |
| 2020 [ | China | 16S rRNA (V3-V4) | 45 | 24–28 | 45 | |
| 2021 [ | China | 16S rRNA (V3-V4) | 23 | ~35.26 | 29 | |
| 2021 [ | Australia | 16S rRNA (V6-V8) | 29 | ~27.7 | 29 | |
| 2021 [ | Thailand | _ * | 28 (successful | ~24.75 | 38 | |
| ~38.43 | 38 | |||||
| 13 (fail to control) | ~25.23 | 38 | ||||
| ~38.23 | 38 | |||||
| 2022 [ | Israel | 16S rRNA (V4) | 28 | 11–14 | 236 | |
| 2022 [ | Czechia | 16S rRNA (V3-V4) | 29 (early | T1 | 22 | |
| 31 (late | T1 | 22 | ||||
| 22 (late | T1 | 22 | ||||
| 2023 [ | China | 16S rRNA (V4) | 49 | 24–28 | 41 | |
| 49 | ~39 | 39 | ||||
| 2023 [ | China | Whole genome | 120 | ~10.2 | 120 | |
| 120 | ~23.9 | 120 | ||||
| 120 | ~33.6 | 120 | ||||
| 2024 [ | Italy, | Varied | 570 | 6–40 | 10378 |
| Year | Country | Sequencing |
| Gestational day | Compared to | Microbial Alterations |
|---|---|---|---|---|---|---|
| 2015 [ | Canada | 16S rRNA (V3) | 5 C57BL/6J mice | Post-conception | Pre-conception | |
| 2016 [ | Saudi | 16S rRNA (V4) | 6 Sprague- | 20 | 6 Non-preg-nant | |
| 2017 [ | USA | 16S rRNA (V4) | 5 C57BL/6 mice | Late | Early/ | |
| 2018 [ | The Netherlands | Phylogenetic microarray (GI chip) | 5 B6 mice | 18 | 5 Non-pregnant | |
| 5 | 18 | 5 Non-pregnant | ||||
| 2018 [ | Canada | Targeted qPCR * | 19–20 C57BL/ | Full term | Pre-conception | |
| 2019 [ | China | 16S rRNA (V4) | 12 MRL mice | Postnatal | 12 Non-pregnant | |
| 2019 [ | Israel | 16S rRNA (V4) | 10 Swiss | 18 | E0 | |
| 2019 [ | USA | 16S rRNA (V4) | 2 C57BL/6 (3–5 foetus per dam) | 17–18, | E14–16 | |
| 2020 [ | China | 16S rRNA (V3-V4) | 6 C57BL/6 | 18 | 6 Pre-pregnant | |
| 2020 [ | The Netherlands | 16S rRNA (V3-V4) | 12 C57BL/6J-OlaHsd mice | 18 | Pre-pregnant | |
| 2023 [ | Spain | 16S rRNA (V3-V4) | 9 C3H mice | 12.5 | 9 Non-pregnant | |
| 2023 [ | Japan | 16S rRNA (V3-V4) | 6 Avy × | 18 | E11 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Gestational Diabetes Research and Management · Reproductive tract infections research
1. Introduction
The human gastrointestinal (GI) tract provides a massive interface (250–400 m^2^) between the internal body and external environmental factors [1]. It processes approximately 1–2.5 kg of food daily, breaking it down into absorbable units whilst simultaneously functioning as a barrier against pathogenic invasion [2,3]. These functions are conducted in collaboration with about 100 trillion microorganisms within the gut, collectively known as the gut microbiota [4], which play diverse roles in a range of processes including food and xenobiotic metabolism, antimicrobial protection, and immunomodulation [5].
The gut microbiota is a rich ecosystem that has co-evolved in the human body symbiotically [6,7]. Its primary establishment commences at birth and is shaped by multiple factors including maternal health status, delivery mode, breastfeeding practice, maternal diet, antibiotics and early environmental exposures [8,9]. This early microbial composition established during infancy appears to influence host metabolism and immune development throughout life [8,9,10,11]. Disruptions in normal infant microbial colonisation, termed dysbiosis, have been associated with increased risk of both intestinal disorders and longer-term metabolic and neurological conditions, including type 1 diabetes, obesity, asthma and autism spectrum disorders [12,13,14,15,16,17]. Given that maternal microbiota directly influences infant gut colonisation and has demonstrable long-term health consequences [8,9,10,11], investigations of maternal microbiota dynamics during pregnancy have become increasingly important.
Microbiota research during pregnancy is critical for both metabolically healthy women and those with pregnancy complications such as diabetes. Understanding how diabetes during pregnancy affects the gut microbiota is particularly timely given concerning epidemiological trends showing that, despite public health efforts, the prevalence of pre-existing diabetes is expected to increase due to advancing maternal age at conception, combined with rapidly rising global incidence of type 2 diabetes (T2D) (affecting more than 530 million adults in 2024) [18] and early-onset T2D (diagnosed before age 40) [19]. Meanwhile, the rate of gestational diabetes mellitus (GDM) also continues to rise—currently affecting about 18% of pregnancies in Australia and 14% worldwide [20,21]. Both pre-existing diabetes and GDM are known to increase the likelihood of infants having an increased risk of obesity and type 2 diabetes later in life [22,23]. These conditions are also associated with gut dysbiosis, which has shown significant links to health outcomes not only in infancy but also well into adulthood [24,25,26,27]. Despite the critical health implications of understanding the pregnancy microbiome, variations in geographic regions, dietary habits, gestational stages, sequencing depth and resolution have limited our understanding of the microbial profiles of pregnant individuals, either with or without diabetes.
In non-pregnant adults, studies exploring the relationship between metabolic syndrome and gut microbiome have expanded therapeutic opportunities and clinical significance. A meta-analysis of 15 randomised controlled trials demonstrated that probiotic supplementation significantly reduced HbA1c, fasting blood glucose, and HOMA-IR (homeostatic model assessment for insulin resistance) compared to placebo in individuals with T2D [28]. Another meta-analysis of 14 randomised controlled trials showed that combining metformin with probiotics produced greater reductions in fasting blood glucose and HbA1c, and lower gastrointestinal side effects, compared to metformin monotherapy alone in adults aged 45–65 with T2D [29]. More directly demonstrating causality, allogenic faecal microbiota transplantation from lean donors to male recipients with metabolic syndrome significantly improved insulin sensitivity and increased colonisation by commonly recognised beneficial bacteria such as Akkermansia and short chain fatty acid (SCFA)-producing Eubacterium within 6 weeks [30,31]. In animal studies, Backhed and colleagues discovered that the colonisation of Clostridium scindens on germ-free female mice increased deoxycholic acid, which was positively correlated with HOMA-IR, HbA1c, triglycerides, and negatively associated with high-density lipoprotein, indicating impaired glucose and lipid metabolism [32]. Additionally, the transfer of unfractionated cecal microbiota—predominantly containing opportunistic bacteria Bacteroides and Clostridium—from conventionally raised to germ-free mice induced body fat accumulation and insulin resistance within 2 weeks, despite reduced food intake [33]. These findings directly demonstrate that dysbiotic microbiota composition can drive metabolic dysfunction independent of dietary intake, highlighting the pathogenic potential of specific bacterial taxa on host metabolism.
Despite the well-established role of the gut microbiome in metabolic health and disease pathogenesis, substantial evidence gaps limit clinical translation and precision therapeutic strategies in pregnancy. The current literature lacks: (1) comprehensive synthesis of pregnancy microbiota dynamics across the full spectrum from healthy gestation to gestational diabetes, integrating both physiological adaptations and pathological remodelling; (2) comparative analysis of human and murine pregnancy microbiota, which would bridge translational mechanistic research with clinical insights; (3) identification of converging microbiota signatures across heterogeneous populations and methodologies, which would establish robust biomarkers despite study variability; and (4) pregnancy-specific mechanistic frameworks distinct from type 2 diabetes models, recognising the unique physiological and metabolic challenges specific to gestation, placental development, and foetal nutrient demands.
Geographic variation, diverse sequencing methodologies, limited longitudinal data, and the tendency to extrapolate directly from T2D models further complicate synthesis of existing evidence and hinder the development of pregnancy-specific therapeutic strategies. Therefore, this narrative review synthesises existing literature on how the gut microbiota shifts through different stages of pregnancy in diverse populations under both healthy and diabetic conditions. By identifying convergent microbiota signatures across heterogeneous studies, integrating human and animal model evidence, and articulating pregnancy-specific mechanistic insights, this review establishes the evidence base necessary for developing clinical strategies, nutritional guidelines, and microbiota-targeted interventions tailored to maternal care and GDM prevention.
2. Literature Search Strategy
This narrative review synthesised all available evidence on gut microbiota composition during healthy pregnancy and gestational diabetes in human and mouse models. A structured search of the NCBI PubMed database was performed to identify studies published between January 2005 and February 2025 using keywords and combinations: “gut microbiome”, “gut microbiota”, “gut bacteria”, “pregnancy”, “trimester 1”, “trimester 3”, “gestational diabetes”, “GDM”, “human”, “mouse”, “murine”.
All studies reporting gut microbial composition during pregnancy or GDM were included, regardless of sequencing platform (16S rRNA or whole genome metagenomics) or study location. Given the high variability of reported taxa across studies and the heterogeneity of reported microbial signatures across cohorts and platforms, comprehensive data synthesis was prioritised over formal inclusion and exclusion criteria. Conference abstracts, studies unrelated to pregnancy or GDM and microbiome analyses only focusing on oral or vaginal microbial composition or discussing microbial metabolites but lacking sequencing data of microbial composition in the gut were excluded. To ensure completeness, database searches were supplemented with reference list screening of included studies to identify additional relevant publications. Consequently, a total of 87 studies were included summarising gut microbial compositional across different stages of healthy pregnancy and gestational diabetes in both human and murine models.
3. Overview and Establishment of the Neonatal Microbiome
A dynamic and complex ecosystem comprising bacteria, archaea, eukaryotic microbes and viruses, the gut microbiota contains as many cells as the number of human cells in the body [34]. Its microbial genome (so-called “microbiome”) represents 150-fold more genetic varieties than the entire human genome [8,35,36,37,38]. The compilation of the MetaHit (Metagenomics of the Human Intestinal Tract), Human Microbiome Project and Chinese cohort, providing the most comprehensive and intercontinental view on the human microbial repertoire, revealed twelve bacterial phyla and one archaeal taxon (Euryarchaeota) of Prokaryotes as commensals or pathogens in human beings [39,40]. Meanwhile, according to rRNA sequencing and GutFeelingKB, a reference of curated metadata for healthy gut microbiome, healthy human gut microbiota can be taxonomically classified into 8 phyla, 18 families, 38 orders and 59 genera, representing more than 7000 strains—belonging to at least 1000 specific phylogenetic types (or “phylotypes”) [41,42,43]. The eight gut microbial phyla, in order of decreasing phylogenetic abundance, are: Bacillota and Bacteroidota (formerly known as Firmicutes and Bacteroidetes), Actinomyceota and Pseudomonadota (formerly named Actinobacteria and Proteobacteria), Verrucomicrobia, Fusobacteria, Synergistetes and Euryarchaeota [44].
The GI tract of the foetus was initially believed to be sterile until the first microbial colonisation at birth [3,38]. However, this belief has been challenged by several findings over the past decades which have demonstrated the existence of bacterial DNA or certain SCFAs in the first-pass meconium, placenta, umbilical cord blood and amniotic fluid in healthy pregnancy [45,46,47,48,49,50,51,52,53]. The placenta has been identified as a niche for commensal species such as Bacillota, Bacteroidota, Pseudomonadota, and Fusobacteriota, and largely resembles the mother’s oral microbiota [52]. The observed similarities among the microbial profiles of the mother’s mouth, placenta, amniotic fluid, and first-pass stool suggest the bacteria cross the placenta—a selective barrier between foetus and mother—and their potential translocation to the developing gastrointestinal tract [45]. This underscores the critical role of the maternal microbiota in shaping the initial establishment of the infants’ microbiome, regardless of differing views on its origin.
Although it is now known that colonisation commences during intrauterine life, the period immediately after birth plays a significant role in determining the initial gut microbial composition. Mode of delivery is a major factor influencing the initial gut microbial composition [3,38,54]. The gut microbiota of vaginal delivered newborns has been shown to inherit the ecosystem of their mother’s vagina, which typically includes Bacteroidota species, especially Lactobacillus, Prevotella genera or Sneathia genera under the Fusobacteriota phylum [54,55]. Meanwhile, the gut microbiota from newborns delivered via caesarean section is dominated by Clostridium, Staphylococcus, Corynebacterium, and Propionibacterium genera—which resembles the microbiota of maternal skin or the hospital environment [55,56,57]. Caesarean-delivered infants also had less diverse species and experienced delayed colonisation of Bacteroidota microorganisms compared to babies delivered vaginally [55,56]. Therefore, the establishment of the infants’ gut microbiome can vary substantially and has the potential to significantly influence ongoing health in childhood and adulthood.
4. Baseline Gut Microbiota Composition: Non-Pregnant Individuals
4.1. Dominant Bacterial Phyla in Healthy Adults
Establishing the baseline gut microbiota composition in non-pregnant individuals is essential for understanding how it adapts and changes during pregnancy. While microbiota data specifically from non-pregnant women of reproductive age remain limited, data from healthy non-pregnant adults provide a valuable reference point for understanding pregnancy-related microbial shifts. While differences in the core microbiome across continents and ethnicities should be acknowledged, it is well recognised that Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria are the dominant phyla in healthy human gut communities [3,58,59,60,61]. In healthy adults, the gut microbiota is dominated by Firmicutes (40–70%), followed by Bacteroidetes (20–45%) and Actinobacteria (1–10%), though these proportions may vary with diet, geography, and other factors [39,44,61,62,63,64,65], which is consistent with reports specifically describing the proportions in healthy women of child-bearing age, particularly Firmicutes (50–70%), Bacteroidetes (20–35%), Actinobacteria (2–9%), and Proteobacteria (3–4%) [66,67].
4.2. Lower Taxonomic Ranks: Dominant Genera and Enterotypes
At the genus level and below, specific dominant taxa have been consistently identified across independent studies. Key genera include Faecalibacterium, Clostridium, Lachnospiraceae, Roseburia, Ruminococcus (under Firmicutes); Bacteroides, Prevotella, Alistipes (under Bacteroidetes); and Bifidobacterium (under Actinobacteria) [39,44,61,62,63,64]. Among these, studies from different cohorts have reliably identified key taxa that underpin three major gut microbiota enterotypes representing distinct microbial community structures: Ruminococcus-dominant, Bacteroides-dominant, and Prevotella-dominant networks [44,68,69,70,71]. Whilst these enterotypes likely reflect stable, long-term microbial configurations, they can be shaped by individual factors including diet and host genetics. When interpreting microbiota findings across studies, several methodological factors warrant careful consideration. Cross-study differences in participant age, geographic location, dietary background, and sequencing methodologies can substantially influence reported microbiota composition. Additionally, faecal samples serve as a convenient proxy for gut microbiota but do not fully represent the complexity of the entire gastrointestinal tract, which exhibits compositional variation both axially (mucosal vs. luminal communities) and longitudinally (proximal vs. distal regions) [72,73]. These limitations underscore the importance of cautious interpretation when generalising findings across populations and study designs.
5. Gut Microbiota Composition in Early Pregnancy
The gut microbiota is shaped by multiple factors, including host genetics, age, diet, antibiotic use, and geography [74,75,76,77]. Alterations in its composition are also commonly reported in inflammation-related disorders such as Crohn’s disease, inflammatory bowel disease (IBD), allergies, as well as in conditions linked to host adiposity and insulin resistance [78,79,80,81,82]. Therefore, in response to substantial metabolic and inflammatory changes during a healthy pregnancy to support implantation, placentation, and foetal growth, corresponding shifts in the gut microbiota are expected [83]. There is limited research directly comparing the gut microbiota of healthy women in the first trimester of pregnancy with the microbial profile before conception. Most existing studies examine changes from early to late pregnancy rather than comparing to the non-pregnant state, and many focus on women with high body mass index (BMI) and/or varying GDM status. Although data are limited, early pregnancy (from the first to early second trimester, before week 20) is typically characterised by a proinflammatory state, with or without insulin resistance [84,85]. This stage likely exerts a smaller influence on gut microbiota composition compared to later pregnancy, when pronounced metabolic changes such as increased insulin resistance and anti-inflammatory adaptations become more evident (Table 1). According to the microbial analysis conducted by Koren et al. in pregnant women across trimesters, although late pregnancy showed significant shifts, early pregnancy microbiota resembled that of non-pregnant individuals, with comparable β-diversity and differences in Firmicutes and Bacteroidetes abundance [86].
5.1. Phylum Level Composition in Early Pregnancy
Altered relative abundance of Firmicutes and Bacteroidetes has been observed in early pregnancy [86]; though direct evidence linking their abundance to specific physiological changes during early pregnancy remains limited. In healthy non-pregnant adults, Firmicutes has been positively associated with increased adiposity and triglyceride levels, whereas Bacteroidetes shows a negative association [87,88]. This relationship may partly explain findings from healthy early pregnancies, where weighted UniFrac analysis revealed that first-trimester samples clustered toward a higher Firmicutes gradient and a lower Bacteroidetes gradient compared to non-pregnant controls [86]. However, a case–control study of 98 healthy women at gestational weeks 10–15 reported comparable relative abundances of these phyla [25]. Such discrepancies highlight ongoing controversies regarding F/B ratios in early pregnancy. These inconsistencies are further complicated when overweight (non-GDM) pregnancies are included, in which greater variation in F/B ratio was observed. For instance, at gestational week 16, overweight (non-obese, non-GDM) women exhibited a F/B ratio of approximately 3:1 [87]. In contrast, Mokkala et al. found a higher proportion of Bacteroidetes compared to Firmicutes at week 14 among 270 overweight/obese women (including 203 without GDM onset). This finding is consistent with their earlier research on 60 overweight/obese non-GDM women and the sub-unit value of the F/B ratio in a matched case–control study of 120 healthy controls at week 10 [89,90,91]. Overall, phylum-level composition during early pregnancy closely resembles that of non-pregnant individuals; particularly, Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria remain predominant during the first trimester, with Firmicutes and Bacteroidetes together accounting for over 80% of total microbial abundance, Actinobacteria 2–10% and Proteobacteria roughly 2% [25,61,86,87,89,90,91,92,93,94,95,96,97,98]. However, the relative dominance hierarchy among these phyla, particularly the ranking of Firmicutes over Bacteroidetes, or Actinobacteria over Proteobacteria, is not consistently reported across first trimester studies, reflecting underlying variability across cohorts.
5.2. Lower Taxonomic Ranks and Enterotype Patterns in Early Pregnancy
At the genus level, microbial hierarchies also mirror those of non-pregnant cohorts: Bacteroides, Prevotella, and Alistipes dominate within Bacteroidetes, while Faecalibacterium, Blautia, Agathobacter, and Roseburia dominate within Firmicutes, establishing Clostridia as the leading class both before and during early pregnancy [25,61,86,87,89,90,91,92,93,94,95,96,97,98]. Among all genera, Bacteroides consistently represents the most abundant taxon, followed by Faecalibacterium, Prevotella, and Blautia [25,61,92]. These genera show broad consistency across geographic regions, with the notable exception of Prevotella, which exhibits marked variation between U.S. and Chinese cohorts and even among different Chinese populations, driven primarily by dietary differences and host genetics [25,61,92,93]. In normoglycemic pregnancies, the gut microbiota is characterised by increased abundance of Prevotellaceae, Fusobacteriales, and Sutterella [92]. Intriguingly, later GDM onset correlates negatively with Prevotella abundance and serum cholesterol levels [92], suggesting a potential protective role for Prevotella in early pregnancy.
Analysis by Xiao et al. identified three distinct clusters from gut microbial analysis among 1715 pregnant women worldwide: one Firmicutes-dominated cluster and two dominated by Bacteroides and Prevotella, respectively [61]. These patterns align with enterotypes observed in non-pregnant populations [44]. Specifically, in the first trimester, cluster 1 was enriched in Bacteroidetes (>50%), cluster 2 featured Prevotella (~50%) and Bacteroides (~12%), and cluster 3 was Firmicutes-dominated, with Faecalibacterium (~18%) and unclassified Lachnospiraceae (~30%). These findings reinforce the concept of a relatively stable gut microbial structure during the transition from non-pregnant to early gestation, despite some variation in Firmicutes and Bacteroidetes abundance, a feature likely linked to mild metabolic and pro-inflammatory changes with pregnancy onset.
6. Gut Microbiota Composition in Late Pregnancy
Beyond the first 20 weeks of gestation, progressive metabolic and immunological adaptations support rapid foetal growth. Elevated circulating concentrations of progesterone, oestradiol, and placental lactogen contribute to the regulation of insulin and promote leptin resistance, facilitating maternal–foetal glucose transfer to meet the increasing demands of the foetus [99]. By the late second and third trimesters, insulin sensitivity can decline by up to 50%, accompanied by low-grade mucosal inflammation. These physiological changes are likely to influence the composition of the commensal gut microbiota during late pregnancy [26,100,101,102].
Gut microbial remodelling in late pregnancy is well documented, with significant β-diversity differences compared to non-pregnant women or early-pregnancy samples across diverse populations in Asia, Europe, the Americas, and Africa (Table 2) [61,86,95,103]. After adjusting for BMI, GDM status and technical factors, the magnitude of β-diversity alteration from first to third trimester were attributed mainly to physiological adaptations of pregnancy [86]. In contrast, α-diversity shows inconsistent patterns with nearly half of studies reporting marked reductions in indices such as Shannon, Faith’s phylogenetic diversity, observed OTUs, or species counts—representing within-sample microbial richness and/or evenness, whilst others find no significant differences [61,86,89,91,93,95,96,97,103,104,105].
6.1. Phylum Level Remodelling: Progressive Shifts Toward Dysbiosis Markers
At the phylum level, Actinobacteria is the only group consistently reported to increase as gestation advances [86,94,102], evidenced by frequent enrichment of its dominant genera, Bifidobacterium and Collinsella, both lactic acid producers [61,93,94,95,106]. Maternal fasting glucose levels have been positively associated with Actinobacteria abundance, while Bifidobacterium, known to promote placental morphogenesis and regulate foetal growth, has been linked to healthy gestational weight gain and increases in abundance in response to elevated circulating progesterone levels during late pregnancy [94,106,107]. Certain Bifidobacterium species are recognised as early colonisers of the infant gut, contributing to neonatal immune development [108,109,110,111].
Changes in Firmicutes and Bacteroidetes during healthy pregnancy show less consistency between studies. However, a decline in the F/B ratio during the third trimester is more reproducible and likely represents a physiological adaptation to support foetal glucose demands and gestational weight gain in healthy pregnancies, though whether it constitutes true dysbiosis or predisposes to metabolic complications in at-risk populations remains unclear [89,91,102,104]. The functional significance of this F/B decline becomes evident when compared across metabolic states: higher Firmicutes abundance correlates with excessive gestational weight gain, whereas higher Bacteroidetes is associated with normal weight gain [106,112]. This contrasts sharply with obesity-related dysbiosis, which shows elevated Firmicutes and reduced Bacteroidetes (increased F/B ratio) [62,113,114,115]. Conversely, type 2 diabetes (T2D) is linked to reduced Firmicutes and Clostridia—one of the largest and predominant classes within Firmicutes [116]. Interestingly, a longitudinal case–control study of 120 pairs of matched pregnant controls and women with GDM reported an inverse correlation between F/B ratio and maternal fasting plasma glucose throughout gestation [89]. Thus, despite variability in absolute abundances, the consistent decline in F/B ratio during healthy late pregnancy may reflect adaptive sub-phylum shifts (e.g., increased Bacteroidetes, decreased Firmicutes) that support normal physiological weight gain and glucose metabolism. Proteobacteria, including Enterobacteriaceae, frequently implicated in inflammation-related disorders such as Crohn’s disease, IBD, colorectal cancer, and obesity [117,118,119,120], shows no consistent pattern from early to late pregnancy, but an increasing trend is more well-documented [86,89,97,103,106].
6.2. Changes in Lower Taxonomic Ranks: Functional Trade-Offs Between Lactic Acid and Butyrate Producers
At the genus level, randomised controlled trials in metabolically healthy women report increased lactic acid-producing bacteria and decreased butyrate producers as pregnancy progresses [121,122]. For example, Ruminococcus, a common butyrate producer within Firmicutes, consistently declines during the third trimester [86,94,103]. Notably, Ruminococcus gnavus produces an inflammatory polysaccharide that stimulates TNF-α, and high abundances of certain Ruminococcus species have been associated with allergies in children (4–6 years old), inflammatory disorders and mucosal damage in non-pregnant adults [123,124,125,126]. Conversely, Streptococcus (Firmicutes), a lactic acid producer, shows consistent increase during pregnancy, similar to Bifidobacterium and Collinsella (Actinobacteria) [61,86,95,103]. Both genera are considered beneficial for gestational outcomes, as reduced abundance has been associated with spontaneous preterm delivery, as reported in a case–control study from Eastern Norway [127].
Another notable trend is the rise in Bilophila throughout gestation [95,102,103], a hydrogen sulfide-producing genus within Desulfovibrionales, typically present at low abundance in healthy guts [128]. Bilophila derived from human stool can induce systemic inflammation in pathogen-free mice and impaired glucose tolerance [95,102,103,129,130]. Moreover, Bilophila has also been associated with high-fat-diet-induced metabolic dysfunctions in mice [130]. However, a meta-analysis further highlighted the contrasting roles of sulfate-reducing versus lactic acid-producing bacteria in gut inflammation: excessive hydrogen sulfide can provoke intestinal toxicity, whereas lactic acid bacteria enhance nutrient bioavailability, promote beneficial microbiota composition, and help prevent IBD [131]. This nuanced interplay between these bacterial groups is critical for maintaining gut function and barrier integrity during late pregnancy—a period marked by heightened nutritional demands and inflammatory responses.
6.3. Mechanistic Insights: Experimental Evidence and Functional Significance
Experimental evidence supports a causal role for microbiota in metabolic adaptations during late pregnancy. Germ-free mice transplanted with third-trimester microbiota exhibited greater weight gain and insulin resistance compared to those receiving first-trimester microbiota [86]. Additionally, multiple studies report correlations between specific taxa and metabolic parameters, including glucose-related markers (FPG, OGTT, HbA1c, HOMA-IR, hyperglycaemia) and adiposity-related measures (HDL, cholesterol, weight gain, dyslipidaemia) [61,89,95,98,102,106,132,133,134]. However, most research has focused on compositional changes, with limited exploration of underlying mechanisms. It remains unclear whether these shifts represent physiological adaptations to meet foetal nutritional demands or protective responses to pregnancy-induced adiposity, insulin resistance, and inflammation. Further mechanistic studies are needed to clarify the functional significance of these taxonomic changes and their contribution to either healthy gestational outcomes or the development of gestational pathologies.
7. Gut Microbiome Composition in Gestational Diabetes
Gut dysbiosis is well documented in T2D, characterised by increased opportunistic pathogens and reduced beneficial taxa [142]. Healthy individuals typically harbour higher levels of butyrate-producing bacteria, whereas T2D shows marked reductions in Clostridium species and Akkermansia muciniphila—taxa with negative correlations with fasting glucose and HbA1c, suggesting a potential role in disease pathogenesis [143,144,145]. Given that GDM is a pregnancy-specific form of diabetes, similar microbial alterations might be expected; however, GDM-associated microbiota changes remain poorly understood (Table 3). Early evidence from a comprehensive multi-omics study of 394 pregnant women identified gut dysbiosis prior to GDM diagnosis [146], suggesting that pathological microbiota remodelling may precede clinical disease manifestation.
7.1. Diversity Patterns: α and β-Diversity as Dysbiosis Markers
Microbial diversity patterns in GDM show contrasting results depending on the reported diversity metric. α-diversity (within-sample richness and evenness) generally shows no significant differences between GDM and healthy controls in most studies, aside from a declining trend during the first four months of pregnancy [61,86,89,91,92,95,132,133,136,137,140,141,146,147]. Conversely, β-diversity differences (between-sample compositional variation) between GDM and non-GDM groups are significant in more than half of studied populations [61,89,92,97,104,134,135,139,140,141,146]. During late pregnancy, women with GDM exhibit similar α and β-diversity trends compared to non-GDM groups, with persistent β-diversity differences (in four of five studies) and significant declines in microbial richness (in three of five studies) [26,61,86,91,104], suggesting potential dysbiosis throughout GDM progression. Importantly, reduced Shannon index and richness have been correlated with higher insulin resistance (HOMA-IR) in T2D cohorts [148], and similar inverse correlations between Shannon index and fasting plasma glucose, triglycerides, and cholesterol have been observed in GDM [89,138]. These findings align with the trends observed in non-pregnant cohorts, for example Chatelier et al., who reported that lower microbial richness in 292 Danish volunteers (including 169 with obesity) was associated with inflammation, insulin resistance, adiposity, and dyslipidaemia [149].
7.2. Phylum Level Alterations: Elevated F/B Ratio as a Dysbiosis Marker
At the phylum level, GDM pregnancies frequently show Firmicutes enrichment and Bacteroidetes reduction, resulting in an elevated F/B ratio in either early or late pregnancy compared to the non-GDM group [26,95,97,104,132,134,136,150,151]. This elevated F/B ratio has been proposed as a marker for pathological conditions, including obesity, T2D, and adverse pregnancy outcomes, contrasting sharply with the declining F/B ratio observed in healthy pregnancies [100,101,152,153]. Notably, among women who develop GDM by week 28, a higher F/B ratio was repeatedly detected at week 16 in both obese and overweight women, though Firmicutes abundance was significantly higher in obese than overweight women [87]. Reduced Bacteroidetes abundance was also observed in overweight compared to normal weight pregnancies [106], suggesting an association between early F/B ratio elevation and abnormal weight gain—a potential compounding risk factor. Recently, increased F/B ratio has also been shown to significantly correlate with FPG and 1h-OGTT [154].
Other phyla such as Actinobacteria, Proteobacteria, and Verrucomicrobia show no consistent trends in GDM. Critically, distinctive features of healthy pregnancy such as reduced F/B ratio and increased Actinobacteria (particularly Bifidobacterium) are often absent or reversed in GDM [26,61,95,104], suggesting that GDM is characterised by distinct inflammatory and metabolic adaptations driven by heightened insulin resistance that diverge from normal pregnancy physiology.
7.3. Lower Taxonomic Rank Alterations: Compensatory Responses and Predictive Biomarkers
At the genus level, alterations in Faecalibacterium and Roseburia, two major butyrate producers, are inconsistent in GDM and do not mirror the reductions commonly observed in T2D [142,155,156]. Conversely, other butyrate-producing genera, including Blautia [97,104,132,137,138,140,147], Ruminococcus [90,95,132,136,137], and Lachnospiraceae [104,132,136,138], are consistently elevated in GDM compared to healthy controls. This enrichment may represent a compensatory response attempting to maintain metabolic homeostasis. Supporting this hypothesis, a GDM mouse model where live Lachnospiraceae and butyrate supplementation improved gut barrier integrity, reduced placental inflammation, and mitigated GDM phenotypes—effects lost when bacteria were heat-inactivated or treated with a butyrate inhibitor [157]. These findings suggest that butyrate-producing capacity, rather than absolute bacterial abundance, may be critical for metabolic control during GDM.
Within Bacteroidetes, genera such as Bacteroides [89,90,136,137,140], Prevotella [92,95,146], and Alistipes [89,135,139] generally follow the phylum-level decline observed in GDM, whereas Parabacteroides often shows the opposite trend [61,104,135]. Notably, Bacteroides and Parabacteroides demonstrated strong predictive power for GDM in shotgun metagenomics analysis of 43 GDM and 81 healthy pregnancies [135]. High maternal Prevotellaceae abundance was associated with euglycemic pregnancies in a Czech cohort [92]. While Parabacteroides, typically enriched in overweight women, correlates positively with glucose tolerance [135,137], Alistipes shows inverse associations with hsCRP, fasting glucose, and OGTT results [26,89,135,137,139].
Bifidobacterium, which normally increases during healthy pregnancy, is notably reduced in GDM compared to normoglycaemic controls [89,104,135]. Metagenomic analysis of 27 GDM pregnancies identified Bifidobacterium as a potential protective factor, with its abundance rising after glycemic intervention, possibly enhancing SCFA production and improving glucose homeostasis [158,159]. Similarly, Sutterella (Proteobacteria) was more abundant in normoglycemic pregnancies than in GDM cases in a Czech cohort [92]. In T2D rat models, Sutterella improved glucose tolerance post-gastric bypass [160]. Interestingly, CRP—a well-established biomarker of systematic inflammation and proposed indicator for future onset of T2D—was also positively correlated with Sutterella during GDM progression [26], though this association is absent in healthy pregnancies, suggesting Sutterella may have distinct functional roles in different metabolic contexts.
7.4. Outstanding Questions: Methodological Variability and Transgenerational Effects?
Despite these observations, GDM-related microbial changes remain inconsistent across taxonomic levels, likely reflecting differences in gestational timing of sampling, cohort sample size, geographic locations, dietary patterns and sequencing methodologies. Further research is needed to clarify mechanistic pathways and establish causal relationships. Importantly, emerging evidence suggests that maternal gut dysbiosis may influence neonatal microbiota composition and immune development: reduced α-diversity and absence of unique taxa have been observed in newborns of GDM mothers compared to controls [46,161], raising questions about whether GDM-associated dysbiosis perpetuates metabolic dysfunction across generations through early-life microbial exposure.
8. Animal Models for Studying Gut Microbiota Composition During Pregnancy
Given the variability of microbial shifts across human studies due to experimental design and population characteristics, studies on controlled animal models have become essential, disentangling intrinsic mechanism of microbial changes and providing valuable complementary approaches to human studies for investigating gut microbiota dynamics during pregnancy (Table 4). However, such research remains relatively limited. Substantial differences also exist between human and animal microbial profiles due to variations in diet, physiology, and genetics [162]. Additionally, the much shorter gestational periods in rodents—19–21 days in mice and 22–23 days in rats compared to approximately 280 days in humans—pose challenges for translating findings to human conditions. Despite these limitations, animal models offer distinct advantages unavailable in human research. They enable rigorous control over diet and pregnancy induction, which is often constrained in human studies by ethical and practical considerations [162,163,164,165]. Furthermore, confounding variables such as lifestyle, medication, and environmental exposures can be standardised by controlling housing conditions and using uniform genetic backgrounds [162,163,164,165]. Critically, many tissues essential for microbial investigation during pregnancy such as segments of the gastrointestinal tract, placenta and maternal organs cannot be ethically collected from pregnant women, making animal models indispensable for mechanistic research.
8.1. Model Selection and Microbiota Comparability
Among available models, mice are the most widely used, either in direct studies or through human faecal microbiota transplantation experiments, to explore microbial composition and host–microbiota interactions. This preference reflects structural similarities between human and murine gut microbiota. Mouse gut bacteria is also dominated by four major phyla: Firmicutes (~55.8%), Bacteroidetes (~37.0%), Proteobacteria (~4.1%), and Actinobacteria (~2.0%) [166]. Several prevalent genera are also shared, including Lactobacillus, Roseburia, and Ruminococcaceae UCG-014 within Firmicutes; Bacteroides, Alloprevotella, and Alistipes within Bacteroidetes; and Escherichia-Shigella within Proteobacteria [166]. Murine models have also provided substantial mechanistic insights into disease processes. For example, microbiota from individuals with IBD exacerbates colitis in mice [121], and gut dysbiosis has been implicated in type 1 diabetes, acute pancreatitis, and neurobehavioral disorders, mirroring patterns observed in humans [122,167,168,169,170]. Critically, as pregnancy progresses, microbial diversity patterns in mice closely parallel those observed in humans, characterised by minimal changes in α-diversity but significant shifts in β-diversity [94,171,172,173,174,175,176].
8.2. Phylum Level Changes: Consistent Bacteroidetes Reduction and Proteobacteria Enrichment
At the phylum level, murine pregnancy studies consistently report a decrease in Bacteroidetes accompanied by an increase in Proteobacteria [173,174,175,176,177,178]. Findings on the F/B ratio show greater variability across studies, though an overall upward trend is suggested, primarily driven by Bacteroidetes reduction [173,175,177]. Proteobacteria enrichment aligns with observations in healthy human pregnancies and has been associated with increased inflammation in late gestation [86,179]. Notably, mono-colonisation with E. coli (a Proteobacteria) in germ-free Swiss Webster mice promoted macrophage accumulation and disrupted glucose metabolism [86,179], suggesting potential metabolic consequences of Proteobacteria expansion during pregnancy.
8.3. Lower Taxonomic Rank Dynamics: From Nutrient Extractors to Cross-Species Conserved Taxa
Genus-level remodelling appears more consistent across murine studies compared to human studies. Many Firmicutes genera involved in nutrient extraction and energy metabolism, such as Faecalibacterium, Roseburia, Allobaculum, Turicibacter, Ruminococcus, Streptococcus, and Lactococcus, consistently increase during murine gestation. These taxa are recognised producers of short-chain fatty acids (SCFAs), glucose, and lactic acid, which play key critical roles in energy metabolism and immune homeostasis. Their enrichment may reflect adaptive responses supporting anti-inflammatory properties and immune tolerance during pregnancy [175,180].
8.4. Cross-Species Conserved Taxa: Biomarkers for Functional Investigation
Only a limited number of genera show similar enrichment patterns during late pregnancy in both humans and mice, notably Streptococcus (Firmicutes), Bifidobacterium, and Collinsella (Actinobacteria). In humans, Bifidobacterium and Collinsella are consistently enriched during late pregnancy, and comparable increases have been observed in mice at embryonic days 15–18 across strains including C57BL/6J, Swiss Webster, and BALB/c [94,171,180,181]. The enrichment of Bifidobacterium appears particularly protective in mice. Supplementation with specific strains has been shown to improve insulin sensitivity and reduce fat accumulation, partly through modulation of metabolic pathways in the gut and adipose tissue [182,183]. Oral administration of Bifidobacterium spp. also improved glucose tolerance by lowering inflammatory adipokines in both mice and rat L6 myoblast cells [184,185], suggesting conserved metabolic functions across rodent species and potential relevance to human pregnancy.
8.5. Rodent Specific Changes: Understanding Model Limitations and Opportunities
Other consistent changes in murine gut microbiota during pregnancy include increases in the opportunistic pathogen Helicobacter and the commensal genus Mucispirillum, which is abundant in rodents but much less common in humans [172,174,176,180,181]. The functional significance of Mucispirillum enrichment during pregnancy remains debated and is likely context dependent. In some contexts, Mucispirillum can antagonise Salmonella virulence and protect against colitis by limiting pathogen invasion [185]. Conversely, in Nod2/Cybb-deficient mice (Crohn’s disease-prone model), Mucispirillum exacerbated colitis, highlighting its pathobiont potential under dysbiotic or immune compromised conditions [186]. These species-specific patterns underscore the importance of interpreting murine findings within an appropriate biological context.
8.6. Translational Implications and Future Directions
Murine models remain among the most suitable systems for studying pregnancy-related microbiota dynamics, offering precise environmental control and access to tissues and mechanistic insights unavailable in human research. Specifically, microbial shifts consistently observed in mice that have not yet been systematically characterised in humans warrant targeted investigation in human cohorts. Conversely, taxa showing overlapping patterns in both species represent promising targets for mechanistic studies exploring their functional roles in supporting healthy pregnancy outcomes. Integration of human and murine data through carefully designed comparative studies may ultimately identify universal principles governing pregnancy-induced microbiota remodelling and reveal opportunities for microbiota-targeted interventions to prevent or mitigate pregnancy complications.
9. Discussion
Pregnancy is a physiological state characterised by profound metabolic and inflammatory adaptations which also bi-directionally influence gut microbiota composition [83] (Figure 1). During the first trimester, gut microbial composition remains largely comparable to that of non-pregnant individuals [25,61,86,92]. However, from the second trimester onwards, notable compositional shifts emerge, reflected by significant changes in β-diversity compared to the first trimester or non-pregnant controls [61,86,103]. At the phylum level, healthy pregnancy has often been associated with a decline in the F/B ratio, driven by reduced Firmicutes and increased Bacteroidetes abundance. These changes correlate with physiological gestational weight gain and elevated maternal fasting glucose levels in metabolically healthy pregnancies [62,106,112,113,114,115]. A distinctive hallmark of healthy pregnancy is increased Actinobacteria abundance, particularly its dominant genus Bifidobacterium [86,94,102]. Bifidobacterium appears especially beneficial during pregnancy, promoting placental morphogenesis, enhancing nutrient transport, and contributing to early neonatal immune development [106,107,108,109,110,111].
At the genus level, healthy pregnancy exhibits two contrasting patterns. First, Ruminococcus (a butyrate-producing genus) consistently declines during gestation [86,94,103]. This reduction may reflect selective suppression of certain Ruminococcus species that produce inflammatory polysaccharides activating immune responses and compromising gut barrier integrity, highlighting that not all butyrate producers are uniformly beneficial. Conversely, the enrichment of lactic acid-producing genera, such as Bifidobacterium, Collinsella, and Streptococcus, is frequently reported [61,86,95,103]. These taxa may enhance nutrient bioavailability and promote a healthier gut environment; notably, Streptococcus and Bifidobacterium have been associated with reduced risk of preterm birth [127,131]. However, the precise mechanistic roles of these taxa in maintaining healthy pregnancy outcomes remain controversial.
GDM is the most common metabolic disorder during pregnancy. The gut microbiota in GDM exhibits alterations that partially resemble those in T2D [137]. A defining feature is the elevated F/B ratio [26,95,132,136,150], similar to T2D patterns but opposite to healthy pregnancy [89,91,102,104], suggesting pathological rather than physiological remodelling. Importantly, Bifidobacterium—a beneficial strain increased in healthy pregnancies and associated with placental morphogenesis and nutrient bioavailability [94,106,107]—is consistently reduced in GDM [26,61,95,104]. Additional GDM-associated changes include significant β-diversity shifts [61,89,92,97,104,134,135,139,140,141,146], increased Ruminococcus abundance [90,95,132,136,137] and the reduced levels of Bacteroidetes genera [97,104,132,134,136], Clostridium, and Akkermansia compared to normoglycemic pregnancies [26,61,95,104,188]. These microbial patterns closely mirror T2D dysbiosis, suggesting shared pathological mechanisms.
Despite these similarities, important distinctions exist between GDM and T2D dysbiosis that suggest pregnancy-specific metabolic challenges. While butyrate-producing bacteria typically decline in T2D, they remain unchanged (Faecalibacterium, Roseburia) or even increased (Blautia, Ruminococcus, Lachnospiraceae) in GDM [95,97,104,132,136,137,138,140,147]. This divergence indicates a more complex functional role for butyrate producers in pregnancy, potentially driven by factors beyond glucose homeostasis, such as foetal nutrient demands, placental development, or pregnancy-specific immune tolerance mechanisms. These findings underscore the need for pregnancy-specific mechanistic research rather than direct extrapolation from T2D models. Clarifying the underlying mechanisms driving GDM-associated dysbiosis remains critical for developing targeted interventions.
Gut microbiota changes during pregnancy have also been investigated in murine models, which offer value for mechanistic investigation due to similarities between mice and humans in digestive anatomy and predominant gut microbial composition. However, existing murine pregnancy microbiota studies remain limited in number and present conflicting observations. Notably, enrichment of two Actinobacteria genera (Bifidobacterium and Collinsella) have been consistently observed during late pregnancy in both humans and mice [94,171,180,181]. This shared feature underscores the potential biological significance of Proteobacteria and Actinobacteria in pregnancy-related gut microbiota changes across species. Conversely, divergent patterns have been reported for the two predominant phyla, Firmicutes and Bacteroidetes, between pregnant mice and humans, highlighting the importance of cautious interpretation when extrapolating findings from animal models to human physiology. Such inconsistencies may reflect species-specific differences in microbial functional capacity, baseline microbiota composition, or fundamental physiological differences in human versus murine pregnancy (e.g., gestation duration, placental architecture, immune development timelines).
Although the gut microbiota represents a vast and largely invisible ecosystem, it remains a modifiable target and has a well-established association with both healthy pregnancy and GDM pathogenesis. This recognition opens clinical opportunities for microbiota-informed interventions. Clinical strategies should emphasise personalised dietary guidance informed by individual microbiota composition, identification of beneficial versus potentially pathogenic microbial taxa, and targeted supplementation approaches. Emerging therapeutic options include faecal microbiota transplantation, prebiotics, probiotics, synbiotics, and postbiotics—each offering distinct mechanistic advantages. Prebiotics enhance growth of endogenous beneficial bacteria; probiotics and synbiotics introduce or reinforce beneficial taxa; postbiotics provide metabolic byproducts of fermentation without requiring viable organisms. These microbiota-modulating interventions represent promising, practical avenues for mitigating GDM risk, optimising metabolic adaptations during healthy pregnancy, and ultimately supporting maternal and foetal health throughout gestation and beyond.
10. Limitations
This narrative review synthesising gut microbiota studies during healthy pregnancy and gestational diabetes across human and mouse models faces several important limitations. Methodological heterogeneity including variable sequencing technologies (16S rRNA, shotgun metagenomics, genus-specific probes) and inconsistent taxonomic reporting limits cross-study comparability and quantitative synthesis. Population heterogeneity further complicates interpretation, with variations in gestational age, maternal metabolic state, GDM diagnostic criteria, and inconsistent adjustment for confounders (BMI, diet, medications, parity) potentially complicating conclusion strength. The gut microbiota represents a highly interconnected system, yet individual studies typically examine only specific bacterial taxa or functional domains, risking speculative interpretations when direct evidence is insufficient. Additionally, cross-study comparisons may reflect contextual differences (diet, lifestyle, geography, environmental exposures) rather than true pregnancy related microbial signatures. Despite these limitations, this comprehensive synthesis advances the field by comprehensively integrating findings across diverse experimental models and populations, thereby establishing a robust evidence base that identifies critical knowledge gaps, prioritises key research directions, and provides a platform for hypothesis-driven investigations to clarify pregnancy-associated microbiota dynamics and their metabolic consequences.
11. Conclusions
Pregnancy induces profound metabolic and immunological changes that shape gut microbiota composition. Evidence consistently highlights increased Actinobacteria, particularly Bifidobacterium, as a hallmark of healthy pregnancy, while shifts in Firmicutes and Bacteroidetes remain variable. In contrast, GDM exhibits microbial patterns resembling T2D, including elevated F/B ratios and reduced Bifidobacterium, suggesting pathological rather than physiological adaptations. However, inconsistencies across studies driven by differences in geography, methodology, and gestational timing underscore the need for standardised approaches and longitudinal designs.
Animal models provide mechanistic insights but require cautious interpretation due to species-specific differences. Future research should prioritise functional studies linking microbial shifts to metabolic pathways, explore microbiome-based interventions, and assess their impact on maternal and neonatal outcomes. Ultimately, integrating microbiome-informed strategies into clinical care through diet and gut-targeted therapies offers promising avenues for improving pregnancy health and reducing GDM risk.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Srivastava A. Prabhakar M.R. Mohanty A. Meena S.S. Influence of gut microbiome on the human physiology Syst. Microbiol. Biomanuf.2022221723110.1007/s 43393-021-00052-w · doi ↗
- 2De Santis S. Cavalcanti E. Mastronardi M. Jirillo E. Chieppa M. Nutritional keys for intestinal barrier modulation Front. Immunol.2015661210.3389/fimmu.2015.0061226697008 PMC 4670985 · doi ↗ · pubmed ↗
- 3Thursby E. Juge N. Introduction to the human gut microbiota Biochem. J.20174741823183610.1042/BCJ 2016051028512250 PMC 5433529 · doi ↗ · pubmed ↗
- 4Gill S.R. Pop M. De Boy R.T. Eckburg P.B. Turnbaugh P.J. Samuel B.S. Gordon J.I. Relman D.A. Fraser-Liggett C.M. Nelson K.E. Metagenomic analysis of the human distal gut microbiome Science 20063121355135910.1126/science.112423416741115 PMC 3027896 · doi ↗ · pubmed ↗
- 5Jandhyala S.M. Talukdar R. Subramanyam C. Vuyyuru H. Sasikala M. Reddy D.N. Role of the normal gut microbiota World J. Gastroenterol. WJG 201521878710.3748/wjg.v 21.i 29.878726269668 PMC 4528021 · doi ↗ · pubmed ↗
- 6Adak A. Khan M.R. An insight into gut microbiota and its functionalities Cell. Mol. Life Sci.20197647349310.1007/s 00018-018-2943-430317530 PMC 11105460 · doi ↗ · pubmed ↗
- 7Backhed F. Ley R.E. Sonnenburg J.L. Peterson D.A. Gordon J.I. Host-bacterial mutualism in the human intestine Science 20053071915192010.1126/science.110481615790844 · doi ↗ · pubmed ↗
- 8Rinninella E. Raoul P. Cintoni M. Franceschi F. Miggiano G.A.D. Gasbarrini A. Mele M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases Microorganisms 201971410.3390/microorganisms 701001430634578 PMC 6351938 · doi ↗ · pubmed ↗