Probiotic and Postbiotic Interactions of Lactobacillus Strains with Candida albicans: Antifungal Effects Through Microbial Competition
Andrea Vega-Vásconez, Diana Lucinda Castillo-Patiño, Javier Alberto Garza-Cervantes, Arlette Santacruz, José Rubén Morones-Ramírez

TL;DR
This study explores how different forms of lactobacillus bacteria and their byproducts can inhibit the growth of Candida albicans, a harmful fungus, through various mechanisms.
Contribution
The study systematically compares the antifungal effects of live probiotics, heat-inactivated postbiotics, and cell-free supernatants of lactobacillus strains against Candida albicans.
Findings
Lactobacillus plantarum 299V showed the strongest antifungal effect, reducing Candida albicans viability by 2.39 log10 CFU/mL.
Antifungal activity of Lactobacillus acidophilus was largely pH-dependent, with inhibition dropping from 79.01% to 28.35% when pH was neutralized.
Heat-inactivated postbiotics inhibited fungal growth up to 95.14%, but coaggregation with fungal cells may affect measurement accuracy.
Abstract
Background: Candida albicans is the most clinically significant opportunistic fungal pathogen, and the growing resistance to conventional antifungals, particularly azoles and echinocandins, highlights the urgent need for alternative therapeutic strategies. Although lactic acid bacteria (LAB) have shown inhibitory potential against C. albicans, the relative contributions of live probiotics, heat-inactivated postbiotics, and cell-free supernatants (CFSs) have rarely been compared in parallel under physiologically relevant conditions against a clinical oral isolate. Results: This study systematically evaluated the antifungal activity of Lactiplantibacillus plantarum 299V, Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842, and Lactobacillus acidophilus ATCC 4356 using co-culture assays, minimum inhibitory concentration tests, agar well diffusion assays, and optical microscopy. L.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
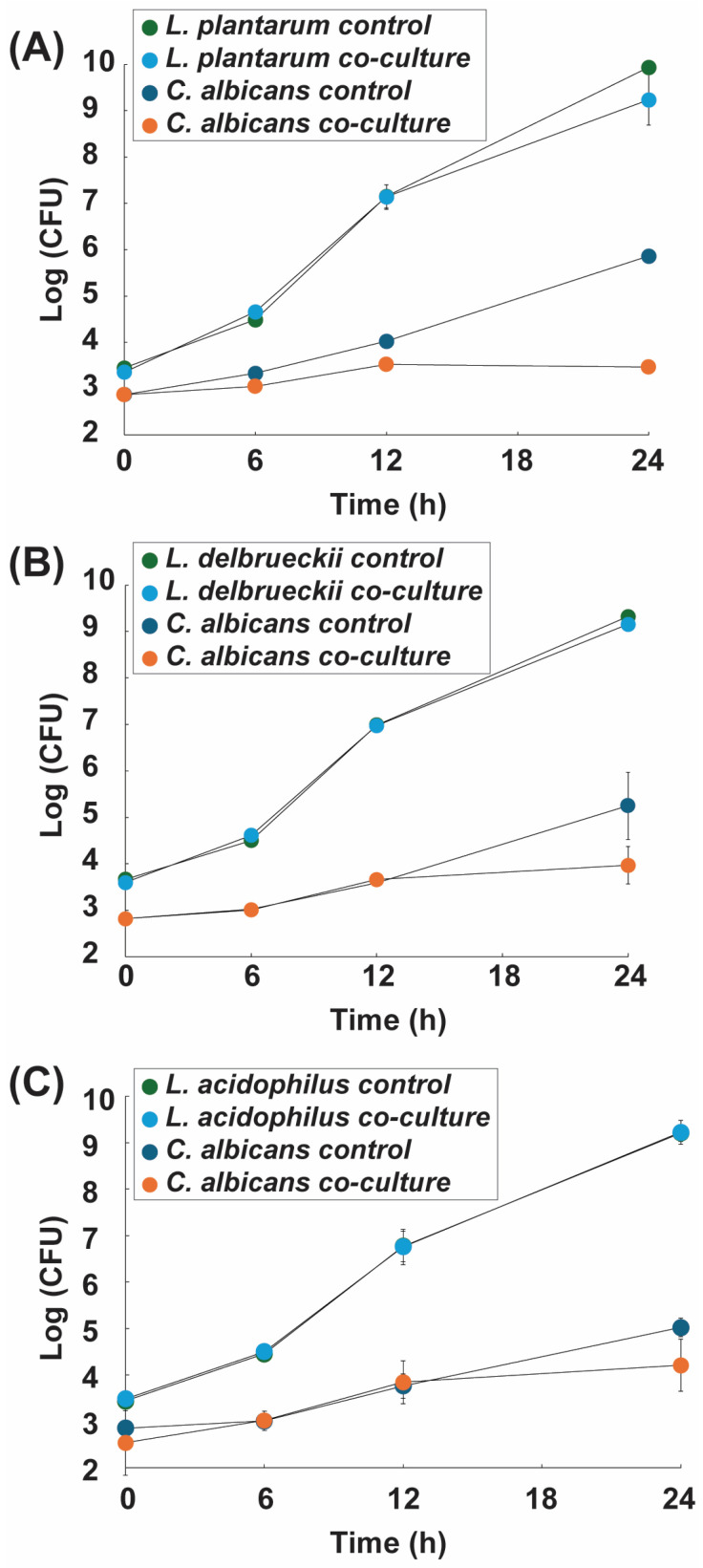 Figure 1
Figure 1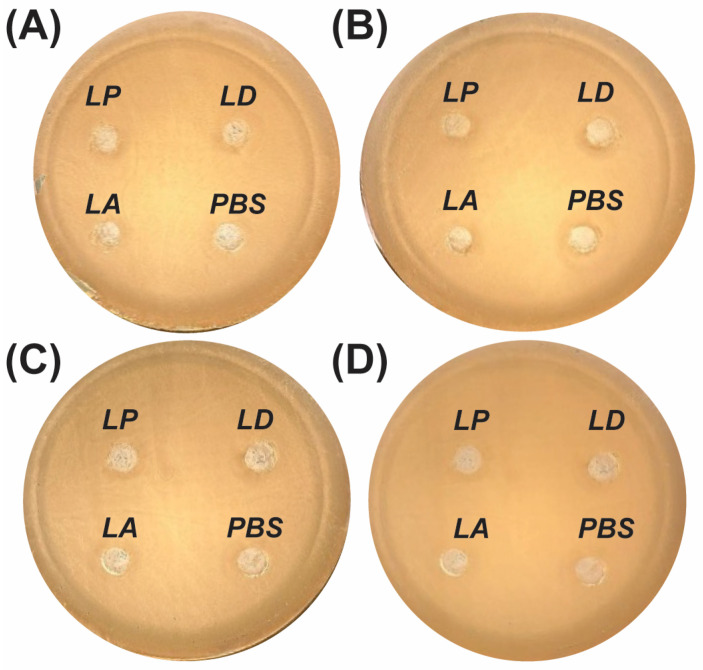 Figure 2
Figure 2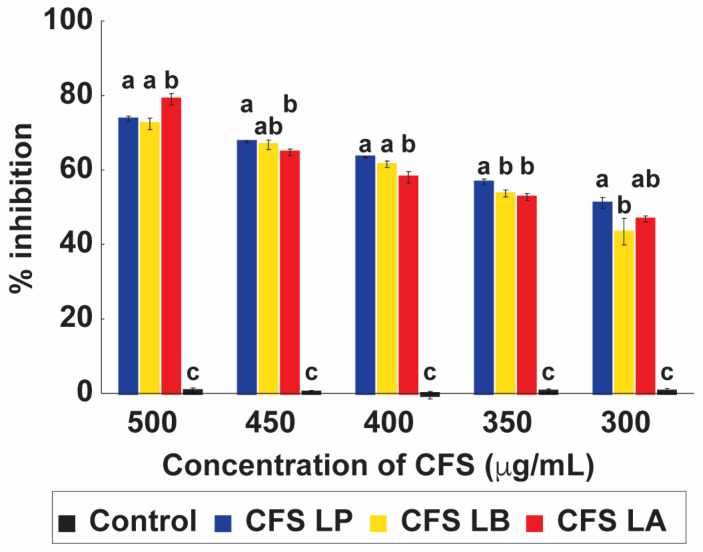 Figure 3
Figure 3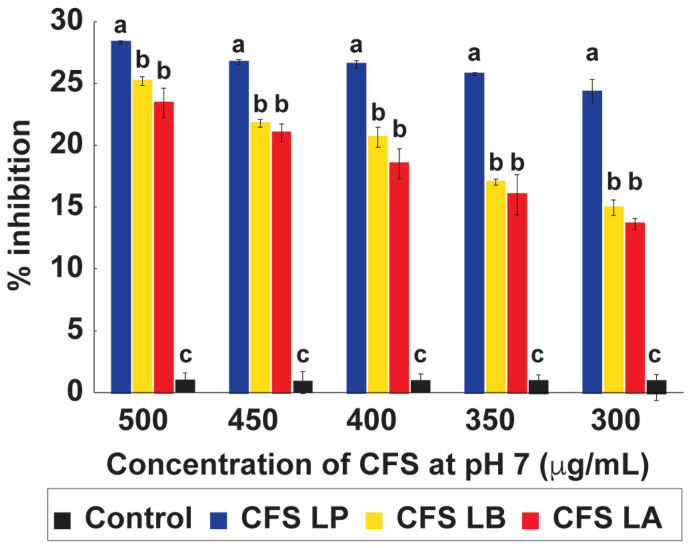 Figure 4
Figure 4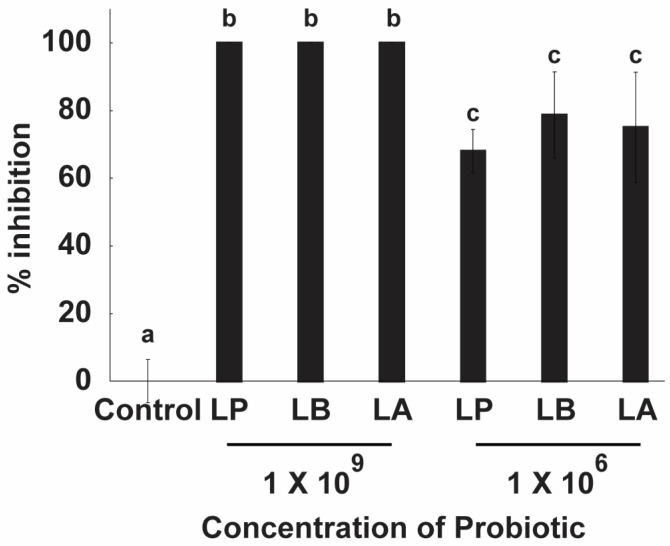 Figure 5
Figure 5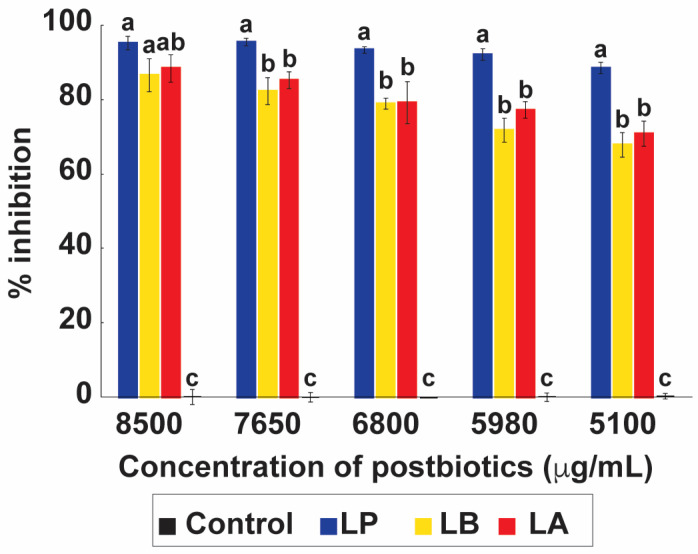 Figure 6
Figure 6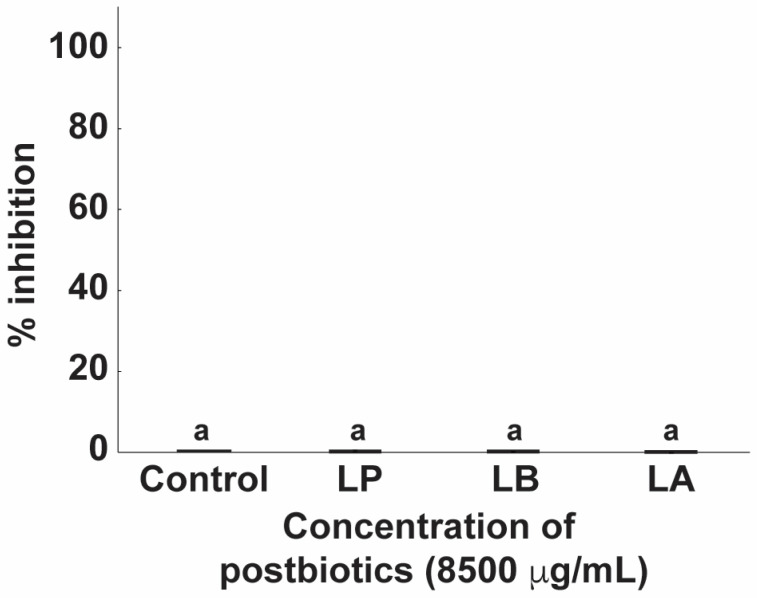 Figure 7
Figure 7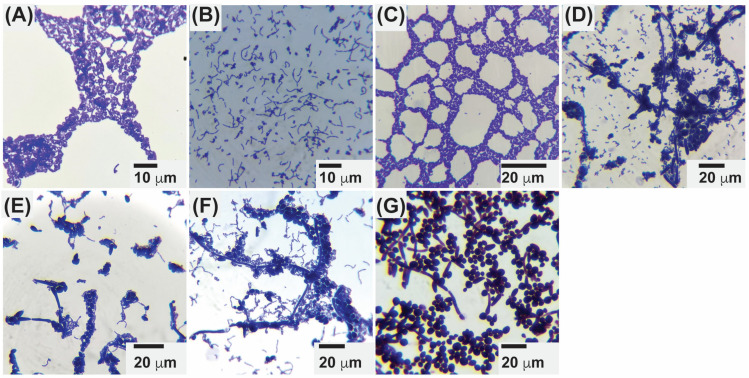 Figure 8
Figure 8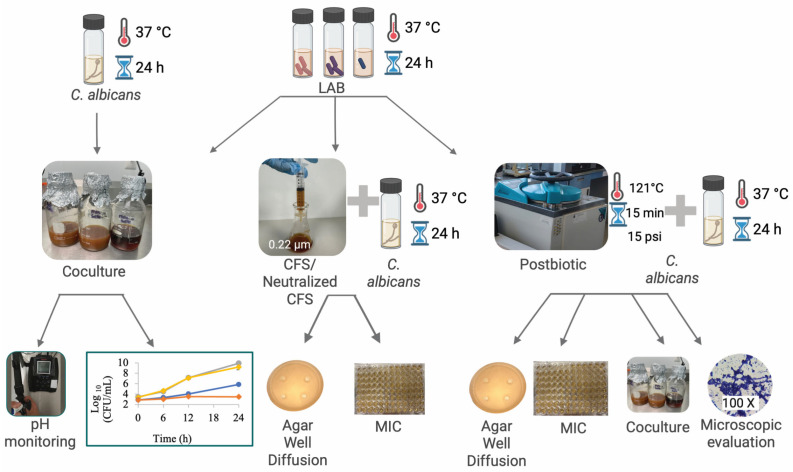 Figure 9
Figure 9- —Universidad Autonoma de Nuevo León and CONACyT
- —Basic science
- —Fronteras de la Ciencia
- —Infraestructura
- —Apoyos a la Ciencia de Frontera
- —Grant a Ciencia de Frontera
- —Beca Nacional de Posgrado from CONAHCyT
- —Becas Nacionales de Postdoctorado from CONAHCyT
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Microbial Metabolites in Food Biotechnology
1. Introduction
Fungal infections represent a major global health burden, responsible for an estimated 1.7 million deaths each year and disproportionately affecting immunocompromised individuals, including patients with diabetes, cancer, or HIV infection [1]. Among opportunistic fungal pathogens, Candida albicans remains the most clinically significant species, responsible for infections that range from superficial mucosal candidiasis to invasive and life-threatening systemic disease [2]. As a polymorphic commensal yeast, C. albicans colonizes the oral cavity, gastrointestinal tract, vaginal mucosa, and skin under normal physiological conditions [3]. However, impairment of host immune defenses or disruption of mucosal barriers can promote its transition from commensal organism to invasive pathogen, characterized by filamentation, tissue invasion, and biofilm development [3,4]. The rising prevalence of antifungal resistance, particularly to azoles and echinocandins, together with adverse drug effects and limited therapeutic options, highlights the urgent need for novel and safer antifungal strategies [5].
In this context, LAB, including Lactiplantibacillus plantarum, Lactobacillus acidophilus, and Lactobacillus delbrueckii subsp. bulgaricus, have emerged as promising biotherapeutic candidates against C. albicans [6,7,8]. These microorganisms are classified as Generally Recognized as Safe and are widely used in fermented foods and probiotic formulations. They naturally colonize gastrointestinal, oral, and urogenital niches [9,10,11]. Beyond their nutritional applications, LAB contribute to mucosal homeostasis through competition with pathogens for nutrients and adhesion sites, reinforcement of epithelial barrier integrity, and modulation of innate and adaptive immune responses [9,11]. Their antifungal activity is mediated by multiple complementary mechanisms, including the production of organic acids, hydrogen peroxide, and bacteriocins that reduce environmental pH and create unfavorable conditions for fungal survival [9]. These metabolites may also interfere with fungal adhesion, morphogenesis, and biofilm formation, thereby enhancing colonization resistance [8,12].
More recently, increasing attention has been directed toward postbiotics, defined as inanimate microorganisms or their bioactive metabolic products, including cell-free supernatants, as alternative therapeutic agents [10]. Postbiotics can exert antimicrobial, immunomodulatory, and anti-inflammatory effects comparable to those of live probiotic cells while reducing potential safety concerns associated with the administration of viable bacteria, particularly in immunocompromised hosts [13]. Despite promising findings, significant knowledge gaps remain regarding the comparative antifungal efficacy of live probiotics, postbiotics, and derived supernatants. Many available studies evaluate single strains, limited experimental endpoints, or reference vaginal isolates of C. albicans under simplified laboratory conditions [7,8,14]. Furthermore, strain variability, environmental parameters such as pH and nutrient composition, and time-dependent interaction dynamics complicate mechanistic interpretation.
To address these limitations, the present study systematically investigated the antagonistic interactions of L. plantarum, L. acidophilus, and L. delbrueckii subsp. bulgaricus against a clinical oral isolate of C. albicans. Using supplemented MRS broth as a competitive model system, inhibition dynamics were characterized through coculture assays, quantitative viable cell enumeration, pH measurement, and microscopic analysis. The antifungal potential of native and pH-neutralized cell-free supernatants, as well as heat-inactivated postbiotic preparations, was further evaluated using agar diffusion and minimum inhibitory concentration assays. By systematically comparing probiotics, postbiotics, and CFS under physiologically relevant conditions, this study provides a mechanistic framework for understanding strain-specific LAB–Candida interactions and offers a comparative foundation for the rational design of probiotic and postbiotic-based antifungal interventions.
2. Results and Discussion
2.1. Co-Culture
The inhibitory activity of LAB strains against Candida albicans was evaluated in co-culture over 24 h (Figure 1A–C). Growth curves revealed that C. albicans growth was markedly reduced in the presence of LAB, with inhibition becoming evident after 12 h. By 24 h, Lactiplantibacillus plantarum achieved the strongest effect, reducing C. albicans viability by 2.39 log_10_ CFU/mL, corresponding to approximately 99% decrease in fungal viability, followed by Lactobacillus delbrueckii (1.28 log_10_ CFU/mL) and Lactobacillus acidophilus (0.81 log_10_ CFU/mL). The reductions observed for L. delbrueckii and L. acidophilus fall within the 1–2 log_10_ range considered biologically relevant probiotic- Candida interaction models [15]. In contrast, all LAB strains maintained robust growth in co-culture, with final cell densities comparable to their monoculture controls [16,17]. The reductions in C. albicans viability observed in co-culture were statistically significant compared with monoculture controls (one-way ANOVA followed by Tukey’s post hoc test, p < 0.05).
This inhibitory effect coincided with a progressive decline in culture pH, which reached 3.94 for L. plantarum, 4.27 for L. delbrueckii, and 4.31 for L. acidophilus after 24 h (Figure S1A–C). Acidification likely enhanced antifungal activity by facilitating the diffusion of undissociated lactic acid across the C. albicans cell membrane, leading to intracellular acidification, disruption of homeostasis, and increased sensitivity to oxidative stress [10,18]. Under acidic conditions, C. albicans activates RIM101-dependent transcriptional responses and stress regulators such as HOG1, which are required for adaptation to low-pH environments [19,20]. Sustained acid stress increases the energetic demand needed to maintain intracellular pH homeostasis, thereby limiting fungal growth [21]. This growth limitation is consistent with activation of pH-stress regulatory pathways and the increased metabolic burden required to preserve intracellular pH balance under weak-acid conditions, collectively constraining fungal proliferation [19,20,21].
Although our study did not directly assess gene expression, previous work has demonstrated that L. plantarum in co-culture with C. albicans upregulates genes linked to environmental sensing and adhesion, while C. albicans downregulates virulence-associated genes, including those linked to hyphal formation and adhesion, such as ALS3, HWP1, CPH1, and BCR1. These findings suggest that beyond acidification, competitive interactions may involve transcriptional reprogramming in both organisms, consistent with the stronger inhibitory effect observed for L. plantarum [8]. These findings highlight that antifungal inhibition in co-culture likely involves direct competition and localized metabolic effects, which may not be fully captured by diffusion-based solid assays.
2.2. Agar Well Diffusion
2.2.1. Inhibition from Postbiotic
No inhibition zones were detected when postbiotics derived from L. plantarum, L. delbrueckii, or L. acidophilus were tested against C. albicans (Figure 2A). Previous studies have reported inhibition zones of 11.5–16.5 mm against several bacterial strains using heat-inactivated L. acidophilus IFFI 6005 [22]. Similarly, heat-inactivated L. delbrueckii subsp. bulgaricus NCDO 2394 produced inhibition zones against E. coli and K. pneumoniae [23]. These discrepancies likely reflect key methodological differences between studies, including the target microorganism (bacterial strains vs. C. albicans), fermentation scale and postbiotic concentration steps, heat-inactivation protocols, agar composition and well diameter, inoculum density, and incubation conditions. Such variables can substantially influence diffusion-based inhibition outcomes.
2.2.2. Inhibition from Live LAB
Consistent with the postbiotic results, no inhibition of C. albicans by live LAB strains was observed in our agar well diffusion assays (Figure 2B). This finding differs from the literature, where inhibition halos of 6–10 mm were observed for L. plantarum using the overlay method [8]. Likewise, other research groups have observed a 19 mm inhibition zone with L. paracasei 28.4 [17]. However, Yocheva et al. reported that even different strains within the same species may not exhibit inhibitory activity against the same C. albicans strain [7]. Since comparable overlay methodologies were employed across studies, strain-dependent variability in antimicrobial metabolite production most likely accounts for the absence of inhibition observed in the present study.
2.2.3. Inhibition from CFS and Neutralized CFS (pH 7)
Neither the native nor the pH-neutralized cell-free supernatants (CFSs) from L. plantarum, L. delbrueckii, or L. acidophilus exhibited inhibitory activity against C. albicans in the agar diffusion assay (Figure 2C,D). Previous studies have similarly reported limited or absent antifungal activity of LAB-derived CFS when assessed using diffusion-based methods [24]. LAB-derived CFS are known to contain diverse bioactive metabolites—including organic acids, hydrogen peroxide, phenyllactic acid, and, in some strains, bacteriocin-like compounds—that may contribute to antifungal effects under appropriate conditions [25,26]. However, the agar well diffusion method presents inherent limitations, particularly for slowly diffusing or high–molecular-weight molecules, which may fail to reach inhibitory concentrations at the fungal growth front [17,27,28]. Because the CFS was not chemically fractionated or metabolically characterized in the present study, the discussion of specific metabolites remains inferential and literature-based. The inhibitory activity observed in liquid-phase MIC assays, therefore, supports the interpretation that the absence of inhibition zones in agar diffusion reflects methodological constraints rather than the absence of antifungal potential.
2.3. Minimum Inhibitory Concentration
2.3.1. Anti-Candida Activity of CFS
The cell-free supernatants (CFSs) from L. plantarum, L. delbrueckii, and L. acidophilus all inhibited C. albicans growth in a dose-dependent manner across the concentration range of 500–300 µg mL^−1^ (Figure 3). At the highest concentration (500 µg mL^−1^), CFS from L. acidophilus exhibited the strongest inhibition (79.01%), followed by L. plantarum (73.72%) and L. delbrueckii (72.41%). Although inhibitory activity declined with decreasing concentrations, significant inhibition was still observed at 300 µg mL^−1^ for all three strains (p < 0.05). Statistical analysis confirmed that differences between treatments were strain- and concentration-dependent.
These results are in line with earlier reports, such as the 74.92% inhibition documented for L. plantarum CFS against C. albicans, though strain-specific variability in antifungal activity has been widely noted [8]. The observed inhibition is likely attributable to organic acids, particularly lactic acid, which are abundant metabolites in LAB supernatants. Supporting this interpretation, prior studies demonstrated that lactic acid at 200 mM reduces both hyphal formation and viability in C. albicans, underscoring its dual role in limiting fungal morphology and survival [29]. Importantly, these effects depend on an acidic environment, as undissociated lactic acid can diffuse across the fungal membrane more effectively under low pH, leading to intracellular acidification and loss of homeostasis [30]. Collectively, these findings reinforce the role of LAB-derived organic acids as central mediators of antifungal activity in CFS preparations.
2.3.2. Anti-Candida Activity of CFS pH 7
The pH-neutralized CFS from L. plantarum, L. delbrueckii, and L. acidophilus retained significant inhibitory activity against C. albicans across all concentrations tested (500–300 µg mL^−1^), with maximum inhibition values of 28.35%, 25.21%, and 23.43%, respectively (Figure 4). Although activity was lower than that observed under acidic conditions (Figure 3), inhibition remained consistent and strain-dependent. These results are comparable to the literature, where inhibition of approximately 33% for L. fermentum 18 A-TV when its CFS was adjusted to pH 7.5 has been observed [18].
The reduced antifungal effect at neutral pH likely reflects the dissociation state of lactic acid, one of the principal metabolites produced by LAB. Under acidic conditions, lactic acid remains largely undissociated, enabling efficient diffusion across the fungal cell membrane [11,31,32]. Once inside the cell, it dissociates, acidifying the cytoplasm and disrupting homeostasis. At neutral pH, however, lactic acid exists predominantly in its dissociated form, which is less membrane-permeable, thereby diminishing its antifungal efficacy [10,18].
2.3.3. Anti-Candida Activity of Probiotics
As shown in Figure 5, all probiotic strains—L. plantarum, L. delbrueckii, and L. acidophilus—completely inhibited C. albicans growth at a concentration of 1 × 10^9^ CFU/mL. At a lower concentration of 1 × 10^6^ CFU/mL, inhibition was reduced but remained significant, with L. plantarum achieving 68.09%, L. delbrueckii 78.72%, and L. acidophilus 75.04%. These findings indicate that higher probiotic densities are required for full suppression of fungal growth, while lower densities result in moderate and strain-dependent inhibition, with L. delbrueckii showing the greatest efficacy under reduced concentrations.
Our results differ from the literature, where a 1.5 log-cycle inhibition has been observed using L. plantarum at 1 × 10^8^ CFU/mL in mixed culture with C. albicans and S. mutans at 1 × 10^3^ CFU/mL. The lower inhibition observed in their study may be related to the growth medium (TSBYE) and the additional presence of S. mutans [29]. Conversely, our findings are consistent with research groups that have observed 82.34% inhibition after 24 h of co-culturing L. plantarum with C. albicans in MRS medium supplemented with yeast extract, peptone, and dextrose (YDP) [8].
The inhibitory effect of probiotics appears closely linked to the ratio of LAB to C. albicans cells. Yeast cells (≈6 µm) are considerably larger than bacterial cells (≈1.5 µm), and a higher bacterial density increases the likelihood of coaggregation, enabling LAB to physically cover and interact with fungal cells. This direct interaction enhances both nutrient competition and the local delivery of antifungal metabolites, including organic acids, bacteriocins, biosurfactants, and hydrogen peroxide [33].
Nutrient competition also plays a critical role. LAB have shorter generation times and rapidly deplete essential nutrients such as glucose, thereby restricting availability for C. albicans. This depletion not only limits fungal growth but also alters the culture environment by lowering pH, generating conditions that are less favorable for yeast development and virulence [10,34]. Collectively, these results highlight that both cell density and metabolic competition are central to LAB-mediated inhibition of C. albicans.
2.3.4. Anti-Candida Activity of Postbiotics
The postbiotics derived from L. plantarum (LP), L. delbrueckii (LB), and L. acidophilus (LA) all exhibited a clear concentration-dependent inhibitory effect against C. albicans (Figure 6). At the highest concentration tested (8500 µg mL^−1^), inhibition reached 95.14% for LP, 88.35% for LA, and 86.50% for LB. Although inhibition declined with decreasing concentrations, substantial antifungal activity was still observed at 5100 µgmL^−1^, with LP maintaining 88.43% inhibition. These findings suggest concentration-dependent inhibitory activity in OD-based MIC assays; however, this effect should be interpreted with caution, given the potential interference of coaggregation with optical density measurements. Comparable results have been reported in previous studies. For example, Miao et al. showed that heat-inactivated L. acidophilus IFFI 6005 displayed measurable antimicrobial effects against bacterial pathogens, while Van et al. demonstrated that postbiotics from L. delbrueckii inhibited E. coli and K. pneumoniae [22,23]. Although most of these reports focus on antibacterial activity, our results extend these observations to antifungal activity against C. albicans, highlighting the broader therapeutic potential of postbiotic preparations.
The high inhibition observed with postbiotics in our study may be attributed to multiple factors, including the persistence of bioactive cell wall components (such as S-layer proteins, peptidoglycan, or lipoteichoic acids) and the presence of heat-stable metabolites. Prior studies have shown that S-layer proteins from LAB mediate microbial adhesion and coaggregation [35], processes that could facilitate close interactions with fungal cells even after bacterial inactivation. Furthermore, organic acids and other metabolites may remain active after heat treatment, providing continued antifungal effects [36].
Together, these findings suggest that postbiotic antifungal activity may be assay-dependent and partly indirect, as high inhibition values observed in OD-based MIC assays were not reproduced in co-culture or agar diffusion assays, likely due to coaggregation-mediated interference with spectrophotometric readings. Therefore, the postbiotic data support inhibitory potential under the tested conditions, but do not by themselves establish strong direct antifungal activity. Given their stability and safety advantages over live probiotics, LAB-derived postbiotics represent a promising strategy for antifungal interventions, especially in immunocompromised hosts where live probiotic use may be contraindicated.
2.3.5. Postbiotics in Coculture Assays
In contrast to the strong antifungal activity observed in the MIC assay (Figure 6), no significant inhibition of C. albicans was detected in coculture at the same highest postbiotic concentration (8500 µg/mL; p > 0.05) (Figure 7). This discrepancy may be explained by coaggregation between postbiotics and C. albicans, as observed microscopically (Figure 8). Coaggregation can alter optical density (OD) readings by reducing light scattering relative to individual cells, potentially masking true inhibition when using spectrophotometric methods. Although OD values are commonly used to estimate CFU/mL in microplate assays, the correlation can be influenced by factors such as cell density, size, and light path length [37]. Thus, our results suggest that the lack of measurable inhibition in coculture may reflect methodological limitations rather than the absence of biological activity.
The discrepancy between the MIC and agar well diffusion results can be attributed to fundamental differences between liquid and solid culture systems. In MIC assays, antifungal metabolites are uniformly distributed and maintain continuous contact with C. albicans, allowing consistent exposure. In contrast, in solid agar, compound distribution is spatially restricted, generating concentration gradients that may limit the effective dose reaching fungal cells [38]. Furthermore, pH changes in solid media tend to remain localized around the colony rather than being evenly distributed as in liquid cultures [39,40]. Additionally, lactobacilli can co-aggregate with pathogenic microorganisms, creating localized microenvironments enriched in inhibitory substances [7]. Together, these physicochemical and biological factors may explain why inhibition was observed in MIC assays but not in agar well diffusion tests.
2.4. Microscopic Evaluation of Postbiotic–C. albicans Interactions
Microscopic analysis confirmed clear coaggregation between postbiotics from L. plantarum, L. delbrueckii, L. acidophilus, and C. albicans after 24 h of incubation (Figure 8). These findings are consistent with the literature, where coaggregation of L. plantarum and L. acidophilus has been observed with C. albicans [41], and with other studies reporting L. delbrueckii interactions with Saccharomyces cerevisiae [7].
Coaggregation in LAB is thought to be mediated primarily by S-layer proteins, which interact with microbial cell wall components such as lipids and teichoic acids [35,42]. Removal of these proteins significantly reduces adhesion and coaggregation [43], while S-layer–associated proteins (SLAPs) further contribute to these interactions [44]. Notably, these proteins are highly stable and capable of self-assembly even after heat stress. In our study, probiotics were exposed to 121 °C at 15 psi for 15 min, yet retained coaggregation capacity, suggesting that S-layer proteins remained functional [45].
On the fungal side, C. albicans is also capable of coaggregation through adhesins such as ALS3 and HWP1, which mediate interactions with oral bacteria, including Porphyromonas gingivalis and Streptococcus mutans [46]. Thus, the interactions observed here likely result from the combined contribution of both LAB and C. albicans.
While these physical interactions did not translate into measurable inhibition in coculture assays, they may nonetheless play a role in shaping host–microbe or microbe–microbe dynamics in vivo. Previous work has shown that postbiotics can activate immune receptors and modulate cytokine responses [33]. Although not addressed in the present study, such effects highlight the potential broader benefits of LAB-derived postbiotics beyond direct antifungal activity.
2.5. Strengths and Limitations
Although working with a single isolate naturally limits the generalizability of the findings, the use of a clinical oral isolate of C. albicans represents a notable strength of this study. Unlike standardized reference strains, clinical isolates better reflect the virulence factors, resistance profiles, and adaptive capacity of strains encountered in real patient settings [47,48]. Furthermore, the oral cavity represents an underexplored niche in the context of LAB–C. albicans interactions, as most published work has focused on vaginal or gastrointestinal isolates, making the present study one of the first to evaluate these interactions using an oral clinical strain [49,50,51]. This distinction is relevant given that oral candidiasis presents unique ecological and immunological conditions that differ substantially from other mucosal sites. That said, direct comparison with the existing literature was not always straightforward, and this challenge was further compounded by the fact that, at the time this study was initiated, data on co-culture interactions between C. albicans and LAB were still scarce. While the field has since advanced, certain mechanistic questions remain open [7,8,52]. Although this study offers a comprehensive comparative evaluation through co-culture assays, agar well diffusion, MIC tests, and coaggregation assessment, the specific metabolites responsible for the observed inhibition and the genes involved in LAB–C. albicans interactions were not directly characterized [53,54]. Addressing these molecular aspects through metabolomic and transcriptomic approaches, combined with validation across a broader panel of clinical isolates, represents an important direction for future research.
3. Experimental Section
3.1. Microorganisms and Coculture Conditions
An overview of the experimental workflow, including the integration of co-culture, CFS/neutralized CFS, and postbiotic assays, is presented in Figure 9. Three probiotic strains L. plantarum 299v (DSM 9843, Deutsche Sammlung von Mikroorganismen und Zellkulturen, Braunschweig, Germany), Lactobacillus delbrueckii subsp. bulgaricus (ATCC 11842, American Type Culture Collection, Manassas, VA, USA), and Lactobacillus acidophilus (ATCC 4356, American Type Culture Collection, Manassas, VA, USA)—were obtained from the Nutrionomics Research Group at Tecnológico de Monterrey. We selected the species Lactiplantibacillus plantarum, Lactobacillus acidophilus, and Lactobacillus delbrueckii subsp. bulgaricus as representative, widely used probiotic LAB with documented anti-Candida potential and relevance to oral/gastrointestinal niches, offering complementary functional traits (e.g., organic-acid production, bacteriocin-like activity, and cell-surface adhesion/coaggregation). L. plantarum 299v was selected due to its demonstrated capacity to reduce pathogenic bacteria in the oropharynx and its high production of lactic acid [55,56]. L. acidophilus ATCC 4356 was included based on its inhibitory activity against C. albicans biofilm formation and filamentation, as well as its production of acidocin 4356, a multifunctional bacteriocin with anti-infective capacity through inhibition of virulence factors and biofilm degradation [55,57]. L. delbrueckii was chosen given evidence of its inhibitory activity against Candida spp. through the production of bacteriocins and cell-free supernatants with demonstrated antifungal effects [58]. A clinical oral isolate of C. albicans was provided by Hospital San Vicente (Monterrey, Mexico) and was identified through CHROMagar™ Candida and VITEK^®^ 2 (BioMérieux, Marcy-l’Étoile, France) systems. The isolate was recovered from an oral clinical sample and stored at −70 °C in glycerol stocks; all experiments were performed using freshly revived cultures within five subcultures from the original stock to minimize phenotypic drift and adaptive variation.
Cocultures were established as described in Table S1. Each strain was activated in its respective growth medium (MRS for LAB species and YM for C. albicans) and incubated at 37 °C for 24 h. The resulting cell pellets were harvested by centrifugation (10,000× g for 10 min), washed three times with phosphate-buffered saline (PBS, pH 7.2), and resuspended in a PBS-glycerol solution (18 mL PBS and 2 mL glycerol). The cell suspensions were stored at −70 °C for subsequent use.
For coculture preparation, the activated cultures were added to 250 mL of supplemented MRS broth (Table S2), where each LAB strain was inoculated at 1 × 10^3^ CFU mL^−1^, along with C. albicans at 1 × 10^2^ CFU mL^−1^. These inoculum levels were selected based on previously published LAB–C. albicans interaction models [59]. Higher starting densities (e.g., ≥10^7^ CFU mL^−1^ were avoided because they compress logarithmic growth and reduce the sensitivity of kinetic comparisons [8]. Monocultures of LAB and C. albicans were used as controls, each inoculated at the same concentrations. The cultures were incubated at 37 °C for 24 h, and 30 mL aliquots were collected every 6 h to measure pH, titratable acidity, and microbial growth.
Microbial growth was assessed by plate counting. Serial dilutions of the cultures or cocultures were prepared in buffered peptone water (0.1% bacteriological peptone), and 100 µL aliquots were plated on the appropriate agar. LAB growth in coculture was evaluated on MRS agar with fluconazole (Best Laboratories, 2 mg mL^−1^) to inhibit C. albicans, while C. albicans growth was assessed on YM agar with erythromycin (Alpharma Laboratories, 1 mg mL^−1^) to inhibit LAB. Control cultures were plated on MRS agar for LAB and YM agar for C. albicans, cultured without antibiotics. Prior to coculture experiments, we verified that fluconazole (2 mg mL^−1^) did not affect LAB recovery on MRS agar and that erythromycin (1 mg mL^−1^) did not affect C. albicans recovery on YM agar, ensuring selective enumeration.
The titration of acidity was performed by transferring 10 mL of the culture or coculture to a 250 mL Erlenmeyer flask, followed by the addition of 50 mL of distilled water to ensure proper dilution. Two to three drops of phenolphthalein were added as an indicator, and the solution was titrated with a 1 N NaOH standard solution, added dropwise while stirring continuously. The endpoint was reached when the solution turned a persistent pink color for 30 s, indicating neutralization of the acids in the sample. Lactic acid percentage was calculated using the formula:
V1 = Sample volume;
V2 = NaOH volume used;
N = NaOH normality;
MEq = 0.090 g of lactic acid.
3.2. Preparation of Cell-Free Supernatants (CFSs)
Each LAB strain was cultured in MRS, then centrifuged (10,000× g, 10 min), and the supernatant was filtered (0.22 µm). For neutralized CFS (pH 7), pH was adjusted with NaOH (8 M) before filtration. CFS samples were stored at 5 °C.
3.3. Postbiotic Production
The LAB was prepared as above, autoclaved at 121 °C and 15 psi for 15 min, and resuspended in YM medium at 1.7% (w v^−1^). Viability was confirmed by plating.
3.4. Antifungal Activity Assays
3.4.1. Agar Well Diffusion
Following Yocheva et al. (2024) [7] with modifications, double-layer YM agar was inoculated with C. albicans (1 × 10^7^ CFU mL^−1^), and 6 mm wells were filled with 100 µL of CFS (at native pH or adjusted to pH 7), postbiotics, or LAB suspensions. Plates were incubated at 37 °C for 24 h.
3.4.2. Minimum Inhibitory Concentration
Serial dilutions of antifungal agents (CFS, CFS pH 7, postbiotics) were tested in 96-well microplates at concentrations ranging from 500 to 3000 µg mL^−1^ for CFS and CFS pH 7, and from 8500 to 5100 µg mL^−1^ for postbiotics, in combination with C. albicans (1 × 10^6^ CFU mL^−1^). After 24 h of incubation at 37 °C, OD was measured. For postbiotic evaluation, viable counts were performed at 0 and 24 h using serial dilutions and YM agar plating [46]. For probiotic evaluation, LAB strains were mixed at concentrations of 1 × 10^6^ CFU mL^−1^ or 1 × 10^9^ CFU mL^−1^ with C. albicans (1 × 10^6^ CFU mL^−1^), followed by 24 h of incubation at 37 °C. Growth was then evaluated for both the control and the co-culture with LAB.
3.4.3. Microscopic Evaluation
To evaluate the potential coaggregation between postbiotics derived from postbiotics of LAB and C. albicans, staining was performed using mixed cultures of both organisms. Initially, the samples were heat-fixed onto glass slides, followed by the application of crystal violet for 1 min. The slides were then washed with distilled water and refixed by heat. Finally, the samples were examined under an optical microscope at 100× magnification using mineral oil [7].
3.5. Statistical Analysis
In this study, all treatments and controls were performed in triplicate, with each treatment spaced over time to allow for proper assessment. Data distribution was assessed for normality using the Shapiro–Wilk test prior to statistical analysis. Because the primary objective was to compare treatments within each fixed experimental setting (e.g., among strains at a given concentration/dose within a given assay), groups were defined per assay as the set of strain–concentration conditions compared in that experiment. Accordingly, statistical significance was evaluated using one-way analysis of variance (ANOVA) followed by Tukey’s HSD post hoc test (95% confidence interval) [7]. A p-value < 0.05 was considered statistically significant. While a two-way ANOVA can be used to test main effects and interaction terms (strain × concentration), formal interaction testing was not the primary objective of this work.
4. Conclusions
This study demonstrates that LAB exert antifungal activity against C. albicans through complementary mechanisms, including direct competitive inhibition in co-culture, pH-dependent CFS activity driven by undissociated organic acids, and coaggregation by heat-inactivated postbiotics. Notably, postbiotic inhibition observed in OD-based MIC assays was not confirmed in co-culture or agar diffusion assays, likely reflecting coaggregation-mediated interference rather than direct antifungal activity. Future research should elucidate the molecular mechanisms underlying LAB–Candida interactions and explore the immunomodulatory potential of postbiotics, supporting the development of LAB-based strategies to combat Candida infections and antifungal resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kainz K. Bauer M.A. Madeo F. Carmona-Gutierrez D. Fungal Infections in Humans: The Silent Crisis MIC 2020714314510.15698/mic 2020.06.71832548176 PMC 7278517 · doi ↗ · pubmed ↗
- 2Nikou S.-A. Kichik N. Brown R. Ponde N. Ho J. Naglik J. Richardson J. Candida albicans Interactions with Mucosal Surfaces during Health and Disease Pathogens 201985310.3390/pathogens 802005331013590 PMC 6631630 · doi ↗ · pubmed ↗
- 3Talapko J. JuzbašićM. MatijevićT. Pustijanac E. BekićS. Kotris I. Škrlec I. Candida albicans—The Virulence Factors and Clinical Manifestations of Infection Jo F 202177910.3390/jof 702007933499276 PMC 7912069 · doi ↗ · pubmed ↗
- 4Pappas P.G. Lionakis M.S. Arendrup M.C. Ostrosky-Zeichner L. Kullberg B.J. Invasive Candidiasis Nat. Rev. Dis. Primers 201841802610.1038/nrdp.2018.2629749387 · doi ↗ · pubmed ↗
- 5Soriano A. Honore P.M. Puerta-Alcalde P. Garcia-Vidal C. Pagotto A. Gonçalves-Bradley D.C. Verweij P.E. Invasive Candidiasis: Current Clinical Challenges and Unmet Needs in Adult Populations J. Antimicrob. Chemother.2023781569158510.1093/jac/dkad 13937220664 PMC 10320127 · doi ↗ · pubmed ↗
- 6Huang Z. Zhu J. Bu X. Lu S. Luo Y. Liu T. Duan N. Wang W. Wang Y. Wang X. Probiotics and Prebiotics: New Treatment Strategies for Oral Potentially Malignant Disorders and Gastrointestinal Precancerous Lesionsnpj Biofilms Microbiomes 2025115510.1038/s 41522-025-00688-940199865 PMC 11978799 · doi ↗ · pubmed ↗
- 7Yocheva L. Tserovska L. Danguleva-Cholakova A. Todorova T. Zhelezova G. Karaivanova E. Georgieva R. In Vitro Inhibitory Effects and Co-Aggregation Activity of Lactobacilli on Candida albicans Microbiol. Res.2024151576158910.3390/microbiolres 15030104 · doi ↗
- 8Xu Z. Li Y. Xu A. Xue L. Soteyome T. Yuan L. Ma Q. Seneviratne G. Hong W. Mao Y. Differential Alteration in Lactiplantibacillus Plantarum Subsp. Plantarum Quorum-Sensing Systems and Reduced Candida albicans Yeast Survival and Virulence Gene Expression in Dual-Species Interaction Microbiol. Spectr.202412 e 00353-2410.1128/spectrum.00353-2438717160 PMC 11237386 · doi ↗ · pubmed ↗