Targeting G9a Exerts Pleiotropic Suppression in Triple-Negative Breast Cancer Cells: Cooperatively Inducing Pyroptosis and Apoptosis
Jialin Li, Guijuan Zhang, Tianyang Liu, Xianxin Yan, Min Ma

TL;DR
Blocking G9a in triple-negative breast cancer cells triggers cell death through pyroptosis and apoptosis, offering a new treatment approach.
Contribution
The study reveals a novel mechanism by which G9a inhibition induces pyroptosis via the RIG-I/STAT1/GSDME pathway in TNBC cells.
Findings
Knockdown of G9a suppressed TNBC cell growth, invasion, and migration.
G9a inhibition induced pyroptosis and increased RIG-I, p-STAT1, and GSDME expression.
RIG-I inhibition partially reversed pyroptosis activation caused by G9a knockdown.
Abstract
Background: Pyroptosis, a pro-inflammatory programmed cell death process, is a key player in tumor biology, including in triple-negative breast cancer (TNBC). Inhibiting G9a has been proven to exert anticancer effects; however, the molecular mechanism of the effects remains unclear. The study aimed to illustrate whether inhibiting G9a can suppress the process of TNBC cells by promoting pyroptosis and investigate the underlying mechanisms. Methods: MCF-10A, MDA-MB-231 and SUM159PT cell lines were used for in vitro study. CCK8 and EdU staining assay were used to examine the cell proliferation, and flow cytometry assay was performed to evaluate cell death. Inflammatory factors were measured by ELISA kits. The mRNA and protein expression levels were analyzed by qRT-PCR, Western blot, and immunofluorescence staining. Transmission electron microscopy was used to observe the morphological…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
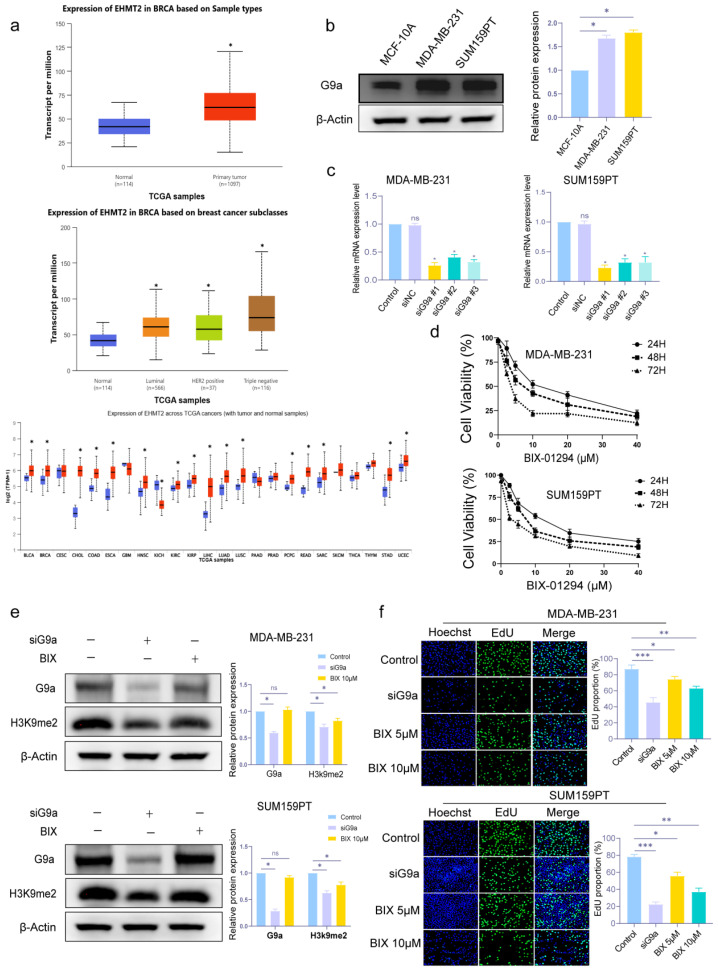 Figure 1
Figure 1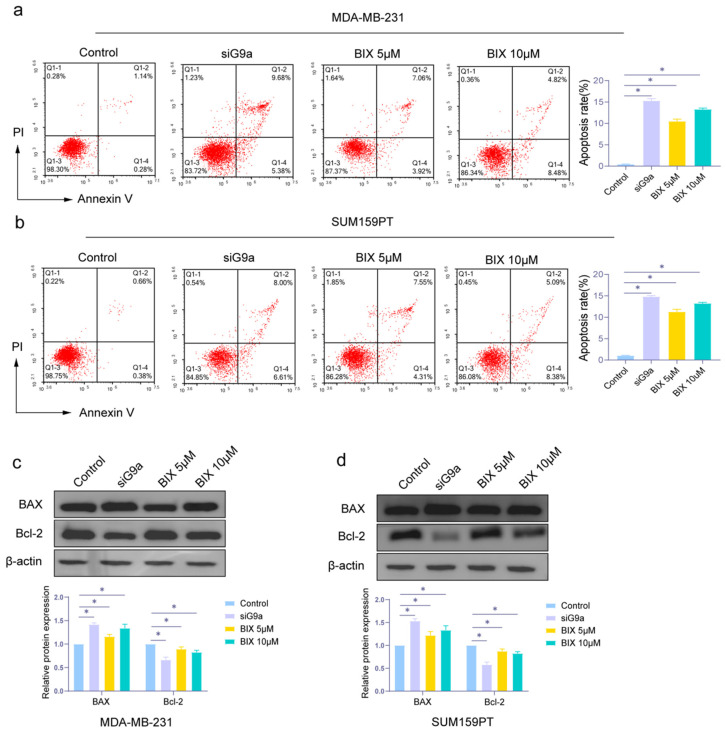 Figure 2
Figure 2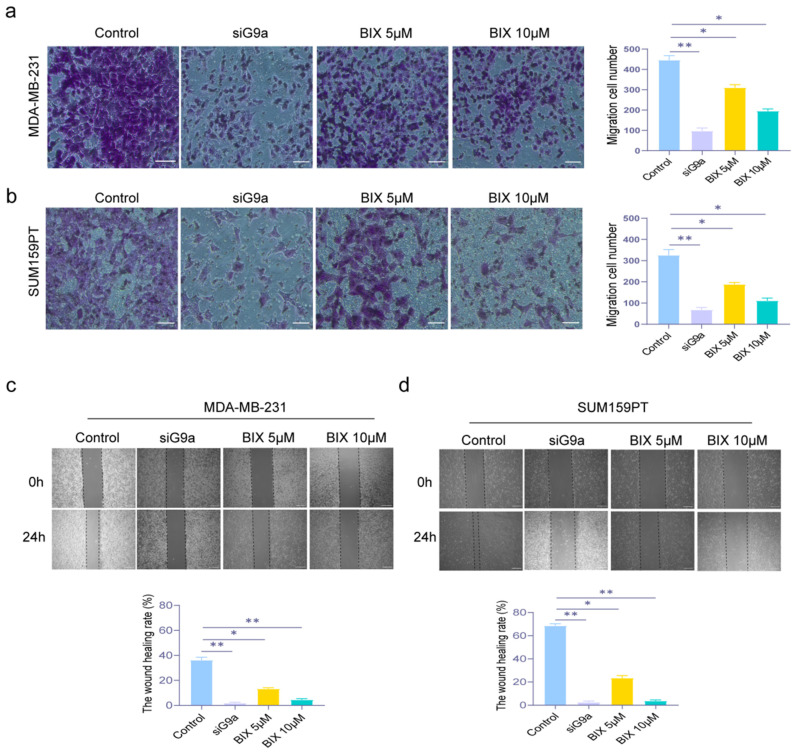 Figure 3
Figure 3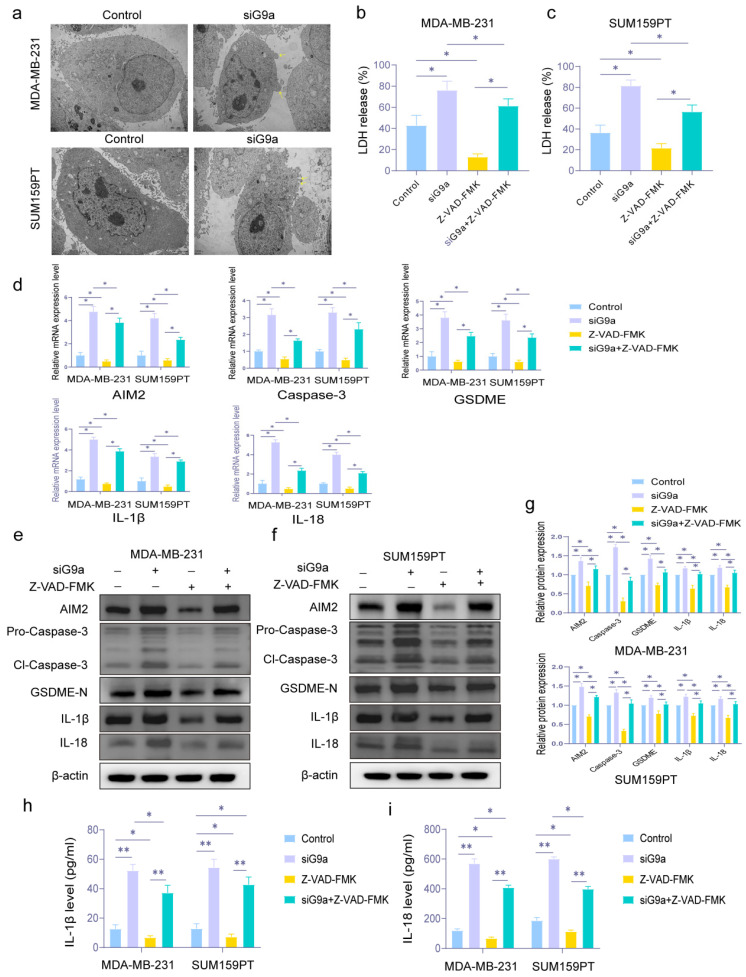 Figure 4
Figure 4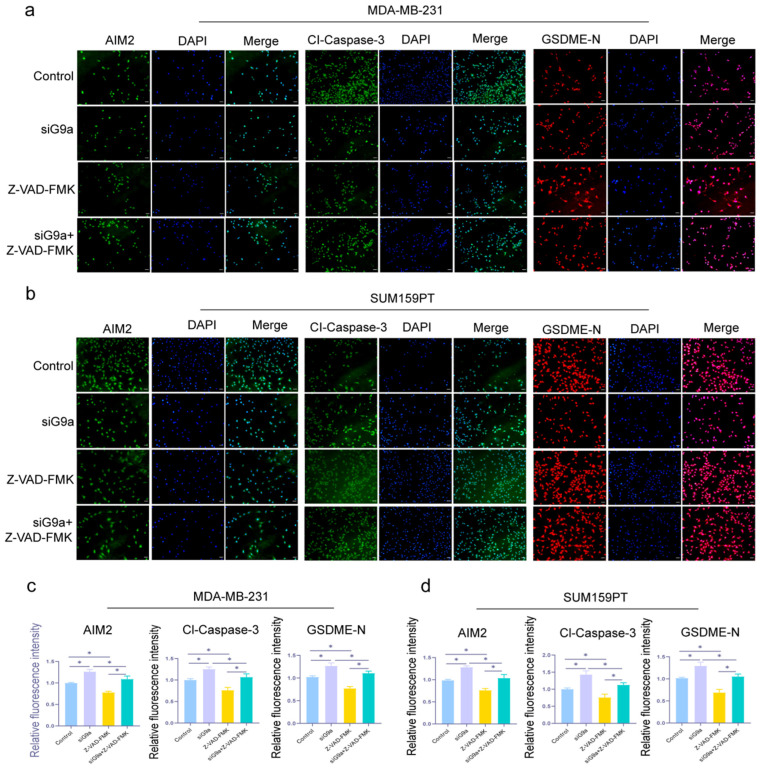 Figure 5
Figure 5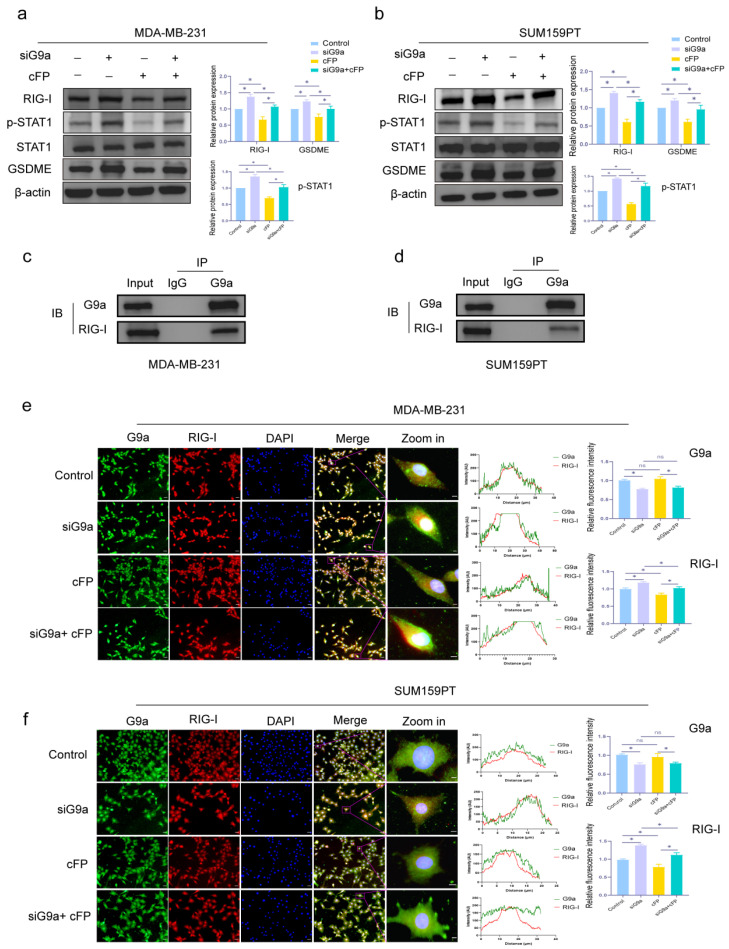 Figure 6
Figure 6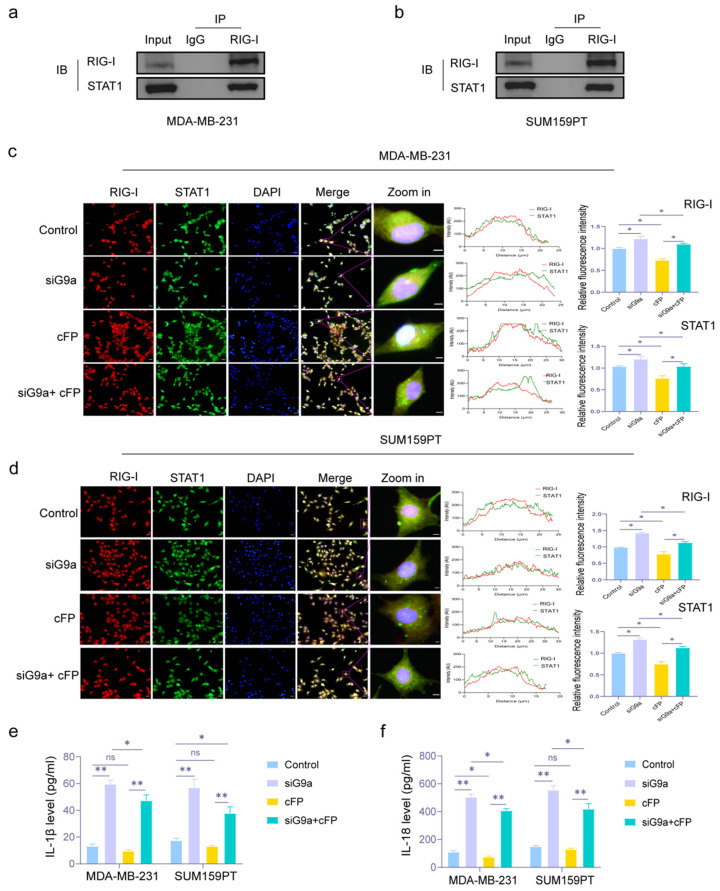 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Guangdong Basic and Applied Basic Research Foundation, China
- —Science and Technology Planning Project of Guangzhou, China
- —Scientific Research Project of Administration of Traditional Chinese Medicine of Guangdong Province of China
- —Fourth Batch of TCM Clinical Outstanding Talent Program of China
- —Fundamental Research Funds for the Central Universities
- —undergraduate innovation and entrepreneurship training program of Guangdong Province
- —Guangdong Basic and Applied Basic Research Foundation in 2026, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammasome and immune disorders · interferon and immune responses · Immune Response and Inflammation
1. Introduction
Breast cancer (BC) is the leading cancer diagnosis among women and still has a high mortality rate in developing and low-income countries [1]. According to three molecular markers, which are known as estrogen receptor (ER), progesterone receptor (PR), as well as human epidermal growth factor receptor 2 (HER2), BC is classified into four types: luminal A, luminal B, HER2-positive, and triple-negative breast cancer (TNBC) [2]. Among them, TNBC is further categorized into six subtypes, which are Basal-Like 1 (BL1), Basal-Like 2 (BL2), Immunomodulatory (IM), Mesenchymal (M), Mesenchymal Stem-Like (MSL), and Luminal Androgen Receptor (LAR) [3]. TNBC accounts for approximately 10–20% of all types but represents an aggressive subtype associated with poor prognosis [1]. Although the therapeutic impact of BC has increased with ongoing treatment updates, patients with TNBC may not respond effectively because of the lack of receptor indicators. For TNBC, there is still a lack of effective therapy so far, and chemotherapy is still the top choice for patients who lost the opportunity for surgery [4]. The prognosis is much worse than that of the other subtypes since fewer patients with TNBC could achieve a full response due to its high invasiveness and recurrence [5,6]. Therefore, seeking safe and efficient methods and therapeutic targets is of great significance for patients with TNBC.
Pyroptosis, a recently identified novel form of programmed cell death mechanism, is related to multiple diseases and plays a significant role in tumors [7,8]. Pyroptosis is executed when the Gasdermin N-terminal forms plasma membrane pores. This results in the massive release of intracellular contents and a potent inflammatory immune response, culminating in this lytic form of cell death [9]. As a member of the Gasdermin family, GSDME is cleaved by Caspase-3. The resulting N-terminal fragment (GSDME-N) then induces pyroptosis [10]. Numerous studies on TNBC indicated that inducing pyroptosis could enhance the efficacy of chemotherapy and radiotherapy, thereby promoting tumor cell death [11,12,13,14].
G9a, also known as EHMT2, is a histone 3 lysine 9 (H3K9) methyltransferase that methylates histone 3 lysine 9 to H3K9me1 and H3K9me2 [15]. It has been discovered that G9a can repress tumor suppressor genes through H3K9 mono-/di-methylation and promote the oncogenes’ functions [16]. Previous studies indicated the role of G9a in tumorigenesis, and G9a is overexpressed in a variety of tumor types, including BC [17,18,19]. In TNBC, G9a knockdown reduces tumor growth and metastasis, suggesting that G9a may be a viable therapeutic target due to its deregulated function [20,21,22]. However, evidence for the role of G9a in TNBC pyroptosis remains scarce, and the underlying molecular mechanisms are poorly understood.
Retinoic acid-induced gene 1 (RIG-I), also known as DDX58, was first identified in 1997 during studies of trans-retinoic acid-induced differentiation in acute promyelocytic leukemia cell lines [23]. Previous studies have shown that RIG-I can regulate the expression of inflammasomes related to pyroptosis [24,25,26] and also takes a great part in cancers [27,28,29,30,31]. Evidence from glioma research suggests a regulatory role for G9a in controlling RIG-I [32].
In this study, we investigated the anti-cancer effects of G9a in TNBC cells, focusing on its role in pyroptosis. We identified the RIG-I/STAT1/GSDME pathway as a potential mechanism through which G9a knockdown induces pyroptosis. These findings position G9a as a promising therapeutic target and offer a novel treatment perspective for TNBC.
2. Materials and Methods
2.1. Cell Culture
MCF-10A, MDA-MB-231, and SUM159PT were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). MCF-10A was cultured in its specific medium (Procell, Wuhan, China). MDA-MB-231 and SUM159PT were cultured in DMEM medium (Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (Procell, Wuhan, China) and 1% penicillin-streptomycin (Gibco, Grand Island, NY, USA). All cells were incubated at 37 °C with 5% CO_2_.
2.2. Reagents
BIX-01294 (HY-108239), Z-VAD-FMK (HY-16658B, purity ≥ 98.78%), and Cyclo (Phe-Pro) (HY-P1934, purity ≥ 98.77%) were purchased from MedChemExpress (MCE, Monmouth Junction, NJ, USA).
2.3. Differential Expression Levels of G9a
The UALCAN database was used to determine the differential expression of G9a in normal and tumor tissues from the TCGA database.
2.4. RNA Interference
The siRNAs used to carry out the G9a knockdown were synthesized by Sangon Biotech (Shanghai, China). Lipofectamine RNAiMAX (Invitrogen, Carlsbad, CA, USA) was used for the transfection following the manufacturer’s instruction. The sequences were as follows: siG9a #1: 5′-AGAAGAAUUCAAGAGAUUCCG-3′; siG9a #2: 5′-AGUAACGGGCAUCAAUGC-3′; siG9a #3: 5′-CCAUGAACAUCGAUCGCAATT-3′.
2.5. Cell Viability Assay
Cells were treated with BIX-01294 (0, 5, 10, 20, and 40 μmol/L) for 24 h, 48 h, and 72 h. Then a Cell Counting Kit-8 (CCK-8) solution (Beyotime, Shanghai, China) was added to each well for another 1 h incubation. The absorbance was measured by a microplate reader (Agilent, Santa Clara, CA, USA) at 450 nm.
2.6. EdU Staining
EdU Staining Kit (C0071S, Beyotime, Shanghai, China) was used to measure the EdU in different treatment groups. Cells were seeded into 96-well plates and treated with different treatments. Subsequently, a 2× EdU working solution was prepared according to the manufacturer’s instructions. Then the working solution was added to each well, followed by an additional incubation for 2 h. After removing the medium, the cells were fixed with 4% paraformaldehyde at room temperature for 15 min. PBS with 0.3% Triton X-100 was then added after the fixation and the mixture was incubated at room temperature for 15 min. After that, a click-reaction solution was added to the washed plates, followed by incubation at room temperature in the dark for 30 min. Finally, Hoechst staining solution was added for nuclear staining for 10 min. A fluorescence microscope (ZEISS, Oberkochen, Germany) was used for observation.
2.7. Flow Cytometric Assay
Adherent cells were collected using trypsin without EDTA (Biosharp, Shanghai, China), washed with PBS, and centrifuged at 2000 rpm for 5 min twice. Then the cells were re-suspended in binding buffer and stained with Annexin V-FITC and PI (Dojindo, Kumamoto, Japan) for 15 min at room temperature in the dark. Then the cells were analyzed with a flow cytometer (NovoCyte, Agilent, CA, USA).
2.8. Transwell Invasion Assay
Cells were re-suspended in medium without FBS and then seeded into 24-well plates. 50 μL of Matrigel (Corning, Corning, NY, USA) mixed with serum-free medium (1:8), and 800 μL of medium with 20%FBS were coated into the upper and lower chamber, respectively. Then 400 μL medium without FBS containing 1 × 10^5^ cells was added into the upper chamber. After incubation for 48 h, each insert was fixed and stained with 4% paraformaldehyde and 0.1% crystal violet. A light microscope (ZEISS, Oberkochen, Germany) was used for the observation after the inserts were completely dried.
2.9. Wound-Healing Assay
Once cells reached confluence in 6-well plates, scratches were created by a sterile 200 µL pipette tip. Then the cells were incubated for 24 h at 37 °C with serum-free medium. An inverted microscope (ZEISS, Oberkochen, Germany) was used to detect the width of the scratch, which was measured at 0 h and 24 h.
2.10. ELISA and LDH Release Assay
The levels of IL-1β and IL-18 were measured by using a Human IL-1β ELISA Kit (JL13662, Jianglai Biotechnology, Shanghai, China) and a Human IL-18 ELISA Kit (JL19261, Jianglai Biotechnology, Shanghai, China). Cell supernatants from different groups were added to a 96-well plate. After applying a plate sealer, the plate was incubated at 37 °C for 60 min. A biotin-antibody solution was added to each well after the liquid was discarded, and the plate was incubated at 37 °C for an additional 60 min. After the incubation, the plate was washed three times, and streptavidin-HRP was added, followed by incubation at 37 °C for 30 min. After that, the plate was washed five times, and the TMB substrate was added. It was then incubated at 37 °C in the dark for 15 min. After adding stop solution to each well, the optical density (OD) was measured at 450 nm through a microplate reader (Agilent, CA, USA).
LDH levels were detected by a LDH Release Assay Kit (C0019S, Beyotime, Shanghai, China). After adding cell supernatants from different treatment groups to a 96-well plate, LDH release assay buffer was added to adjust the total volume in each well to 100 µL. The standard and sample wells were then filled with 100 µL of the LDH assay working solution and incubated at 37 °C in the dark for 30 min. After the incubation, 20 µL of stop solution was added to each well, and the absorbance was measured at 450 nm.
2.11. Transmission Electron Microscopy (TEM)
After treating the cells with different intervention factors, 2.5% glutaraldehyde (G1102, Servicebio, Beijing, China) was used for primary fixation overnight at 4 °C, and 1% osmium tetroxide (02602-AB, Ted Pella, Redding, CA, USA) was used for post-fixation for 2 h at 4 °C in the dark. Subsequently, gradient dehydration was carried out, with each step lasting over 20 min. To enable the resin to completely replace the dehydrating agent and penetrate the cells, the dehydrated samples were placed in a mixture of resin and dehydrating agent before being penetrated in pure resin overnight. The samples were then placed into embedding molds filled with fresh resin and polymerized in an oven at 60 to 70 °C for 48 h to form hard resin blocks. Following this, sectioning (UC-7, Lecia, Wetzlar, Germany) and staining were performed. Finally, the samples were observed at transmission electron microscope (H-7500, HITACHI, Tokyo, Japan).
2.12. RT-qPCR Analysis
Cells were seeded in 6-well plates with different treatments. RNA extraction and complementary DNA (cDNA) synthesis were performed according to the manufacturer’s instructions. RNA extraction kit (TCH020, Takara, Kyoto, Japan) and RT Reagent Kit (RR092A, Takara, Kyoto, Japan) were used to extract RNA from the cells and reverse-transcribed into cDNA. TB Green FAST qPCR (RR820A, Takara, Kyoto, Japan) was adopted to detect the quantitative PCR from the 2^−ΔΔCt^ method. The primers were as follows: AIM2: forward 5′-TTGAGAAGAAGGCAAGCCCA-3′, reverse 5′-CGTGAGTATTTACCTCGCGC-3′; Caspase-3: forward 5′-GGACCAAACCTGAAATGTGG-3′, reverse 5′-GACCCTTTGAATTCTGTCCC-3′; GSDME: forward 5′-GGAGGTGCTGGAAGATAGAA-3′, reverse 5′-GTGCGTTGGAGTCCACATTG-3′; IL-1β: forward 5′-CCAGGGACAGGATATGGAGCA-3′; reverse 5′-TTCAACACGCAGGACAGGTACAG-3′; IL-18: forward 5′-CTGCCACCTGCTGCAGTCTA-3′; reverse 5′-TCTACTGGTTCAGCAGCCATCTTTA-3′; β-actin: forward 5′-AGCGAGCATCCCCCAAAGTT-3′; reverse 5′-GGGCACGAAGGCTCATCATT-3′.
2.13. Western Blot Assay
Cells were lysed with RIPA Lysis Buffer (Epizyme, Shanghai, China). And BCA assay (Epizyme, Shanghai, China) was used to detect protein concentration. Then 6–10% SDS-PAGE (Epizyme, Shanghai, China) was used to separate proteins depending on the protein molecular weight. After transferring to PVDF membranes (Millipore, Billerica, MA, USA), the membranes were blocked with 5% skim milk to make the primary antibody a better bind. After incubating with primary antibodies at 4 °C overnight, the membranes were then incubated with secondary antibodies for 1 h at room temperature. ECL solution (Biosharp, Shanghai, China) and ChemiDoc MP (Bio-Rad, Hercules, CA, USA) were used to visualize the membranes. The primary antibodies were as follows: anti-G9a (1:5000, #66689-1-Ig, Proteintech, Wuhan, China), anti-H3K9me2 (1:5000, #39041, Active Motif, Shanghai, China), anti-BAX (1:2000, #T40051, Abmart, Shanghai, China), anti-Bcl-2 (1:2000, #T40056, Abmart, Shanghai, China), anti-AIM2 (1:5000, #20590-1-AP, Proteintech, Wuhan, China), anti-cleaved caspase-3 (1:1000, #9664S, CST, Danvers, MA, USA), anti-GSDME (1:5000, #13075-1-AP, Proteintech, Wuhan, China), anti-IL-1β (1:1000, #PC0812, Abmart, Shanghai, China), anti-IL-18 (1:1000, #M027287, Abmart, Shanghai, China), anti-RIG-I (1:5000, #25068-1-AP, Proteintech, Wuhan, China), anti-STAT1 (1:5000, #10144-2-AP, Proteintech, Wuhan, China), anti-p-STAT1 (1:1000, #28977-1-AP, Proteintech, Wuhan, China), anti-β-actin (1:40,000, #66009-1-Ig, Proteintech, Wuhan, China). The secondary antibodies were as follows: HRP-conjugated Goat Anti-Rabbit IgG(H + L) (1:5000, #SA00001-2, Proteintech, Wuhan, China), HRP-conjugated Goat Anti-Mouse IgG(H + L) (1:5000, #SA00001-1, Proteintech, Wuhan, China).
2.14. Co-Immunoprecipitation (CO-IP)
The protein lysate was divided into three portions: one as the input and the other two were supplemented with IgG (B900620, Proteintech, Wuhan, China) and the primary antibodies, respectively. After incubating overnight, magnetic beads were added to each portion and incubated overnight at 4 °C again. The next day, the lysate was mixed with loading buffer and boiled at 100 °C for 5 min. We then followed the steps as described for Western blot.
2.15. Immunofluorescence (IF) Staining
Cells with different treatments were fixed with 4% paraformaldehyde and penetrated with 0.5%TritonX-100 (IT9100, Solarbio, Beijing, China). After blocking with 5%BSA for 1 h at room temperature, the cells were incubated with primary antibodies overnight at 4 °C. The next day, cells were incubated with secondary antibodies and stained with DAPI (S2110, Solarbio, Beijing, China). Then the samples were observed by a fluorescence microscope (ZEISS, Oberkochen, Germany). The primary antibodies were as follows: anti-AIM2 (1:200, #PH11499, Abmart, Shanghai, China), anti-caspase-3 (1:200, #TA6311, Abmart, Shanghai, China), anti-GSDME (1:200, #P79886, Abmart, Shanghai, China), anti-G9a (1:2000, #66689-1-Ig, Proteintech, Wuhan, China), anti-RIG-I (1:400, #20566-1-AP, Proteintech, Wuhan, China), anti-STAT1 (1:400, #10144-2-AP, Proteintech, Wuhan, China). The secondary antibodies were as follows: CoraLite594-conjugated Goat Anti-Rabbit IgG(H + L) (1:500, #SA00013-4, Proteintech, Wuhan, China), CoraLite488-conjugated Goat Anti-Mouse IgG(H + L) (1:500, #SA00013-1, Proteintech, Wuhan, China).
2.16. Statistical Analysis
All data are presented as the mean ± standard deviation (SD) from three independent experiments. A normality test was performed on the data distribution. Student’s t-test and one-way ANOVA were performed to compare two or more groups, p values less than 0.05 were regarded as statistically significant. GraphPad Prism software 8.0 was used to analyze data.
3. Results
3.1. Knockdown of G9a and Its Inhibitor Suppress Cells Proliferation
G9a can catalyze H3K9me2 [15], and numerous investigations have shown that G9a is overexpressed in BC [18,19]. In Figure 1a, we detected G9a expression in BC and normal tissues from the TCGA database, and it showed that G9a expression is significantly up-regulated in breast cancer tissues compared to normal breast tissues, especially in TNBC. To validate this finding, we detected the levels of G9a by Western blot. Consistent with the TCGA data, G9a expression was higher in TNBC cell lines (MDA-MB-231 and SUM159PT) than in the normal breast epithelial cell line MCF-10A (Figure 1b). Then we used siRNAs to knock G9a down. Among all siRNAs, siG9a #1 showed the highest efficiency and was employed in the following experiments (Figure 1c). CCK8 assay showed that BIX-01294 (BIX), the inhibitor of G9a, could inhibit the proliferation of TNBC cells in a time- and dose-dependent manner (Figure 1d), and the IC50 values in MDA-MB-231 were 12.65 μmol/L, 7.69 μmol/L, and 3.62 μmol/L for 24 h, 48 h, and 72 h, while the corresponding values in SUM159PT were 11.84 μmol/L, 7.26 μmol/L, and 3.19 μmol/L.
We could also find that G9a knockdown can inhibit the expression of G9a and the methylation of H3K9me2, and BIX can suppress the methylation of H3K9me2 but has no significant effect on the expression of G9a (Figure 1e) (* p < 0.05). In EdU assay (Figure 1f), both G9a knockdown and BIX groups significantly reduced EdU proportion compared to the control group (* p < 0.05, ** p < 0.01, *** p < 0.001). These results showed that G9a knockdown and its inhibitor BIX suppressed cell proliferation, whereas BIX influenced the catalytic effect of G9a instead of its expression.
3.2. Knockdown and Inhibition of G9a Promote Apoptosis
In order to investigate the influence of G9a in cell apoptosis, we used flow cytometric analysis to evaluate. As shown in Figure 2a,b, apoptosis rate was increased upon G9a knockdown in TNBC cells, and the pharmacological inhibition of G9a activity also induced apoptosis. The expression levels of BAX (a pro-apoptotic protein) and Bcl-2 (an anti-apoptotic protein) [33] were measured by Western blot assay. As can be seen in Figure 2c,d, BAX had higher expression in the knockdown group and the BIX group, compared with the control group. The results demonstrated that inhibition of G9a function could induce cell apoptosis in TNBC cells.
3.3. Knockdown and Inhibition of G9a Enhance the Suppression of Invasion and Migration in TNBC Cells
The potential to metastasize is a crucial trait of malignant tumors, including TNBC, which is known for its high invasiveness and recurrence. To evaluate the two critical processes in metastasis—cell invasion and migration—we employed the Transwell (for invasion) and Wound Healing (for migration) assays. In Figure 3a,b, Both G9a knockdown and BIX groups significantly reduced the number of invasive cells compared to the control group. And there was the same trend in Figure 3c,d, that knockdown and inhibition of G9a resulted in lower wound healing rates relative to the control group (* p < 0.05, ** p < 0.01). The results indicated that G9a knockdown and BIX inhibited the invasion and migration abilities in TNBC cells.
3.4. G9a Affects Pyroptosis in TNBC Cells MDA-MB-231 and SUM159PT
In terms of pyroptosis, we used TEM to observe the changes in cell membranes. It is generally recognized that the most obvious morphological alteration of pyroptosis is the swelling and blebbing of the cell membrane. As shown in Figure 4a, knockdown of G9a in MDA-MB-231 and SUM159PT exhibited membrane blebbing.
LDH is also released through membrane permeabilization during pyroptosis. In Figure 4b,c, when G9a was knocked down, the level of released LDH increased. We also used Z-VAD-FMK, known as an inhibitor of pyroptosis, for secondary verification. The Z-VAD-FMK group decreased LDH release and reduced the rise in LDH brought on by G9a knockdown (* p < 0.05).
Considering the major proteins of pyroptosis (AIM2, Cl-Caspase-3, GSDME-N, IL-18, and IL-1β), we estimated the mRNA level and protein expression by qRT-PCR and Western blot. In Figure 4d, we could see that the knockdown group had greater relative mRNA expression levels of genes related to pyroptosis. As above, the addition of the pyroptosis inhibitor reversed the pro-pyroptosis effect of G9a (* p < 0.05). The expression level of these proteins, shown in Figure 4e to g, also had the same performance as the result of qRT-PCR (* p < 0.05).
ELISA assay was used to test the levels of IL-18 and IL-1β. As shown in Figure 4h,i, the levels of IL-18 and IL-1β in TNBC cells were increased in the knockdown group. These trends were consistent with the Western blot results. (* p < 0.05, ** p < 0.01). Similarly, pyroptosis inhibitors had the potential to counteract this impact.
We also examined the three proteins (AIM2, Cl-Caspase-3, and GSDME-N) by IF assay, and the results confirmed the tendencies shown in the previous experiments. In Figure 5, these proteins had higher fluorescence intensity in the knockdown group versus the control group (* p < 0.05). The results above suggested that G9a played a role in the process of pyroptosis, and pyroptosis was induced when the function of G9a had been suppressed in TNBC cells.
3.5. G9a Regulates Pyroptosis in TNBC Cells via RIG-I/STAT1/GSDME Signaling Pathway
RIG-I has been implicated in cancer development and progression [34,35,36], and also plays a role in pyroptosis [25,37]. As shown in Figure 6a,b, the expression of RIG-I, p-STAT1, and GSDME increased in G9a knockdown group, and decreased in the Cyclo (Phe-Pro) (cFP) group. Moreover, when cFP was added to the cells which had been knocked G9a down, the promoting effect of GSDME caused by inhibiting G9a was partially reversed (* p < 0.05).
To explore if G9a could interact with RIG-I, we performed CO-IP assay. In Figure 6c,d, the results showed that G9a and RIG-I might interact in MDA-MB-231 and SUM159PT. The IF assay in Figure 6e,f also revealed that the fluorescence intensity of RIG-I was higher in the knockdown group, and the promoting effect was diminished with the addition of cFP (* p < 0.05).
Then we also tested whether RIG-I and STAT1 interacted. CO-IP assay showed that there was an interaction between RIG-I and STAT1 (Figure 7a,b). In Figure 7c,d, we also observed that G9a knockdown increased the fluorescence intensity and co-localization (yellow spot) of RIG-I and STAT1, suggesting their enhanced interaction. And cFP could partially reverse this effect (* p < 0.05). Then we estimated the expression of IL-1β and IL-18 by ELISA. In Figure 7e,f, the levels of IL-1β and IL-18 were increased in the siG9a group but reduced in the cFP group. cFP effectively reduced the up-regulation of the two pyroptosis-related inflammatory factors induced by G9a knockdown (* p < 0.05).
The above results revealed that G9a regulated pyroptosis in TNBC cells by RIG-I/STAT1/GSDME pathway.
4. Discussion
In this study, we revealed that inhibiting G9a suppresses the proliferation, decreases invasive and migratory capacities, and induces cell death, including apoptosis and pyroptosis in TNBC cells (MDA-MB-231 and SUM159PT). Moreover, we also discover that the RIG-I/STAT1/GSDME pathway may be responsible for the underlying mechanism of G9a involved in the pyroptosis process. We provide the first evidence elucidating how G9a regulates pyroptosis at the cellular level in TNBC cells, thereby revealing a novel mechanistic insight.
Due to a paucity of receptor indicators, TNBC has the worst prognosis among all subtypes of BC, and chemotherapy remains the important option for advanced TNBC [38]. Hence, identifying a therapeutic target is critical for patients with TNBC.
Studies demonstrated that G9a is overexpressed in various tumors and its exhaustion helps to alleviate the malignant manifestations of cancer [39,40,41,42]. In the present study, we found that the level of G9a expression in TNBC cells (MDA-MB-231 and SUM159PT) was higher than that in normal breast cells (MCF-10A). When G9a was knocked down, the abilities of proliferation, invasion, and migration of TNBC cells were reduced, while cell death was promoted. We also noticed that G9a knockdown reduced both G9a expression and H3K9me2 methylation levels, whereas the inhibitor (BIX) reduced H3K9me2 methylation but had no significant effect on G9a expression, which was consistent with the prior work [43].
Pyroptosis is a specific form of cell death characterized by cell swelling, membrane pore formation, and the release of pro-inflammatory cytokines that plays an important role in cancer. Three major pathways have been identified to activate pyroptosis: the canonical pathway mediated by Caspase-1, the non-canonical pathway directed by caspase-4/5/11, and the other pathways mediated by Caspase-3/GSDME and Caspase-8 [44]. GSDME can be cleaved by Caspase-3, and forms holes on cell membranes, causing the loss of membrane integrity and releasing the IL-1β and IL-18, which leads to cell death [45,46]. In BC, GSDME expression was reported downregulated [47]. Li et al. [48] found that Dihydroartemisinin could promote the expression of AIM2, caspase-3, and GSDME led pyroptosis in BC. In this study, we also found that G9a knockdown could stimulate the expression of the pyroptosis-related proteins, and TEM revealed changes in the cell membranes caused by pyroptosis. At the same time, we used an inhibitor of pyroptosis (Z-VAD-FMK), for secondary validation. We were surprised to find that the effects caused by knockdown and inhibitor of G9a could be partially reversed by Z-VAD-FMK. These results confirmed that suppressing the function of G9a could induce pyroptosis in TNBC cells.
RIG-I is a cytoplasmic pattern recognition receptor (PRR) that plays an important role in the innate immune response [26]. A meta-analysis showed that RIG-I was involved in the regulation of development and metastasis of BC [30], while Elion and co-authors found that RIG-I agonists could induce cell death in BC [31].
Signal transducer and activator of transcription 1 (STAT1) is a transcription factor involved in multiple intracellular signaling pathways [49]. STAT1 also plays an anti-cancer role in the progression of BC and pyroptosis [50]. BC patients with higher STAT1 activation had longer relapse-free and overall survival [51], and a longer survival period without distant metastasis [52]. While another study showed that STAT1 expression in BC cells was significantly lower than that in normal breast epithelial cells, and STAT1-deficient mice developed spontaneous breast tumors [53]. Various studies indicated that cytokines could induce pyroptosis by activating STAT1 [54,55]. Wang et al. also proved that STAT1 upregulated the inflammasome related to pyroptosis [56].
RIG-I induced cell death by activating STAT1 in hepatocellular carcinoma [57], and knockdown of RIG-I suppressed STAT1 expression and promoted radiotherapy resistance in Nasopharyngeal Carcinoma [58]. It also has been proven that RIG-I agonists activated STAT1 and NF-κB pathway in BC [31]. Interestingly, the efficacy of immunotherapy for hepatocellular carcinoma could be facilitated by promoting the STAT1/GSDME pyroptotic circuitry [59]. In the present study, we discovered that G9a, RIG-I, and STAT1 form an interaction network in TNBC cells. G9a knockdown activated the expression of RIG-I and p-STAT1 to promote GSDME. We also found that inhibiting the function of RIG-I suppressed the activation of p-STAT1 and GSDME. However, when an inhibitor of RIG-I (cFP) was given to the cells that had been knocked G9a down, the expression of RIG-I, p-STAT1, and GSDME were rebounded compared to the cFP group, suggesting that the G9a knockdown could partially reverse this decrease. Similarly, suppressing RIG-I function could also reverse the increase in pyroptosis-related inflammatory factors caused by G9a knockdown. These results suggested that RIG-I/STAT1/GSDME pathway might be the potential mechanism of G9a-induced pyroptosis in TNBC cells.
These findings would provide a theoretical basis for G9a being a therapeutic target, which would be of great significance for TNBC patients with poor treatment responses. However, there are still some limitations in this study. Firstly, the effects of G9a knockdown were exclusively investigated in TNBC cell lines in vitro; further studies in other BC subtypes and in vivo are needed to verify these results. Secondly, our findings indicated that there was an interaction between G9a and RIG-I; however, how G9a epigenetically regulates RIG-I warrants additional examination. Thirdly, as a pan-caspase inhibitor, Z-VAD-FMK could inhibit both pyroptosis and apoptosis. Consequently, the role of apoptosis in the effects of pyroptosis following the inhibitor treatment requires further investigation.
5. Conclusions
In conclusion, the study indicates that G9a is highly expressed in TNBC cells. Inhibiting G9a can suppress cell proliferation, invasion, and migration, and can induce cell apoptosis and pyroptosis. The mechanism by which G9a engages in the process of pyroptosis may be related to the RIG-I/STAT1/GSDME pathway. The findings demonstrate the therapeutic potential of inhibiting the function of G9a in TNBC cells.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li Y. Zhang H. Merkher Y. Merkher Y. Chen L. Liu N. Leonov S. Chen Y. Recent advances in therapeutic strategies for triple-negative breast cancer J. Hematol. Oncol.20221512110.1186/s 13045-022-01341-036038913 PMC 9422136 · doi ↗ · pubmed ↗
- 2Harbeck N. Gnant M. Breast cancer Lancet 20173891134115010.1016/S 0140-6736(16)31891-827865536 · doi ↗ · pubmed ↗
- 3Newport-Ratiu P.A. Hussein K.A. Carter T. Panjarian S. Jonnalagadda S.C. Pandey M.K. Unveiling the intricate dance: Obesity and tnbc connection examined Life Sci.202435712308210.1016/j.lfs.2024.12308239332488 · doi ↗ · pubmed ↗
- 4Ni Y. Schmidt K.R. Werner B.A. Koenig J.K. Guldner I.H. Schnepp P.M. Tan X. Jiang L. Host M. Sun L. Death effector domain-containing protein induces vulnerability to cell cycle inhibition in triple-negative breast cancer Nat. Commun.201910286010.1038/s 41467-019-10743-731253784 PMC 6599020 · doi ↗ · pubmed ↗
- 5Waks A.G. Winer E.P. Breast cancer treatment: A review JAMA 201932128830010.1001/jama.2018.1932330667505 · doi ↗ · pubmed ↗
- 6Foulkes W.D. Smith I.E. Reis-Filho J.S. Triple-negative breast cancer N. Engl. J. Med.20103631938194810.1056/NEJ Mra 100138921067385 · doi ↗ · pubmed ↗
- 7Nagarajan K. Soundarapandian K. Thorne R.F. Li D. Li D. Activation of pyroptotic cell death pathways in cancer: An alternative therapeutic approach Transl. Oncol.20191292593110.1016/j.tranon.2019.04.01031085408 PMC 6518321 · doi ↗ · pubmed ↗
- 8Thi H.T.H. Hong S. Inflammasome as a therapeutic target for cancer prevention and treatment J. Cancer Prev.201722627310.15430/JCP.2017.22.2.6228698859 PMC 5503217 · doi ↗ · pubmed ↗