The Papilla Stage as a Critical Molecular Transition: Antp and Sex-Regulatory Network Orchestrate Cheliped Regeneration in Eriocheir sinensis
Benzhen Li, Yanan Yang, Mengqi Ni, Yourong Liu, Zhaoxia Cui

TL;DR
This study reveals how male and female Chinese mitten crabs regenerate claws differently at the molecular level, with key differences emerging early in the process.
Contribution
The study identifies the papilla stage as a critical molecular transition and highlights the role of Antp and sex-regulatory networks in cheliped regeneration.
Findings
Sexual dimorphism in cheliped regeneration is established at the molecular level as early as 4 days post autotomy.
The Hox gene Antp is a master regulator of appendage patterning and exoskeleton assembly during regeneration.
RNAi knockdown of Antp disrupts joint differentiation and confirms its regulatory role in regeneration.
Abstract
The ability to regenerate chelipeds is vital for the survival of the Chinese mitten crab (Eriocheir sinensis), yet the molecular mechanisms driving sexual dimorphism during this process remain unclear. This study combined morphological and transcriptomic analyses to investigate regenerative stages and sex-related differences. We found that while physical differences, such as larger regenerating chelipeds in males, only become prominent at 28 days post autotomy (dpa), the molecular foundation for this divergence is established as early as 4 dpa. This early divergence is driven by sex-specific endocrine networks, with males and females showing distinct gene expression patterns and hormonal activities. Furthermore, we identified the Hox gene Antennapedia (Antp) as a master regulator. RNAi-mediated knockdown of Antp resulted in impaired joint differentiation and confirmed its role in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1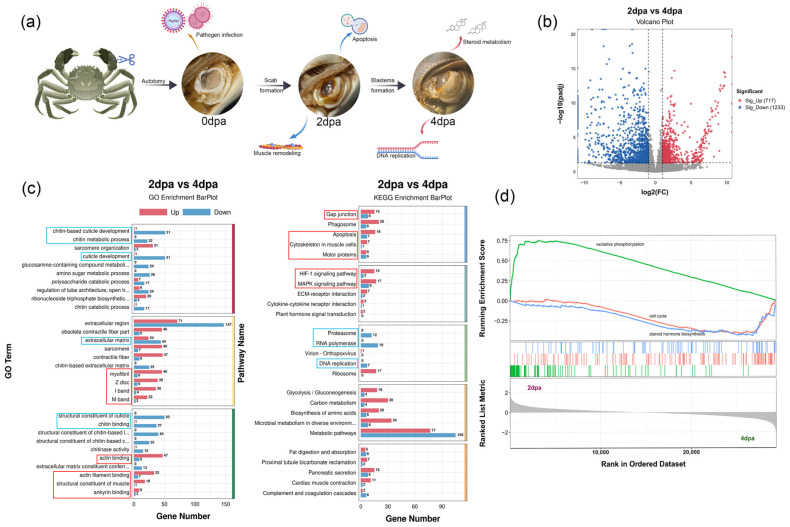 Figure 2
Figure 2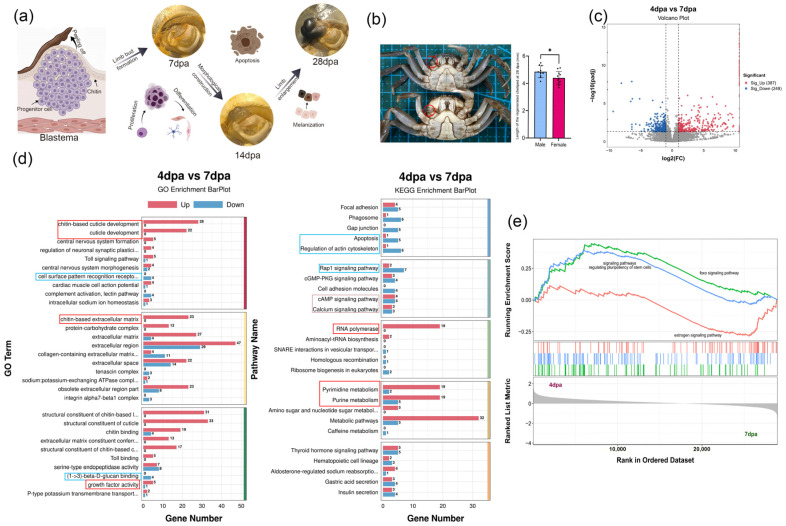 Figure 3
Figure 3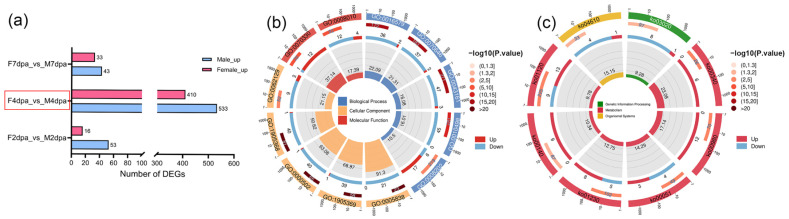 Figure 4
Figure 4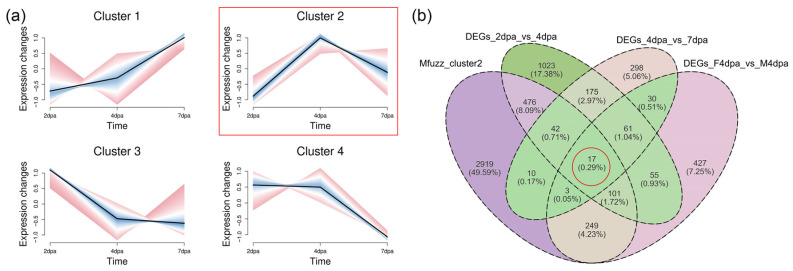 Figure 5
Figure 5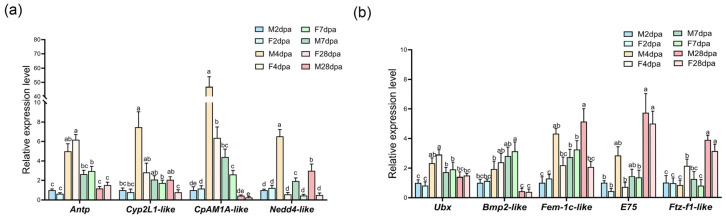 Figure 6
Figure 6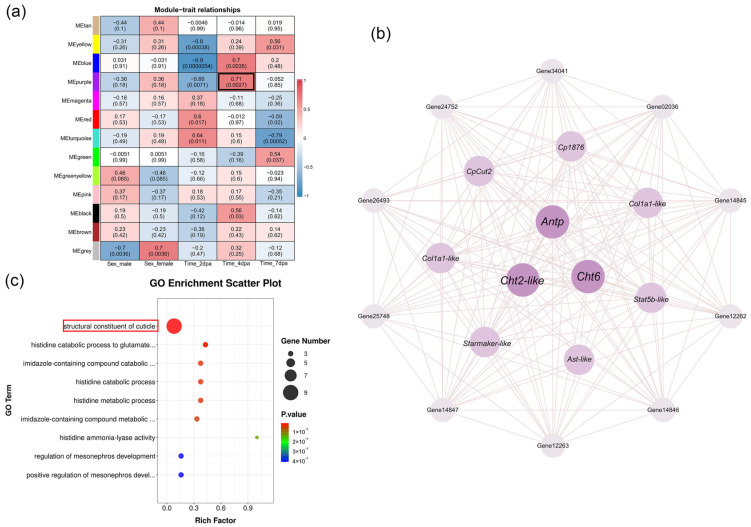 Figure 7
Figure 7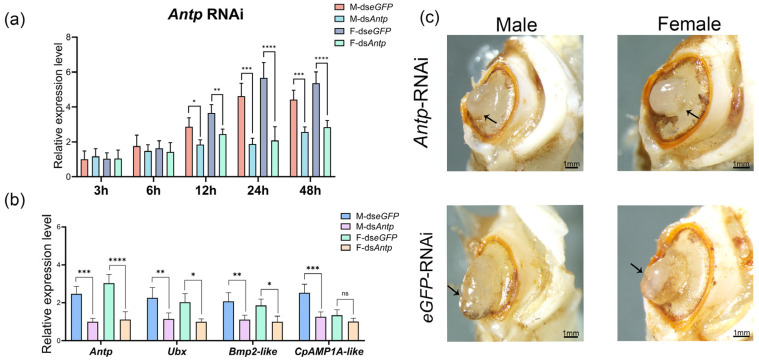 Figure 8
Figure 8- —National Natural Science Foundation of China
- —Ten Thousand Talents Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Planarian Biology and Electrostimulation · Echinoderm biology and ecology
1. Introduction
Regeneration, the spellbinding biological feat of mending damaged bodies by restoring functional tissues and complex organs, stands as one of the most enthralling puzzles in developmental biology [1,2]. The core of this process hinges on a precise cellular transition: mature cells at the wound site first undergo dedifferentiation, through which they shed their specialized phenotypic traits and undergo reprogramming to revert to a progenitor cell state [3,4]. These multipotent cells subsequently aggregate to form a proliferative blastema, and then proceed to undergo redifferentiation to faithfully recapitulate the missing tissue structure [5]. While the core stages from dedifferentiation to redifferentiation are conserved across a broad range of taxa, the molecular switches governing these transitions, particularly regarding their interaction with biological sex, have yet to be fully elucidated.
Crustaceans represent excellent models for studying the mechanisms of appendage regeneration [6]. A notable example is the Chinese mitten crab, E. sinensis, which is an economically significant aquaculture species widely distributed across East Asia [7]. In 2024, the production of Chinese mitten crab in China reached 894,395 tonnes, accounting for 75.75% of the total crustacean aquaculture output in the country [8]. In this species, the chelipeds (pincer-like appendages) are indispensable not only for foraging and defense, but also as important secondary sexual characteristics. Male individuals begin to gradually develop larger claws and denser setae during the juvenile crab stage [9,10]. Both in natural habitats and intensive aquaculture systems, E. sinensis frequently undergoes cheliped autotomy resulting from intraspecific agonistic interactions. Such impairments substantially compromise the organism’s survival, growth performance, and overall economic value [11,12].
The regenerative cycle of the cheliped encompasses multiple sequential physiological phases, among which three key stages are recognized as core drivers of the regenerative process: the scab stage, papilla stage, and limb bud stage [13,14]. Notably, the papilla stage serves as a core molecular regulatory node, marking the transition from wound healing to morphogenesis [15]. While substantial foundational research has been conducted on limb regeneration in crustaceans, and core signaling pathways including Wnt, Notch, and TGF-β have been well-documented as key regulators in this process, studies focusing on potential sexual dimorphism during the early phase from cellular reprogramming to redifferentiation remain scarce [16,17,18]. In particular, the sex-differential activation patterns and regulatory effects of these pathways in male and female individuals have not yet been thoroughly elucidated. Given that the chelipeds of male E. sinensis are significantly more robust and morphologically complex than those of females, investigating how sex-differential regulatory networks modulate the regenerative process is of great importance.
The genetic identity and spatial patterning of segments are governed by the highly conserved Hox gene family [19]. Among these, Antp acts as a master regulator of appendage identity and growth in crustacean [20]. Sex combs reduced (Scr), another critical Hox gene, is involved in the regulation of appendage patterning in Drosophila and plays a central role in the development of sex combs, a structure associated with sexual dimorphism [21]. As a member of the Hox gene family, Antp may similarly function in the development of chelipeds, which represent secondary sexual characteristics, in E. sinensis. However, the functional requirement of Antp during the early stages of cheliped regeneration in E. sinensis, as well as its potential synergy with sex differentiation-related genes, has yet to be characterized.
In this study, comparative transcriptomic profiling was employed to systematically analyze the regenerative progression across three pivotal stages in both male and female E. sinensis. Our results identify the papilla stage as the primary window for sex-biased gene expression, featuring key candidate regulators such as Antp, Cyp2L1-like, CpAMP1A-like, Fem-1c-like, Ftz-f1-like, E75, and Nedd4-like. Functional validation via RNAi further demonstrated that Antp is indispensable for the transition from the papilla stage to the growth stage; silencing of Antp not only significantly retards the regeneration rate but also downregulates the expression of Ubx, Bmp2-like and CpAMP1A-like. These findings provide novel insights into how Hox genes and sex-biased pathways coordinately govern the intricate transition from cellular dedifferentiation to redifferentiation in sexually dimorphic organs.
2. Materials and Methods
2.1. Tissue Sampling and RNA Isolation
The juvenile Chinese mitten crabs used in this experiment were obtained from Nanjing, Jiangsu Province, with an average body weight of 15 ± 3.8 g. The crabs were housed in 6 rectangular tanks (1 m × 0.6 m × 0.4 m) for a 3 day acclimation period, under conditions of a constant water temperature of 20 ± 1 °C, natural photoperiod, and dissolved oxygen content above 5 mg/L. During acclimation, the crabs were fed a mixed diet of corn, soybean meal, and manila clam (Ruditapes philippinarum) at an approximate ratio of 1:1:1, with a daily feeding allowance of roughly 5% of their body weight and daily water renewal. After acclimation, 100 males and 100 females in the intermolt stage with intact appendages were selected, anesthetized on ice, and right cheliped autotomy was induced by manual pressure. After no hemolymph exuded from the wound, the crabs were stocked at 40 individuals per tank. To simulate the real aquaculture environment, each tank was co-cultured with 20 males and 20 females. Morphological changes were recorded by photography after autotomy. Intact regenerating cheliped tissues were collected separately by sex at 2, 4, 7 and 28 dpa, with 6 individuals sampled for each sex at each stage, and the regenerating tissue from a single crab served as one biological replicate.
Total RNA was isolated from regenerating cheliped tissues using RNAiso Plus (TaKaRa, Shiga, Japan) per the manufacturer’s protocol. RNA quality and concentration were quantified with a NanoDrop™ One C Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). For samples at 2, 4 and 7 dpa, three biological replicates were prepared for both sexes and submitted to LC-Bio Technologies (Hangzhou, China) for transcriptome sequencing, designated as M/F2dpa1-3, M/F4dpa1-3, and M/F7dpa1-3.
2.2. Transcriptome Sequencing and Data Processing
Paired-end sequencing (PE150, 2 × 150 bp read length) was conducted on the Illumina NovaseqTM 6000 platform (Illumina, San Diego, CA, USA). Raw sequencing reads were first subjected to quality filtering using Trimmomatic-0.39 to generate clean data [22], followed by quality assessment of the filtered reads with FastQC-v0.11.5. The qualified clean data were then aligned to the reference genome using Hisat2-v2.2.1 [23,24]. The resulting SAM alignment files were converted to binary BAM format via Samtools-1.2.0 [25]. Finally, Stringtie-v2.2.3 was employed to quantify gene expression levels [26], with Transcripts Per Kilobase of exon model per Million mapped reads (TPM) values calculated accordingly [27].
2.3. Transcriptome Data Analysis
DEG analysis was performed on the raw count gene expression matrix using the DESeq2-3.22 package [28]. DEGs were screened with the thresholds of |log2 fold change (log2FC) > 1 and false discovery rate (FDR) < 0.05. Volcano plots visualizing DEGs were constructed using the ggplot2 package [29]. Subsequently, functional annotation of DEGs was conducted via sequence alignment against the non-redundant database (https://ftp.ncbi.nlm.nih.gov/blast/db/FASTA/nr.gz, accessed on 27 April 2025) using Diamond-2.1.8 and Blast-2.14.0 [30,31]. For functional classification and pathway enrichment, DEGs were annotated based on the GO (http://geneontology.org, accessed on 12 May 2025) and Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.kegg.jp, accessed on 12 May 2025). GO, KEGG, and Gene Set Enrichment Analysis (GSEA) were performed using the OmicStudio tools (https://www.omicstudio.cn/tool), with all expressed genes serving as the background gene set. For GSEA, the entire gene expression matrix was used to identify coordinated biological changes and significant enrichment patterns. In addition, some figures were generated using BioGDB (https://biogdp.com) [32].
2.4. Cluster Analysis of Gene Expression Patterns and Validation by qRT-PCR
Cluster analysis and visualization of gene expression patterns across transcriptome data from different time points were conducted via the Mfuzz 2.54.0 package [33]. Venn diagrams were constructed to visualize the overlap of DEGs among different time points using the VennDiagram v1.7.4 package [34]. Primers were designed using SnapGene 6.02 (https://www.snapgene.com) (Table S1). β-actin served as the reference gene for normalization. Following total RNA extraction, reverse transcription was performed, and quantitative reverse transcription polymerase chain reaction (qRT-PCR) was carried out using the PerfectStart^®^ Green qPCR SuperMix (TransGen Biotech, Beijing, China). Each experiment group included 4 biological replicates and 3 technical replicates to ensure reliability. Relative gene expression levels were computed using the 2^−ΔΔCt^ method [35]. All data were presented as the mean ± standard deviation (SD). Normality of the data distribution was assessed using the Shapiro–Wilk test via GraphPad Prism 10.1.12. Homogeneity of variances was examined by Levene’s test with SPSS 17.0 software [36]. According to the characteristics of the gene-related data, subsequent statistical analysis was performed using mixed-model analysis of variance (ANOVA) with GraphPad Prism. Bar graphs were constructed using GraphPad Prism 10.1.12.
2.5. Weighted Gene Co-Expression Network Analysis
WGCNA was constructed using the WGCNA Shiny v0.0.6.230118 embedded in TBtools-II v2.360 [37,38]. Prior to network establishment, TPM values were subjected to logarithmic transformation for data normalization. To guarantee high-quality data input, genes with expression levels below 1 in over 50% of the samples were filtered out. The key parameters for network construction were set as follows: soft threshold was fixed at 10, module tree height was set to 0.25, and the minimum number of genes per module was defined as 30. The cytohubba v0.1 plugin was used to screen hub genes of the modules and construct the corresponding network graphs (https://apps.cytoscape.org/apps/cytohubba, accessed on 21 May 2025).
2.6. RNA Interference
Primers for dsRNA synthesis of Antp and eGFP were designed (Table 1). dsRNA was synthesized via in vitro transcription using a dsRNA Synthesis Kit (Sangon Biotech, Shanghai, China). The dsRNA was dissolved in sterile, enzyme-free PBS, with a small amount of phenol red added as an indicator. To avoid degradation, the mixture of dsRNA and phenol red was freshly prepared immediately before each injection.
After cheliped autotomy, crabs were monitored for scab formation to ensure all individuals entered the scab stage; those that failed to form scabs were excluded before injection. At 3 dpa, dsRNA was injected into the coxopodite of the regenerating cheliped at a dose of 3 μg per gram of crab body weight.
First, RNAi efficiency was verified. Then, according to the effect of injection frequency on survival rate, an injection interval of 72 h was determined, and a total of three rounds of injections were performed. The dsRNA-Antp group included 30 males and 30 females, while the dsRNA-eGFP control group included 25 males and 25 females.
3. Results
3.1. Morphological Observation and Sequencing Data of Cheliped Regeneration of E. sinensis
The majority of E. sinensis individuals regenerated a fully functional cheliped after a single molting event post autotomy. Within hours post amputation, the wound site appeared transparent, accompanied by a minor exudation of hemolymph. Subsequently, black punctate structures emerged at the wound surface (Figure 1a). By 2 dpa, a black scab had completely covered the amputation site (Figure 1b). Around 4 dpa, this scab began to detach, revealing a transparent, papillary structure enclosed by a thin, translucent membrane (Figure 1c). By 7 dpa, a prominent regenerative limb bud became visible (Figure 1d). By approximately 14 dpa, a structurally intact but transparent new cheliped had formed (Figure 1e). During the following two weeks, the segments of the nascent cheliped became fully encased in a black membrane and exhibited significant swelling (Figure 1f).
Transcriptome sequencing was performed on the regenerating chelipeds of E. sinensis at 2, 4, and 7 dpa based on the morphological observation. Following quality filtering, a total of 129.38 GB of high-quality data were generated for subsequent bioinformatic analysis. The average proportion of Q30 bases (corresponding to a sequencing error rate of <0.1%) reached 95.76%, with an average GC content of 49.63% and an average mapping rate of 78.62% (Table S2).
3.2. Molecular Transition from Wound Healing to Blastema Formation During Early Limb Regeneration in E. sinensis
Following cheliped autotomy, E. sinensis underwent a sequence of wound healing and blastema formation, marking a pivotal transition from wound repair to the initiation of regeneration. The 2 dpa stage represented the scab phase of healing, whereas by 4 dpa, the formation of a papilla was observed, which set the stage for subsequent morphological patterning (Figure 2a). Transcriptomic profiling identified 1950 DEGs at 2 dpa compared to 4 dpa, comprising 717 up-regulated and 1233 down-regulated genes (Figure 2b).
At 2 dpa, the molecular signature was defined by tissue remodeling and acute stress responses. DEGs were significantly associated with myofibril assembly and cytoskeletal dynamics involving actin and ankyrin binding activities (Figure 2c). The activation of HIF-1 and MAPK pathways, coupled with high oxidative phosphorylation in GSEA, suggested intense cellular homeostasis and oxidative stress during initial wound closure (Figure 2c,d).
By 4 dpa, the transcriptional landscape shifted toward structural reconstruction and rapid cell proliferation. This stage was characterized by enriched chitin metabolic processes and cuticle development, signaling the onset of new tissue formation (Figure 2c). The activation of DNA replication and cell cycle-related pathways reflected the high proliferative capacity of nascent blastema cells. Notably, the upregulated steroid metabolism observed in GSEA indicated that the initiation of regeneration might be regulated by steroid hormones (Figure 2d).
3.3. Molecular Transition from Blastema Development to Morphogenesis in the Regenerating Cheliped of E. sinensis
Following the peeling off of the scab, the regenerative papilla initiates formal regeneration by proliferating and differentiating into a limb bud with indistinct segments. As regeneration progressed, coordinated morphogenesis meticulously sculpted the definitive cheliped structure, followed by segmental expansion and cuticular melanization. Notably, the early development of the papilla was regulated by steroid hormones, which potentially drove the sexual dimorphism observed in regenerated chelipeds by 28 dpa (Figure 3a,b). Male and female transcript profiles were combined for this comparison to exclude sex effects and highlight changes across regeneration stages. Comparative transcriptomic profiling revealed 636 genes differentially expressed between 4 dpa and 7 dpa, with 387 genes up-regulated and 249 genes down-regulated in the 4 dpa group (Figure 3c).
At 4 dpa, the molecular signature focused on structural biosynthesis and robust metabolism. GO analysis showed that DEGs were primarily enriched in chitin-based cuticle development and growth factor activity (Figure 3d). KEGG analysis revealed a high demand for material synthesis, specifically in pyrimidine/purine metabolism and RNA polymerase pathways. Notably, GSEA identified activation of signaling pathways regulating stem cell pluripotency and the foxo signaling pathway (Figure 3e), consistent with the blastema’s role as a convergence of dedifferentiated progenitor cells. By 7 dpa, the landscape shifted toward immune recognition and cytoskeletal remodeling. GO terms were significantly associated with (1 → 3)-β-D-glucan binding and cell surface pattern recognition receptor signaling. At the pathway level, 7 dpa exhibited enrichment in apoptosis and actin cytoskeleton regulation, suggesting active morphological sculpting (Figure 3d). While cAMP and Calcium signaling were enriched at both stages, GSEA specifically highlighted the estradiol pathway at 7 dpa (Figure 3e). This indicates that, following early steroid-regulated initiation, subsequent re-differentiation is further governed by sex hormone signaling.
3.4. Sexual Differences in the Transcriptomic Landscape During the Papilla Stage
To investigate the sexual differences in E. sinensis during early limb regeneration, we analyzed the DEGs between sexes across three stages. The results showed a male-biased expression pattern, with more up-regulated genes in males than in females at all stages; the number of DEGs peaked at 4 dpa (Figure 4a). Functional enrichment analysis revealed that male-upregulated genes were significantly enriched in protein modification processes, such as protein deubiquitination (GO:0016579) and protein modification by small protein removal (GO:0070646). In contrast, female-upregulated genes were enriched in aromatase activity (GO:0070330). Regarding cuticle-related terms, males showed enrichment in the structural constituent of chitin-based larval cuticle (GO:0008010) (Figure 4b). Furthermore, KEGG enrichment analysis revealed distinct enrichment patterns between sexes: the complement and coagulation cascades (ko04610) and fructose and mannose metabolism (ko00051) were enriched in males, while steroid hormone biosynthesis (ko00140) and metabolism of xenobiotics by cytochrome P450 (ko00980) were enriched in females (Figure 4c).
3.5. Antp and Sex-Differential Key DEGs Revealed by Cluster-Based Screening
All genes identified by transcriptome sequencing that met the screening criteria were categorized into 4 clusters based on their expression patterns (Figure 5a). Given that 4 dpa was a critical transitional phase during the early stage of cheliped regeneration and also a pivotal period for sexual differentiation, we focused on the genes with relatively high expression levels at 4 dpa. Among these clusters, the genes in cluster 2 exhibited the highest expression levels at 4 dpa in terms of their expression patterns, and this module contained 3817 genes. By taking the intersection of DEGs across different regeneration stages, DEGs between sexes at 4 dpa, and genes within Cluster 2, we identified 17 key genes (Figure 5b, Table 2). Among these were Antp, Cyp2L1-like, CpAMP1A-like and Nedd4-like.
3.6. Validation of the Key DEGs and Their Expression Dynamics During Regeneration
To verify the reliability of the transcriptomic data, several representative genes were selected from the intersection analysis for validation, including Antp, CpAMP1A-like, Cyp2L1-like, and Nedd4-like. All these genes reached their peak expression levels at 4 dpa and exhibited significant sex-based differences. Notably, Nedd4-like expression was consistently higher in males than in females throughout the cheliped redifferentiation stage (Figure 6a). Furthermore, we validated the expression levels of sex-differential DEGs identified at 4 dpa, such as Fem-1c-like, Ftz-f1-like, and E75, along with genes showing significant differential expression across different regeneration stages, including Ubx and Bmp2-like (Figure 6b). The qRT-PCR results were highly consistent with the expression trends observed in the RNA-Seq data, confirming the accuracy of our sequencing results. Given the distinct phenotypic differences observed between male and female regenerating claws at 28 dpa, we also performed qPCR analysis on these genes at this later stage to explore their potential roles.
3.7. The Key Module and Hub Genes at the Regenerative Papilla Stage
WGCNA was performed on transcriptome-derived genes meeting the preset criteria, yielding 12 valid modules. Among these modules, the purple module showed the strongest correlation with 4 dpa (r = 0.71, p < 0.05) and comprised a total of 101 genes (Figure 7a). Gene ranking using cytoHubba identified Antp as a hub gene with high connectivity within this module. Furthermore, other highly connected genes, such as Cht6, Cht2-like, and Cp1876, were predominantly associated with crustacean cuticle development (Figure 7b). GO enrichment analysis of all genes in the purple module revealed that they were primarily enriched in the functional category of structural constituent of cuticle (Figure 7c).
3.8. Effects of Antp RNAi on Cheliped Regeneration in E. sinensis
Based on mfuzz analyses and WGCNA, Antp was identified as a key gene characterized by both differential expression across regeneration stages and significant sex-based differences. To investigate the functional significance of Antp, RNAi was performed via dsRNA injections at the scab stage, achieving peak silencing efficiency at 24 h post-injection. Compared to the control group (dsRNA-eGFP), Antp expression levels in males and females were significantly reduced to 40.56% and 36.40%, respectively (Figure 8a). The results demonstrated that the silencing of Antp led to a significant down-regulation of Ubx and Bmp2-like in both males and females. Notably, the regulatory effect on CpAMP1A-like showed a sex-based difference, with its expression being significantly suppressed only in males. These findings suggest that Antp exerted sex-based regulatory control over downstream targets. Specifically, Antp appeared to play a more predominant or direct role in the regenerative regulatory network of males compared to females, particularly concerning the modulation of cuticle-related genes such as CpAMP1A-like (Figure 8b). Following three rounds of dsRNA-Antp treatment, the segmentation boundary between the propodus and carpus of regenerating chelipeds was obscure in several individuals, in sharp contrast to the well-defined segmentation observed in the dsRNA-eGFP control group (Figure 8c). Specifically, only 16.7% of males and 13.3% of females in the dsRNA-Antp group exhibited distinct segment differentiation, whereas 46.2% of males and 44% of females showed clear segmentation in the control group. These results indicate a significant inhibitory effect of Antp knockdown on cheliped segment differentiation (Figure S1).
4. Discussion
Cheliped regeneration in E. sinensis progresses from wound healing to structure-specific morphogenesis. The wound healing stage (2 dpa) is driven by a defensive-reconstructive network, where MAPK activation likely serves as a dual regulator of stress responses and tissue repair [39]. Concurrently, the upregulation of muscle remodeling genes supports blastema formation, consistent with observations in Cherax destructor and Exopalaemon carinicauda [14,40]. As regeneration advances to the limb bud stage (7 dpa), the role of apoptosis shifts from cellular clearance to active morphogenetic sculpting, a mechanism essential for segmentation that parallels development in Drosophila [41,42]. Ultimately, the definitive sexual dimorphism of the regenerated chelipeds become morphologically distinct by 28 dpa.
The papilla stage (4 dpa) represents a pivotal turning point in the transcriptional landscape, marking the transition from an undifferentiated state to rapid proliferation and epidermal specification. This stage is characterized by a strategic redirection of metabolic energy toward chitin biosynthesis and epidermal morphogenesis. Given that chitin and cuticle proteins are indispensable for morphogenesis in insects [43], a similar conserved mechanism likely operates in E. sinensis as a decapod crustacean. The concomitant activation of DNA replication and RNA polymerase pathways signify an exponential surge in cellular proliferation, a fundamental prerequisite for blastema establishment that mirrors the conserved proliferative strategies observed in Ambystoma mexicanum [44]. Consequently, we identify 4 dpa as the peak of proliferative transcriptional activity. The regenerative program diverges from a shared wound-healing response to a sex-differential trajectory during this critical window. The distinct transcriptional profiles suggest that the papilla stage serves as the mechanistic foundation where physiological resources are allocated differently between sexes to support dimorphic limb regeneration.
Integrated analysis suggests that Antp may act as a central hub gene orchestrating limb patterning. Its temporal expression is low at 2 dpa and peaks at 4 dpa. This pattern implies that the subsequent upregulation potentially drives structure-specific differentiation. Studies in mice (Mus musculus) have indicated that Hox gene expression may serve as a mechanism that sustains periosteal stem/progenitor cells in a more primitive tripotent state [45]. RNAi-mediated silencing of Antp leads to arrested segmental differentiation and the significant downregulation of Ubx, Bmp2-like, and CpAMP1A-like. These results indicate that Antp acts synergistically with Ubx to define segmentation [46] and may regulate epidermal development via the TGF-β signaling pathway to further modulate limb size [47]. Furthermore, the observation that Antp regulates various epidermal-related genes mirrors conserved mechanisms in Drosophila and Bombyx mori. Antp directly binds to cis-regulatory elements of epidermal protein genes to modulate organ development, and Antp knockdown-induced downregulation of cuticle proteins in E. sinensis likely follows this conserved mode of action [48]. Collectively, our findings position Antp as a master regulator that integrates structural patterning with the expression of epidermal genes.
The pronounced sexual dimorphism in cheliped regeneration is driven by sex-specific regulatory networks recruited from the papilla stage. Male regeneration prioritizes the chitinous exoskeleton pathway (CpAMP1A-like) and actively recruits male differentiation factors, specifically Fem-1c-like and Nedd4-like. The transcriptional profiles align with their established biological functions: Fem-1c plays a crucial role in male sexual differentiation in E. sinensis, whereas Nedd4 deficiency leads to male-to-female sex reversal in mice [49,50]. Furthermore, the male-biased fructose and mannose metabolism may be associated with the development of a rigid exoskeleton, a pattern that has also been observed in transcriptomic studies of the silkworm Bombyx mori [51]. In contrast, the female program emphasizes systemic signaling, characterized by elevated aromatase activity and Ftz-f1-like expression. The enrichment of aromatase and Ftz-f1-like in females contrasts sharply with the male-biased expression of Cyp2L1-like [52,53]. This distinct pattern suggests that the regenerating tissue actively mediates systemic hormonal signals, mirroring mechanisms observed in Danio rerio [54]. Notably, cheliped regeneration in E. sinensis is inextricably linked to molting behavior. Given this close association, the nuclear receptors E75 and Ftz-f1-like likely integrate ecdysteroid and sex-steroid signaling [55,56]. Consequently, we propose a hierarchical model wherein sex-biased hormonal signals modulate the expression of Antp and other development-related genes, which subsequently influence downstream phenotype-related genes to ensure sexually dimorphic morphogenesis.
5. Conclusions
This study provides a comprehensive temporal and sex-differential characterization of early cheliped regeneration in E. sinensis. Our findings identify the papilla stage (4 dpa) as a pivotal physiological and transcriptional crossroad that underpins the molecular foundation of sexual dimorphism. This divergence is governed by sex-differential networks: males exhibit preferential expression of Fem-1c-like, Cyp2L1-like, and Nedd4-like, while females are characterized by elevated aromatase activity. Within this regulatory framework, the Hox gene Antp serves a critical role in orchestrating appendage patterning and exoskeleton assembly with highly co-expressed cuticle-related genes. Ultimately, we speculate a hierarchical regulatory model in which systemic hormonal signals may integrate Antp and other sex-biased regulators to promote the precisely structured regeneration of chelipeds. These findings provide novel insights into the interplay between developmental plasticity and sex-differential development in decapod crustaceans.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Poss K.D. Advances in understanding tissue regenerative capacity and mechanisms in animals Nat. Rev. Genet.20101171072210.1038/nrg 287920838411 PMC 3069856 · doi ↗ · pubmed ↗
- 2Tanaka E.M. Reddien P.W. The cellular basis for animal regeneration Dev. Cell 20112117218510.1016/j.devcel.2011.06.01621763617 PMC 3139400 · doi ↗ · pubmed ↗
- 3Jopling C. Boue S. Izpisua Belmonte J.C. Dedifferentiation, transdifferentiation and reprogramming: Three routes to regeneration Nat. Rev. Mol. Cell Biol.201112798910.1038/nrm 304321252997 · doi ↗ · pubmed ↗
- 4Galliot B. Ghila L. Cell plasticity in homeostasis and regeneration Mol. Reprod. Dev.20107783785510.1002/mrd.2120620602493 · doi ↗ · pubmed ↗
- 5Mokalled M.H. Poss K.D. A regeneration toolkit Dev. Cell 20184726728010.1016/j.devcel.2018.10.01530399333 PMC 6373444 · doi ↗ · pubmed ↗
- 6Almazán A. ÇevrimÇ. Musser J.M. Averof M. Paris M. Crustacean leg regeneration restores complex microanatomy and cell diversity Sci. Adv.20228 eabn 982310.1126/sciadv.abn 982336001670 PMC 9401613 · doi ↗ · pubmed ↗
- 7Che S. Gao J. Zhu H. Du J. Cao L. Zheng Y. Xu G. Liu B. Integrated application of transcriptomics and metabolomics provides insights into the different body-size growth in Chinese mitten crab (Eriocheir sinensis)Int. J. Mol. Sci.202526461710.3390/ijms 2610461740429760 PMC 12110897 · doi ↗ · pubmed ↗
- 8The Bureau of Fisheries and Fishery Administration, Ministry of Agriculture National Fisheries Technology Extension Center Chinese Society of Fisheries China Fisheries Statistical Yearbook 20251 st ed.China Agriculture Press Beijing, China 20252734