Frog Skin Peptides: Nature’s Dual-Action Weapons Against Infection and Cancer
Eleonora Grisard, Carlo Vetrano, Ali Benour, Eeva Tortellini, Dania Al Ismail, Giacomo Cappella, Bruno Casciaro, Maria Luisa Mangoni, Milena Mechkarska

TL;DR
Frog skin peptides show promise as dual-action treatments for both infections and cancer due to their unique properties and mechanisms.
Contribution
This review identifies and emphasizes frog skin peptides with dual antimicrobial and anticancer properties and their translational potential.
Findings
Frog skin peptides like dermaseptins, temporins, and brevinins disrupt microbial membranes and induce cancer cell death.
Key features like net positive charge and α-helical conformation contribute to their dual functionality.
Peptide engineering and delivery advances have improved their stability and therapeutic potential.
Abstract
The rise of antimicrobial resistance and the global burden of cancer demand innovative therapeutic strategies. Frog skin secretions offer a rich source of bioactive peptides, some of which exhibit remarkable dual functionality—potent antimicrobial activity coupled with selective anticancer effects. This review highlights frog skin-derived peptides that bridge the gap between antimicrobial and anticancer therapeutics, emphasizing their structural diversity, mechanisms of action, and translational potential. A comprehensive literature search was conducted to identify peptides isolated from diverse anuran species, with emphasis on studies reporting structural features, activity against Gram-positive and Gram-negative bacteria, including multidrug resistant clinical isolates, anticancer effects, and underlying molecular mechanisms of cytotoxicity. Peptides such as dermaseptins, temporins,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
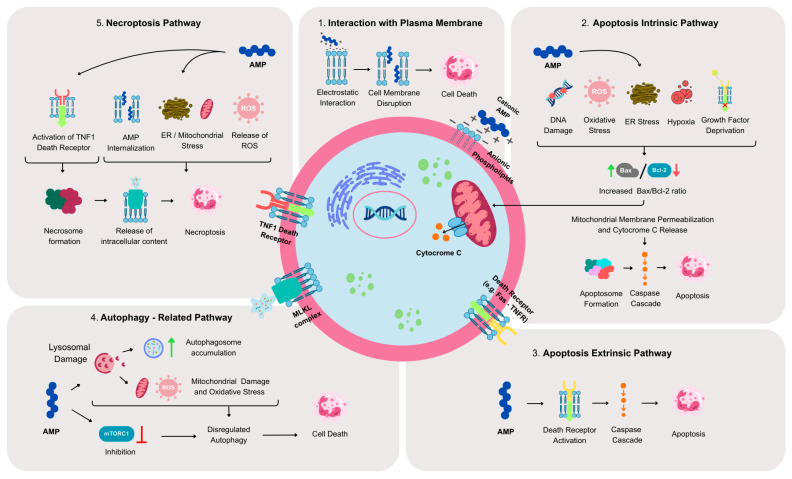 Figure 1
Figure 1- —Sapienza University
- —Italian Ministry of University and Research
- —European Union
- —PNRR PhD scholarship
- —UWI St. Augustine Campus
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Peptides and Activities · Microbial Natural Products and Biosynthesis · Biochemical and Structural Characterization
1. Introduction
The COVID-19 pandemic has made the world acutely aware of the impact of global health emergencies [1]. Yet, another ongoing crisis—often described as a “silent pandemic”—is projected to cause 10 million deaths per year by 2050: antimicrobial resistance (AMR) [2]. The extensive use of antibiotics in clinical settings and in agriculture has accelerated the emergence and environmental spread of multidrug-resistant (MDR) bacteria. In parallel, the discovery of new antibiotics has markedly declined, creating a critical threat to global health [3,4,5]. At the same time, cancer-related mortality is also expected to rise dramatically. By 2050, cancer is projected to cause 18.5 million deaths, representing an 89.7% increase compared with the 2022 estimate of 9.7 million deaths [6]. Cancer is not a single disease but rather a heterogeneous group of diseases comprising more than 250 clinico-pathological types and thousands of described neoplastic variants [7,8]. In this increasingly complex landscape, the links between microbial infections—caused by viruses, bacteria, and fungi—and tumorigenesis have become more evident. Major infectious contributors to cancer development include Helicobacter pylori (5%), human papillomaviruses (HPV) (5%), hepatitis B and C viruses (HBV, HCV) (5%), Epstein–Barr virus (EBV) (1%), and HIV together with human herpesvirus (HSV) (1%). Moreover, chemotherapy can profoundly disrupt the host microbiome, increasing susceptibility to infections and potentially promoting cancer development [9]. Persistent microbial infections that induce chronic inflammation are now recognized as key drivers of carcinogenesis in affected tissues [10]. Given the dangerous interplay between microbial infection and cancer development, discovering compounds with dual antibacterial and anticancer properties is a highly valuable strategy—potentially reducing toxicity, limiting the need for multiple drugs, and mitigating resistance.
Antimicrobial peptides (AMPs) represent a promising class of such therapeutics. AMPs are gene-encoded bioactive peptides produced by virtually all living organisms. Synthesized as pre-propeptides, they are processed into their active form through proteolytic cleavage [11]. They are typically cationic, short (10–50 amino acids), and amphipathic, and they exert antimicrobial activity primarily by interacting with and disrupting microbial membranes [12]. Nonetheless, AMPs also act through a variety of alternative mechanisms: for instance, proline-rich AMPs inhibit protein synthesis by binding to ribosomes [13]; others interfere with membrane protein functions, such as the LPS transport system [14]; and compounds like darobactin target components of the β-barrel assembly machinery [15]. This mechanistic diversity reduces the likelihood of resistance development [16]. Beyond their antimicrobial role, AMPs often possess a wide array of biological functions; therefore, they are referred to as “host defense peptides” (HDPs) [17]. They can also act as drug-delivery vectors, signaling molecules, contraceptive agents, immunomodulators, mitogenic factors, and, importantly, antitumor agents [18].
2. Amphibian Skin Peptides: Blueprint for Development of Antimicrobial and Anticancer Therapeutics
Frog skin secretions have captivated researchers for decades as a natural pharmacy of bioactive molecules. Among their most fascinating components are the AMPs, small, cationic, and amphipathic molecules that serve as the first line of defense in amphibian immunity. The story began in the 1980s, when Michael Zasloff’s discovery of magainins from the African clawed frog Xenopus laevis unveiled a new class of natural antibiotics capable of killing a wide range of pathogens [19]. Since then, hundreds of structurally diverse AMPs have been isolated from frog species across the families Ranidae, Hylidae, and Phyllomedusidae [20,21]. Their remarkable structural plasticity—ranging from α-helices to β-sheets and looped forms—has made them invaluable templates for designing novel antimicrobial agents at a time when infections caused by MDR bacteria pose a mounting global threat [22].
In recent years, a new role has emerged for these peptides: several frog skin AMPs also exhibit potent anticancer activity. Peptides such as dermaseptins, temporins, and brevinins can selectively destroy cancer cells while sparing healthy ones, exploiting differences in membrane charge and composition [23,24]. For instance, dermaseptin-B2 (from Phyllomedusa bicolor) not only shows broad antimicrobial effects but also inhibits the growth of human prostate and breast cancer cells by inducing apoptosis and disrupting cellular membranes [24,25]. Likewise, temporin-1CEa (from Rana chensinensis) and brevinin-2R (from Pelophylax ridibundus) have demonstrated anticancer activity through mitochondrial damage and caspase activation [26]. The physicochemical features that empower these peptides—cationicity, amphipathicity, and α-helical conformation—enable them to interact with negatively charged surfaces common to both microbial and tumor cells. This dual functionality places frog-derived AMPs at the crossroads of antimicrobial and anticancer drug discovery, positioning them as natural blueprints for multifunctional therapeutics in an era of widespread hospital and community-associated infections, antimicrobial resistance and oncologic challenges.
A novel and rapidly expanding approach has emerged that leverages artificial intelligence (AI) and machine learning (ML) models to identify recurring sequence–structure–function relationships, thereby accelerating the discovery of peptides as both antimicrobial and anticancer therapeutic leads. Key physicochemical features, such as those discussed earlier, which underpin selective membrane disruption in microbial and cancer cell membranes, can be effectively predicted using ML models trained on large datasets of AMPs and anticancer peptides [27,28]. These AI-driven strategies enable rapid in silico screening and rational peptide design, allowing prioritisation of candidates with high predicted efficacy and reduced host toxicity prior to costly synthesis and biological evaluation. Most published studies report AI-guided peptide discovery workflows that employ computational tools typically focused on individual prediction tasks—either antimicrobial or anticancer activity. While such studies establish a strong foundation for dual-function peptide discovery, combined experimental validation of peptides predicted to possess both activities remains an emerging area. To date, most validation studies are limited to in vitro evaluation of antimicrobial activity against multiple bacterial species and anticancer effects in tumor-derived cell lines [29], whereas direct in vivo validation of AI-designed peptides with dual antimicrobial and anticancer activity in animal models remains rare. Consequently, ML/AI-based prediction and design strategies were not a central focus of this article.
This review summarizes the experimentally derived data available to date on selected AMPs with dual activity, including their names and primary structures, the frog species from which they were isolated [30], and their anticancer and antibacterial activities as per the original publications (Table 1). Anticancer activity is reported as lethal concentration 50% (LC_50_) or inhibitory concentration 50% (IC_50_) values for cancer cells, while antibacterial activity is expressed as minimal inhibitory concentrations (MICs) or lethal concentrations (LCs) for bacterial growth. We then focus in detail on the principal physicochemical and molecular mechanisms underlying their anticancer activity (Section 2.1), followed by an analysis of the functional consequences of AMP treatment on tumor cell behavior (Section 2.2), conserved mode of AMP action across species (Section 2.3), and translational implications and safety concerns (Section 2.4).
2.1. Mechanisms of Anticancer Action
Several frog skin-derived AMPs have been evaluated for both antimicrobial and anticancer activities. Their anticancer potential has been investigated predominantly in vitro using a wide range of human tumor-derived cell lines and, to a lesser extent, in vivo in murine cancer models. The principal molecular mechanisms underlying the anticancer activity of AMPs are summarized (Figure 1) and are discussed in detail in the following sections.
2.1.1. Cytolysis Mediated by Physical Interaction with the Tumor Cell Plasma Membrane
Most AMPs are enriched in positively charged amino acids—arginine, lysine, and histidine—which confer a strong cationic character. This property underlies their antimicrobial activity by enabling electrostatic interactions with negatively charged bacterial membranes [155]. In contrast, plasma membranes of normal eukaryotic cells are dominated by neutral phospholipids such as phosphatidylcholine. Many tumor cells, however, undergo compositional remodeling that results in an increased abundance of negatively charged lipids—including phosphatidylserine, phosphatidic acid, and altered cholesterol distribution—resulting in a predominantly more anionic membrane surface [156]. As a consequence, cationic AMPs from both vertebrate and invertebrate organisms often display selective cytolytic activity toward cancer cells via mechanisms analogous to those used against bacterial membranes [157].
Two examples of such peptides, bombinin H4 and temporin A, were recently tested on non-small cell lung cancer (NSCLC) cell lines and normal bronchial epithelial cells [43]. Both peptides exhibited selective cytotoxicity toward NSCLC cells, though to different degrees. Lipidomic profiling revealed significant differences in membrane phospholipid composition between normal and tumor cells, with NSCLC cells displaying higher levels of negatively charged phospholipids. These findings support the idea that the selective anticancer action of cationic AMPs is linked, at least in part, to preferential binding to and disruption of anionic tumor membranes. Bombinin H4 also inhibited the growth of S. epidermidis strains 1457 and 5179-R1, highlighting its dual antimicrobial and anticancer functionality [43].
However, membrane-mediated tumor killing is not restricted only to cationic AMPs. The anionic amphibian peptide maximin-H5, active against Gram-positive S. aureus [122], selectively killed human glioblastoma cells while sparing normal glial cells. The potent cytotoxicity is attributed to its ability to adopt an α-helical conformation in proximity to the anionic glioblastoma membrane, leading to non-membranolytic, yet selective, tumor cell death [120].
Many anticancer AMPs from both vertebrate and invertebrate organisms exhibit membranolytic activity [157]. The most common assay used to evaluate plasma membrane integrity is the lactate dehydrogenase (LDH) release test, which detects extracellular LDH as an indicator of membrane damage [158]. For frog-skinderived anticancer AMPs, membrane lysis is a frequently reported consequence of peptide treatment.
In studies where AMPs induce tumor cell death through defined intracellular cell-death pathways (see sections below), LDH release is often only one among several functional assays [88,134,150]. Due to the inherent limitations of in vitro experiments, which cannot easily resolve the chronological order of events following peptide exposure, it remains unclear whether membrane disruption is a downstream consequence of programmed cell death or the initiating event that triggers such pathways.
In other studies, LDH release represents the sole measure of membrane integrity, accompanied only by general cytotoxicity assays [87,90,91,129,135,143,159]. This makes it even more challenging to determine whether membrane permeabilization arises as part of a defined cellular/molecular death program or as a primary effect of a direct AMP–membrane interaction.
Notably, all of these peptides also exhibit broad-spectrum antimicrobial activity, strengthening the hypothesis that membrane disruption may be a conserved mechanism across microbial and cancer targets. The antimicrobial spectra are reported mostly for strains of the ESKAPE+ pathogens (highly virulent MDR bacteria responsible for the majority of hospital-acquired infections worldwide, including: Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.), as well as for MRSA and various bacterial MDR clinical isolates [87,90,91,110,129,135,140,159].
2.1.2. Activation of Cell-Death Pathways
Most amphibian AMPs with anticancer properties described in this review exert their effects through the direct or indirect activation of specific cell-death programs. The predominant pathways engaged by these peptides include: (i) the intrinsic apoptotic pathway, (ii) the extrinsic (death receptor-mediated) apoptotic pathway, (iii) autophagy-related cell-death mechanisms, and (iv) the necroptosis pathway (see also Figure 1).
(i)Apoptosis Intrinsic Pathway (Mitochondrial Pathway)
The intrinsic, or mitochondrial, apoptotic pathway is activated by internal cellular stress signals, including DNA damage, hypoxia, ER stress, oxidative stress, or deprivation of growth factors. These stimuli are sensed primarily by Bcl-2 family proteins, which regulate mitochondrial membrane integrity [160]. Activation of the pro-apoptotic member Bax promotes mitochondrial outer membrane permeabilization, cytochrome-c release, and initiation of the apoptotic cascade; thus, an increased Bax/Bcl-2 ratio is widely used as an indicator of mitochondrial apoptosis activation [161].
Temporin-SHf, a peptide with strong potency against E. coli, was shown to selectively induce mitochondrial apoptosis in lung cancer cells [150]. Initial 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assays revealed selective toxicity toward various tumor cell lines (lung, breast, prostate, liver) but not toward normal endothelial cells. A549 lung cancer cells, identified as the most sensitive, displayed a dose-dependent increase in p53 and Bax expression following treatment, leading to elevation of the Bax/Bcl-2 ratio. The mitochondrial apoptotic program was further supported by increased levels of initiator and effector molecules—including total and cleaved caspase-9, -8, -3, and PARP—as well as chromatin condensation and fragmentation. Temporin-SHf also caused membranolytic effects (measured by LDH release), reduced tumor cell growth in soft agar, and impaired tumor-cell migration in a pseudo-wound-healing assay, highlighting its broad functional impact on cancer cell behavior [150].
Hymenochirin-1B, active against several Gram-positive and Gram-negative bacteria [105,106,162], similarly exhibited selective toxicity toward hepatoma and lung carcinoma cells by MTT assay, with lung cancer cells again being the most sensitive [105]. Unlike temporin-SHf, hymenochirin-1B did not induce membrane lysis by LDH assay. Instead, it triggered G0/G1 cell-cycle arrest and apoptosis (Annexin V/PI staining) through direct interaction with mitochondria. This was accompanied by mitochondrial membrane potential dissipation, elevated ROS production, an increased Bax/Bcl-2 ratio, and enhanced expression of cleaved caspase-3. Functionally, hymenochirin-1B reduced tumor-cell migration and clonogenicity.
Dermaseptin-PS1, moderately active against S. aureus, E. coli, and C. albicans, was evaluated in several tumor cell lines and in normal endothelial cells [88]. At low concentrations (10^−6^ M), it showed selective toxicity toward U215MG glioblastoma cells while sparing normal cells. At this concentration, LDH release was minimal, yet caspase-3 cleavage was evident, indicating early activation of apoptosis. This cleavage was fully prevented by a pan-caspase inhibitor, confirming the involvement of caspases. Expression analyses revealed upregulation of multiple pro-apoptotic markers—including Bax—and increased Bax/Bcl-2 ratio, p53 phosphorylation, and cytosolic cytochrome-c release. Autophagy-related and extrinsic apoptotic pathways were excluded, demonstrating that dermaseptin-PS1 acts specifically through mitochondria-dependent apoptosis [88].
Ranatuerin-2PLx, particularly active toward S. aureus and E. coli, was tested across a range of tumor and normal cell lines [134]. At high concentrations, it was selectively toxic to tumor cells—especially PC3 prostate cancer cells—while sparing normal cells. In PC3 cells, the peptide caused membrane lysis (measured by LDH release) and induced apoptosis as demonstrated by Annexin V staining. Increased caspase-3 activity suggested involvement of an intrinsic apoptotic pathway, although it is shared by both intrinsic and extrinsic pathways; therefore, additional analyses would be needed to define the mechanism conclusively.
In some cases, frog-derived AMPs exhibit cytotoxic profiles compatible with mitochondrial apoptosis, even when no specific molecular analyses were performed. For example, temporin-La and palustrin-Ca were tested on cervical, gastric, and hepatic cancer cells [126]. Both peptides exhibited strong cytotoxicity in MTT assays. Transmission electron microscopy (TEM) analysis in HeLa cells revealed membrane damage, cytoplasmic leakage, and blurred cell boundaries, along with conspicuous mitochondrial vacuolization and loss of cristae—features consistent with mitochondrial dysfunction and suggestive of intrinsic apoptosis. Both peptides were more active against Gram-positive than Gram-negative bacteria.
The AMP pentadactylin, active against Gram-negative and Gram-positive bacteria [127], showed selective toxicity toward B16-F10 melanoma cells [128]. Cytological analyses demonstrated membrane disruption, formation of cellular protrusions, and altered nuclear morphology. The peptide also induced S-phase cell-cycle arrest, mitochondrial membrane potential loss, and DNA fragmentation—strong indicators of mitochondrial apoptosis.
(ii)Apoptosis Extrinsic Pathway (Death Receptor Pathway)
The extrinsic pathway of apoptosis is initiated by extracellular signals, typically through the binding of death ligands—such as FasL, TNF-α, or TRAIL—to their cognate receptors on the plasma membrane. This receptor-mediated mechanism plays a central role in immune regulation and in the removal of damaged, infected, or superfluous cells [160].
The peptides caerin 1.1 and caerin 1.9, exhibit broad spectrum activity against Gram-negative and Gram-positive bacteria, including MRSA strains and in vivo antimicrobial effect using murine MRSA skin infection model [68,163]. When tested on HeLa cervical cancer cells, both peptides inhibited cell proliferation as determined by MTT assay [63]. Microscopy-based studies showed that the peptides interact directly with the plasma membrane and become internalized, localizing to perinuclear regions within the first 5 min of treatment.
Comprehensive proteomic profiling of HeLa cells treated with caerin 1.1, caerin 1.9, or a combination of the two revealed substantial modulation of cellular pathways. Pathway-enrichment analyses indicated that caerin 1.1—alone or combined with caerin 1.9—upregulated biological processes related to mRNA stability and the cellular response to unfolded or misfolded proteins. Most importantly, both peptides significantly enhanced pathways linked to programmed cell death, leading to upregulation of proteins associated with apoptotic signaling. In particular, caerin 1.1 or 1.9 treatment selectively increased the apoptosis-related branch of the TNF-α signaling pathway, which was experimentally validated by elevated expression of caspase-3 and -9, consistent with activation of the extrinsic apoptosis pathway [63].
AMP-induced modulation of EGFR signaling has been described in non-tumor contexts, especially during wound healing [164,165]. Interestingly, in HeLa cancer cells, caerin peptides had the opposite effect: they downregulated the EGFR1 pathway, as confirmed by a dose-dependent reduction of PI3K/AKT pathway components [63]. These observations indicate that AMPs may either promote or inhibit EGFR signaling—along with other major pathways—depending on the physiological or pathological state of the target cells (healthy versus cancerous).
In some cases, AMPs induce cancer cell death through mechanisms bridging intrinsic and extrinsic apoptotic programs. The frog-skin peptide SSTP1 (also referred to as Temporin 1IDau1) exhibited selective cytotoxicity against human tongue squamous carcinoma HSC-4 cells, as shown by MTT assay, while sparing non-tumor cells [166]. Annexin V/PI staining revealed that SSTP1 markedly increased the proportion of cells in early apoptosis. This was supported by activation of caspase-3, -7, -9, and PARP cleavage, whereas caspase-8 remained unaffected, pointing to dominant involvement of mitochondrial (intrinsic) apoptosis.
RNA-sequencing analysis further demonstrated that SSTP1 significantly regulated TNF, JAK-STAT, and cytokine–receptor interaction pathways, suggesting participation of receptor-mediated signaling. Indeed, the peptide was shown to physically associate with the active IL-6/IL-6R/gp130 complex on the plasma membrane. This interaction selectively suppressed the pro-proliferative JAK/STAT cascade while simultaneously activating a pro-apoptotic IL-6/JNK/AP-1 pathway, revealing a complex network of signaling events converging on caspase-mediated apoptosis [166].
(iii)Autophagy-Related Pathway
The brevinin superfamily of peptides comprises the brevinin-1 and brevinin-2 groups, both known for broad-spectrum antimicrobial activity against diverse bacterial and fungal pathogens [167]. Within this family, brevinin-2R has demonstrated activity against K. pneumoniae [168] and Leishmania species [169].
Beyond its antimicrobial effects, brevinin-2R has emerged as a promising anticancer peptide. It exhibits semi-selective cytotoxicity toward multiple tumor cell lines, including T cell leukemia, B cell lymphoma, fibrosarcoma, breast and lung cancer cells, while exerting minimal or no toxicity toward peripheral blood mononuclear cells (PBMCs), normal T cells, or healthy human lung fibroblasts [59]. Remarkably, its cytotoxic potency exceeded that of standard chemotherapeutics such as doxorubicin and cisplatin in highly sensitive cancer models, including MCF-7 breast cancer and Jurkat T cell leukemia cells. Mechanistic analyses revealed that brevinin-2R does not induce caspase-dependent apoptosis. Co-treatment with a pan-caspase inhibitor failed to impair its cytotoxic effects, and no increase in expression or activation of caspase-3, -8, or -9 was detected following peptide exposure.
The mitochondrial pathway appears only partially involved in brevinin-2R-mediated cell death. In several tumor cell lines, treatment with this peptide triggered ROS generation and reduced ATP levels—hallmarks of mitochondrial dysfunction. Consistently, overexpression of the anti-apoptotic protein Bcl-2 markedly attenuated brevinin-2R cytotoxicity. However, because cell death proceeded independently of caspases, the authors examined the involvement of mitochondrial release of AIF and EndoG—key mediators of caspase-independent apoptosis [170]. Neither of these factors was released during brevinin-2R-induced cell death.
Given these observations, the authors investigated whether autophagy-associated lysosomal pathways contributed to cytotoxicity. Lethal signaling events can activate autophagy and promote cell death through lysosomal permeabilization and release of their lytic enzymes [171]. Indeed, brevinin-2R caused pronounced lysosomal swelling and permeabilization, accompanied by cytosolic translocation of cathepsin-B. The peptide also co-localized with both early and late endo-lysosomal markers, suggesting direct interaction with lysosomal membranes. Additional ultrastructural changes—including ER disintegration, cytoplasmic vacuolization, and autophagosome formation—further supported the involvement of autophagy.
Collectively, these findings indicate that brevinin-2R exerts its anticancer activity primarily through induction of autophagy and lysosome-mediated cell death, with a secondary contribution from partial activation of mitochondrial dysfunction [59].
(iv)Necroptosis Pathway
Necroptosis is a form of programmed cell death positioned at the interface between unregulated necrosis and caspase-dependent apoptosis. It is typically activated during chronic inflammatory conditions or viral infections. Central to this pathway are the RIPK3 kinase and its downstream effector MLKL, whose phosphorylation triggers membrane permeabilization and cell disruption [172].
Among amphibian AMPs, tigerinin-1 shows potent activity against S. aureus [173] and displays notable cytotoxicity toward various cancer cell types—including hepatoma, breast, lung, and prostate cancer cells. In particular, tigerinin-1 exhibits pronounced efficacy against A549 lung cancer cells [153]. In this cell line, the peptide reduces viability by promoting plasma membrane rupture and compromising lysosomal integrity, as demonstrated by LDH release and neutral red uptake (NRU) assays, respectively.
At the molecular level, tigerinin-1 selectively enhances the expression and phosphorylation of necroptosis-associated proteins RIP/p-RIP and MLKL/p-MLKL, while leaving levels of classical apoptotic markers (total or cleaved caspase-3 and -8) unchanged. These findings indicate that tigerinin-1 cytotoxic activity proceeds independently of apoptosis. Because necroptosis is frequently accompanied by elevated production of reactive oxygen species (ROS) [172], it is notable that tigerinin-1 induces a dose-dependent increase in ROS and related oxidative-stress markers—including lactoperoxidase (LPO), nitric oxide (NO), protein carbonyl (PC), and hydrogen peroxide (H_2_O_2_)—in lung cancer cells. This oxidative burden contributes to significant DNA damage, further compromising tumor cell viability.
Functionally, tigerinin-1 markedly reduces colony formation of A549 cells in soft agar, indicating inhibition of anchorage-independent growth. Moreover, when tested on HUVEC endothelial cells, the peptide suppresses capillaries formation, suggesting a potential angiostatic effect that may augment its antitumor activity [153].
2.2. Functional Consequences of AMP Treatment on Tumor Cells
For several of the anticancer AMPs listed in Table 1, the molecular mechanisms underlying their cytotoxic effects have not yet been fully elucidated. Nonetheless, a range of in vitro experimental studies has provided valuable insights into how these peptides functionally modulate tumor cell behavior. Several AMPs have been evaluated using different in vivo experimental tumor models, including syngeneic murine leukemia systems and human tumor xenografts in immunodeficient mice, highlighting the diversity of preclinical approaches used to assess their anticancer therapeutic potential.
2.2.1. Regulation of Tumor Cell Proliferation, Migration, and Clonogenicity
The Xenopus-derived AMP magainin-2 and its two analogues—MSI-136 (“all-L”) and MSI-238 (“all-D”)—were initially tested in vitro on Ehrlich ascites tumor cells and A549 lung cancer cells using MTT assays. In both cell lines, the two analogues exhibited a more pronounced cytotoxicity compared with native magainin-2 [118].
To further evaluate their anticancer potential, the three peptides were tested in vivo in mouse models of leukemia, sarcoma, and spontaneous ovarian teratoma. In a syngeneic P388 leukemia ascites model in DBA/2 mice, where peptides were administered intraperitoneally, MSI-136 and MSI-238 modestly extended animal survival, though to a lesser extent than cisplatin. Magainin-2 produced a comparable effect only at higher doses. In syngeneic S180 sarcoma model, both analogues—but not magainin-2—achieved 100% increases in life span relative to untreated controls. The most compelling results came from studies in spontaneous ovarian teratoma in LT/Sv mice, where magainin-2 and its analogues significantly increased survival and delayed ascites tumor growth, reaching efficacy levels comparable to doxorubicin. Histopathological analyses showed that MSI-136 and MSI-238 also reduced tumor colonization of the peritoneum and mesenteric lymph nodes.
In terms of antimicrobial activity, magainin-2 is active against S. aureus mprF mutants [117] and Batrachochytrium dendrobatidis [20].
The AMPs dermaseptin-B2 and dermaseptin-B3, purified from Phyllomedusa bicolor skin extracts, were active against Gram-negative and Gram-positive bacteria [174] including S. aureus, P. aeruginosa, and E. coli [83]. Additionally, they have also shown significant anticancer activity, inhibiting PC-3 prostate cancer cell proliferation. They markedly suppressed anchorage-independent colony formation in soft agar, achieving effects comparable to 5-fluorouracil, and displayed anti-angiogenic activity in endothelial cell assays. Crucially, these antitumor effects were reproduced with synthetic versions of the peptides, emphasizing their drug-development potential [175].
The anticancer role of dermaseptin-B2 has been confirmed further in breast cancer cells [81]. In this study, MCF-7 tumor cells and non-tumor MCF-10A cells were transduced with a plasmid expressing dermaseptin-B2. Growth assays using crystal violet staining showed that dermaseptin-B2 expression significantly impaired MCF-7 proliferation while exerting only minor effects on MCF-10A cells. Consistently, tumor-specific reductions in colony formation in soft agar were observed.
Supporting these findings, wound-healing assays demonstrated a substantial inhibition of cell migration in dermaseptin-B2-expressing MCF-7 cells. Flow-cytometry analyses further revealed that dermaseptin-B2 expression selectively increased apoptotic susceptibility and induced G0/G1 cell-cycle arrest in tumor but not normal cells. At the molecular level, dermaseptin-B2 reduced the expression of anti-apoptotic genes, increased pro-apoptotic gene expression, and inhibited components of the pro-proliferative AKT1/AKT3 signaling pathway, highlighting its multi-level suppression of tumor growth and survival.
2.2.2. Regulation of Immune Response
The AMP esculentin-2CHa and its double-substituted [D20K,D27K] analog exhibited antimicrobial activity against S. aureus and E. coli, and displayed dose-dependent cytotoxicity toward A549 lung cancer cells [176]. Beyond its antimicrobial and anticancer effects, esculentin-2CHA also shows immunomodulatory properties. When administered to mouse peritoneal macrophages, it did not alter IL-1β or IL-6 expression, either in the presence or absence of LPS stimulation. However, it significantly increased TNF-α expression compared with untreated controls and further enhanced TNF-α levels when combined with LPS. The peptide also modulated lymphoid cells, inducing a significant upregulation of the anti-inflammatory cytokine IL-10 alone or in combination with concanavalin A (known T cells mitogen), relative to their respective controls.
Two additional frog-skin-secretion-derived peptides, pseudohymenochirin-1Pb and pseudohymenochirin-2Pa, were examined for cytotoxicity in several tumor cell lines, including lung, metastatic breast, and colon cancer cells [131]. Among them, pseudohymenochirin-1Pb displayed selective, dose-dependent cytotoxicity toward cancer cells, whereas pseudohymenochirin-2Pa showed non-selective toxicity affecting non-tumor cells even at low doses. Their immunomodulatory potential was assessed in mouse macrophages: both peptides inhibited the expression of the anti-inflammatory cytokine IL-10 while increasing the production of the pro-inflammatory cytokine IL-23, including under LPS stimulation. Neither peptide affected IL-1β or TNF-α levels. Interestingly, IL-6—whose inflammatory role can vary with context [177]—was consistently downregulated. Both peptides displayed strong activity against Gram-positive bacteria, including multidrug-resistant clinical isolates, but showed only moderate effects against Gram-negative bacteria and C. albicans [131].
The peptides frenatin-2.1S and frenatin-2.2S also demonstrated potent antimicrobial activity against clinical isolates of MRSA and S. epidermidis, though they exhibited weaker activity against Gram-negative bacteria [103]. Both peptides displayed moderate cytotoxicity toward A549 lung cancer cells and were evaluated for immunomodulatory effects in mouse peritoneal macrophages. Consistent with trends observed for other frog-skin AMPs, frenatin-2.1S enhanced TNF-α expression in LPS-stimulated macrophages. Moreover, both frenatin-2.1S and frenatin-2.2S increased IL-1β and IL-23 levels in the presence of LPS. Notably, frenatin-2.2S downregulated IL-10 expression under LPS stimulation, while IL-6 remained unaffected by either peptide.
The antitumor and immunomodulatory potential of frenatin-2.1S was further investigated in vivo [104]. Although the peptide showed no effect on the viability of 4T1 mouse mammary carcinoma cells in vitro, as measured by real-time impedance-based cell index [178], its administration to a human A549 lung cancer xenograft model in BALB/c nude mice caused significant inhibition of tumor growth and markedly influenced immune-cell recruitment. Frenatin-2.1S increased the numbers of CD3^+^ T cells and CD11c^+^ dendritic cells in the peritoneal cavity. In addition, it elevated both the abundance and activation status of NK cells, as evidenced by upregulation of the cell surface markers NKG2D, FasL, CD69, and CD107a. Functionally, NK cells isolated from frenatin-2.1S-treated mice, but not peritoneal leukocytes, induced significant cytolysis of 4T1 cells in co-culture. These findings suggest that frenatin-2.1S exerts indirect antitumor effects by promoting the enrichment and activation of cytolytic NK cells [104].
2.3. Conservation of AMP Mechanisms of Action Across Species
Amphibian-derived AMPs represent a broad class of antimicrobial peptides whose anticancer mechanisms have been outlined in detail in the previous sections. The literature also contains abundant experimental data, compiled in both recent and earlier reviews, derived from studies of AMPs originating from a variety of other species, including humans, bovines, insects, and fish, as well as peptides of synthetic origin [28,179,180]. Notably, the mechanisms of action described for these dual-function peptides are largely comparable to those reported by us and others for amphibian AMPs [181].
These mechanisms frequently fall into several major categories, including membrane permeabilization or lysis; induction of apoptosis (via both intrinsic and extrinsic pathways); necrosis; cytolysis; regulation of the cell cycle; inhibition of angiogenesis; and modulation of immune responses. An exception is represented by certain amphibian AMPs, such as buforin, whose activity also involves mechanisms related to the inhibition of protein synthesis and interference with DNA/RNA replication [182].
Taken together, these observations suggest that the mechanisms underlying the dual antimicrobial and anticancer functions of many AMPs are largely conserved across species, highlighting their potential as a broad-spectrum defense strategy against both infections and tumor progression.
2.4. Translational Implications and Safety Concerns of Dual-Action Amphibian AMPs
AMPs with dual activity generally offer the advantage of being potentially applicable to cancers that are driven or promoted by bacterial infections. For instance, gastric cancer can arise as a consequence of infection with H. pylori [183], gallbladder cancer may be originated by infection with S. typhi [184] or bladder cancer can be caused by infection of different pathogens, including Streptococcus, Anaerococcus, Aerococcus and Pseudomonas [185]. Furthermore, the use of AMPs with dual functionality may be beneficial during anticancer therapies that render patients particularly susceptible to opportunistic infections, either due to prolonged hospitalization or as a result of therapy-induced immunosuppression [186,187].
As previously mentioned, the tumor microbiota plays an important role in tumor progression. Dual-function AMPs may therefore be employed to eliminate harmful bacteria that are part of the tumor-associated microbiota [188]. In addition, AMPs generally display a relatively low propensity to induce drug resistance and are reasonably biocompatible, including those of non-natural origin that are synthetically produced to confer increased specificity, improved tolerability, or other advantageous properties [189]. Importantly, they can also be used in combination with conventional anticancer drugs, potentially enhancing their therapeutic efficacy through synergistic effects [190,191].
The disadvantages represent the other side of the same coin. While dual-function AMPs may target harmful components of the microbiota and contribute to slowing tumor progression, they may also disrupt beneficial microbial communities, such as those of gut or cervical microbiota [192]. Moreover, they may pose risks to normal host cells, exhibiting cytotoxic and hemolytic effects or triggering excessive immune responses or inflammatory reactions [193,194].
An additional drawback is the relatively high peptide concentrations that are often required to achieve a significant therapeutic effect, also due to the high susceptibility to proteolytic degradation [195,196]. Intratumoral delivery of peptides, when feasible, is generally more efficient compared to systemic administration [197,198].
With regard to natural amphibian AMPs, at present there are relatively few clinical trials and limited studies employing relevant tumor models, although some investigations have been conducted using modified or enhanced amphibian peptides [199,200,201]. The modification of specific amino acid residues, the addition of functional groups, and/or the use of these peptides in combination with conventional anticancer drugs are likely to represent key strategies for maximizing the translational potential of dual-function antimicrobial peptides.
3. Concluding Remarks
Overall, analysis of the available literature indicates that amphibian-derived AMPs represent a promising and versatile class of anticancer candidates with clear translational potential. Although the body of work addressing this topic remains heterogeneous in terms of experimental approaches, tumor models, and analytical methodologies, the accumulated evidence consistently supports the notion that amphibian AMPs exert reproducible and biologically meaningful antitumor effects. Importantly, these effects converge on a limited number of conserved mechanisms, suggesting that AMP-based anticancer strategies could be rationally optimized for therapeutic development.
A more detailed analysis of the literature revealed an important structural feature of AMPs exhibiting both antimicrobial and antitumor activities: most of the peptides analyzed contain at least one α-helical segment within their sequence (see Table 1). It is well established that α-helical secondary structure is one of the factors that enhance the ability of AMPs to interact with and disrupt bacterial and/or mammalian cellular membranes [202]. This structural characteristic appears to be a common motif among anticancer amphibian AMPs and may therefore partly explain the mechanisms underlying their dual biological activity.
From a translational perspective, the preferential interaction of many amphibian AMPs with tumor cell membranes—largely attributed to their net positive charge and the altered lipid composition of cancer cell surfaces—offers a mechanistic basis for tumor selectivity. While membrane disruption and cytolysis have been reported for both cationic and anionic AMPs, membranolysis emerges as a recurrent and therapeutically relevant mechanism. Nonetheless, distinguishing between direct membrane damage and secondary membrane alterations arising from downstream cell death pathways remains challenging and highlights the need for standardized assays and time-resolved analyses in future preclinical studies.
Although the cationic nature of AMPs remains a valid parameter for predicting tumor selectivity, an important challenge for membranolytic peptides is to discriminate between genuine tumor selectivity and nonspecific cytotoxicity. This difficulty is largely attributable to the common practice of administering peptides either to bacterial strains or to in vitro cultures of cell lines, with cell viability typically used as the primary readout in both cases. Whenever possible, we sought to provide information regarding the effects of AMPs on normal versus cancer cells in order to assess whether tumor specificity was indeed supported by the available data.
Crucially, the prevailing anticancer activity of amphibian AMPs appears to rely on the activation of regulated cell death programs rather than nonspecific cytotoxicity. Comprehensive molecular investigations across multiple tumor cell lines have demonstrated that these peptides induce apoptosis via both intrinsic (mitochondrial) and extrinsic (death receptor-mediated) pathways, mechanisms that are already clinically exploited by several approved anticancer agents. The additional involvement of alternative cell death modalities, including autophagy and necroptosis, is of particular clinical relevance, as it suggests that AMPs may overcome resistance mechanisms associated with apoptosis-defective or therapy-refractory tumors.
Functionally, amphibian AMPs consistently suppress tumor cell proliferation, survival, and clonogenic expansion in vitro, including under three-dimensional and anchorage-independent growth conditions that more closely recapitulate in vivo tumor architecture. Their ability to impair tumor cell migration and motility further underscores their potential to limit invasion and metastatic spread—key determinants of poor clinical outcome. Although still limited in number, in vivo studies using mouse tumor models provide compelling proof-of-concept evidence that AMP treatment can reduce tumor growth and dissemination without overt systemic toxicity, thereby supporting continued preclinical development.
Beyond their direct effects on tumor cells, amphibian AMPs also exhibit immunomodulatory properties that may enhance their therapeutic value. Studies using immune cell cultures, particularly macrophages, indicate that anticancer AMPs promote a pro-inflammatory milieu by increasing the production of pro-inflammatory cytokines while suppressing anti-inflammatory signals. This dual activity—combining direct tumor cell killing with immune activation—positions amphibian AMPs as attractive candidates for multimodal cancer therapies, including combination strategies with chemotherapy, radiotherapy, or immune checkpoint inhibitors.
A crucial point is to determine to which extent the dual antimicrobial and anti-tumor activities of AMPs are truly coupled or simply converge on membrane-targeting mechanisms. Defining this would require specific experimental settings or in vivo models, because conventional 2D in vitro cultures of tumor cells fail to accurately reproduce the complexity of the in vivo tumor microenvironment, e.g., tissue organization, vascularization, immune infiltration, and the microbiota. This latter is a key regulator of tumor progression and therapeutic response [203].
Thus, a proof of concept of full functional duality would be identifying AMPs capable of concurrently suppressing tumor growth and bacterial proliferation in adequate oncology models where microbiota alterations and tumor progression are linked. This aspect, at least for amphibian AMPs, remains insufficiently explored.
Consequently, the lack of such specific studies makes it difficult to predict whether antimicrobial potency correlates with tumor selectivity. Notably, a substantial portion of the literature addressing the application of AMPs in oncology is based on synthetic derivatives that preserve the core structural features of AMPs—predominantly an α-helical conformation—while incorporating additional modifications designed to enhance tumor specificity [28]. Taken together, the available evidence supports amphibian-derived AMPs as promising leads for anticancer drug development. Future translational efforts should focus on improving peptide stability, bioavailability, and tumor selectivity, as well as on defining optimal delivery strategies and combination regimens. Addressing these challenges through rigorous preclinical and early-phase clinical studies will be essential to advance amphibian AMPs from experimental systems toward clinically viable anticancer therapeutics.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Khurana P. Binti Raja Ibrahim R.Z.A. Omar K. Ahmad A.B. Frontline Mental Resilience: Lessons Learned from the Pandemic Experience Australas. Emerg. Care 2025 in press 10.1016/j.auec.2025.09.00240946005 · doi ↗ · pubmed ↗
- 2Ruiz-Ramos J. Integrating Global Surveillance, Local Action, and Innovative Stewardship Against Antimicrobial Resistance Antibiotics 20251483510.3390/antibiotics 1408083540868029 PMC 12382669 · doi ↗ · pubmed ↗
- 3Butler M.S. Capon R.J. Blaskovich M.A.T. Henderson I.R. Natural Product-Derived Compounds in Clinical Trials and Drug Approvals Nat. Prod. Rep.202543208810.1039/D 5NP 00031 A 40923383 · doi ↗ · pubmed ↗
- 4Larsson D.G.J. Flach C.F. Antibiotic Resistance in the Environment Nat. Rev. Microbiol.20222025726910.1038/s 41579-021-00649-x 34737424 PMC 8567979 · doi ↗ · pubmed ↗
- 5Renwick M.J. Brogan D.M. Mossialos E. A Systematic Review and Critical Assessment of Incentive Strategies for Discovery and Development of Novel Antibiotics J. Antibiot.201669738810.1038/ja.2015.98PMC 477554026464014 · doi ↗ · pubmed ↗
- 6Bizuayehu H.M. Ahmed K.Y. Kibret G.D. Dadi A.F. Belachew S.A. Bagade T. Tegegne T.K. Venchiarutti R.L. Kibret K.T. Hailegebireal A.H. Global Disparities of Cancer and Its Projected Burden in 2050 JAMA Netw. Open 20247 e 244319810.1001/jamanetworkopen.2024.4319839499513 PMC 11539015 · doi ↗ · pubmed ↗
- 7Denisov E.V. Litviakov N.V. Zavyalova M.V. Perelmuter V.M. Vtorushin S.V. Tsyganov M.M. Gerashchenko T.S. Garbukov E.Y. Slonimskaya E.M. Cherdyntseva N.V. Intratumoral Morphological Heterogeneity of Breast Cancer: Neoadjuvant Chemotherapy Efficiency and Multidrug Resistance Gene Expression Sci. Rep.20144470910.1038/srep 0470924736530 PMC 3988480 · doi ↗ · pubmed ↗
- 8Travis W.D. Brambilla E. Noguchi M. Nicholson A.G. Geisinger K. Yatabe Y. Ishikawa Y. Wistuba I. Flieder D.B. Franklin W. Diagnosis of Lung Cancer in Small Biopsies and Cytology: Implications of the 2011 International Association for the Study of Lung Cancer/American Thoracic Society/European Respiratory Society classification Arch. Pathol. Lab. Med.201313766868410.5858/arpa.2012-0263-RA 22970842 PMC 4509741 · doi ↗ · pubmed ↗