OsOFP3 Negatively Regulates Heat Stress Tolerance by Modulating H2O2 Homeostasis and Stomatal Aperture in Rice
Guiyuan Yu, Yingfeng Wang, Guilian Zhang, Huabing Deng, Wenbang Tang, Lifeng Wang, Yunhua Xiao

TL;DR
This study shows that the OsOFP3 protein in rice reduces heat stress tolerance by affecting hydrogen peroxide balance and stomatal behavior.
Contribution
OsOFP3 is identified as a novel negative regulator of heat stress tolerance in rice through its molecular interactions and physiological effects.
Findings
OsOFP3 overexpression reduces heat tolerance and increases reactive oxygen species in rice.
OsOFP3 mutants show enhanced heat stress resistance and improved stomatal closure.
OsOFP3 interacts with OsHTAS, a positive regulator of heat tolerance, through its RING domain.
Abstract
Rice (Oryza sativa) is a staple crop that is highly susceptible to heat stress (HS), which severely impairs growth and yield. In this study, we identified the rice Ovate Family Protein OsOFP3 as a novel negative regulator in response to heat. Our results demonstrate that the expression of OsOFP3 is suppressed at both the transcriptional and protein levels under HS. Overexpression of OsOFP3 significantly reduces the survival rate of rice seedlings under HS and exacerbates chlorophyll degradation, membrane damage, and the accumulation of reactive oxygen species (H2O2 and O2−). In contrast, OsOFP3 mutants exhibit enhanced heat tolerance. Moreover, OsOFP3-overexpressing plants display increased stomatal opening and decreased stomatal closure under HS. Molecular interaction analysis further reveals that OsOFP3 interacts with the C-terminal domain of OsHTAS, a known positive regulator of heat…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
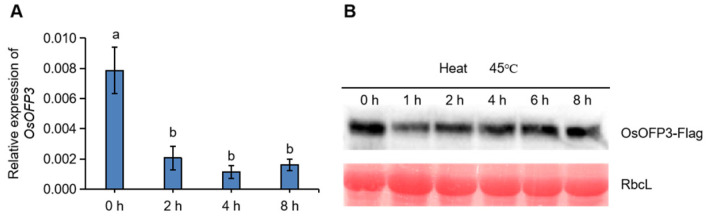 Figure 1
Figure 1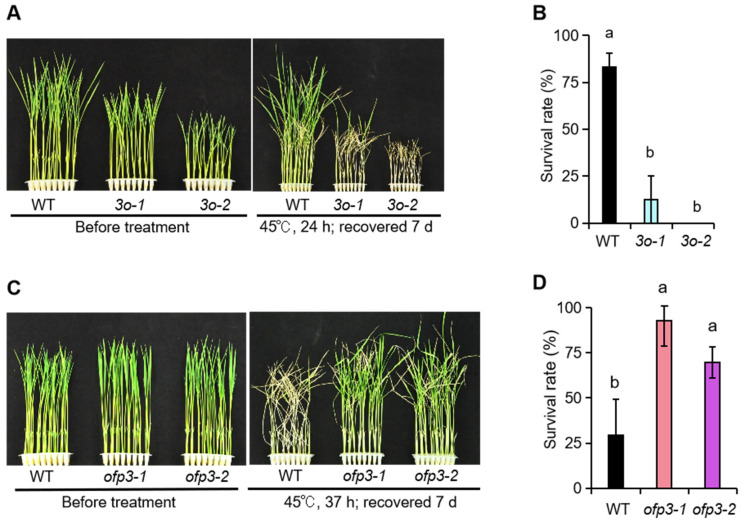 Figure 2
Figure 2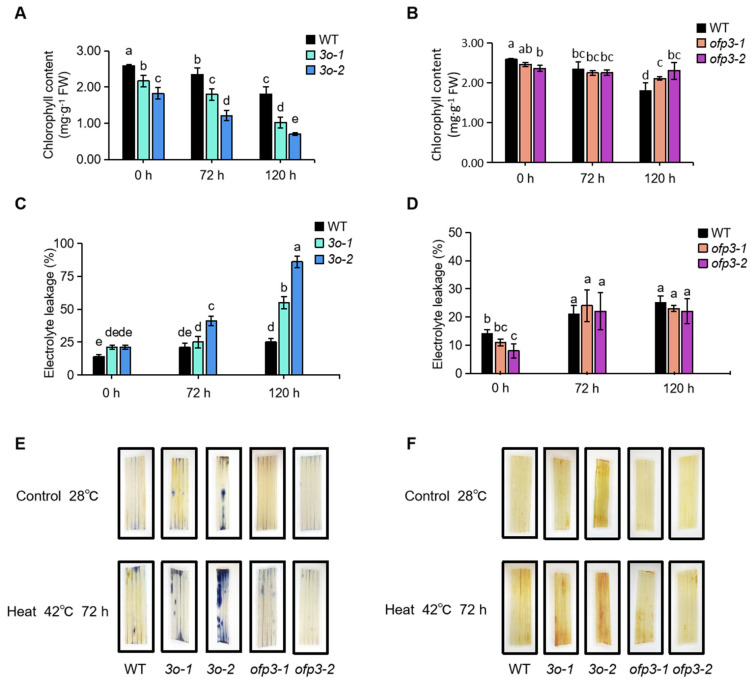 Figure 3
Figure 3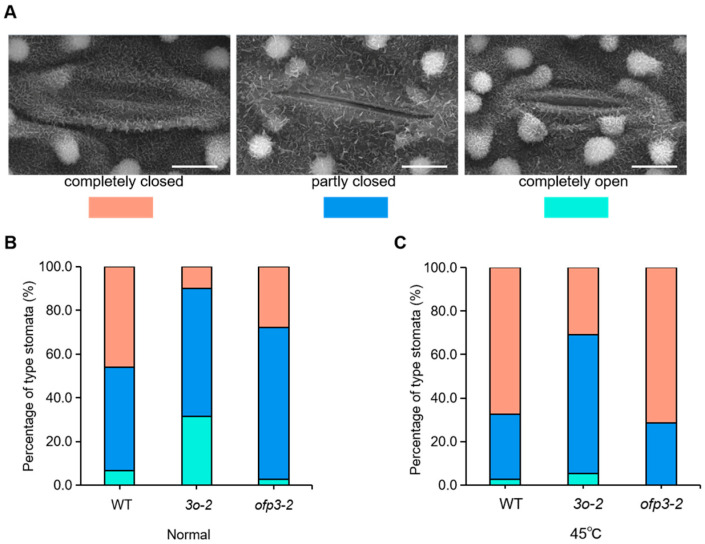 Figure 4
Figure 4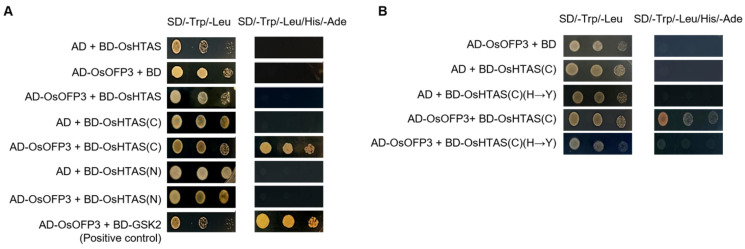 Figure 5
Figure 5- —National Natural Science Foundation of China
- —Natural Science Foundation of Hunan Province
- —Natural Science Foundation of Changsha
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant responses to water stress · Heat shock proteins research · Plant Stress Responses and Tolerance
1. Introduction
Under the exacerbation of global climate change, HS has become one of the major abiotic stresses limiting crop yield and quality [1]. Rice is the staple food for more than half of the world’s population [2]. The stability of rice production is crucial to food security. HS affect multiple critical growth stages of rice, directly disrupting the physiological metabolic balance. For photosynthesis, heat damages the structure and function of the thylakoid membrane and inhibits the activity of Rubisco activase (RCA), resulting in the decreased efficiency of photosystem II and carbon assimilation [3,4]. Scafaro also suggested that overexpression of RCA in wild rice significantly ensured the normal growth and increased the yield of cultivated rice under HS, suggesting that engineering RCA may be an effective way for heat tolerance breeding [5]. Studies have shown that when rice is exposed to HS, reactive oxygen species (ROS) accumulate within the cells, intensifying lipid peroxidation and protein oxidation of the cell membranes, thereby damaging the structure and function of the biological membranes and ultimately leading to cell death [6], increased chalky grain rate [7], seedling death [8], and even spikelet sterility [9,10]. In addition, the content of endogenous hormones will change when rice is subjected to stress. Research has demonstrated that under HS, the ethylene-mediated signaling pathway mitigates oxidative damage in rice seedlings, preserves chlorophyll content, and enhances plant heat stress tolerance [11]. HS can also induce an increase in the abscisic acid (ABA) content in rice grains and pollen, which leads to pollen abortion and yield reduction [12].
When plants are subjected to HS, they will initiate a series of complex physiological and molecular response mechanisms. The movement of stomata is the core link in the regulation of leaf water transpiration and body temperature; stomatal closing can alleviate water loss caused by HS, which is an important strategy for plants to improve heat tolerance [13]. Hydrogen peroxide (H_2_O_2_), as a kind of stable ROS, plays a dual role in this process: at low concentrations, it acts as a signaling molecule, participates in acclimatory signaling, and triggers tolerance to various biotic and abiotic stresses; and at high concentrations, it causes programmed cell death [14]. In stomatal guard cells, H_2_O_2_ is a key secondary messenger in the ABA signaling pathway, which can activate calcium channels and eventually induce stomatal closure [15]. Interestingly, an ABA-independent mechanism of stomatal closure was also revealed in rice: the drought and salt tolerance factor DST, functioning as a zinc finger transcription factor, negatively regulates H_2_O_2_-induced stomatal closure by directly modulating the expression of genes involved in hydrogen peroxide scavenging [16]. Therefore, maintaining cellular H_2_O_2_ homeostasis is a critical determinant of heat tolerance in rice.
The regulation of H_2_O_2_ homeostasis and stomatal movement relies on a sophisticated signaling network. The ubiquitin-26S proteasome system (UPS)-mediated post-translational modification is a core mechanism for plants to rapidly respond to environmental stress and precisely regulate signaling pathways [17]. The E3 ubiquitin ligase-containing RING domains play a critical “executor” role in this process by specifically recognizing and ubiquitinating substrate proteins [18]. Numerous studies have indicated that RING finger E3 ligases are extensively involved in the post-translational regulatory networks of plant hormone signaling and abiotic stress responses [19]. For instance, the RING E3 ligase XERICO positively regulates drought resistance by promoting ABA biosynthesis in Arabidopsis thaliana [20], while HOS1 participates in low-temperature responses by negatively regulating cold signal transduction [21]. The C3HC4 RING finger E3 ligase OsDIS1 negatively regulates drought stress tolerance by modulating multiple stress-related genes and potentially through the post-translational modification of OsNek6 in rice [22]. Hot pepper RING MEMBRANE-ANCHOR 1 HOMOLOG1 functions as an E3 ligase, ubiquitinating the plasma membrane aquaporins PIP2;1 under water-deficient conditions [23]. It is worth noting that the RING finger E3 ligase OsHTAS, as a positive regulator of heat tolerance, enhances the heat tolerance of rice seedlings by promoting H_2_O_2_ accumulation and inducing ABA-dependent stomatal closure [24].
The Ovate Family Proteins (OFPs) are a class of plant-specific transcription factors that are extensively involved in various processes such as plant growth and development and hormone signal transduction [25]. The OFP family was originally identified in tomato as a key quantitative trait locus controlling fruit shape; overexpression of OFP leads to phenotypic changes such as plant dwarfing and organ size reduction [26]. A total of 19 OFP gene family members have been identified in Arabidopsis thaliana, 33 in rice, and 45 in maize (Zea mays) [27]. Overexpression of OsOFP2 resulted in plant dwarfing and leaf morphological changes [28]. Overexpression of OsOFP3 resulted in BR insensitivity and decreased plant height, while deletion of OsOFP3 promoted seedling growth [29]. OsOFP6 is expressed in various tissues of rice, and OsOFP6-RNAi plants exhibited faster water loss and higher H_2_O_2_ contents under drought conditions [30]. OsOFP8 plays a positive role in BR signaling [31]. Overexpression of OsOFP19 leads to reduced plant height, thicker leaves, and sturdier stems in rice [32]. These studies revealed the multiple functions of the OFP family in plant growth and development. Although a few OFP members have been confirmed to participate in abiotic stress responses, their specific functions and molecular mechanisms in heat stress response remain poorly understood.
In this study, we identified a rice gene, OsOFP3, that was significantly inhibited by HS. Analysis revealed that overexpression of OsOFP3 aggravated chlorophyll degradation, membrane damage, and reactive oxygen species accumulation under HS. Furthermore, OsOFP3 interacts with OsHTAS, which may negatively regulate heat tolerance by modulating the H_2_O_2_ signaling pathway.
2. Materials and Methods
2.1. Plant Materials and High-Temperature Stress Treatments
Zhonghua 11 (Oryza sativa L. subsp. japonica cv. ZH11) was used as a control, and the overexpressed lines (3o-1, 3o-2) and mutant lines (ofp3-1, ofp3-2) were constructed on ZH11 as the genetic background. We produced OsOFP3 knockout mutants using CRISPR/Cas9 editing. By targeting a sequence at the 50-end of the coding sequence, some independent homozygous mutant lines were obtained. Two alleles, ofp3-1 and ofp3-2, containing a 1 bp insertion and a 386 bp deletion, respectively, were selected for the detailed analysis. The OsOFP3-Flag fusion overexpressed material was used to examine protein expression under HS. The seeds were immersed at 37 °C to accelerate germination (changing water for 12 h). The well-germinated seeds were seeded in a 96-well PCR plate with holes in the bottom, and then hydroponic culture was carried out in a light incubator at 28 °C. The culture was changed to modified Kimura B nutrient solution from the fourth day of seeding, and the nutrient solution was replaced on days 4, 6, 8, 9, and 10. The light incubator was set to a light cycle of 12 h light/12 h dark, light intensity of 540 μmol m^−2^ s^−1^, relative humidity of 70%, normal temperature treatment of 28 °C, and heat treatment of 42 °C or 45 °C. The materials could not survive for a long time at 45 °C. In order to observe the change trends of the plant morphology and physiological indexes under the high-temperature treatment for 5 days, the temperature was changed to 42 °C.
2.2. Expression Analysis of OsOFP3 Under High-Temperature Stress
The wild-type ZH11 was used as the material, and the seedlings were treated at 45 °C for 0, 2, 4, and 8 h. The extraction of total RNA, reverse transcription, and qRT-PCR detection were performed according to the method described previously [33]. The gene expression was quantified using the 2^−∆∆CT^ method, and three biological replicates were applied throughout the assay. The primer sequences used for qRT-PCR are shown in Supplemental Table S1. The OsOFP3-Flag fusion overexpressed material was used for Western blotting. After sampling at the same time points, the protein was extracted, separated by SDS-PAGE, wet transferred to the membrane, incubated with antibody, and protein expression changes were ultimately analyzed by ECL chemiluminescence [34].
2.3. NBT and DAB Staining
NBT and DAB staining were performed according to the method described by Kaur [35]. The specific steps were as follows: Rice leaves treated at 42 °C for 72 h were cut into approximately 3 cm segments and separately immersed in DAB solution (1 mg/mL) and NBT solution (1 mg/mL) prepared with double-distilled water (pH 3.8) and were penetrated under vacuum at 60 kpa for 10 min. The samples were subsequently immersed in absolute ethanol and then stored in a water bath at 100 °C until the chlorophyll faded completely. Finally, images were captured using a microscope.
2.4. Chlorophyll and Electrolyte Leakage Detection
Leaf samples of 0.1 g were weighed and cut into pieces, 95% acetone-ethanol extract was added, and the samples were wrapped in tin foil and placed in the refrigerator at 4 °C overnight until the filaments turned white. The absorbance values were measured at wavelengths of 645 nm and 663 nm. The chlorophyll concentration was calculated according to the formulas, Ca = 12.7 × A_663_ − 2.69 × A_645_, Cb = 22.9 × A_645_ − 4.86 × A_663_, Total chlorophyll concentration = Ca + Cb. A_645_ is the absorbance value of the extract at 645 nm, A_663_ is the absorbance value of the extract at 663 nm. Chlorophyll content = CV/(1000A), where C represents the chlorophyll concentration (mg·L^−1^), V represents the total volume of extract (mL), and A represents the fresh weight of leaves (g).
The leaves were washed with distilled water, blotted to dry the surface water, and cut into about 4 cm long strips avoiding the main vein. Next, 0.1 g of sample was placed in 10 mL of distilled water for 12 h before the conductance value (R_1_) was determined. Then, the samples were heated in a water bath at 100 °C for 30 min, cooled to room temperature, shaken, and the conductance value (R_2_) was measured again. The relative electrical conductivity was calculated as R_1_/R_2_ × 100%. The conductivity meter was calibrated at two points with a standard solution before use.
2.5. Measurement of the Degree of Stomatal Opening
Seedlings were sampled after 2 h of heat treatment at 45 °C. To reduce the influence of position, individuals with uniform growth that were located in the middle region of the culture apparatus were selected, and the use of edge row materials was avoided. About 3–5 mm of tissue was taken from the shoot tip, and three biological replicates were set for each material. Immediately after sampling, samples were transferred to 2 mL EP tubes containing electron microscope fixative, evacuated for 5 h, and subsequently stored wrapped in tin foil at 4 °C until examination. After scanning electron microscope observation, three pictures of each treatment group (room temperature/high temperature) were selected under the same field of view, and 50 stomata were counted in each picture. The stomata were classified and counted according to their opening state (completely open, partly open, completely closed).
2.6. Yeast Two-Hybrid Assays
The coding sequences of the C-terminus containing the RING finger domain (amino acids 338–414), the N-terminus (amino acids 1–337), and the full-length OsHTAS protein were amplified and cloned in frame with the GAL4 DNA binding domain of the pGBKT7 vector to generate GAL4 DNA-BD fusion constructs, called BD-OsHTAS(C), BD-OsHTAS(N) and BD-OsHTAS, respectively. Then, the coding sequence of OsOFP3 was amplified and cloned in pGADT7, called AD-OsOFP3. The prey and bait proteins were co-transformed into AH109 yeast cells using the lithium acetate method, and transformants were selected on synthetic dextrose/–Ade/–His/–Leu/–Trp plates. Colony growth was observed after 3–5 days of incubation at 30 °C. AD-OsOFP3 + BD-GSK229 was used as a positive control, and the empty vector combination was used as a negative control.
2.7. Statistical Analysis
All results are presented as the mean ± SD (n = 3). Statistical analyses were performed using DPS (v7.05). One-way ANOVA was used to statistically analyze the data, and differences were considered significant at p < 0.05 by Duncan’s multiple range test.
3. Results
3.1. OsOFP3 Expression and Protein Accumulation Decreased Under HS
We used Plant CARE (http://bioinformatics.psb.ugent.be/, accessed on 15 March 2025) to analyze the cis-acting elements in the OsOFP3 promoter region, which is rich in multiple response elements related to plant hormones, such as ABA and Gibberellic acid (GA), as well as to anaerobic and low temperature stress (Supplemental Figure S1). The result implies that OsOFP3 may be involved in plant growth and development and stress resistance. To explore the possible regulatory mechanism of OsOFP3 in HS, we analyzed the changes in OsOFP3 at the transcriptional and protein levels by using quantitative real-time PCR and Western blotting, respectively. The results showed that the expression level of OsOFP3 significantly decreased over time in rice plants treated at 45 °C (Figure 1A). Western blot analysis showed that protein accumulation significantly decreased after one hour of heat treatment, followed by a gradual recovery, although it remained lower than the control (Figure 1B). These results indicate that HS suppresses OsOFP3 accumulation at both the transcriptional and protein levels, suggesting that OsOFP3 may participate in the heat response by down-regulating the expression and protein degradation.
3.2. OsOFP3 Negatively Regulates Heat Tolerance During the Seedling Stage in Rice
To further investigate the effect of OsOFP3 on rice under HS, we subjected WT, OsOFP3-overexpressing plants (3o-1, 3o-2), and OsOFP3 mutants (ofp3-1, ofp3-2) to high-temperature treatment. The results showed that the survival rate of both 3o-1 and 3o-2 was significantly lower than that of the WT. The survival rate of the WT was more than 80%, while that of 3o-1 was less than 25%, and all the 3o-2 plants died (Figure 2A,B). The survival rates of both ofp3-1 and ofp3-2 were significantly higher than the WT (Figure 2C,D). These results suggest that OsOFP3 negatively regulates the tolerance of rice seedlings to HS.
3.3. OsOFP3 Overexpression Leads to Accelerated Chlorophyll Decomposition, More ROS, and Enhanced Ion Efflux Under HS
To understand the physiological mechanism of OsOFP3 under HS, the chlorophyll content was measured after 72 h and 120 h. The results showed that the chlorophyll content of all plants decreased with the extension of high-temperature treatment time. After 72 h of treatment, the chlorophyll content of 3o-1 and 3o-2 was significantly lower than WT, while there was no significant difference between the knockout plants and the WT. After 120 h of treatment, the chlorophyll content of ofp3-2 was significantly higher than WT, while 3o-2 was less than half of the wild type (Figure 3A,B). This suggests that the overexpression of OsOFP3 accelerates chlorophyll decomposition under HS.
Under the same treatment conditions, the electrolyte leakage of 3o-2 was significantly higher than the WT at 72 h of heat treatment, but there was no significant difference in the knockout plants. After 120 h of heat treatment, the electrolyte leakages of 3o-1 and 3o-2 reached 50% and 80%, respectively, which were significantly higher than the WT, while the electrolyte leakages of ofp3-1 and ofp3-2 were slightly lower than the WT (Figure 3C,D). This result indicates that the cell membrane of the overexpressing plants was more severely damaged under HS, resulting in more ion efflux.
The accumulation of H_2_O_2_ and O_2_^−^ in leaves was detected by DAB and NBT staining after 72 h of heat treatment. The results showed that all the plant leaves exhibited increased brown and blue spots. However, the spots in the overexpressing plants were significantly darker than the WT, while the knockout plants showed a staining intensity similar to the WT (Figure 3E,F). This indicates that HS induces higher levels of ROS in overexpressing plants, potentially leading to more severe damage to the cell membrane system.
3.4. The Stomatal Aperture of the OsOFP3-Overexpressing Plants Was Higher
To investigate whether OsOFP3 is involved in this process, WT, 3o-2 and ofp3-2 were treated at 45 °C for 2 h, followed by scanning electron microscopy. The results showed that the number of stomata that were opened at 45 °C was lower than that at room temperature. However, the stomatal opening rate of 3o-2 at 45 °C was 69.3%, which was about twice that of the WT (34.6%), while the opening rate of ofp3-2 was only 28.7%, which was slightly lower than the WT (Figure 4). This suggests that OsOFP3 may regulate rice heat tolerance by affecting stomatal opening and closing. Excessive stomatal opening in overexpressing plants under HS may lead to increased water loss, thereby reducing heat tolerance.
3.5. Interaction Between OsOFP3 and OsHTAS in Yeast
Previous studies have shown that the RING finger E3 ligase OsHTAS enhanced heat tolerance by promoting H_2_O_2_ accumulation-induced stomatal closing in rice [24]. To explore whether there is an interaction between OsOFP3 and OsHTAS, we conducted a yeast two-hybrid experiment. The N-terminus of OsHTAS contains four transmembrane (TM) domains, and the C-terminus contains a RING finger domain. We constructed vectors by fusing the full-length OsHTAS (HTAS, amino acids 1–414), N-terminal region (OsHTAS(N), amino acids 1–337), or C-terminal region (OsHTAS(C), amino acids 338–414) to the BD domain. The results showed that OsOFP3 did not interact with HTAS or OsHTAS(N) but has positive interactions with OsHTAS(C) (Figure 5A). To test whether the intact RING finger domain (amino acids 338–378) of OsHTAS was indispensable for the interaction between OsHTAS(C) and OsOFP3, We introduced a point mutation by substituting histidine (His-358) with tyrosine (Tyr-358), which disrupted the catalytic core of the RING finger domain. New vectors were reconstructed and denoted as BD-OsHTAS(C)(H→Y). The results showed that BD-OsHTAS(C)(H→Y) did not interact with OsOFP3 (Figure 5B), indicating that the RING finger domain is required for the interaction between OsHTAS(C) and OsOFP3.
4. Discussion
4.1. OsOFP3 Is Heat-Suppressive and Negatively Regulates Heat Tolerance in Rice
In this study, phenotypic analysis clearly confirmed that OsOFP3 is a key factor that negatively regulates heat tolerance at the rice seedling stage. In contrast to OsHTAS, a positive regulator of heat tolerance, overexpression of OsOFP3 significantly reduced plant survival under HS, whereas mutant plants showed greater heat tolerance. This “overexpression-sensitive, mutant-tolerant” phenotypic pattern clearly defines the negative regulatory function of OsOFP3. More importantly, we found that the expression of OsOFP3 was suppressed by HS both at the transcriptional and protein levels, indicating that OsOFP3 is a heat-suppressive protein. The expression pattern is consistent with its negative regulatory function: when exposed to high temperature, the inhibition of heat stress tolerance was relieved by down-regulating OsOFP3 expression. This coordinated regulation at the transcriptional and protein levels is a common pattern found in other heat-reactive factors such as HTS1 [36].
4.2. OsOFP3 Exacerbates Heat Damage by Disrupting Chloroplast Stability, ROS Homeostasis, and Stomatal Aperture
Overexpression of OsOFP3 aggravated chlorophyll degradation, enhanced cell membrane damage (increased relative conductivity), and led to a large accumulation of reactive oxygen species (H_2_O_2_ and O_2_^−^), which are typical characteristics of plant oxidative damage under HS. First, a decrease in the chlorophyll content is typically linked to thylakoid damage and impaired photosynthetic function [3]. Overexpression of OsOFP3 may have impaired chloroplast stability and disrupted chlorophyll metabolism under HS. Second, excessive reactive oxygen species (ROS) act as a central hub in various stress responses. This study confirmed that OsOFP3-overexpressing plants accumulated more H_2_O_2_ and O_2_^−^ under HS, which may be due to the inhibition of antioxidant enzyme activity or the promotion of enzymes that produce ROS, as observed in the heat-sensitive rice variety IR64 [37]. Excessive ROS attack biofilms, proteins, and nucleic acids, leading to membrane lipid peroxidation and ion leakage, ultimately accelerating cell death [38]. Third, stomata are critical for regulating transpiration, heat dissipation and water balance. We found that stomatal opening was greater in OsOFP3-overexpressing plants under HS, indicating that stomatal closure was inhibited, thereby destroying the water retention capacity, and may be a direct cause of their reduced heat tolerance. This phenotype is consistent with the functions of several genes that modulate stomatal closure by regulating H_2_O_2_ signaling, thereby altering heat tolerance. For instance, the positive regulators OsHTAS and OsMDHAR4 promote heat tolerance by inducing stomatal closure through facilitating and mediating H_2_O_2_ signaling [39], respectively. In contrast, OsOFP3 functions oppositely.
4.3. The Interaction Between OsOFP3 and OsHTAS May Reveal a Novel Regulatory Pathway for Heat Tolerance
The important finding of this study was the identification of OsHTAS as an interacting protein with OsOFP3 by the yeast two-hybrid assay. OsHTAS is a characterized E3 ligase that positively regulates rice heat tolerance by inducing stomatal closure through the promotion of H_2_O_2_ accumulation and the ABA signaling pathway. The identification of this interaction provides a key clue for unraveling the negative regulatory mechanism of OsOFP3. We hypothesized that OsOFP3 may function as a negative regulatory component or antagonist in the OsHTAS signaling pathway. The specific mechanism may include the following hypothesis: OsHTAS may target OsOFP3 through ubiquitination modification and direct its degradation through the 26S proteasome. The present study found a rapid decrease in OsOFP3 protein levels under HS, a process that is most likely mediated by OsHTAS. The degradation of OsOFP3 is a key step in relieving heat resistance.
5. Conclusions
In this study, we systematically identified OsOFP3 as a novel negative regulator of heat tolerance in rice. The expression of OsOFP3 was inhibited by HS, and its overexpression significantly reduced plant heat tolerance, while the mutant showed enhanced heat tolerance. At the molecular physiological level, OsOFP3 plays a negative regulatory role by aggravating chlorophyll degradation, promoting membrane system damage, enhancing ROS burst, and inhibiting stomatal closure under HS. Further mechanistic studies showed that OsOFP3 interacted with the E3 ligase OsHTAS, suggesting that OsOFP3 may function by modulating OsHTAS-mediated H_2_O_2_ signaling and the stomatal closure pathway.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Quint M. Delker C. Franklin K.A. Wigge P.A. Halliday K.J. van Zanten M. Molecular and genetic control of plant thermomorphogenesis Nat. Plants 201521919010.1038/nplants.2015.19027250752 · doi ↗ · pubmed ↗
- 2Tanaka N. Kawahara Y. Ebana K. Shenton M. Genome wide association study of rice agronomical traits and seed ionome with the NARO Open Rice Collection Plant J.2025122 e 7015210.1111/tpj.7015240245413 · doi ↗ · pubmed ↗
- 3Huve K. Bichele I. Rasulov B. Rasulov B. Niinemets U. When it is too hot for photosynthesis: Heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H 2O 2 formation Plant Cell Environ.20113411312610.1111/j.1365-3040.2010.02229.x 21029116 · doi ↗ · pubmed ↗
- 4Perdomo J. Capó-BauçàS. Carmo-Silva E. Galmés J. Rubisco and rubisco activase play an important role in the biochemical limitations of photosynthesis in rice, wheat, and maize under high temperature and water deficit Front. Plant Sci.201713849010.3389/fpls.2017.00490 PMC 539049028450871 · doi ↗ · pubmed ↗
- 5Scafaro A. Atwell B. Muylaert S. Reusel B.V. Ruiz G.A. Rie J.V. GalléA. A thermotolerant variant of rubisco activase from a wild relative improves growth and seed yield in rice under heat stress Front. Plant Sci.201820166310.3389/fpls.2018.01663 PMC 625628630524456 · doi ↗ · pubmed ↗
- 6Niu Y. Xiang Y. An Overview of biomembrane functions in plant responses to high temperature stress Front. Plant Sci.2018991510.3389/fpls.2018.0091530018629 PMC 6037897 · doi ↗ · pubmed ↗
- 7Suriyasak C. Harano K. Tanamachi K. Matsuo K. Tamada A. Iwaya-Inoue M. Ishibashi Y. Reactive oxygen species induced by heat stress during grain filling of rice (Oryza sativa L.) are involved in occurrence of grain chalkiness J. Plant Physiol.2017216525710.1016/j.jplph.2017.05.01528575747 · doi ↗ · pubmed ↗
- 8Fang Y. Liao K. Du H. Xu Y. Song H. Li X. Xiong L. A stress-responsive NAC transcription factor SNAC 3 confers heat and drought tolerance through modulation of reactive oxygen species in rice J. Exp. Bot.2015666803681710.1093/jxb/erv 38626261267 PMC 4623689 · doi ↗ · pubmed ↗