miRNA-146b Targets TRAF6 and Inhibits LTA-Induced Inflammation of Bovine Mammary Epithelial Cells
Yangyang Song, Peng Liu, Mingxue Li, Xiaolin Li, Huaxue Song, Yutong Zhang, Fanzhi Kong, Changyuan Wang, Binglei Shen

TL;DR
This study shows that miR-146b reduces inflammation in cow mammary cells by targeting TRAF6, suggesting it could help treat mastitis.
Contribution
The study identifies miR-146b as a novel regulator of inflammation in bovine mammary epithelial cells through TRAF6 and NF-κB signaling.
Findings
miR-146b targets TRAF6 and inhibits inflammation in bovine mammary epithelial cells.
Overexpression of miR-146b reduces pro-inflammatory cytokines like TNF-α, IL-1β, and IL-6.
Knockdown of miR-146b abolishes its anti-inflammatory effects.
Abstract
The aim of this study was to investigate the regulatory effect of miR-146b on lipoteichoic acid-induced inflammatory response in dairy cow mammary epithelial cells. Our results indicate that miR-146b targets TRAF6, inhibits inflammation in dairy cow mammary epithelial cells, and promotes their cell viability. miR-146b and its target gene TRAF6 play a certain role in mammary gland development in dairy cows, which is expected to become an effective target for the treatment of mastitis. Mastitis is an inflammatory disease of mammary tissue that impairs milk production and quality, thereby seriously threatening the economic viability of dairy farms. miRNAs, such as miR-146b, are emerging as new candidates for anti-inflammatory therapy, and their activity modulation may provide a basis for controlling inflammation. However, the exact role and underlying mechanisms of miR-146b in bovine…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
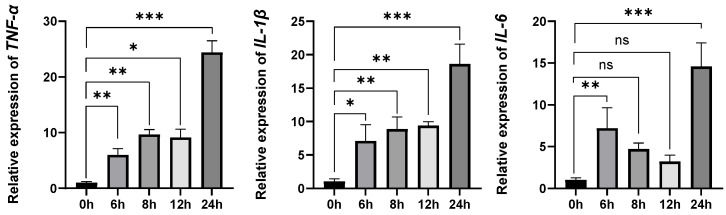 Figure 1
Figure 1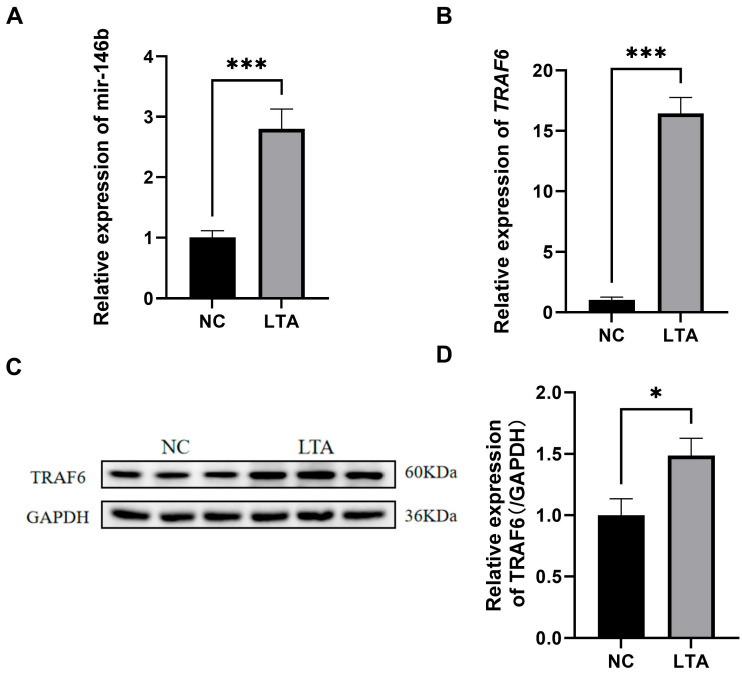 Figure 2
Figure 2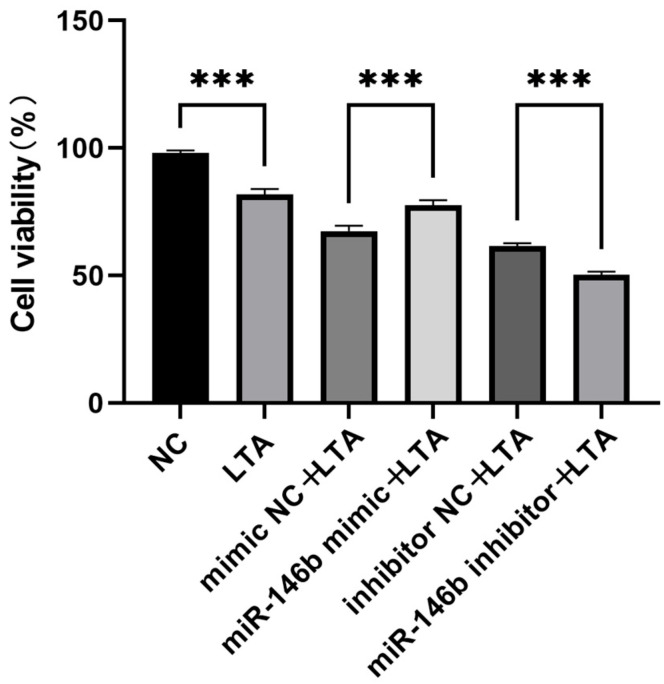 Figure 3
Figure 3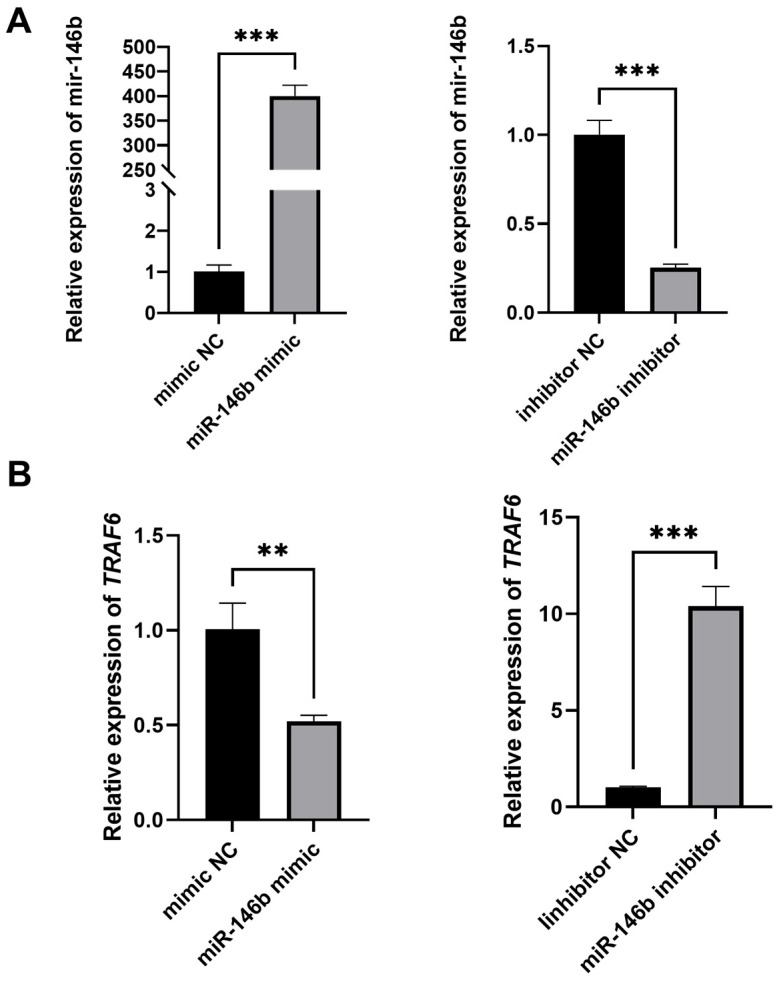 Figure 4
Figure 4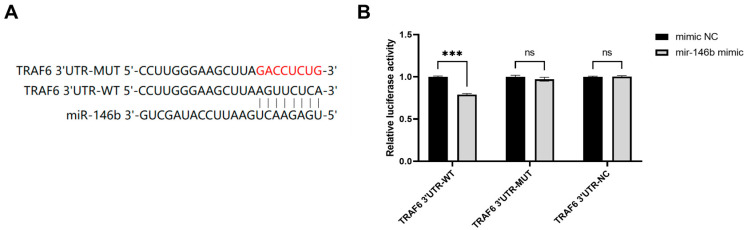 Figure 5
Figure 5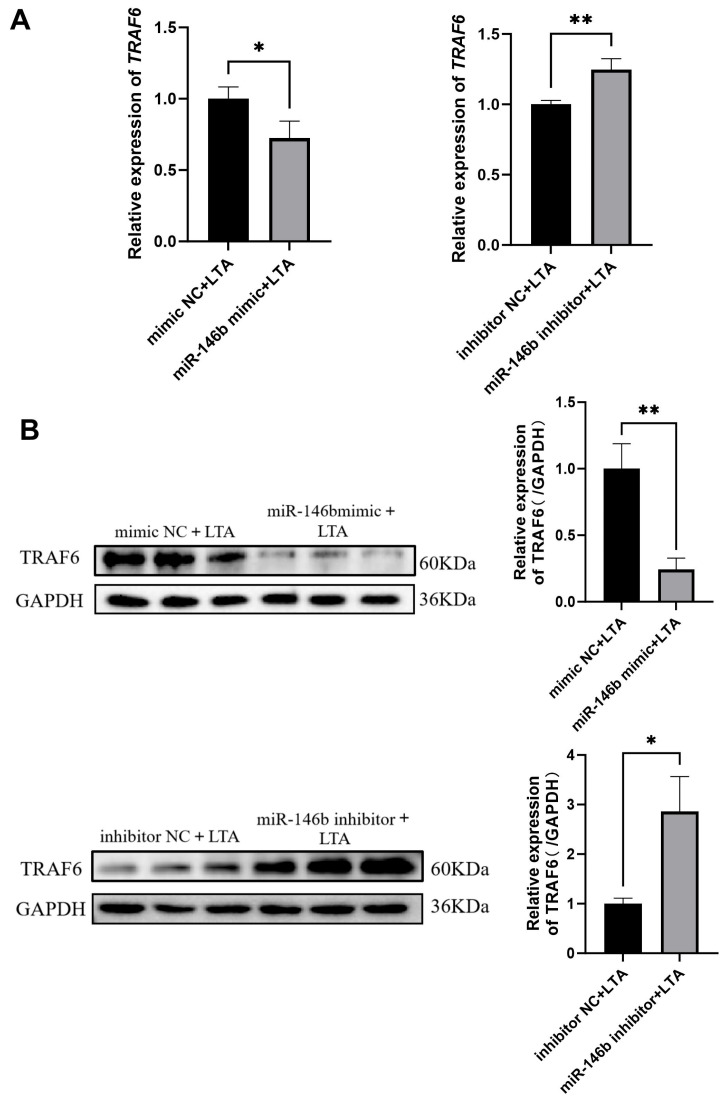 Figure 6
Figure 6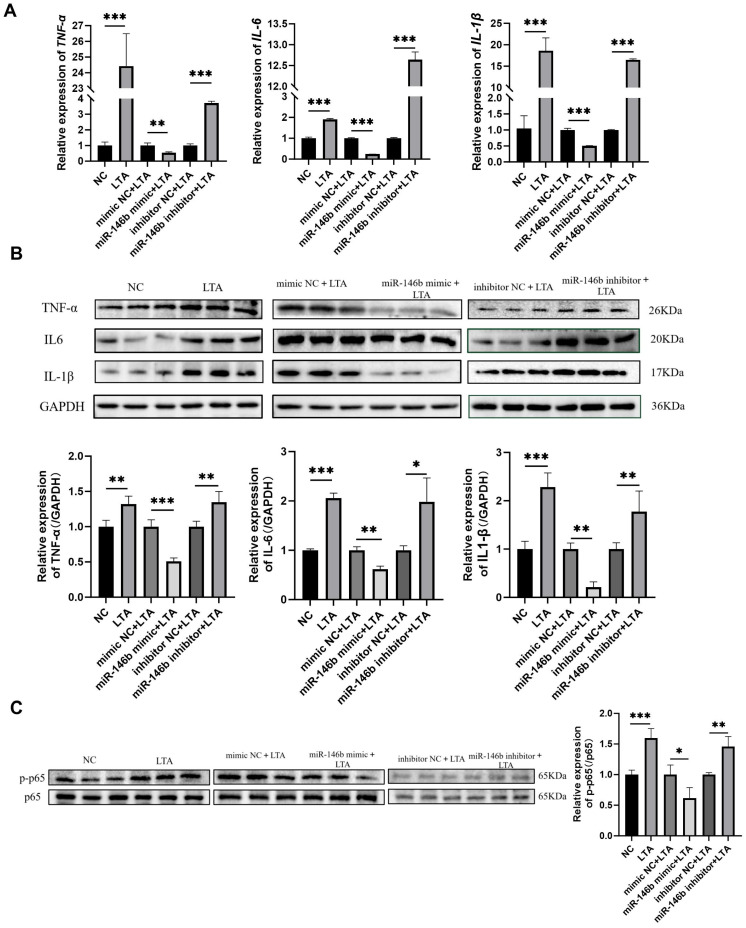 Figure 7
Figure 7- —Natural Science Foundation of Heilongjiang Province
- —Heilongjiang Province Postdoctoral Research Start-up Fund Project
- —Heilongjiang Province Collaborative Innovation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMilk Quality and Mastitis in Dairy Cows · MicroRNA in disease regulation · Reproductive Physiology in Livestock
1. Introduction
Bovine mastitis is an inflammatory disease of the mammary gland induced by physical, chemical, or microbial factors and is considered one of the most economically devastating diseases in the worldwide dairy industry [1,2]. Subclinical mastitis is difficult to diagnose because of the absence of apparent symptoms. Staphylococcus aureus is known to be one of the predominant pathogens causing subclinical mastitis in dairy cattle [3]. One of the major virulence components of this pathogen is Lipoteichoic acid (LTA), which strongly provokes the secretion of various inflammatory factors (TNF-α, IL-6, and IL-1β) from host cells [4]. Thus, the inhibition of subsequent inflammatory signaling pathways provides a potential approach for alleviating mammary glands damage.
Immune balance within the mammary gland is maintained by coordinated interactions among bovine mammary epithelial cells (MAC-T), lymphocytes, macrophages, and neutrophils present in the tissue [5]. Besides their primary role in milk production, the MAC-T cells form an important barrier against pathogenic invasion and play a central role in udder immune defense [6]. These cells participate in the early recognition of pathogens and initiate innate immune reactions, thus organizing further immune responses at both the cellular and molecular levels [7]. A number of studies have shown that MAC-T in culture are able to sense bacteria or bacterial products and respond by upregulating several sets of genes involved in the innate immune response [8]. MAC-T cell is widely regarded as a reliable in vitro model for studies of mammary gland functions because of its expression of milk-specific proteins [9]. While antibiotics remain the main treatment for mastitis, their use is associated with negative consequences such as drug residues and antimicrobial resistance [10]. For these reasons, developing alternative non-antibiotic treatments for bovine mastitis is considered an urgent priority.
MicroRNA is one class of non-coding small RNA molecules, approximately 18–22 nucleotides in length, that play an important role in regulating several pathological processes [11]. The evidence accumulated thus far has demonstrated that miRNAs may be secreted and subsequently taken into specific effector cells to exert functional regulation, mainly by modulating post-transcriptional gene expression [12]. Among these, miR-146b was implicated as a key regulator in immune responses, inflammatory processes, and cytokine signal transduction. Lei Zhang et al. (2020) found that MIR-146b prevented inflammatory damage through the MyD88/NF-κB signaling pathway in pediatric pneumonia [13]. Shuying He et al. (2025) found that miR-146b alleviated inflammatory bowel disease (IBD) by inducing anti-inflammatory effects of IL-10 reprogrammed macrophage polarization [14]. Further studies by Chou Lou et al. (2023) showed that miR-146b-5p inhibited inflammatory responses by targeting TRAF6 [15]. S. Matis et al. (2022) documented that miR-146b-5p targets the IL-23 receptor and its downstream signaling components in the process of regulation of immune responses and cell survival [16]. Ruoxi He et al. (2019) found that overexpression of miR-146b in murine alveolar macrophages reduced LPS-induced TNF-α and IL-1β release [17]. These results suggest that miR-146b plays an important role in immune regulation and inflammation.
Tumor necrosis factor receptor-associated factor 6 (TRAF6), a member of the TRAF family with E3 ubiquitin ligase activity [18], acts as a pivotal node in activating various inflammatory signaling pathways [19,20]. In addition to its established functions in innate and adaptive immune responses, TRAF6 is important in processes such as embryonic development and tissue homeostasis [18]. Until now, the relationship between miR-146b and TRAF6 in the pathogenesis of bovine mastitis has not been reported. Based on these previous studies, we hypothesized that miR-146b targets TRAF6 to exert anti-inflammatory effects in MAC-T cells. In this study, LTA-induced dairy cow mammary epithelial cells were used to explore the effect of miR-146b on inflammation in dairy cow mammary epithelial cells, and to provide a new direction for analyzing the molecular regulatory network and finding molecular therapeutic targets for dairy cow mastitis.
2. Materials and Methods
2.1. Cell Culture and Treatment
MAC-T and HEK293T cell were both obtained from Heilongjiang Bayi Agricultural University. MAC-T and HEK293T cell lines were cultured in Dulbecco’s Modified Eagle Medium (c11995500, Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (164210, Pricella, Wuhan, China). The cells were incubated at 37 °C in a humidified atmosphere of 5% CO_2_. MAC-T cells were cultured to 80% cell density and then stimulated with 20 μg/mL LTA for 0, 6, 8, 12, and 24 h to determine the optimal stimulation time for the LTA model. The dose of 20 μg/mL LTA can stimulate MAC-T cells [21,22].
2.2. Cell Transfection
For transfection experiments, cells were seeded in 6-well plates at a density of 2.0 × 10^5^ cells per well. The cells were transfected with either a miR-146b mimic or inhibitor (Songon, Shanghai, China) using an appropriate mRNA transfection reagent, following the manufacturer’s instructions. The experimental design included the following groups: negative control (NC), LTA-induced inflammation in MAC-T cells (LTA), mimic NC, miR-146b mimic, inhibitor NC, and miR-146b inhibitor.
2.3. Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
Extraction of total RNA was carried out using TRIzol reagent. Reverse transcription of miRNAs was performed with the PrimeScript™ RT reagent Kit with gDNA Eraser (RR047Q, Takara, Beijing, China) in accordance with the manufacturer’s protocol. Amplification was achieved by qPCR on a CFX96 RT-PCR detection system utilizing a TB Green Premix Ex Taq^TM^ (RR420Q, TaKaRa, China). The expression levels of miR-146b, TRAF6, TNF-α, IL-1β, IL-6, U6, and GAPDH (Sangon, Shanghai, China) were quantified. The sequences of the primers employed are presented in Table 1. Analysis of relative expression was conducted using the comparative Ct (2^−ΔΔCt^) method.
2.4. Western Blot
Protein samples (10 μL) were separated by SDS-PAGE. Subsequently, the separated proteins were electrophoresed and transferred to a polyvinylidene difluoride (PVDF) membrane. Blocked with 5% skimmed milk for 2 h. After washing three times with TBST for 15 min, the membranes were incubated with primary antibodies, including anti-TRAF6 antibody (1:1000, A16991, ABclonal, Wuhan, China), anti-TNF-α antibody (1:1000, EPR19147, Abcam, Cambridge, UK), anti-IL-6 antibody (1:1000, EPR21711, Abcam, Waltham, MA, USA), Anti-il-1β antibody (1:1000, EPR16805-15, Abcam, USA), anti-GAPDH antibody (1:50,000, 1E6D9, China) were incubated at 4 °C for 12 h. After incubation, the membranes were washed three times with tris-buffered saline (TBST) containing Tween-20 for 10 min. The membranes were incubated with HRP Conjugated AffiniPure Goat Anti-rabbit IgG (H + L) (1:10,000, BA1054, Boster, Wuhan, China) for 60 min and washed with TBST for another 10 min for three times. Finally, the cells were immersed in enhanced chemiluminescence (ECL) substrates. Protein bands were visualized using a Bio-Rad ChemiDoc imaging system (Bio-Rad, Hercules, CA, USA), and images were quantified using ImageJ software 1.8.0.
2.5. CCK-8 Cells Viability Assay
MAC-T cells were plated in 96-well plates and allowed to adhere for 24 h. Subsequently, the cells were treated with specific stimuli as required by the experimental protocol. After a 48 h incubation period, 10 μL of CCK-8 (MA0218, MeilunBio, Dalian, China) solution was added to each well. Finally, following a 2 h incubation at 37 °C, the absorbance was recorded at 450 nm.
As: absorbance of experimental wells (medium containing cells, CCK-8, drug to be tested).
Ac: absorbance of control wells (medium containing cells, CCK-8, and no drug under test).
Ab: absorbance of blank wells (medium without cells and drug to be tested, CCK-8).
2.6. Dual-Luciferase Reporter Gene Assay
HEK293T cells were seeded into 96-well plates at a density of 1 × 10^4^ cells per well and cultured under standard conditions (37 °C, 5% CO_2_, saturated humidity). The wild-type (WT) and mutant (MUT) 3′UTR sequences of TRAF6 mRNA, along with a negative control (NC) sequence, were cloned into a reporter vector. According to the manufacturer’s protocol, the recombinant vectors were co-transfected with either miR-146b mimic or mimic NC into the cells using Lipofectamine 3000 (BL623, Biosharp, Beijing, China). After 6 h of incubation, the culture medium was replaced with fresh complete medium. At 48 h post-transfection, firefly and Renilla luciferase activities were measured using a dual-luciferase reporter assay kit (RG088, Beyotime, Shanghai, China), following the manufacturer’s instructions. All experiments were performed in triplicate.
2.7. Statistical Analysis of Data
Data are presented as mean ± standard deviation (SD) from at least three independent experiments. The normality of data distribution was assessed with the Shapiro–Wilk test. Statistical analysis was performed using GraphPad Prism 8.0. A two-tailed Student’s t-test was used for two-group comparisons. Multiple group comparisons were conducted using one-way ANOVA followed by Tukey’s post hoc test. Statistical significance was defined as a p-value ≤ 0.05.
3. Results
3.1. Time-Dependent Screening for LTA-Induced MAC-T Inflammatory Model Establishment
The qRT-PCR assay was used to detect the expression of inflammatory factors in MAC-T cells after stimulation with 20 μg/mL LTA at 0 h, 6 h, 8 h, 12 h, and 24 h. The results showed that after LTA stimulation of MAC for 24 h, the expressions of inflammatory factors TNF-α, IL-6, and IL-1β were significantly up-regulated (Figure 1).
3.2. Effect of LTA on the Expression of miR-146b and TRAF6 in MAC-T Cells
To further investigate the role of miR-146b in LTA-induced bovine mastitis, we first examined its expression in MAC-T cells following LTA stimulation. MAC-T cells were treated with LTA for 24 h to establish an inflammatory model. Compared with the control group, LTA challenge significantly up-regulated the expression of miR-146b (Figure 2A). Additionally, TRAF6 mRNA levels were also increased (Figure 2B). Western blot analysis further revealed that TRAF6 protein expression was elevated in LTA-induced MAC-T cells relative to the NC group (Figure 2C).
3.3. Effect of miR-146b on the Viability of LTA-Induced MAC-T Cells
The effect of miR-146b on the viability of LTA-induced MAC-T cells was assessed using the CCK-8 assay. MAC-T cells were seeded in 96-well plates and treated with either an miR-146b mimic or an miR-146b inhibitor. The results showed that compared with the NC group, cell viability was significantly decreased in the LTA-treated group. In contrast, overexpression of miR-146b markedly increased cell viability relative to the mimic NC group, while inhibition of miR-146b led to a significant reduction in cell viability compared to the inhibitor NC group (Figure 3).
3.4. Effect of miR-146b on TRAF6 Expression in HEK293T Cells
To investigate the effect of miR-146b on TRAF6 expression, HEK293T cells were transfected with either an miR-146b mimic or inhibitor. Transfection efficiency was confirmed by the increased level of miR-146b in the mimic group and the decreased level in the inhibitor group (Figure 4A). Compared with the mimic NC group, TRAF6 mRNA levels were significantly reduced in cells overexpressing miR-146b. Conversely, inhibition of miR-146b led to an increase in TRAF6 mRNA levels relative to the inhibitor NC group (Figure 4B). These results indicate a negative correlation between miR-146b and TRAF6, suggesting that TRAF6 is a potential target gene of miR-146b in bovine mammary epithelial cells.
3.5. TRAF6 Expression Is Regulated by miR-146b Through 3′-UTR Binding
TargetScan analysis predicted that the 3′-untranslated region (3′-UTR) of TRAF6 mRNA contains conserved binding sites for miR-146b (Figure 5A). To validate this prediction, a dual-luciferase reporter assay was performed in HEK293T cells. The results showed that co-transfection with the miR-146b mimic and the wild-type TRAF6 (TRAF6-WT) reporter plasmid significantly reduced luciferase activity compared to co-transfection with the mutant TRAF6 (TRAF6-MUT) reporter plasmid or the negative control (Figure 5B). These findings demonstrate that miR-146b can directly bind to the 3′-UTR of TRAF6 and suppress its expression at the post-transcriptional level.
3.6. miR-146b Negatively Regulates TRAF6 mRNA and Protein Expression in LTA-Induced MAC-T Cells
To further validate the regulatory role of miR-146b on TRAF6 in LTA-induced MAC-T cells, we assessed TRAF6 expression following miR-146b overexpression or knockdown using qRT-PCR and Western blot. The results demonstrated that miR-146b negatively regulates both the mRNA and protein levels of TRAF6 in inflammatory MAC-T cells (Figure 6A,B). Consistent with the dual-luciferase reporter assay results, we confirmed that TRAF6 is a direct molecular target of miR-146b in MAC-T cells.
3.7. miR-146b Inhibits the Expression of Inflammatory Factors in LTA-Induced MAC-T Cells
RT-qPCR analysis demonstrated that in the LTA-induced MAC-T inflammatory model, the mRNA expression levels of the inflammatory factors TNF-α, IL-6, and IL-1β were significantly elevated compared to the NC group. Conversely, transfection with an miR-146b mimic reduced the mRNA levels of TNF-α, IL-6, and IL-1β, whereas the miR-146b inhibitor increased their expression (Figure 7A). Furthermore, Western blot analysis revealed a consistent trend at the protein level (Figure 7B). Moreover, overexpression of miR-146b suppressed the expression of phosphorylated p65 (p-p65), while inhibition of miR-146b enhanced p-p65 levels (Figure 7C). These results indicate that miR-146b attenuates the expression and activity of phosphorylated NF-κB (pNF-κB), thereby downregulating the intracellular expression of the inflammatory factors TNF-α, IL-6, and IL-1β.
4. Discussion
Mastitis is an inflammatory process that induces pathological changes in the mammary tissue of dairy cows and leads to the impairment of bovine mammary epithelial cells [23]. While many therapeutic strategies have improved in recent years, the incidence rate of bovine mastitis remains at a high level. Because of the absence of effective treatment, infected cows are usually culled, leading to great economic losses for farmers [23]. Thus, investigating the mechanisms underlying inflammatory injury in bovine mastitis is necessary for developing strategies that will reduce its incidence. Staphylococcus aureus is considered one of the most important pathogens responsible for bovine mastitis. LTA is the major component of the Gram-positive bacterial cell wall, is capable of triggering immune signaling pathways in host cells [3]. This activation leads to the occurrence of oxidative stress, autophagy, and apoptosis in mammary epithelial cells. These properties make LTA an ideal agent for simulating inflammation caused by Gram-positive bacteria [24,25]. Thus, we established an inflammatory model with LTA in MAC-T cells in order to further study the pathogenesis process of mastitis and find possible strategies for preventing and treating mastitis.
Changes in the miRNA expression are one of the early cellular responses to extracellular stimuli. It has been reported that miRNAs may directly target pro-inflammatory cytokines and genes of the crucial signaling pathway, for example, JAK/STAT and MAPK signaling [26,27]. As one important member of the inflammation-associated miRNA family, the function of miR-146b in regulating the TLR/NF-κB signaling pathway has been confirmed in humans, mice, and other species [28]. However, its specific role in bovine mammary epithelial cells remains to be determined. In the current study, miR-146b expression was found to be significantly up-regulated in inflammatory MAC-T cells. These results indicated that miR-146b may participate in the development of bovine mastitis. Further overexpression of miR-146b resulted in a decrease in TRAF6 expression, while inhibition of miR-146b promoted the expression of TRAF6. This suggests that TRAF6 is a putative target gene of miR-146b. Therefore, we further conducted dual-luciferase reporter assays and proved that TRAF6 is a direct molecular target of miR-146b. Additionally, this result was consistent with several studies conducted on other species or cell types. For example, several studies have shown that miR-146b inhibits inflammatory responses through Interleukin 1 Receptor Associated Kinase 1 (IRAK1) and TRAF6 in human and murine immune cells [29,30]. Overexpression of miR-146b notably inhibited the expression of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) in LTA-stimulated MAC-T cells, whereas inhibiting miR-146b showed the opposite effect. These findings suggest that miR-146b exerts its anti-inflammation effect through the downregulation of TRAF6. In addition, we found that miR-146b also affected the phosphorylation of NF-κB p65, and we speculated that, mir-146b inhibited the activation of the NF-κB pathway. CCK-8 assays showed that miR-146b promotes the viability of cells in the LTA-induced inflammatory model. It hinted that miR-146b could not only alleviate inflammation but also protect the injury of mammary epithelial cells mediated by LTA [31,32].
It is important to note that a specific “synchronous upregulation” expression pattern of miR-146b and TRAF6 was displayed in MAC-T cells during inflammation. This can be explained mainly by the temporal control of inflammatory signaling and peculiar features of miRNA-mediated post-transcriptional regulation. Mechanistically, binding of LTA to TLR2 triggers the activation of a downstream MyD88-dependent pathway, which leads to recruitment and activation of TRAF6 [33]. Because TRAF6 is a key adaptor protein in the NF-κB signaling pathway and also becomes a target of transcriptional regulation by NF-κB [34], inflammatory activation thus initiates a positive feedback loop: NF-κB nuclear translocation promotes the transcription of proinflammatory genes, including TRAF6, IL-6, and TNF-α [35], causing a rapid transcriptional induction of TRAF6. Meanwhile, a negative feedback mechanism was elicited to maintain immune homeostasis. In the early to intermediate stage of inflammation, the rate and strength of NF-κB-mediated TRAF6 transcription may overcome the suppressive activity of miR-146b, which was also induced by NF-κB. Consequently, both exhibited an overall up-regulation at the population level. Of note, the induction of miR-146b should not be misinterpreted as proinflammatory. It represents an adaptive regulatory response. Once the level of miR-146b increases to a functional threshold, it suppresses the expression of TRAF6, resulting in restraint of NF-κB signaling, which limits inflammatory amplification and protects mammary epithelial cells from injury.
In the present study, our findings link bovine mastitis to miR-146b; its overexpression in LTA-challenged MAC-T cells curbs inflammatory responses while enhancing cell viability, and this protection is mediated through direct targeting of TRAF6 to suppress the inflammatory cascade.
5. Conclusions
In conclusion, this study demonstrates that miR-146b plays a critical anti-inflammatory role in LTA-stimulated bovine mammary epithelial cells by targeting TRAF6. Our findings show that miR-146b binds with high specificity to the seed sequence in the 3′-untranslated region (3′-UTR) of TRAF6 mRNA, leading to the repression of TRAF6 translation and subsequent reduction in protein levels, with a significant reduction in the expression of its downstream proinflammatory cytokines. Overexpression of miR-146b enhances cell viability while markedly reducing inflammatory responses, highlighting its potential as a therapeutic target for mastitis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nielsen C. Ostergaard S. Emanuelson U. Andersson H. Berglund B. Strandberg E. Economic consequences of mastitis and withdrawal of milk with high somatic cell count in Swedish dairy herds Anim. Int. J. Anim. Biosci.201041758177010.1017/S 175173111000070422445130 · doi ↗ · pubmed ↗
- 2Hogeveen H. Huijps K. Lam T.J. Economic aspects of mastitis: New developments N. Z. Vet. J.201159162310.1080/00480169.2011.54716521328153 · doi ↗ · pubmed ↗
- 3Dong W. Chen Y. Zhang Q. Zhao X. Liu P. He H. Lu T. He Y. Du X. Hu J. Effects of lipoteichoic and arachidonic acids on the immune-regulatory mechanism of bovine mammary epithelial cells using multi-omics analysis Front. Vet. Sci.2022998460710.3389/fvets.2022.98460736090174 PMC 9450935 · doi ↗ · pubmed ↗
- 4Wang J. Qi L. Mei L. Wu Z. Wang H.C. butyricum lipoteichoic acid inhibits the inflammatory response and apoptosis in HT-29 cells induced by S. aureus lipoteichoic acid Int. J. Biol. Macromol.201688818710.1016/j.ijbiomac.2016.03.05427020942 · doi ↗ · pubmed ↗
- 5Bannerman D.D. Paape M.J. Goff J.P. Kimura K. Lippolis J.D. Hope J.C. Innate immune response to intramammary infection with Serratia marcescens and Streptococcus uberis Vet. Res.20043568170010.1051/vetres:200404015535958 · doi ↗ · pubmed ↗
- 6Sun Y. Li L. Li C. Wang G. Xing G. Gene microarray integrated with i TRAQ-based proteomics for the discovery of NLRP 3 in LPS-induced inflammatory response of bovine mammary epithelial cells J. Dairy Res.20198641642410.1017/S 002202991900076131722754 · doi ↗ · pubmed ↗
- 7Aitken S.L. Corl C.M. Sordillo L.M. Immunopathology of mastitis: Insights into disease recognition and resolution J. Mammary Gland Biol. Neoplasia 20111629130410.1007/s 10911-011-9230-421938490 · doi ↗ · pubmed ↗
- 8Griesbeck-Zilch B. Meyer H.H. Kühn C.H. Schwerin M. Wellnitz O. Staphylococcus aureus and Escherichia coli cause deviating expression profiles of cytokines and lactoferrin messenger ribonucleic acid in mammary epithelial cells J. Dairy Sci.2008912215222410.3168/jds.2007-075218487644 · doi ↗ · pubmed ↗