Effects of Enzyme–Microbe Co-Fermented Ganoderma lucidum Spent Substrate on Growth Performance, Apparent Nutrient Digestibility, Organ Indices, and Gut Microbiota in Yellow-Feathered Broilers
Bo Fan, Mengyun Li, Zhifang Shi, Xuanyang Li, Tongshuai Liu, Pu Cheng, Lei Xi

TL;DR
This study shows that adding a fermented Ganoderma lucidum product to broiler chicken feed improves growth and gut health, with 1.5% being the most effective level.
Contribution
The novel contribution is demonstrating that enzyme–microbe co-fermented Ganoderma lucidum spent substrate improves broiler growth and gut microbiota structure.
Findings
EFGLS supplementation improved growth performance and feed efficiency in broilers.
A 1.5% EFGLS diet enhanced thymus development and enriched beneficial gut microbes.
Cecal microbial diversity increased with 3.0% EFGLS, but 1.5% was more consistently beneficial.
Abstract
This study evaluated the use of enzyme–microbe co-fermented Ganoderma lucidum spent substrate (EFGLS) as a feed additive for yellow-feathered broiler chickens. In a six-week feeding trial, broilers were fed diets supplemented with 1.5% or 3.0% EFGLS, partially replacing corn and soybean meal in a conventional basal diet. Both supplementation levels improved growth performance and feed efficiency compared with a control diet, while apparent nutrient utilization also trended upwards. A dietary inclusion level of 1.5% EFGLS was associated with improved thymus development, whereas the 3.0% supplementation level increased cecal microbial diversity. Notably, beneficial dominant microbial populations were more consistently enriched in broilers fed the 1.5% EFGLS supplemented diet. Overall, EFGLS represents a promising functional feed ingredient for broiler production, and under the conditions…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
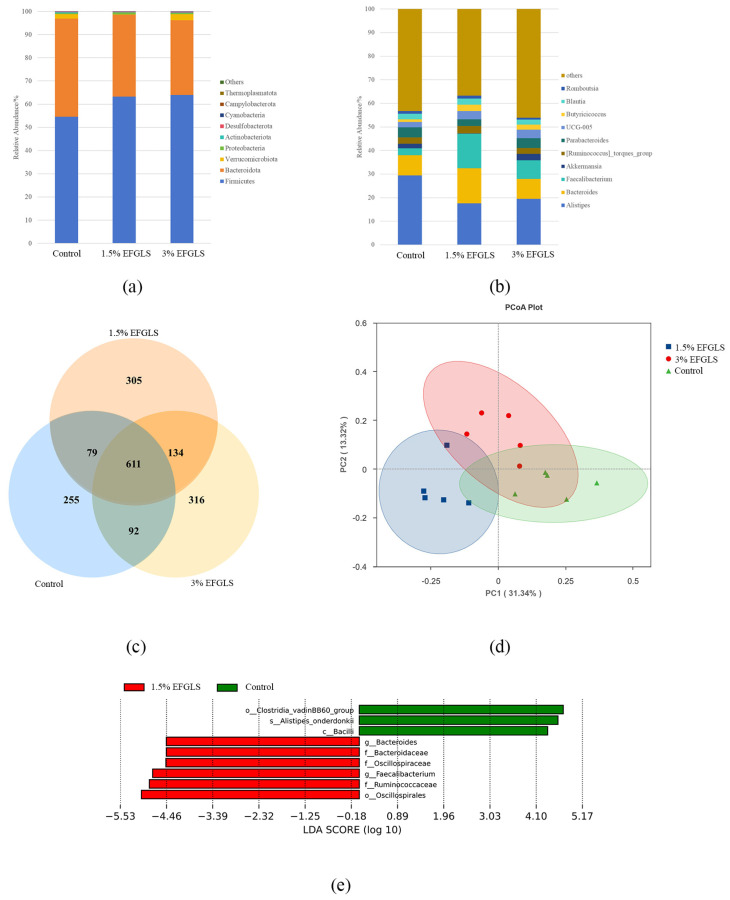 Figure 1
Figure 1- —Subproject of China’s National Key Research and Development Program
- —Key Project of Science and Technology of Henan Province
- —Natural Science Foundation of Henan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology · Fungal Biology and Applications · Insect Utilization and Effects
1. Introduction
With increased demand for high-quality animal protein, poultry meat has gained widespread consumer acceptance because of its low fat content, high protein value, and relatively affordable price. The rapid expansion of the poultry industry has consequently intensified challenges related to nutrient utilization efficiency and environmental sustainability.
Ganoderma lucidum is a traditional Chinese precious medicinal material that is rich in bioactive compounds, including polysaccharides, triterpenoids, phenolic substances, and trace elements, etc., that have substantial pharmacological value [1]. According to the China Edible Fungi Association, the total production of G. lucidum in China reached approximately 150,000 tons in 2023 [2]. Alongside large-scale production, substantial quantities of G. lucidum spent substrate are generated as by-products. This spent substrate, derived from spent mushroom substrates, contains abundant organic matter and mineral elements [3], and has been recognized as a promising feed resource for livestock and poultry [4]. Poultry are characterized by a relatively short digestive tract and the absence of separation between feces and urine, which often results in incomplete digestion of dietary nutrients. As a result, poultry manure contains high levels of organic matter, and the subsequent microbial fermentation of this excreta produces malodorous gases such as ammonia and hydrogen sulfide that create significant environmental problems [5]. Previous studies have shown that dietary inclusion of 3% G. lucidum spent substrate to laying hens reduced ammonia and hydrogen sulfide emissions from manure [6], while supplementation in laying ducks decreased serum lipid levels [7]. In ruminants, the inclusion of fermented G. lucidum mycelia enhances immune function and antioxidant capacity in dairy cows [8].
Microbial fermentation is an effective strategy for the valorization of agricultural by-products and for alleviating pressure on conventional feed resources. Fermentation can degrade fibrous components, reduce anti-nutritional factors, and enrich substrates with bioactive metabolites such as peptides, enzymes, and beneficial microorganisms. These fermentation-derived compounds may influence gut microbial ecology and host nutrient metabolism [9]. Numerous studies have demonstrated that fermented feeds can improve growth performance, modulate intestinal microbiota, reduce nitrogen and phosphorus excretion, and mitigate environmental pollution in poultry production systems [10,11]. The application of fermentation technology to G. lucidum spent substrate can potentially enhance its nutritional value and functional properties. Positive effects of fermented Flammulina velutipes spent substrate on growth and health parameters have already been reported in meat sheep [12]. Anti-nutrient substances are compounds originating in most animal feed materials that are toxic to animals and limit the accessibility of nutrients to the animal body [13]. Combined microbial–enzyme fermentation can efficiently degrade anti-nutritional factors. Based on solid-state fermentation technology, this approach is generally classified into one- and two-step fermentation processes, and it is considered to be a key technique in the development and utilization of unconventional feed resources [14]. Although the underlying mechanisms remain incompletely understood, this approach has attracted considerable research attention because of its favorable fermentation performance.
While fermentation of other agricultural spent substrate has been investigated [15,16], data on fermented G. lucidum spent substrate in broilers, especially yellow-feathered broilers, is limited. In particular, for these broilers, its effects on growth performance, nutrient utilization, immune organ development, and cecal microbiota are not fully known. Therefore, we investigate the effects of supplementing diet with two levels of enzyme–microbe co-fermented G. lucidum spent substrate (EFGLS) on growth performance, nutrient utilization, immune organ development, and cecal microbiota in yellow-feathered broilers.
2. Materials and Methods
2.1. Preparation of Fermented Ganoderma lucidum Spent Substrate
EFGLS was produced by Henan dong-fangjianbiotech Co., Ltd. (Puyang, China). The fermentation procedure and microbial strain selection were performed in accordance with Chen and Heng [17,18], with minor modifications. The compound probiotic preparation was provided by the laboratory of Henan dongfangjianbiotech Co., Ltd., and the compound enzyme preparation (Item number: Xinyangmei 1819E11) was purchased from Smistyle (Neiqiu) Bio-tech Co., Ltd. (Xingtai, China). The collected G. lucidum spent substrate (the spent cultivation substrate remaining after G. lucidum fruiting body harvest, consisting primarily of sawdust, wheat bran, sugarcane bagasse, residual mycelia, and their metabolic products) was oven-dried at 60–70 °C to constant weight then ground to pass through a 40-mesh sieve. The resulting powder was mixed with corn flour, soybean meal, and wheat bran at a ratio of 7:1:1:1 (w/w). After thorough mixing, its moisture content was adjusted to approximately 50%. Subsequently, 2% (w/w) of a compound probiotic preparation and 0.02% (w/w) of a compound enzyme preparation were added, and the mixture was homogenized, packed into breathable fermentation bags, de-aerated, and incubated at 37 °C for 7 days to complete fermentation.
The compound probiotic preparation mainly consisted of Bacillus subtilis (viable count ≥ 2.5 × 10^8^ CFU/g), Saccharomyces cerevisiae (≥6.0 × 10^8^ CFU/g), and Lactobacillus plantarum (≥2.0 × 10^9^ CFU/g). The compound enzyme preparation contained xylanase (≥20,000 U/g), β-mannanase (≥1200 U/g), β-glucanase (≥700 U/g), amylase (≥200 U/g), protease (≥1000 U/g), and cellulase (≥100 U/g). A synergistic interaction exists between microbial inoculants and enzyme preparations [19]. The compound enzyme breaks down the starch, non-starch polysaccharides, cellulose, and structural proteins in the fungus spent substrate, providing energy for the fungus, accelerating the start of fermentation, and providing microbial nitrogen sources. Ultimately, this improves the fermentation quality, making the nutrients easier for animals to absorb and utilize.
2.2. Birds, Diets and Experiment
A total of 450 mixed-sex 22-day-old yellow-feathered broilers of similar body weight were randomly assigned to three dietary treatments. Each treatment included five replicates, with 30 birds per replicate, and each replicate served as an independent experimental unit and was housed individually. Dietary groups consisted of (1) a corn–soybean meal basal diet (Control); (2) basal diet supplemented with 1.5% EFGLS partially replacing corn and soybean meal (1.5% EFGLS); and (3) basal diet supplemented with 3% EFGLS partially replacing corn and soybean meal (3% EFGLS). Inclusion levels of 1.5% and 3.0% EFGLS were selected to represent moderate and relatively high supplementation levels in the diet. Previous studies investigating the application of fermented spent substrates and related by-products in broiler nutrition have mostly investigated inclusion levels of 0.5–3.0% [20,21], with higher supplementation levels reported to negatively affect broiler growth performance [21]. Based on these findings and discussions within our research group, we considered inclusion levels of 1.5% and 3.0% to be appropriate to evaluate potential dose-dependent effects while maintaining practical feed formulation. All experimental diets were formulated to be isonitrogenous and isoenergetic. The basal diet was formulated according to the Feeding Standard of Chicken (NY/T 33–2004); ingredient composition and nutrient levels are detailed in Table 1. The experimental period lasted for six weeks, from 22–63 days of age.
2.3. Animal Management
The feeding trial was performed at an experimental poultry facility. Throughout the experiment, broilers had ad libitum access to feed and water. The ambient temperature was maintained at 27–30 °C during the first week, then reduced by 2–3 °C per week until reaching approximately 20 °C, where it was maintained until the end of the experiment. Natural ventilation was provided, and birds were vaccinated according to standard farm immunization procedures. Manure was removed regularly, and feed intake and mortality were recorded daily.
2.4. Measurements and Sample Collection
2.4.1. Growth Performance
Broilers were weighed on days 22, 42, and 62 of age after feed withdrawal, on a replicate basis. Average daily gain (ADG) was calculated accordingly. Feed intake was recorded on a replicate basis throughout the experimental period to calculate average daily feed intake (ADFI). The feed-to-gain ratio (F/G) was calculated using ADFI and ADG.
2.4.2. Apparent Nutrient Digestibility
From days 60–62, excreta samples were collected continuously for three consecutive days on a replicate basis. Samples were thoroughly mixed, acidified with 10% dilute hydrochloric acid to fix nitrogen, air-dried, ground, and then passed through a 40-mesh sieve before analysis. Crude protein contents in feed and excreta were determined according to GB/T 6432–2018 [22], crude fat following GB/T 6433–2006 [23], sulfur following GB/T 17776-2016 [24], crude ash following GB/T 6438–2007 [25], and total phosphorus following GB/T 6437–2018 [26]. Acid-insoluble ash (AIA) was determined following GB/T 23742–2009 [27] and used as an internal marker to calculate apparent nutrient digestibility. Apparent nutrient digestibility was calculated using Formula (1):
2.4.3. Organ Indices
On day 63, one broiler of body weight close to the replicate mean was selected from each replicate after a 12 h feed withdrawal (with free access to water) and humanely slaughtered. The heart, liver, thymus, spleen, bursa of Fabricius, gizzard, proventriculus, and intestines were excised, and adherent fat was removed before weighing. Organ indices were calculated as the ratio of organ weight to body weight. Organ indices were calculated using Formula (2):
2.4.4. Cecal Microbiota Analysis
After slaughter, cecal contents were aseptically collected into sterile cryogenic tubes, immediately frozen in liquid nitrogen, and stored at −80 °C until analysis. High-throughput sequencing of the 16S rRNA gene was performed by Novogene Co., Ltd. (Beijing, China). Amplification of the 16S rRNA gene targeted the V3–V4 regions and used primers 341F (5′-CCTAYGGGRBGCASCAG-3′) and 806R (5′-GGACTACNNGGGTATCTAAT-3′). Bioinformatic analyses were performed using NovoMagic (Novogene Co., Ltd., Beijing, China).
2.5. Statistical Analysis
All data were analyzed using SPSS software (version 27.0; SPSS Inc., Chicago, IL, USA). One-way analysis of variance (ANOVA) was performed to evaluate differences among treatments, and Duncan’s multiple range tests were used for post hoc comparisons. Differences were considered highly significant at p < 0.01 and significant at p ≤ 0.05.
3. Results
3.1. Effects of EFGLS on Growth Performance
From 43–63 days of age, the final body weights and ADGs of broilers in 1.5% and 3% EFGLS groups were higher than those of the control (p < 0.001), while F/G was markedly reduced (p = 0.001) (Table 2). Over the entire experimental period (22–63 days of age), broilers in 1.5% and 3% EFGLS treatments exhibited greater final body weights and ADGs than controls (p < 0.001), and a lower F/G (p < 0.05).
3.2. Effects of EFGLS on Apparent Nutrient Digestibility
No significant differences in apparent nutrient digestibility were observed among groups (p > 0.05) (Table 3). However, compared with control broilers, those fed diets supplemented with EFGLS had numerically higher apparent digestibility of all measured nutrients.
3.3. Effects of EFGLS on Organ Indices
The thymus index in the 1.5% EFGLS group was higher than that in the 3% EFGLS group (p < 0.05) (Table 4). For indices of other organs there were no significant differences among groups.
3.4. Effects of EFGLS on Cecal Microbiota
3.4.1. Alpha Diversity
Pielou’s evenness and Shannon indices of cecal microbiota in the 1.5% EFGLS treatment were higher than those in the control (p < 0.05) (Table 5). Simpson’s index was lower, and the dominance index was higher in the control than in supplementation groups (p < 0.05). No significant differences were observed among treatments in Chao1 or observed features indices.
3.4.2. Amplicon Sequence Variant (ASV) Clustering Analysis
A total of 611 ASVs were shared among the three groups (Figure 1c). The control group had 1037 ASVs, of which 255 were unique (24.59%). Group 1.5% EFGLS had 1129 ASVs, of which 305 were unique (27.02%), and the 3% EFGLS group had 1153 ASVs, of which 316 were unique (27.41%).
3.4.3. Taxonomic Composition at Phylum and Genus Levels
At the level of phylum (Figure 1a), Firmicutes and Bacteroidota were dominant in the cecal microbiota of all groups (Table 6). The relative abundance of Bacteroidota in the control group was higher than that in the 3% EFGLS group (p < 0.05), while that of Actinobacteriota was higher in the control group than in either supplementation group (p < 0.05). The five most abundant genera across all groups were Alistipes, Bacteroides, Faecalibacterium, Akkermansia, and [Ruminococcus]_torques_group (Figure 1b). The relative abundance of Alistipes was higher in the control group than in either supplementation group (p < 0.05). The 1.5% EFGLS group had higher relative abundances of Faecalibacterium compared with the control and 3% EFGLS groups (p < 0.05).
3.4.4. Beta Diversity Analysis
Principal coordinates 1 (PC1) and PC2 explained 31.34% and 13.32% of the total variation, respectively (Figure 1d). Distinct clusters were observed between control and supplementation groups. Adonis analysis revealed community variation to be 38% between the 1.5% EFGLS and control groups (R^2^ = 0.38, p < 0.01), 25% between the 3% EFGLS and control groups (R^2^ = 0.25, p < 0.01), and 28% between the 1.5% EFGLS and 3% EFGLS groups (R^2^ = 0.28, p > 0.01).
3.4.5. Differential Taxa Analysis
Linear discriminant analysis effect size (LEfSe) analysis with an LDA score threshold of 4 identified distinct microbial biomarkers among treatments (Figure 1e). Differentially abundant taxa were identified in the control and 1.5% EFGLS groups.
4. Discussion
Fermented feed ingredients have been widely reported to enhance animal growth performance. Previous studies have demonstrated that dietary supplementation with fermented mushroom spent substrate improves ADG in pigs [28], sheep [29], and broilers [30], while liquid fermentation products of G. lucidum have also shown growth-promoting effects in broilers [31]. Consistent with these findings, we report the inclusion of EFGLS in the diet of yellow-feathered broilers benefits their growth performance.
The growth-promoting effects of EFGLS were more evident during later growth stages, consistent with Xu et al. [32], who reported fermented feeds to exert stronger effects during latter growth phases of animals. Although ADFI was not significantly affected, the numerically higher feed intake observed in supplementation groups may be attributed to improved diet palatability resulting from fermentation.
The digestibility and utilization of dietary nutrients in poultry are influenced both by the birds’ digestive capacity and the nutritional composition and processing characteristics of the diet. Appropriate fermentation techniques improve feed quality [33], enhance the utilization efficiency of certain feed ingredients [34], reduce the content of anti-nutritional factors in raw materials [35], improve protein and lipid utilization, and promote mineral and amino acid absorption by modulating gut pH and microbial balance [36,37]. However, we found that supplementation with EFGLS did not significantly affect apparent nutrient digestibility. Although this finding differs from most previous reports, supplementation with fermented feed does not always significantly affect nitrogen digestibility in chickens [38]. This may be due to the relatively modest influence of EFGLS on apparent nutrient digestibility, with any beneficial effect primarily reflected in improvements in intestinal health. Moreover, variability in the recovery of AIA and the analytical procedures involved may affect the sensitivity of apparent digestibility measurements. Considered alongside growth-performance results, dietary supplementation with EFGLS improved the productive performance of broilers without exerting adverse effects on nutrient digestibility. Moreover, the partial replacement of conventional feed ingredients may help to reduce feeding costs, indicating its potential applicability in practical poultry production.
Organ indices are commonly used as indicators of immune development and digestive function in animals. Among them, the bursa of Fabricius, thymus, and spleen are the principal immune organs. Because viral infection in chickens can induce atrophy of immune organs [39], the state of their development can, to an extent, reflect immune competence in poultry. The thymus is important in the differentiation and maturation of T lymphocytes, and its development is highly sensitive to nutritional status and gut-derived immunoregulatory signals. Fermented feed ingredients can positively modulate immune organ development in poultry [40]. We report the thymus index in the 1.5% EFGLS group to trend upwards against the control. Although this difference was not statistically significant, it may indicate that a moderate level of fermented substrate supplementation benefits thymic development and immune maturation. In contrast, a significantly lower thymus index occurred in broilers that received the 3% supplementation level, suggesting that excessive inclusion of EFGLS may be of no further benefit, and may even compromise immune organ development. Similar dose-dependent effects have been reported for other fermented feed ingredients [41]. Ganoderma lucidum is rich in bioactive components such as triterpenoids and saponins—compounds with immunomodulatory activities [4]. However, excessive accumulation of these bioactive substances in vivo may exert cytotoxic effects on various cell lines, significantly reducing cell viability [42]. Excessive dietary supplementation with EFGLS may lead to the accumulation of these bioactive compounds in broilers and exert inhibitory effects on the development of immune organs, possibly explaining the reduced thymus index observed in the 3% EFGLS treatment. The underlying mechanisms, however, remain unclear and warrant further investigation.
The heart plays an important role in meeting the increased metabolic demands associated with rapid growth, while the liver serves as the primary organ for nutrient metabolism and detoxification. The small intestine is the primary site of digestion and nutrient absorption in chickens. Fermented feed can improve digestive capacity by enhancing intestinal structure in broilers [43]. However, we report that supplementation with EFGLS did not significantly affect intestine, heart, and liver indices.
Alpha diversity indices are widely used to evaluate microbial richness and evenness, which are closely linked to ecosystem stability and functional redundancy of the intestinal microbiota [44]. We report dietary supplementation with EFGLS to be generally associated with increased microbial richness and a more even cecal microbial community, suggesting a beneficial modulation of gut microbial ecology. Similar improvements in intestinal microbial diversity have been reported in poultry fed fermented diets at appropriate inclusion levels [45]. The observed changes in alpha diversity were further supported by ASV analysis, indicating that EFGLS supplementation reshaped cecal microbial composition. The greater overlap in ASVs between the two EFGLS-supplemented groups compared with the control suggests that EFGLS may exert a directional effect on microbial community assembly and selectively enrich specific microbial members. The specific functional roles of these taxa require further investigation.
Principal coordinates analysis revealed clear separations between the control and EFGLS-supplemented groups. These differences were further supported by Adonis analysis, confirming that EFGLS supplementation altered overall cecal microbial community structure. Such alterations may be attributed to the combined effects of fermentation-derived enzymes, probiotics, and bioactive compounds present in G. lucidum spent substrate (we did not investigate the underlying mechanisms).
Previous studies have shown that dietary inclusion of mushroom spent substrates or herbal by-products can modify dominant microbial phyla and genera in ruminants and monogastric animals [28,46]. At the taxonomic level, Firmicutes and Bacteroidota dominated the cecal microbiota across all treatments, consistent with previous reports [47]. Although the interpretation of the F/B ratio remains controversial, associations between higher F/B ratios and improved growth performance have been reported for poultry [48,49]. We observed a similar pattern, suggesting that EFGLS supplementation may influence the relative abundance of Firmicutes and Bacteroidota, potentially reflecting changes in energy utilization and metabolic efficiency. Combined with species abundances at phylum and genus levels and LEfSe analysis, EFGLS supplementation, particularly at 1.5%, favored enrichment of fiber-degrading and short-chain fatty acid-producing taxa, including Bacteroides, Faecalibacterium, Oscillospiraceae, and Ruminococcaceae. These microbial groups play key roles in polysaccharide degradation, butyrate production, and maintenance of intestinal barrier integrity [50,51,52]. Short-chain fatty acids produced by microbial fermentation play important roles in regulating immune homeostasis and development of immune organs [53,54]. This may partially explain the association between changes in dominant cecal microbiota and the improved thymus development observed in the low-dose EFGLS supplementation group. However, some members of Bacteroides are also involved in tryptophan metabolism, producing indole and skatole [55], which could lead to deterioration of the housing environment and negatively affect broiler welfare and performance. Therefore, further studies are required to identify functionally distinct Bacteroides species or strains in the broiler cecum and to clarify their specific roles. We report such enrichment in the 1.5% EFGLS group but not the 3% EFGLS group, indicating a potential dose-dependent response. Similar findings have been reported in broilers and other monogastric animals, where excessive inclusion of fermented feeds attenuated the proliferation of beneficial taxa or reduced microbial diversity [40,56,57].
5. Conclusions
Dietary supplementation with EFGLS improved growth performance, enhanced thymus development, and modulated cecal microbial composition in yellow-feathered broilers. Based on overall results, a dietary inclusion level of 1.5% EFGLS is recommended under the conditions of this study. As a fermented agricultural by-product, EFGLS offers advantages in terms of resource reutilization and sustainability, demonstrating practical potential as an alternative feed ingredient in commercial broiler production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ran J. Yao Z.B. Xu H. Zhang W. Bai X.Y. Research Progress on Chemical Composition and Pharmacological Effects of Lingzhi (Ganoderma) and Prediction Analysis of Quality Marker Chin. Arch. Tradit. Chin. Med.202543172182+29110.13193/j.issn.1673-7717.2025.05.032 · doi ↗
- 2China Edible Fungi Association Analysis of the National Statistical Survey Results of Edible Fungi Production in China in 2023 Edible Fungi China 20254412012910.13629/j.cnki.53-1054.2025.01.017 · doi ↗
- 3Xie X.H. Liang Y.J. Li Y. Effect of Waste Material Cultured Auricularia auricula on Soda Alkali-Saline Soil Improvement J. Soil Water Conserv.200822130133+15210.13870/j.cnki.stbcxb.2008.05.031 · doi ↗
- 4Fan M.J. Qi J.W. Wang Y. Wang R.F. Song M. Yang Y.P. Liu N. An X.P. Application of Ganoderma lucidum fungus chaff in animal production Feed Res.20214415615910.13557/j.cnki.issn 1002-2813.2021.02.037 · doi ↗
- 5Liu J.Z. Zhao X.Y. Yang G.Q. Research progress on generation and reduction of odor-causing compounds in excreta of poultry Feed Ind.201940586410.13302/j.cnki.fi.2019.07.010 · doi ↗
- 6Jiang Y.X. Li S.M. Fan J.H. Zheng Y.Q. Lu J. Qu L. Wang K.H. Dou X.H. Effects of Dietary Supplementation with Ganoderma lucidum Residue on Apparent Nutrient Digestibility and Emissions of Ammonia and Hydrogen Sulfide in Laying Hens China Poult.201941616310.16372/j.issn.1004-6364.2019.22.013 · doi ↗
- 7Huang X. Yao Y.L. Li C. Zhang X. Jiang G.T. Hu Y. Dai Q.Z. Effects of Dietary Ganoderma lucidum Fungus Chaff Supplemental Level on Laying Performance, Egg Quality and Serum Biochemical Indices of Laying Ducks Chin. J. Anim. Nutr.2020323654366110.3969/j.issn.1006-267x.2020.08.023 · doi ↗
- 8Zheng D. Shi J. Lin Z.X. Effect of the Fermentation Products by Fresh JUNCAO and Ganoderma lucidum on Immune Function and Antioxidant Capacity of Dairy Cows China Dairy Cattle 20177464910.19305/j.cnki.11-3009/s.2017.07.012 · doi ↗