Molecular bases of circadian magnesium rhythms across eukaryotes
Helen K. Feord, Gerben van Ooijen

TL;DR
This paper explores how magnesium levels in cells follow daily rhythms and suggests ancient transport proteins might control these rhythms.
Contribution
The paper proposes that ancestral prokaryotic Mg2+ transport proteins and a feedback model could explain circadian magnesium rhythms in eukaryotes.
Findings
Circadian rhythms in intracellular Mg2+ exist across eukaryotic kingdoms.
Ancestral prokaryotic Mg2+ transport proteins are conserved and may mediate Mg2+ rhythms.
A reciprocal feedback model between Mg2+ usage and transport activity is proposed.
Abstract
Circadian clocks allow for the physiological anticipation of daily environmental changes. A circadian rhythm in intracellular Mg2+ was recently discovered in multiple eukaryotes. Given the pivotal role for Mg2+ in metabolism, Mg2+ rhythms could affect cellular energy expenditure over the daily cycle. To probe the potential mechanisms underlying the generation of cellular Mg2+ rhythms, we present a phylogenetic analysis of Mg2+ transport proteins. Extensive conservation was observed for ancestral prokaryotic proteins, identifying these as candidate proteins mediating Mg2+ rhythms across eukaryotes. We also posit that shared allosteric regulation of Mg2+ transport proteins might underlie Mg2+ rhythms and propose a reciprocal feedback model between the rhythmic usage of Mg2+ and rhythmic transport activity. Circadian rhythms in intracellular [Mg2+] exist across eukaryotic kingdoms.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
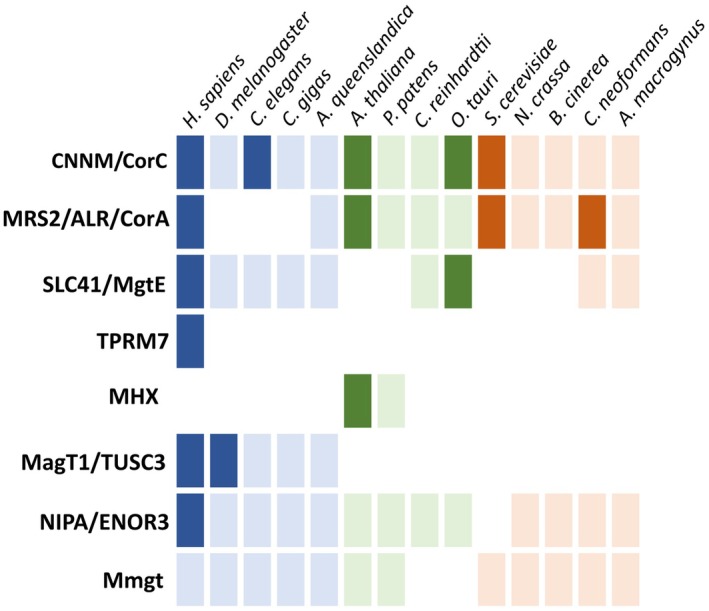 Fig. 1
Fig. 1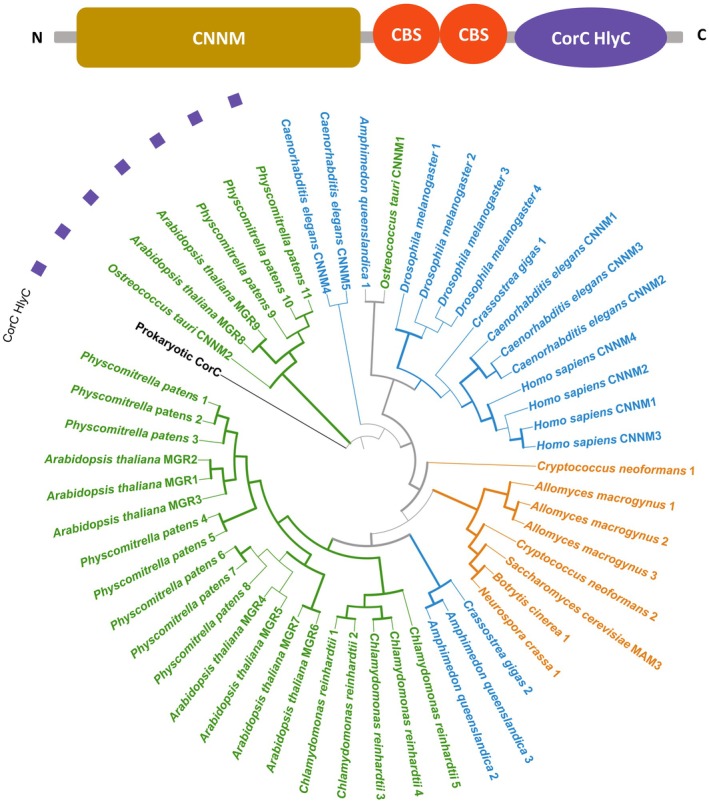 Fig. 2
Fig. 2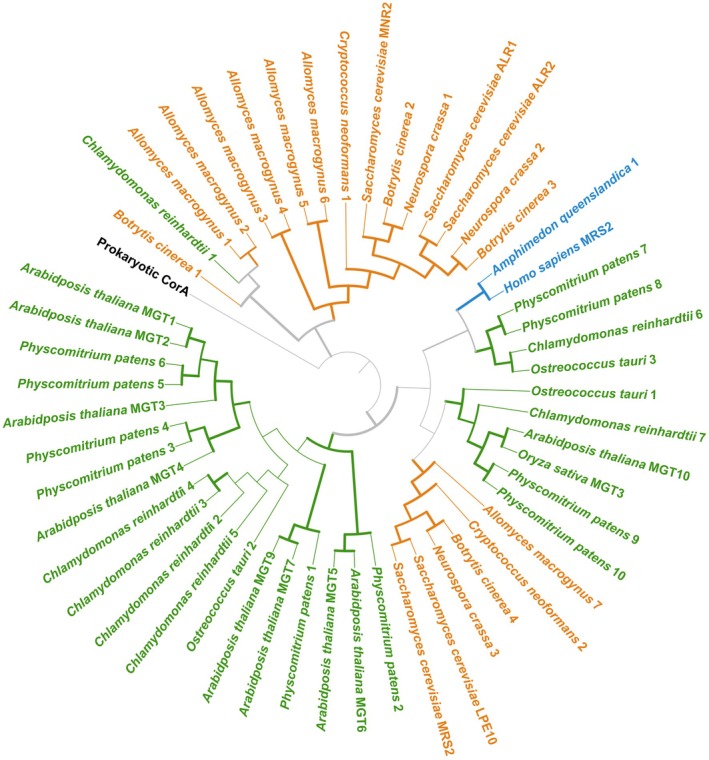 Fig. 3
Fig. 3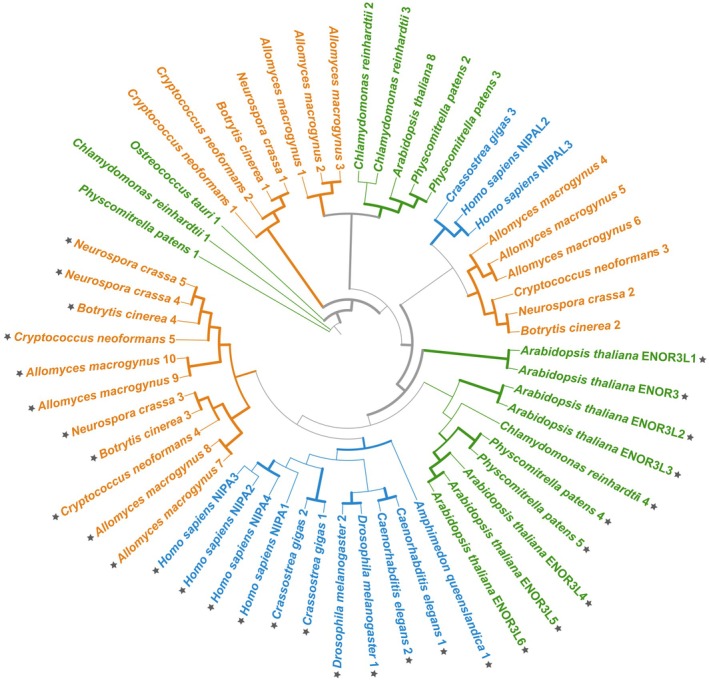 Fig. 4
Fig. 4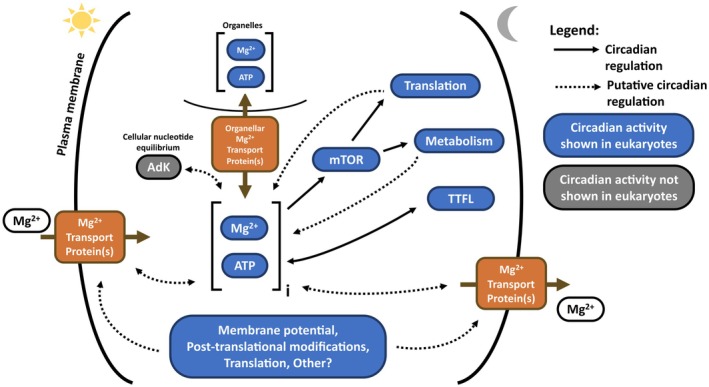 Fig. 5
Fig. 5- —Helmholtz Foundation
- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
- —H2020 European Research Council10.13039/100010663
- —Wellcome Trust10.13039/100010269
- —Leverhulme Trust10.13039/501100000275
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMagnesium in Health and Disease · Circadian rhythm and melatonin · Plant Stress Responses and Tolerance
Abbreviations
AdK adenylate kinase
AI artificial intelligence
BT bootstrap
CBS cystathionine beta synthase
LECA last eukaryotic common ancestor
mTOR mammalian target of rapamycin
TTFL transcriptional/translational feedback loop
Circadian clocks evolved to equip organisms with the capacity to anticipate predictable daily environmental changes resulting from Earth's 24‐h rotation on itself, such as rhythmic light and temperature. Most eukaryotic and some prokaryotic species have a circadian clock [1], and invariably, circadian timekeeping is a property of an individual cell. These cellular oscillators are coupled in multicellular organisms to ensure appropriate synchrony in a tissue or whole organism. For example, in humans the rhythmic environment is perceived by the brain and temporal information is spread across the body by neuronal and hormonal networks. In any organism, the circadian clock involves the rhythmic expression of clock genes, which feedback to regulate their own expression through a complex system of transcriptional‐translational feedback loops (TTFLs). Clock gene expression drives the majority of circadian rhythms controlling organismal and cellular physiology by direct transcriptional regulation [2]. Rhythmic outputs of the clock are often modelled as a sine wave, with critical parameters such as period (time it takes for one oscillation), phase (the timing of a rhythm relative to the environment), and amplitude (relating to the relative difference between the peak and trough levels). Understanding the regulation of these biological rhythms is not only crucial to our understanding of fundamental cell biology: Circadian rhythms are highly important to overall organismal health and fitness, and therefore, circadian biology has immediate applications in fields such as agriculture and medicine.
Before the molecular era of cell biology, it was postulated that the circadian clock could be a ‘feedback system involving ions and ion transport channels’ in which ions and transmembrane ion transporters were considered to be the components of a biochemical circadian clock [3, 4]. This model (known as the ‘Njus‐Sulzman‐Hastings membrane model’) needed to include changes in the activity of transmembrane ion transporters and circadian changes to transmembrane ion gradients, predicting that any manipulation of gradients and transporters must influence the clock [5]. While this field of study was overshadowed by the identification of clock genes [6, 7, 8] and the study of the TTFL system they are part of, there is a convincing body of work that provides evidence for a contribution of rhythmically regulated ion levels to cellular rhythmicity. In 1978, rhythms in K^+^ uptake were reported in duckweed, accompanied by a reverse movement of Mg^2+^ [9]. As expected from a circadian rhythm, these ion rhythms responded to environmental cues such as temperature and light [10]. In a molluscan eye, treatments affecting ionic conductance or gradients influenced the period and phase of clock gene expression [5]. In plants, the transcriptomic response to Mg^2+^ starvation included changes in the expression of plant clock genes [11], providing the first evidence that the availability of Mg^2+^ ions affects the functioning of the circadian TTFL. Mg^2+^ has also previously been highlighted as a cycling nutrient in humans [12], and diurnal rhythms of ionic Mg^2+^ have also been observed in serum [13]. However, as treatments affecting ion rhythms did not stop clock gene expression rhythms, none of these experiments conclusively supported the Njus‐Sulzman‐Hastings membrane model.
In 2016, circadian rhythms in the intracellular concentrations of magnesium and potassium were resolved in eukaryotic cells ranging from the algal minimal model cells of Ostreococcus tauri to mammalian cell types and the fungus Neurospora crassa [14]. Consistent with earlier results in duckweed [10], other biologically relevant ions like calcium did not follow circadian rhythms. The observation in many cell types that free calcium levels in the cytosol and organelles are rhythmic [15, 16] while whole‐cell levels remain identical [10, 14] provides evidence for subcellular circadian ion fluxes. While differential levels of other biologically relevant ions such as iron or sodium were observed over 24 h in Ostreococcus, only potassium and magnesium were rhythmic across eukaryotic kingdoms. These potassium and magnesium rhythms occurred cell‐autonomously and entrain to relevant environmental signals [14]. As treatments that changed intracellular magnesium levels affected the period, phase, and amplitude of clock gene expression and could act as an input pathway, magnesium was posited to be a clock component in itself. Notably, results were near‐identical in algal and mammalian cells, for which the last common ancestor existed more than a billion years ago. Interestingly, ion rhythms persist in human red blood cells, which do not have a nucleus and therefore no circadian gene expression [17]. Circadian rhythms in potassium in these cells were physiologically relevant as they mediated the electrophysiological properties of the cell such as cytoplasmic conductivity and membrane conductance. Recently, ion transport rhythms were shown to be compensatory to allow for circadian rhythms in overall cellular protein synthesis: As protein abundance is rhythmic, the contribution of proteins to osmotic potential is also rhythmic, and ion rhythms in cardiomyocytes maintained osmotic balance in the cytosol over the circadian cycle [18].
Since 2016, several studies on the topic of circadian Mg^2+^ transport have been published in different model organisms. In cyanobacteria, a direct role for Mg^2+^ in the control of the KaiC‐based oscillator was identified [19]. In mice, a magnesium transport system based on PRL2 and CNNM3 was found to affect timekeeping: PRL2‐KO mice exhibited altered circadian behaviours like timing of food intake, activity, and average respiratory quotient, providing evidence that control of [Mg^2+^]i regulates circadian energy and metabolism [20]. Rhythmic transcriptional and translational regulation of TRPM7, a mammalian Mg^2+^ transporter, was observed in mice [21], and TRPM7‐deficient cells had reduced cellular [Mg^2+^]i. In Drosophila, Mg^2+^ cycling was observed in brain cells [22]. In plants, Mg^2+^ deficiency affected not only clock gene expression, but also the pathways feeding environmental information into the clock as well as clock output rhythms [23]. Low extracellular Mg^2+^ lengthened plant circadian rhythms [24], similarly to what is reported in human and algal cells [14]. In rice, the chloroplast Mg^2+^ transporter MGT3 was identified to contribute to diurnal Mg^2+^ rhythms in the chloroplast stroma, directly contributing to circadian rhythms in photosynthetic efficiency and rhythmic carbon fixation rates [25]. In the minimal model cells of Ostreococcus, overexpression of homologues of bacterial magnesium transporters MgtE [26] or CorC [27] altered [Mg^2+^]i and circadian gene expression.
Overall, there is a sizeable body of evidence for a tight interaction between Mg^2+^ rhythms and the full circadian system that integrates environmental information into cellular and organismal rhythmicity. Given the extraordinary importance for magnesium in metabolism (Box 1), understanding the molecular bases and functional consequences of rhythms in its availability is highly relevant. However, while it has been established that these Mg^2+^ rhythms are shared across distant eukaryotic taxa, it is not clear if these rhythms are generated by shared Mg^2+^ transport proteins. The distribution of Mg^2+^ transport protein families across eukaryotes has not been comprehensively reviewed at the time of writing. In addition, the fact that magnesium is notoriously difficult to trace in living cells or tissues [28] has hampered coherent efforts to assess the conservation of protein function across plants, animals and fungi. Using a combination of phylogenetic analyses and a review of published research into magnesium transport, this Perspective article will explore the potential molecular bases of circadian rhythms in magnesium concentrations. Are circadian magnesium rhythms across highly evolutionarily diverse cell types generated by shared Mg^2+^ transport proteins? Or are they mediated by shared regulatory networks acting on divergent transporters? Answering these questions required us to revisit the conservation of Mg^2+^ transport proteins and their regulation in the three eukaryotic kingdoms currently known to exhibit Mg^2+^ rhythms in at least one representative species: animals, plants and fungi [14].
Box 1Metabolic role of rhythmic cellular Mg^2+^ In the context of the circadian control of cell biology, intracellular magnesium levels, or [Mg^2+^]i, are particularly important. Mg^2+^ is a cofactor for ATP and is therefore involved in all metabolic reactions. In addition, more than 600 human enzymes directly require Mg^2+^ for activity [28]. Among other roles, Mg^2+^ is essential for the stabilisation of RNA structures (including tRNA) and for RNA–protein interactions such as between ribosomal RNAs and proteins [95, 96]. These factors imply that most Mg^2+^ is bound to macromolecules such as ribosomes, polynucleotides and ATP [28]. The small amount of freely available [Mg^2+^]i is tightly regulated and functions as a secondary messenger in mammalian cells [89, 97]. Small changes in available free Mg^2+^ can therefore cause a great change in reaction rates of Mg^2+^‐dependent enzymes because of NTPs' sensitivity to and requirement for Mg^2+^ [89]. For example, the master regulator of translation, mTOR, is modulated by Mg^2+^ to contribute to circadian rhythms in the overall cellular translation rates in human U2OS cells [14]. Indeed, a functional consequence of daily fluxes in intracellular Mg^2+^ appears to be the overall control of cellular energy metabolism [14], or in other words, ‘a rhythm in [Mg^2+^]i means a rhythm in effective energy charge’ [98]. Biologically, this could signify that the ‘effective energy charge’ increases throughout the day, as rising magnesium levels from dawn to dusk are observed in all cell types tested to date [14]. It is therefore plausible that Mg^2+^ is a ‘meta‐regulator of metabolic state’ [93]. Because of their potentially large functional relevance, the molecular bases of generating circadian oscillations in [Mg^2+^]i are the main focus of this Perspective paper.
An overview of magnesium transporting proteins in eukaryotes
The identification of Mg^2+^ transport proteins has generally been tied to their functional characterisation in key model cell types, mostly with Homo sapiens for animals, Arabidopsis thaliana for plants and Saccharomyces cerevisiae for fungi. Despite the identity of several Mg^2+^ transporters being known for a long time (see subsections below), the functional characterisation of these proteins is mostly limited to the model cell types that they were initially identified in (Fig. 1; Table S1). Here, we review information regarding the animal, plant and fungal proteins experimentally validated to transport Mg^2+^. Furthermore, using evidence of previous phylogenetic work as well as providing new analyses, we also outline the phylogeny of protein families using the predicted proteomes of model taxa representative of different lineages of animals (H. sapiens, Drosophila melanogaster, Caenorhabditis elegans, Crassostrea gigas, Amphimedon queenslandica), plants (A. thaliana, Physcomitrella patens, Chlamydomonas reinhardtii, O. tauri) and fungi (S. cerevisiae, Neurocrassa crassa, Botrytis cinerea, Cryptococcus neoformans, Allomyces macrogynus). Throughout this review, when referring to ‘plants’, this is inclusive of all Viridiplantae (streptophytes, chlorophytes and prasinophytes). Eukaryotic Mg^2+^ transport proteins can be categorised into two groups: Firstly, the proteins of prokaryotic ancestry and secondly those of nonprokaryotic ancestry that are often lineage‐specific (Fig. 1) [29]. These two groups will be discussed here in that order.
Homologues for different Mg2+ transport protein families across model eukaryotes. Animal proteins are in blue, plant and algal proteins in green, and fungal proteins in orange. Opaque colours indicate proteins experimentally characterised to transport Mg2+, while lighter colours indicate only the presence of a putative homologue. Empty boxes indicate no homologue is present for this taxon. Information about the genome versions and accession numbers used in this paper is available in Table S1. Identification of homologues either by previous studies (TRPM7, MHX, MatT1/TUSC3) or by new phylogenetic analysis (CNNM/CorC, MRS2/ALR/CorA, NIPA) is explained in the following subsections. MgtE homologues were identified with the MgtE domain (PF01769) and Mmgt proteins were identified with the Mmgt domain (PF10270) after sequence annotation with InterProScan (v5.65.97) [99].
Prokaryotic ancestry: CNNM/CorC protein family
The CNNM/CorC family is a family of transporter proteins mediating Mg^2+^ efflux, characterised in prokaryotic (CorB/C proteins, with a characterised crystal structure; [30, 31]) as well as eukaryotic species (CNNM/MGR/MAM3 proteins, Figs 1 and 2). Eukaryotic proteins from this family experimentally validated to transport Mg^2+^ include CNNM1‐4 in mammals [32], CNNM1‐5 in C. elegans [33], MAM3 in yeast [34, 35], CNNM proteins in algae [27] and MGR proteins in higher plants (Figs 1 and 2) [34, 36]. CNNM/CorC proteins have a cyclin M transmembrane N‐terminal domain (CNNM, previously known as domain of unknown function 21, DUF21, Pfam number PF01595) and two cystathionine beta synthase (CBS) domains (Pfam number PF00571, Fig. 2). The CNNM domain is, along with the CBS domains, the characteristic domain of CNNM/CorC proteins, constituting the transmembrane part of the protein (Fig. 2). Some homologues, including prokaryotic proteins, have a CorC HlyC ion transport domain (Pfam PF03471, Fig. 2). The regulation of CNNM/CorC protein activity is known to occur through the CBS domains by Mg‐ATP binding [37] and by direct Mg^2+^ binding to Mg^2+^ binding sites [30]. In the case of mammalian proteins, physical interaction with PRL proteins also contributes to the regulation of CNNM protein activity [38]. A recent study using artificial intelligence (AI) to determine the transport mechanics of CNNM/CorC proteins identified that human CNNM proteins and prokaryotic CorC retained conserved hydrophilic residues involved in transport regulation, but also provided evidence that CNNM/CorC proteins are Mg^2+^/Na^+^ antiporters [39].
Protein topography and phylogenetic analysis of CNNM/CorC proteins across animals, plants and fungi. Protein topography shows the CNNM and CBS domains existing on all homologues, and the CorC HlyC domain which exists only for some homologues. Evolutionary relationship between CNNM/CorC proteins is based on a consensus tree inferred from 1000 bootstrap trees. Protein sequences with a DUF21 domain (PF01595) were selected for analysis after sequence annotation with InterProScan (v5.65.97) [99]. Sequences were aligned with MUSCLE (v3.8.31) [100] and trimmed with TrimAL (v1.5) [101] using the ‐automated1 parameter. Phylogenetic analysis was undertaken with IQ‐TREE (v.2.4.0) [102], with the following parameters: ‐m MFP ‐B 1000 ‐alrt 1000 ‐nt AUTO. The consensus tree was visualised with iTOL (v7) [103]. Bootstrap values above 70 are indicated with a thicker branch. Plant proteins are in green, animal proteins in blue and fungal proteins in orange. Proteins with a CorC HlyC domain are marked with a purple square. The prokaryotic CorC protein sequence (in black) is the Escherichia coli protein (UniProt accession: P0AE78).
CNNM/CorC proteins are ubiquitous across eukaryotic phylogenetic clades (Figs 1 and 2) [37], with significant lineage‐specific diversification for animals, fungi and plants (Fig. 2). We identified a large cluster of multiple A. thaliana, P. patens and C. reinhardtii proteins (Fig. 1), confirming a previously identified [34] expansion of this protein family in a common ancestor of streptophytes and chlorophytes. A. thaliana MGR1‐3 proteins are localised at the vacuole membrane and MGR4‐7 at the plasma membrane [34, 40]. This diversification likely happened during the evolution of land plants, as there is clear differential clustering of MGR1‐3 and MGR4‐7 with P. patens proteins, but not C. reinhardtii proteins. In contrast, a homologue from the prasinophyte model O. tauri is positioned close to the animal proteins, indicating a significant evolutionary distance between this protein and the other plant proteins. A. thaliana MGR8‐9 are chloroplast localised [36], and cluster separately from AtMGR1‐7 (Fig. 2), alongside homologues of O. tauri and P. patens (but not C. reinhardtii) as well as the prokaryotic protein [27, 36]. Each of the proteins in this cluster has a CorC HlyC domain (with purple annotations on Fig. 2). It is possible that a second ancestral prokaryotic protein was acquired through the endosymbiosis which gave rise to the chloroplast and the genes encoding these CorC HlyC domain‐containing proteins.
Prokaryotic ancestry: The MRS2/ALR/CorA protein family
The CorA/MRS2/ALR family is a second family with both a prokaryote ancestor and characterised homologues in the three eukaryotic lineages highlighted here (Fig. 1). Mediating Mg^2+^ influx, members of this family of Mg^2+^ transport proteins have two transmembrane domains, and a GMN motif. In eukaryotes, homologues have been characterised as Mg^2+^ transport proteins in mammals (the mitochondrial MRS2 protein), yeast (the five MRS2, LPE10 and ALR proteins) and Arabidopsis (the MRS2/MGT proteins) [41]. The conservation of function between proteins of this family has previously been highlighted by their ability to functionally substitute for each other between species: For example, the human MRS2 protein can functionally substitute for the yeast MRS2 protein [42]. Crystal structures of the prokaryotic CorA and the mammalian MRS2 proteins have identified that Mg^2+^ transport activity is regulated by Mg^2+^ binding for both eukaryotic and prokaryotic proteins [43, 44, 45] and also Ca^2+^ [46]. Changes to membrane potential can also contribute to MRS2 transport activity [47].
MRS2 homologues are known to exist in animal, plant and fungal taxa [41, 48]. However, in animals, while conserved across vertebrates (one mitochondrial‐located copy per species), MRS2 is absent from most other animal lineages, except at its base with sponges (Figs 1 and 3) [41]. In contrast, plant species have a higher copy number of CorA/MRS2/ALR proteins (Fig. 3). The phylogeny of plant CorA/MRS2/ALR proteins (known as MGT proteins) with additional angiosperm species, including rice, has been addressed in other studies [49, 50, 51, 52, 53]. The Arabidopsis proteins have been well described: MGT1‐3 localise at the tonoplast, MGT6 and 9 localise to the plasma membrane, MGT4 and 7 localise to the ER membrane and MGT5 at the mitochondrial membrane [53, 54]. The Arabidopsis MGT10 protein, localised at the chloroplast [25, 55], shows clear clustering with homologues from the three other plant proteomes, possibly suggesting that all these are chloroplast localised. The rice homologue of MGT10, OsMGT3, is also a chloroplast protein and is involved in diel Mg^2+^ rhythms in the chloroplast (Fig. 3) [25]. Fungal MRS2 proteins exist in two distinct groups (Fig. 3), with homologues of all proteomes queried identified in both. One group, which clusters with the animal proteins, includes the yeast LPE10 and MRS2 proteins. The human MRS2 protein and the yeast MRS2 and LPE10 are both known to localise at the mitochondrial membrane [56] and are likely to be a product of shared opisthokont evolution. The other three characterised yeast proteins (MNR2, ALR1 and ALR2) are evolutionarily very distinct (Fig. 3). ALR1 and ALR2 both localise at the plasma membrane, while MNR2 localises at the vacuole membrane [57]. However, this diversification appears to be basidiomycete‐specific (Fig. 3), limiting in silico predictions of nonbasidiomycete homologues of MNR2/ALR1/ALR2.
Phylogenetic analysis of MRS2/ALR/CorA proteins across animals, plants and fungi. Protein topography shows the GMN motif existing on all protein homologues. The evolutionary relationship between MRS2/ALR/CorA proteins is based on a consensus tree inferred from 1000 bootstrap trees. Homologues were identified by BLASTp searches (v2.14.1, with ‐evalue 1e‐5 parameters) [104], using all previously identified Arabidopsis (MGT 1‐10), yeast (MRS2, LPE10, ALR1‐2, MNR2) and human (MRS2) proteins. Only sequences with a GMN motif were retained for phylogenetic analysis. Sequence alignment, alignment trimming, phylogenetic analysis and tree building were undertaken as in Fig. 2. Bootstrap values above 70 are indicated with a thicker branch. Plant proteins are in green, animal proteins in blue and fungal proteins in orange. This tree also includes the rice (Oryza sativa) OsMGT3 chloroplast‐localised protein (UniPort accession: Q9AUK4). The prokaryotic CorA protein sequence (in black) is the Escherichia coli CorA protein (UniProt accession: P0ABI4).
Prokaryotic ancestry: SLC41/MgtE protein family
The MgtE protein family is characterised by a shared MgtE domain (PF01769), with most homologues having one MgtE domain, except animal proteins (called SLC41 proteins) and some prokaryotic proteins that have two [26]. Prokaryotic proteins are channels with two CBS domains, similar to CNNM/CorC proteins [58]. MgtE proteins are not represented in the proteomes of yeast or higher plants, but homologues are found in algae and some fungal species (Fig. 1) [26]. The O. tauri homologue has been experimentally shown to contribute to timekeeping and Mg^2+^ homeostasis [26]. The regulation of the activity of these proteins is not conserved between prokaryote and animal homologues: The bacterial proteins are regulated by Mg‐ATP interaction with the CBS domains and direct Mg^2+^ binding to the cytoplasmic domain mediating Mg^2+^ import into the cell [58, 59], while the human proteins (SLC41A1–3) mediate Na^+^‐dependent Mg^2+^ efflux [60]. The method of regulation of the algal protein, which does not have CBS domains [26], is unknown. Further structural studies are required to fully establish the regulatory mechanism in the eukaryotic proteins [58].
Proteins of nonprokaryotic ancestry
Mg^2+^ transport in eukaryotic cells is also mediated by proteins that do not have prokaryotic ancestry, and their characterisation is generally limited to the taxa they were originally identified in, for example with plant (MHX proteins) or animal cell types (TRPM6‐7, NIPA, MmgT and MagT1). However, while some nonprokaryotic protein families appear to mostly be limited to the taxonomic group they were identified in, for example MHX [61] and TRPM6‐7 (Fig. 1), others may actually display a level of functional conservation across eukaryotic clades, such as NIPA (nonimprinted in Prader‐Willi/Angelman syndrome; Pfam domain PF05653).
NIPA proteins have been experimentally characterised as Mg^2+^ transporters in mammals [62]. However, we do not know how NIPA is regulated, or indeed which protein motif/domains are responsible for their Mg^2+^‐transport activity. Due to their high protein similarity (including the conservation of the nine TM domains), the ENOR3 family of Arabidopsis proteins has been suggested to be NIPA homologues in plants [63]. The list of hits also included the two human NIPA‐like proteins: NIPAL2 and NIPAL3, which are annotated as being similar to the NIPA proteins, without any proven functional similarity (Fig. 4). This clade also includes proteins from all Opisthokonta proteomes (except S. cerevisiae), as well as from A. thaliana, P. patens and C. reinhardtii (these proteins are marked with grey stars in Fig. 4). If plant ENOR3 proteins are functional Mg^2+^ transport proteins, like the human NIPA proteins, it is likely that all the proteins in this clade are involved in Mg^2+^ transport. Protein characterisation beyond the animal proteins in this clade would be essential to confirm this. The exact segments that are responsible for Mg^2+^ transport (like the MgtE domain in MgtE proteins or the GMN motif in MRS2 proteins) are unknown for NIPA proteins, and therefore, we cannot draw conclusions on the potential functional Mg^2+^ transport activity of any of the clades identified in Fig. 4: no experimental characterisation of nonmammalian proteins nor any further insight into NIPA regulation currently exists, which limits our ability to robustly identify putative nonanimal homologues as bona fide Mg^2+^ transporters.
Protein topography and phylogenetic analysis of proteins with a NIPA domain across animals, plants and fungi. Evolutionary relationships between NIPA proteins are based on a consensus tree inferred from 1000 bootstrap trees. Protein sequences with a NIPA domain (PF05653) were selected for analysis after sequence annotation with InterProScan (v5.65.97, [99]). Sequence alignment, alignment trimming, phylogenetic analysis and tree building were undertaken as in Fig. 2. Bootstrap values above 70 are indicated with a thicker branch. Plant proteins are in green, animal proteins in blue and fungal proteins in orange. Putative Mg2+ transport proteins are marked with a dark grey star.
Human TRPM6‐7 proteins are 2 of 8 members of the vertebrate TRPM (transient receptor potential melastatins) cation channel family. TRPM6‐7 have been identified as a channel involved in the transport of not only Mg^2+^, but also Zn^2+^ and Ca^2+^. These two proteins also share the unique characteristic of having a kinase domain and are therefore the only known examples of bifunctional kinase‐coupled channels in vertebrates [64]. The TRPM protein family itself is 1 of 8 families belonging to the TRP superfamily [65]. Based on a phylogenetic study in Metazoa [65], TRPM diversification likely occurred independently in different metazoan lineages. This signifies that the TRPM1‐8 (including TRPM6/7) diversification in vertebrates was specific to that group, clarifying that the expansion of TRPM proteins in vertebrates is responsible for the generation of TRPM proteins that mediate Mg^2+^ transport (TRPM6/7). While TRPM proteins exist in certain nonvertebrate species [65], it is unlikely that other TRPM proteins also transport Mg^2+^, unless this function arose separately in other lineages.
MagT1 and TUSC3 are known mammalian Mg^2+^ transport homologues, sharing 66% amino acid similarity [66]. MagT1 is also known to be a subunit of oligosaccharyltransferase and is known to be localised at the ER [67, 68]. This protein is therefore a biological link between [Mg^2+^]i and cellular glycosylation. This protein has also been characterised in Drosophila melanogaster [69]. MagT1 proteins are conserved in metazoans, share homology with Saccharomyces cerevisiae OST3/OST6 and contribute directly to N‐linked glycosylation [68].
The mammalian protein Mmgt is located at the Golgi apparatus and post‐Golgi vesicles and has two TM domains. Only one study currently exists which experimentally characterised Mg^2+^ transporter activity [70]. The exact method of regulation and transport of these proteins has not been elucidated, but they have been identified as protein channels [71]. The proteins are very short in length (approximately 130 amino acids) and it is unlikely that they have the ability to transport Mg^2+^ without associated proteins [72]. The characteristic MmgT domain (PF10270) does exist in species representative of all three lineages, suggesting possible conservation across eukaryotic lineages (Fig. 1), and however, it is not possible to currently assess the likely extent of Mg^2+^ transport activity of uncharacterised homologues in mammals or other taxa.
Finally, in plants, the MHX protein family consists of Mg^2+^/H^+^ exchangers that mediate vacuolar Mg^2+^ influx [73]. The proteins are ubiquitous in land plants, but absent in other members of the green lineage [61]. It has been suggested that these Mg^2+^/H^+^ transporters evolved from N^+^/Ca^2+^ exchangers after the split from chlorophytes, making it a streptophyte‐specific protein family. It is not currently known if chlorophyte algae evolved other specific Mg^2+^ transporting proteins. Similarly, no fungi‐specific proteins have been characterised, highlighting an important line of research: do fungi or chlorophyte algae only use ancestral prokaryotic proteins (and possibly NIPA) to transport Mg^2+^ (Fig. 1), or have they also evolved their own systems currently unexplored?
Regulation of circadian rhythms in magnesium
In the previous sections, we reviewed the conservation of proteins known to transport Mg^2+^ across Eukarya. We found that the two Mg^2+^ transport protein families with prokaryotic members, CorC/CNNM and CorA/MRS2/ALR, were the most highly conserved families. The MgtE/SLC41 family is less well conserved than the other two and is missing from large sections of Eukarya, including most fungi and land plants. However, it has characterised members in prokaryotes [74], green algae [26] and animals [60, 75], displaying a level of conservation of Mg^2+^ transport function despite gene loss in various lineages and differing modes of regulation. The high level of conservation of sequence and function of the families with prokaryotic ancestry indicates their evolutionary importance for Mg^2+^ transport; an importance which could plausibly extend to rhythmic Mg^2+^ transport. Eukaryotic proteins of all three of the above protein families are likely to have been present in LECA (last eukaryotic common ancestor). As [Mg^2+^]i rhythms are believed to have a common origin [14], they are likely to either have arisen in LECA or were inherited from a (prokaryotic) ancestor. This highlights the relevance of functionally characterising proteins from the CorC/CNNM, CorA/MRS2/ALR, and MgtE/SLC41 families, as modelling the original ‘LECA system’ provides the framework to understand the basic regulation of eukaryotic [Mg^2+^]i rhythms. Furthermore, compared to CorC/CNNM, CorA/MRS2/ALR and MgtE/SLC41, the protein families without prokaryotic ancestry often have biological roles that are not limited to Mg^2+^ transport, such as glycosylation for MagT1 and kinase activity for TRMP6/7, which are functions that are not directly relevant to circadian Mg^2+^ transport. Based on our analyses, the minimal model cells of Ostreococcus tauri would provide a suitable cell type to study the ‘LECA system’, as only homologues of magnesium transport proteins (except for a potential NIPA homologue) from prokaryotic descent appear to be present. Early indications from this model cell are that overexpression of the MgtE homologue [26] or the CNNM homologues [27] affects the properties of clock gene expression, presumably through the observed differences in cellular Mg^2+^ levels. Genetic deletion of the coding sequences in future studies would be required to fully elucidate the contribution of individual proteins to the rhythmicity of Mg^2+^ levels.
However, there is evidence that taxon‐specific proteins can also contribute to [Mg^2+^]i rhythms, for example the TRPM7 protein in mammalian cells [21]. This suggests that proteins do not need to be highly conserved across eukaryotes to contribute to circadian [Mg^2+^]i fluxes. We can hypothesise that [Mg^2+^]i rhythms in different taxa are mediated by different Mg^2+^ transport systems, adapted to account for differences in cellular complexity (such as a different set of organelles) and environmental constraints (e.g., the lack of Na^+^‐dependent transport in land plants). Following this hypothesis, it is possible that divergent proteins respond to shared cellular signals, resulting in the same phenotype. It is therefore important to not only identify the proteins responsible for generating Mg^2+^ rhythms, but also, crucially, to understand their rhythmic regulation. It is well established that the genes coding for transmembrane transporters are generally highly rhythmically regulated by the transcriptional clock system, as can be assessed using databases such as CircaDB (mammals) [76] or AlgaeFun (Ostreococcus) [77]. Promoter analyses could be carried out to find potential clock proteins responsible for this transcriptional regulation. However, in Ostreococcus, [Mg^2+^]i rhythms persist in the absence of transcription [14], indicating that the rhythmic regulation of Mg^2+^ transport can be sustained without rhythmic mRNA. Furthermore, no clear evidence for rhythmic abundance of Mg^2+^ transport proteins was identified in the Ostreococcus proteome [78]. These observations imply that post‐translational regulation of Mg^2+^ transport proteins is likely to drive [Mg^2+^]i rhythms in this alga (Fig. 5). As overwhelming evidence exists that the activity of magnesium transporters is post‐translationally or allosterically regulated, we posit that circadian rhythms in magnesium across eukaryotes are likely generated through rhythmic activity rather than rhythmic abundance of the proteins that make up the transport system. For example, changes to membrane potential can contribute to transport activity, as evidenced, for example, for the mitochondrial MRS2 channel [47]. Therefore, circadian control of membrane potential could contribute to circadian rhythms of Mg^2+^ transport (Fig. 5). The clear circadian rhythms observed for intracellular K^+^ levels across eukaryotes [14, 17, 79, 80] are likely to provide a mechanism for circadian regulation of membrane potential, given that K^+^ is the most abundant cation in any living cell [81]. Indeed, circadian rhythms in potassium and magnesium levels generally peak at the same phase [14].
Possible regulation of eukaryotic [Mg2+]i rhythms and feedback with cellular pathways. [Mg2+]i rhythms generated by rhythmic transmembrane transport could be regulated by a variety of cellular components (such as membrane potential, post‐translational modifications, translation and by Mg2+ and ATP), with an extensive degree of potential feedback. We posit that feedback exists between cytosolic and organellar [Mg2+]/[Mg‐ATP], as well as between [Mg2+]/[Mg‐ATP] and cellular metabolism, translation and clock gene rhythmicity, at least part of which is mediated through mTOR. Cellular nucleotide equilibrium likely plays a vital role in mediating [Mg2+] rhythms by modulating the pool of ATP for Mg2+ binding, possibly via AdK. AdK, adenylate kinase; mTOR, mammalian target of rapamycin; TTFL, transcriptional/translational feedback loop.
However, a key and conserved allosteric regulator of Mg^2+^ transport proteins is Mg^2+^ itself; both free Mg^2+^ and Mg‐ATP (Fig. 5). For most Mg^2+^ transport proteins with a resolved crystal structure, Mg^2+^ and ATP‐binding sites are evident: MgtE [74], CorA/MRS2 [43, 44, 45], and CorC/CNNM [30, 31, 39], including in key conserved domains such as the cyclin domain in CorB/C/CNNMs [31]. The same is true for mammalian TRPM7 [82]. In all these studies, it was shown that Mg^2+^ or Mg‐ATP binding was instrumental in activating or inhibiting the Mg^2+^ transport activity of the proteins. Noncircadian work has indicated that free cytosolic Mg^2+^ is tightly controlled [72]. Therefore, the transfer of Mg^2+^ between the cytosol and organelles and/or between bound forms (such as proteins or ATP) and free forms, likely signals for the uptake or removal of Mg^2+^ ions from the cell (or possibly into the vacuole in the case of plant cells). The original report of circadian Mg^2+^ rhythms [14] was limited to the whole‐cell level, and subcellular dynamics as well as the fraction of bound versus free Mg^2+^ remained unknown. However, observations that Mg^2+^ rhythms affect overall cellular translation in algae and mammals have suggested that total cytosolic Mg^2+^ levels are in fact rhythmic [14], and it is clear that rhythmic transport over the plasma membrane must exist to account for whole‐cell rhythms (Fig. 5). Therefore, it is likely that rhythmic plasma membrane Mg^2+^ transport will act according to the Mg^2+^ status of the cytosol, which is a direct result of rhythmic Mg^2+^ use by organelles and cytosolic binding sites such as proteins and ATP. Therefore, it is possible that the rhythmic activity of Mg^2+^ transport proteins is modulated by Mg^2+^ availability through conserved Mg^2+^ binding sites in important protein domains such as the cyclin domain in CorC/CNNM. However, to investigate this cellular process further we need to ask the question: how do rhythmic changes in the abundance and usage of cytosolic free Mg^2+^ and Mg‐ATP occur?
Investigating the potential feedback mechanism between rhythmic Mg^2+^ use and rhythmic Mg^2+^ transport involves identifying important cellular pathways that rhythmically use Mg^2+^. A first determinant of cytosolic Mg^2+^ availability is the shuttling of Mg^2+^ in and out of the organelles based on organellar Mg^2+^ usage (Fig. 5). An important example of this occurrence is the rhythmic Mg^2+^ contribution to daily photosynthesis rates which enhances the activity of rubisco in plant chloroplasts [25, 83]. A second important determinant of cytosolic Mg^2+^ availability is Mg^2+^ binding and use by various proteins, such as ribosomal proteins requiring Mg^2+^ for stability or enzymes requiring Mg^2+^ for activity. Mg^2+^ use by proteins is intrinsically linked to the activity of mTOR. Modulating mTOR activity, thereby contributing to the overall control of cellular energy metabolism and translation, has already been identified as a functional consequence of circadian rhythms in intracellular Mg^2+^ (Box 1, Fig. 5) [14, 28]. mTOR has many functions in the cell, including balancing metabolism, protein synthesis and cellular growth or proliferation with the availability of cellular energy: all processing requires Mg^2+^ and Mg‐ATP. Therefore, the regulation by Mg^2+^ of metabolic proteins as well as its own transport systems suggests a potential for a feedback loop between Mg^2+^ transport activity and metabolic protein use (Fig. 5).
Finally, as Mg‐ATP contributes to the allosteric regulation of Mg^2+^ transport proteins (see above), ATP abundance plays an important role for Mg^2+^ abundance in the cell. As the most abundant cellular Mg^2+^ binding partner [72], the cellular pathways linked to ATP generation and degradation have a significant impact on the availability of free Mg^2+^. Therefore, investigating the impact on [Mg^2+^]i of proteins involved in the generation of ATP and control of cellular nucleotide equilibrium is crucial. As an example, the enzyme adenylate kinase (AdK) is an interesting protein to investigate further. AdK is responsible for the interconversion of ATP and AMP to ADP, thus modulating nucleotide (or adenylate) levels in cells. AdK is considered an indirect regulator of [Mg^2+^]i and its subcellular distribution through the regulation of cellular adenylate levels and the modulation of organellar membrane potential [78, 79, 80, 84, 85, 86]. However, Mg^2+^ has a role in regulating AdK activity through Mg^2+^ binding and Mg‐ATP [87], indicating that AdK is under feedback control from Mg^2+^ [86]. Indeed, it has previously been suggested that in photosynthetic organisms, AdK is regulated by diurnal changes in stromal Mg^2+^ [88]. AdK could be instrumental in the rhythmic change of [Mg^2+^]i and [Mg‐ATP]i (Fig. 5), and potential circadian control of feedback between rhythmic [Mg^2+^]i and rhythmic AdK activity is therefore worth investigating. Overall, we propose a feedback model in which the quantity of potential binding sites (protein or ATP) for Mg^2+^ over the 24‐h cycle drives rhythmic transport activity (Fig. 5), as opposed to a unidirectional model where rhythmic Mg^2+^ transport exclusively confers rhythmicity onto metabolism.
Conclusions and perspectives
Advancing the frontier of understanding of dynamic regulation of magnesium is significant to many aspects of medical and crop science research, as magnesium ions are essential to biochemistry. Magnesium‐dependent enzymes function in every metabolic pathway, including every step that requires ATP as a source of energy, and dynamic regulation of their function by Mg^2+^ availability has now been observed (Box 1) [14, 89]. There are even indications of the involvement of magnesium extrusion in establishing sleep [90], hibernation [91] and torpor [92], pointing to magnesium as a potential meta‐regulator of metabolic state [93]. As the importance of high‐amplitude circadian rhythms in magnesium is evident, we have provided a perspective that outlines potential transmembrane proteins responsible for circadian magnesium levels across taxa. We conclude that protein families with prokaryotic members have the highest degree of conservation across eukaryotes and hypothesise that these proteins might generate magnesium rhythms. While circadian rhythms in magnesium have not yet been identified in prokaryotes, the cyanobacterial circadian clock entrains to experimentally imposed rhythms in magnesium supply [19, 94]. Alternatively, unknown factors conserved across eukaryotes could act upon the activity of divergent transport systems. This Perspective paper provides directions towards understanding the molecular mediators that underly [Mg^2+^]i rhythms, as well as their rhythmic regulation and potential feedback mechanisms with rhythmic metabolism.
Author contributions
HKF undertook the phylogenetic analyses and made the figures. HKF and GvO conventionalised, structured and wrote the manuscript.
Supporting information
Table S1. Genome versions and protein accessions numbers used in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dunlap JC (1999) Molecular bases for circadian clocks. Cell 96, 271–290.9988221 10.1016/s 0092-8674(00)80566-8 · doi ↗ · pubmed ↗
- 2Bell‐Pedersen D , Cassone VM , Earnest DJ , Golden SS , Hardin PE , Thomas TL and Zoran MJ (2005) Circadian rhythms from multiple oscillators: lessons from diverse organisms. Nat Rev Genet 6, 544–556.15951747 10.1038/nrg 1633 PMC 2735866 · doi ↗ · pubmed ↗
- 3Njus D (1976) The search for the biochemical clock. Trends Biochem Sci 1, 79–80.
- 4Njus D , Sulzman FM and Hastings JW (1974) Membrane model for the circadian clock. Nature 248, 116–120.4818914 10.1038/248116 a 0 · doi ↗ · pubmed ↗
- 5Nitabach MN , Holmes TC and Blau J (2005) Membranes, ions, and clocks: testing the Njus–Sulzman–Hastings model of the circadian oscillator. Methods Enzymol 393, 682–693.15817319 10.1016/S 0076-6879(05)93036-X · doi ↗ · pubmed ↗
- 6Wang Z‐Y and Tobin EM (1998) Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA 1) gene disrupts circadian rhythms and suppresses its own expression. Cell 93, 1207–1217.9657153 10.1016/s 0092-8674(00)81464-6 · doi ↗ · pubmed ↗
- 7Bargiello TA and Young MW (1984) Molecular genetics of a biological clock in drosophila . Proc Natl Acad Sci USA 81, 2142–2146.16593450 10.1073/pnas.81.7.2142 PMC 345453 · doi ↗ · pubmed ↗
- 8Mc Clung CR , Fox BA and Dunlap JC (1989) The Neurospora clock gene frequency shares a sequence element with the drosophila clock gene period. Nature 339, 558–562.2525233 10.1038/339558 a 0 · doi ↗ · pubmed ↗