Fungal microbiota diversity in Aedes, Anopheles and Culex and their potential use as biological tools to control vector-borne diseases, a systematic review
Aicha Fatimata Sodré, Doubé Lucien Lamy, Nouhoun Traore, Abdoulaye Diabaté, Fernand Sankara, Etienne Bilgo, Emma Camacho, Daniel W Pérez-Ramos, Zhen Zou, Holly Nichols, Irene Ricci

TL;DR
This paper reviews how fungi in mosquitoes could be used to control diseases like malaria and dengue by blocking pathogens or reducing mosquito survival.
Contribution
The study systematically reviews fungal microbiota in Aedes, Anopheles, and Culex mosquitoes and their potential for vector control.
Findings
Fungal species like Wickerhamomyces anomalus and Microsporidia MB reduce vector capacity in Anopheles species.
Fungi inhibit larval development and show antivectorial potential in mosquito organs.
Ascomycota, Basidiomycota, and Microsporidiomycota are the predominant fungal phyla in mosquitoes.
Abstract
Mosquitoes harbor diverse and dynamic microbiota which plays a critical role in shaping their development, survival and vector competence. Many studies have focused on the fungal microbiota of mosquitoes to develop new mosquito control strategies and pathogen blocking tools. This review aims to synthesize current knowledge on the composition and functional roles of the mosquito fungal microbiota, highlighting its potential as a tool for innovative vector born disease control strategies. This review was written in accordance with PRISMA guidelines. Studies on the fungal microbiota of species of the genera Aedes, Anopheles, and Culex and their potential in vector control were searched for in two databases (PubMed, Google Scholar). The quality of the included studies was assessed using the Joanna Briggs Institute (JBI) critical appraisal tool. Fungal diversity was investigated according…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
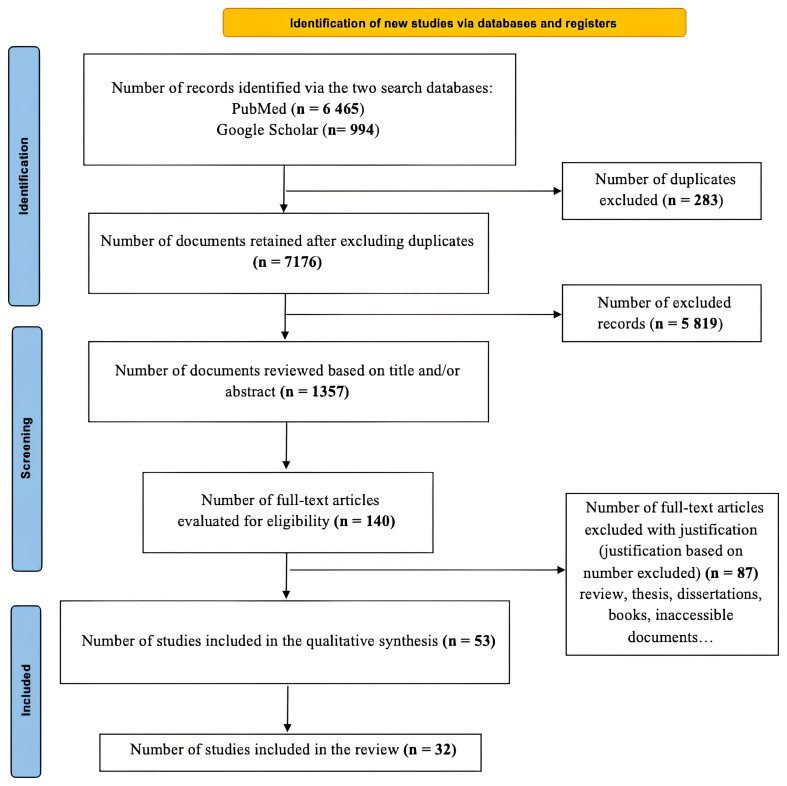 Figure 1
Figure 1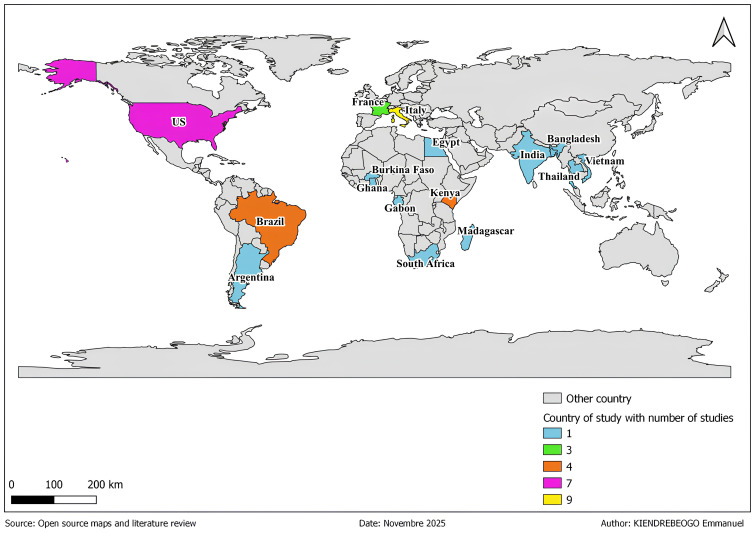 Figure 2
Figure 2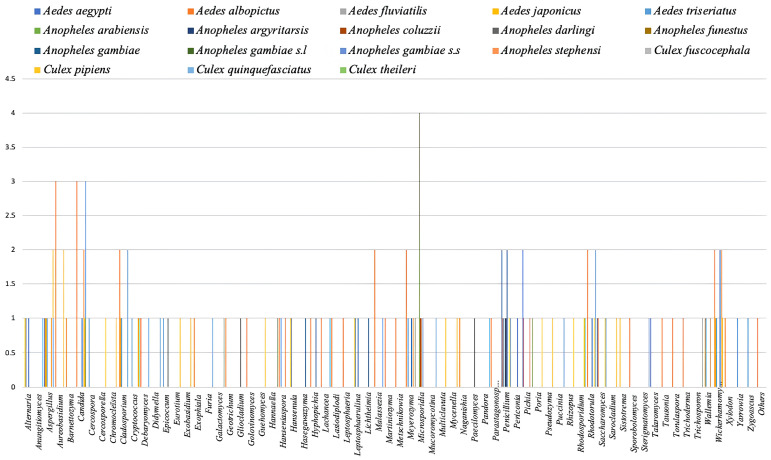 Figure 3
Figure 3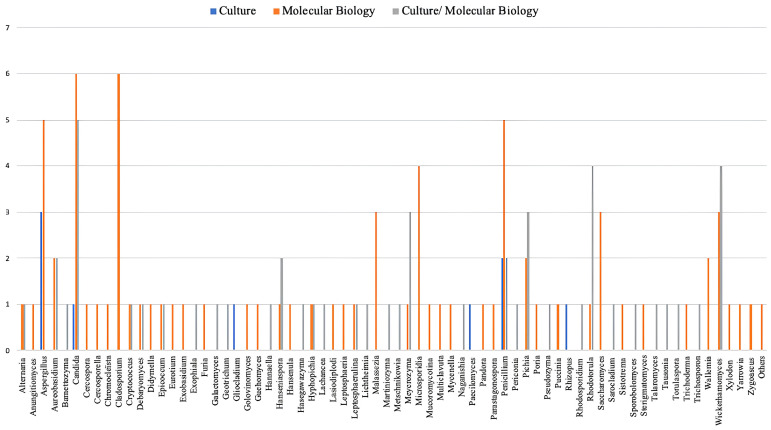 Figure 4
Figure 4| Fungi species | Life history
| Impact
| Pathogen used | Dev stage used | Mosquitoes species | Reference |
|---|---|---|---|---|---|---|
|
| No survival and fecundity effect | Impairs the transmission |
| Adults |
| (
|
|
| Delayed the development time | - | Larvae |
| (
| |
|
| - | Inhibitory activity on the development of early sporogonic stages |
| Adults |
| (
|
|
| Reduced survival rate, inhibits pupation | - | Larvae |
| (
| |
|
| Gradual reduction of larvae |
|
| Larvae |
| (
|
|
| Slows larvae growth | Larvae |
| (
| ||
|
| Slower development, produces smaller adult |
|
| Larvae |
| (
|
|
| Slower development, produces smaller adult |
|
| Larvae |
| (
|
- —Wellcome Trust
- —African Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · Entomopathogenic Microorganisms in Pest Control · Studies on Chitinases and Chitosanases
Introduction
The emergence or re-emergence of mosquito-borne diseases is sometimes linked to factors such as the evolution of pathogens, resistances developed by insect vectors, socio-demographic changes (population growth, human mobility, uncontrolled and often rapid urbanization) and climate change ( Garrido et al., 2023; Zheng et al., 2023). Vector-borne diseases, particularly those transmitted by mosquitoes of the Culicidae family, represent a major public health problem in the world ( de Bitencourt et al., 2021; WHO, 2020). These diseases constitute a heavy burden for populations living especially in tropical and subtropical regions ( Katak et al., 2021; Shaw & Catteruccia, 2019), and as well as a handicap to the socio-economic development of endemic countries ( Saldaña et al., 2017; Shaw & Catteruccia, 2019). Mosquitoes of Culicidae family are widely distributed around the world and the most problematic of these belong to the genus Aedes, Anopheles and Culex ( Gao et al., 2020; Rodgers et al., 2017). Females mosquito vectors after mating necessarily need a blood meal as a source of protein for their eggs maturation ( LaReau et al., 2023; Sarma et al., 2022). When toking this blood meal biting infected human or animal hosts, female mosquitoes ingests at the same time potential pathogens ( Batson et al., 2021; Fontenille et al., 2017). These pathogens are transported into the insect intestines where they infect intestinal epithelial cells, then penetrate into hemolymph and invade the salivary glands, to finally being able to be transmitted to a new host during the next blood-feeding ( Duvallet et al., 2018; Romoli & Gendrin, 2018). Among these pathogens transmitted by mosquitoes malaria protozoa, lymphatic filariasis ( Malassigné et al., 2020; WHO, 2022) an arboviruses including the one of dengue fever, yellow fever, chikungunya, Japanese encephalitis and West Nile fever ( Gao et al., 2020; Malassigné et al., 2020). Bacteria such as Rickettsia felis responsible ( Dieme et al., 2015) and Francisella tularensis responsible of tularemia transmitted by Aedes mosquitoes ( Jonckers Nieboer et al., 2023), and fungi such as Candida parapsilosis probably responsible of candidemia and transmitted by Anopheles, Aedes and Culex mosquitoes ( Bozic et al., 2017). The vector control constitutes a key component of combating vector-borne diseases. Insecticides were for long time the principal component of this strategy ( WHO, 2022). Unfortunately, the excessive use of insecticides agents raised chemical pollution of the environment and insect resistance ti several molecules ( Batson et al., 2021; Chala & Hamde, 2021; Dahmana & Mediannikov, 2020). Innovative, complementary, and effective strategies respectful of environmental health are needed for effective vector control. The diversity and dynamics of microorganisms in mosquitoes including bacteria, fungi, viruses could constitute a base of this innovation needed. They colonize the intestine, the salivary glands, the reproductive organs and could be used for blocking the pathogen development inside the mosquitoes. For example, microorganisms such as Asaia bacteria ( Favia et al., 2007), Wolbachia ( Bian et al., 2013), Pantoea stawartii et Pantoea agglomerans ( Lindh et al., 2008; Wang et al., 2012), Serratia marcescens ( Bando et al., 2013; Wang et al., 2017), entomopahogenic fungi ( Metarhizium anisopliae) ( Fang et al., 2011) and fungal microbiota ( Wickerhamomyces anomalus) ( Cappelli et al., 2021) have been proposed for malaria control.
Entomopathogenic fungi (such as Beauveria, Metarhizium) have proven to be effective biological control agents due to their efficacy and lack of impact on non-target organisms ( Perumal et al., 2024). In Burkina Faso, the virulence of local strains of Metarhizium pingshaense (Met_S10; Met_S26) has been tested against Anopheles coluzzii mosquitoes ( Bilgo et al., 2018). In addition, when combined with a chemical insecticide (Deltamethrin), these different local strains (Met_S10; Met_S26, Met_S31) made the mosquitoes more susceptible to death by the chemical ( Lamy et al., 2025).
Several studies have focused on the diversity of the fungal microbiota of mosquitoes species of genera Aedes, Anopheles and Culex ( Flores et al., 2022; Hegde et al., 2024; Nattoh et al., 2021a) and the interest of certain fungi in the symbiotic fight against malaria ( Angleró-Rodríguez et al., 2016; Cappelli et al., 2019; Herren et al., 2020; Valzano et al., 2016) and certain arbovirosis ( Angleró-Rodríguez et al., 2017)
This systematic review is carried out to synthesize current knowledge on the fungal microbiota diversity in mosquito genera Aedes, Anopheles and Culex and analyze the perspectives for using them as biological control tools against vector-borne diseases.
Methods
Data collection
This review was written in accordance with the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) (2020) protocol for systematic reviews ( Page et al., 2021). A duly completed PRISMA checklist was attached (Supplementary File 1. Table I).
Research papers used in this study were collected in two electronic databases from September 2023 to April 3, 2025 including PubMed and Google Scholar. From the keywords Anopheles spp, Aedes spp , Culex spp, fungi microbiota and vector control we built a research algorithm using Boolean operators as follow (( Anopheles spp) AND ( Aedes spp) AND ( Culex spp) AND (Fungi microbiota) AND (Vector control) OR ( Anopheles spp) OR ( Aedes spp) OR ( Culex spp) OR (Fungi microbiota) OR (Vector control)).
The publication date filter for this review, set at 20 years, aims to provide an up-to-date and in-depth analysis of the fungal communities of mosquitoes of genera Aedes, Anopheles and Culex and their potential in vector control. We also conducted a manual search of the reference lists of identified studies and previously published reviews to identify other relevant studies. No articles from sources were added in order to preserve the systematic nature of this analysis.
The article databases were first imported into Rayyan software for manual filtering ( https://www.rayyan.ai/ consult on April 3, 2025 ( Ouzzani et al., 2016). Using this software, we first eliminated duplicate articles, then eliminated irrelevant articles and sorted the articles to be included according to the inclusion criteria. Rayyan software enabled the development of the PRISMA diagram ( Figure 1).
PRISMA flow diagram showing the study selection process.
Eligibility criteria
Inclusion criteria:
- Studies published in the last 20 years (2005–2025)- Studies published in english in pure reviewed journal- Studies published on any of the target mosquito genera ( Aedes, Anopheles and Culex)- Studies tolking about fungal and vector control
Exclusion criteria:
- All studies not fiting with the inclusion criteria listed- Review studies- Dissertations and theses- Inaccessible articles
Eligibility assessment and data extraction
The articles were sorted manually and independently by four authors. These authors first reviewed the titles and abstracts and discussed any discrepancies until a consensus was reached. Next, each author independently reviewed the titles and abstracts of all extracted articles. In cases of disagreement, consensus was reached by reviewing the full text of the various articles. If necessary, a fifth and sixth author were consulted for the final decision. Four authors then independently reviewed the full-text articles for inclusion. However, in the event of further disagreements, consensus was reached through discussion of the inclusion or exclusion criteria. Otherwise, the fifth and sixth authors were consulted.
Discrepancies were resolved by comparing the extracted data. The authors verified the accuracy of the extracted data.
Quality and risks of bias evaluation of included the studies
The quality and risks of bias of included studies was determined using the Joanna Briggs Institute (JBI) critical appraisal tool, which comprises 9 parameters ( https://jbi.global/critical-appraisal-tools) to cohort, case-control, or cross-sectional studies.
A score of “1” was assigned to each parameter that was well completed and “0” if it was not described. The top-quality score was 9. The quality of each study was assessed as high if it was ≥ 7 points, moderate if the score was between 4 and 6 points, and low if the score was < 4 points. The quality of the studies was assessed independently by three evaluators and a fourth evaluator for consensus in case of disagreement.
Data summary
The following variables were collected: lead author, study location, mosquito origin (field or laboratory), developmental stage (larvae, adults), isolation and identification techniques, dissected tissues (whole larvae, whole adults, organs), mosquito vector species of the genera Aedes, Anopheles, and Culex, and finally, if available, the potential antivector properties of fungal species in the mosquito microbiota.
A map of the geographical distribution of the studies was developed using information on the location where the mosquitoes were collected or just the country of study when articles did not provide the sample origin. This map was created using QCIS 3.4 software. In addition, Microsoft Excel was used to create graphs providing information on fungal diversity according to species of the genera Aedes, Anopheles, and Culex, as well as according to the isolation and identification techniques used in each study included.
Results
Study results
Of the total of 7,458 articles initially were identified, only thirty-two (32) fully met the eligibility criteria after removing duplicates (283) and applying the inclusion and exclusion criteria. Three (3) articles discussing the antivectorial potential of two fungal strains appeared to meet the inclusion criteria but were excluded because the strains came from previous studies that did not meet the selection criteria ( Barnard et al., 2007; Frankel-Bricker, 2020; Frankel-Bricker et al., 2020).
There were 12 studies associating the diversity of microorganisms (fungi and/or bacteria, viruses) in the microbiota of mosquito species. More specifically, only four included studies focused on the anti-plasmodial potential of certain fungal strains ( Cappelli et al., 2014; Cappelli et al., 2019; Herren et al., 2020; Valzano et al., 2016) among articles studying the diversity of the fungal microbiota of Anopheles mosquitoes responsible of malaria. No study on the diversity of the fungal microbiota of mosquitoes in Aedes and Culex responsible of arboviruses infections have been investigated their antiviral potential.
Risks of bias
The Joanna Briggs Institute (JBI) critical appraisal tool, comprising nine points, was used to assess the quality of the studies. The different thresholds of the bias risk checklist are pointed out in the Supplementary File 2. Table II. Of the 32 studies included and evaluated, 28 studies correctly met the 9 points of the checklist with a threshold of 100%, indicating a low risk rate.
With a threshold of 77.78%, two studies each presented a high risk of bias and an uncertain risk of bias due to the non-representativeness of the target population and inappropriate statistical analysis, respectively ( Gusmão et al., 2007; Tajedin et al., 2009). Furthermore, with a threshold of 88.89% each, two studies presented a high risk of bias due to inappropriate statistical analysis ( da S Pereira et al., 2009) and an uncertain risk of bias due to a small sample size ( Chandler et al., 2015), respectively.
In general, all studies from the two databases (PubMed, Google Scholar) evaluated and included in the systematic review were of high quality, thereby reinforcing knowledge about the fungal diversity of species of the genera Aedes, Anopheles, and Culex and their antivectorial potential.
Study characteristics
The detailed characteristics of the studies are presented in the Supplementary file 3. Table III.
** Geographical distribution of studies on fungal diversity in the microbiota of mosquitoes of the genera Aedes, Anopheles, and Culex **
The map in Figure 2 shows the global distribution of studies on the fungal microbiota of Aedes, Anopheles, and Culex mosquitoes. Countries are represented by different colors depending on the number of studies. The number of studies per country ranges from 0 to 9.
Geographic distribution of published studies investigating the fungal microbiota associated with Aedes, Anopheles, and Culex mosquitoes.
The gray color represents countries where no studies were found according to research and inclusion criterias. These resulting countries include several countries in Central and West Africa, in Central Asia and in Oceania.
Countries with 1 to 2 studies includes in this review are represented in color light blue in the map. These countries include South Africa ( Steyn et al., 2016), Argentina ( Flores et al., 2022), South Asia ( Bozic et al., 2017), Burkina Faso ( Bozic et al., 2017), Egypt ( Galal et al., 2017), Gabon ( Zouache et al., 2022), Ghana ( Akorli et al., 2024), India ( Tajedin et al., 2009), Madagascar ( Luis et al., 2019), Thailand ( Thongsripong et al., 2017; Thongsripong et al., 2021), and Vietnam ( Luis et al., 2019).
In France, represented in light green color, three studies have focused on the fungal microbiota of Aedes mosquitoes ( Luis et al., 2019; Guégan et al., 2020a; Malassigné et al., 2025).
Furthermore, in countries such as Brazil ( Gusmão et al., 2007; da S Pereira et al., 2009; Gusmão et al., 2010; Bozic et al., 2017) and Kenya ( Herren et al., 2020; Nattoh et al., 2021a; Onchuru et al., 2024) represented in color orange, four studies focused on the fungal microbiota Aedes and Anopheles mosquitoes.
In addition, seven studies focused on the fungal microbiota of Aedes, Anopheles, and Culex mosquitoes in the United States, represented in color light purple ( Angleró-Rodríguez et al., 2016; Angleró-Rodríguez et al., 2017; Bishop-Lilly et al., 2010; Chandler et al., 2015; Hegde et al., 2024; Muturi et al., 2016; Tawidian et al., 2021).
Finally, Italy, represented in color yellow on the map, is the country where several studies (9) have been conducted on the fungal microbiota of Aedes, Anopheles, and Culex ( Bozic et al., 2017; Cappelli et al., 2014; Cappelli et al., 2019, Cappelli et al., 2023; Ricci et al., 2011a; Ricci et al., 2011b; Valzano et al., 2016).
There is an uneven distribution of studies, with a greater number of studies in Italy, the United States, and Brazil. The lack of data in certain countries such as Central Africa, several West African countries, Central Asia, and Oceania may lead to geographical bias in scientific writing on the fungal microbiota of Aedes, Anopheles and Culex mosquitoes, considering that certain fungal species may be specific ecosystem.
** Fungal diversity associated with the microbiota of Aedes, Anopheles, and Culex species **
The analyse of the distribution of fungal genera in the microbiota according to mosquito genera ( Aedes, Anopheles or Culex) showed variation in the fungal microbiota composition of the microbiota of certain mosquito species. These fungal genera are most often composed of yeast-like fungi and filamentous fungi that can be commensal, symbiotic, or pathogenic to mosquitoes.
Fungal diversity associated with the microbiota of Aedes species
The analyse of the Aedes fungal microbiota according to literature showed that studies were focused on species such as Aedes aegypti ( Ae. aegypti), Aedes albopictus ( Ae. albopictus), Aedes fluviatilis ( Ae. fluviatilis), Aedes japonicus ( Ae. japonicus), and Aedes triseriatus ( Ae. triseriatus), represented by different colors respectively ( Figure 3).
Distribution of fungal genera in the mosquito microbiota according to mosquito genera ( Aedes, Anopheles and Culex).
In total of eight fungal genera, including five genera belonging to yeast-like fungi and three genera belonging to filamentous fungi (molds), Ae. aegypti has a relatively undiversified microbiota, with a predominance of the fungi genera belonging to Candida, Penicillium, Pichia, and Wickerhamomyces. Although weakly associated with this mosquito species, yeast-like fungi such as the genera Meyerozyma and Saccharomyces ( Bishop-Lilly et al., 2010; Bozic et al., 2017) and molds such as the genera Aspergillus and T alaromyces ( Angleró-Rodríguez et al., 2017; Bishop-Lilly et al., 2010) have been identified.
Unlike Ae. aegypti, with more than thirty fungal genera, including 17 genera of yeast-like fungi and 11 genera of filamentous fungi (molds, phytopathogens), Ae. albopictus has a more diverse microbiota with a predominance of Aureobasidium, Candida, Cladosporium, Malassezia, Meyerozyma, Rhodotorula, and Wickerhamomyces. Yeast-like fungal genera weakly associated with Ae. albopictus include Barnettozyma, Debaryomyces, Geotrichum, Hanseniaspora, Hyphopichia, Lachancea, Martiniozyma, Metschnikowia, Naganishia, Pichia, Saccharomyces, Sporobolomyces, Tausonia, and Torulaspora ( Bozic et al., 2017; Hegde et al., 2024; Luis et al., 2019; Malassigné et al., 2025). As for filamentous fungi, the weakly identified genera belong to molds such as Aspergillus, Exophiala, Penicillium, and Trichoderma ( Guégan et al., 2020a; Luis et al., 2019; Malassigné et al., 2025) phytopathogens including Golovinomyces, Lasiodiplodia, Parastagonospora, Leptosphaeria, and Xylodon ( Guégan et al., 2020a).
Furthermore, Ae. fluviatilis harbors a relatively weak microbiota consisting solely of a filamentous fungus of the genus Aspergillus ( da S Pereira et al., 2009).
Finally, the two species Ae. japonicus and Ae. triseriatus appear to harbor microbiota that are not very diverse but identical in filamentous fungi, including molds such as the genera Alternaria, Aspergillus, Cladosporium, and Penicillium, and phytopathogens such as Cercospora, Epicoccum, and Sistotrema ( Muturi et al., 2016). The yeast-like fungi common to these two-mosquito species include the genera Hansenula, Meyerozyma, Wallemia, Wickerhamomyces, Yarrowia, and Zygoascus ( Muturi et al., 2016). Specifically, the genera Cryptococcus and Hanseniaspora have been identified in the microbiota of Aedes triseriatus ( Muturi et al., 2016).
Fungal diversity associated with the microbiota of Anopheles species
The analyse of the Anopheles mosquitoes fungal microbiota according to this review showed that Anopheles included studies were focused on the species including such as Anopheles arabiensis ( An. arabiensis), Anopheles argyritarsis ( An. argyritarsis), Anopheles coluzzii ( An. coluzzii), Anopheles darlingi ( An. darlingi), Anopheles funestus ( An. funestus), Anopheles gambiae ( An. gambiae), Anopheles gambiae s.l. ( An. gambiae s.l.), Anopheles gambiae s.s. ( An. gambiae s.s.) and Anopheles stephensi ( An. stephensi), represented respectively in different colors ( Figure 3).
Species such as An. arabiensis, An. coluzzii, An. funestus, An. gambiae s.l, and An. gambiae s.s. each harbor a microbiota poor fungal microbiota, with only one genus ( Microsporidia) and specifically intracellular ( Akorli et al., 2024; Herren et al., 2020; Nattoh et al., 2021b; Onchuru et al., 2024). This genus Microsporidia was predominant in An. arabiensis.
In addition, An. argyritarsis and An. darlingi showed a poor microbiota, with only one genus ( Penicillium) and three genera ( Penicillium, Gliocladium, Paecilomyces) respectively, all of which are filamentous fungi ( da S Pereira et al., 2009).
On the other hand, with a predominance of the genus Wickerhamomyces, An. gambiae hosts a fairly diverse fungal microbiota with six genera of yeast-like fungi and seven genera of filamentous fungi. Yeast-like fungi such as the genera Candida, Hasegawazyma, Hyphopichia, Meyerozyma, and Rhodotorula ( Bozic et al., 2017; Nattoh et al., 2021a) as well as filamentous fungi including molds (genera Alternaria, Penicillium, Periconia) ( Angleró-Rodríguez et al., 2016; Nattoh et al., 2021a) and certain phytopathogens ( Epicoccum, Leptosphaerulina, Lichtheimia) ( Nattoh et al., 2021a) have been weakly associated with An. gambiae.
Finally, An. stephensi harbors a less diverse fungal microbiota including six genera of yeast-like fungi and two genera of filamentous fungi. While the genera Candida and Wickerhamomyces are predominant, other yeast-like genera such as Hanseniaspora, Meyerozyma, Pichia, and Wallemia ( Bozic et al., 2017; Ricci et al., 2011a) and a few filamentous genera such as Aspergillus and Penicillium ( Angleró-Rodríguez et al., 2016; Tajedin et al., 2009) are weakly associated with An. gambiae.
Fungal diversity associated with the microbiota of Culex species
The results showed that for Culex mosquitoes fungal microbiota, the included studies were focused on species including such as Culex fuscocephala ( Cx. fuscocephala), Culex pipiens ( Cx. pipiens), Culex quinquefasciatus ( Cx. quinquefasciatus), and Culex theileri ( Cx. theileri), represented by different colors ( Figure 3).
Cx. pipiens appears to harbor a microflora rich in fungi of eight yeast-like genera (Candida, Cryptococcus, Guehomyces, Malassezia, Pseudozyma, Rhodosporidium, Rhodotorula, Saccharomyces, Wickerhamomyces), 10 filamentous genera with molds (Aspergillus, Aureobasidium, Cladosporium, Eurotium, Penicillium, Rhizopus) and phytopathogens ( Cercosporella, Exobasidium, Mycenella, Sarocladium) ( Chandler et al., 2015; Galal et al., 2017; Steyn et al., 2016).
lichenized ( Multiclavuta) and lignivorous ( Mycenella, Poria) fungi genera have been also found ( Chandler et al., 2015). All other fungal genera are found weakly associated to Cx. Pipiens except two predominant genera Aspergillus and Aureobasidium.
Some fungi with unspecified genera belonging to various phyla, including Ascomycota, Basidiomycota, and Zoopagomycota, have been identified in the Cx. fuscocephala microbiota ( Thongsripong et al., 2021).
Furthermore, microbiota undiversified Cx. Pipiens and Cx. quinquefasciatus species even undiversified, harbors fungi from seven yeast-like genera ( Candida, Debaryomyces, Lachancea, Malassezia, Meyerozyma, Rhodotorula, and Sterigmatomyces) ( Bozic et al., 2017; Flores et al., 2022; Hegde et al., 2024), eight filamentous genera with molds ( Anungitiomyces, Aspergillus, Cladosporium, Mucoromycotina, and Penicillium), phytopathogens ( Didymella, Puccinia), and entomopathogens ( Furia) ( Flores et al., 2022; Thongsripong et al., 2017). A predominance of Candida, Cladosporium, and Rhodotorula fungi was observed among the genera associated with Cx. quinquefasciatus.
Finally, only yeast-like fungi from nine genera ( Candida, Cryptococcus, Galactomyces, Hannaella, Meyerozyma, Pichia, Rhodosporidium, Rhodotorula, Trichosporon) were found in microbiota Cx. theileri ( Steyn et al., 2016).
** Fungal diversity associated with the microbiota of Aedes, Anopheles, and Culex species according to culture techniques **
The analyses of fungal genera according to the techniques used for isolation and identification such as culture-dependent techniques involving homogenate culture on agar media, molecular biology techniques on homogenate and molecular biology techniques coupled culture- techniques of homogenate on agar media ( Figure 4).
Distribution of fungal genera in Aedes, Anopheles and Culex mosquito microbiota by isolation and identification techniques.
Fungal diversity according to culture-dependent technique
The culture-dependent technique for isolating fungal genera appears to be the least used in studies. Thus, Aspergillus and Penicillium are the two predominant filamentous fungal genera isolated from the microbiota using the culture technique. In addition, this technique allows the isolation of small proportions of other fungal genera, including 1 yeast-like species such as Candida and 2 filamentous species ( Gliocladium, Paecilomyces, Rhizopus) ( Galal et al., 2017; da S Pereira et al., 2009; Tajedin et al., 2009).
Fungal diversity according to molecular biology techniques coupled with culture
In numerous studies, it has been possible to isolate several fungal genera using molecular biology techniques combined with culture. Most of the fungal genera isolated are yeast-like (7 in total, including Candida, Rhodotorula, Hanseniaspora, Meyerozyma, Pichia, Wickerhamomyces, and Hanseniaspora) and filamentous fungi (including Aureobasidium and Penicillium ( Angleró-Rodríguez et al., 2016; Bozic et al., 2017; Cappelli et al., 2023; Gusmão et al., 2007; Hegde et al., 2024; Malassigné et al., 2025; Ricci et al., 2011a; Steyn et al., 2016).
As for the other minority isolated genera, they include 19 yeast-like species ( Barnettozyma, Cryptococcus, Debaryomyces, Galactomyces, Geotrichum, Hannaella, Hasegawazyma, Hyphopichia, Lachancea, Martiniozyma, Metschnikowia, Naganishia, Pseudozyma, Rhodosporidium, Sporobolomyces, Tausonia, Torulaspora, Trichosporon, Yarrowia) ( Bozic et al., 2017; Malassigné et al., 2025; Nattoh et al., 2021a; Steyn et al., 2016). In addition to these yeast-like genera, nine filamentous genera composed of molds and phytopathogens including Alternaria, Epicoccum, Exophiala, Leptosphaerulina, Lichtheimia, Periconia, Sarocladium, Talaromyces and Trichoderma are weakly represented ( Angleró-Rodríguez et al., 2017; Nattoh et al., 2021a; Steyn et al., 2016).
Fungal diversity according to molecular biology techniques
Given the sensitivity of this technique, it allowed the identification of several other fungal genera in addition to those isolated by the association of molecular biology technique with culture. technique The predominant fungi consist of seven yeast-like genera such as Candida ( Bozic et al., 2017; Flores et al., 2022; Gusmão et al., 2010; Hegde et al., 2024; Luis et al., 2019; Ricci et al., 2011a), Malassezia ( Chandler et al., 2015; Guégan et al., 2020a; Hegde et al., 2024), Sacharomyces ( Bishop-Lilly et al., 2010; Chandler et al., 2015; Hegde et al., 2024) Wickerhamomyces ( Muturi et al., 2016; Ricci et al., 2011a; Ricci et al., 2011b), Hansenula ( Muturi et al., 2016), Pichia ( Gusmão et al., 2010; Ricci et al., 2011a), Wallemia ( Muturi et al., 2016; Ricci et al., 2011a). In addition, an obligate intracellular genus ( Microsporidia) and filamentous fungi (5 genera) such as Cladosporium ( Chandler et al., 2015; Flores et al., 2022; Guégan et al., 2020a; Luis et al., 2019; Muturi et al., 2016; Thongsripong et al., 2017), Aspergillus ( Bishop-Lilly et al., 2010; Chandler et al., 2015; Flores et al., 2022; Luis et al., 2019; Muturi et al., 2016), Penicillium ( Bishop-Lilly et al., 2010; Flores et al., 2022; Guégan et al., 2020a; Muturi et al., 2016; Zouache et al., 2022), Alternaria ( Muturi et al., 2016), Aureobasidium ( Guégan et al., 2020a; Luis et al., 2019) are mostly identified.
The fungal genera weakly identified by this technique include several fungal genera (23), composed of molds such as Mucoromycotina ( Flores et al., 2022), Trichoderma ( Guégan et al., 2020a), phytopathogens ( Anungitiomyces, Cercospora, Cercosporella, Didymella, Epicoccum, Eurotium, Exobasidium, Golovinomyces, Lasiodiplodi, Leptosphaeria, Leptosphaerulina, Parastagonospora, Puccinia, Sistotrema, Xylodon) ( Chandler et al., 2015; Flores et al., 2022; Guégan et al., 2020a; Muturi et al., 2016), one entomopathogen ( Furia, Pandora) ( Thongsripong et al., 2017), and other fungi ( Multiclavuta, Mycenella, Poria) ( Chandler et al., 2015; Thongsripong et al., 2017).
In addition, yeast-like genera (9) such as Cryptococcus, Debaryomyces, Guehomyces, Hanseniaspora, Hyphopichia, Meyerozyma, Rhodotorula, Sterigmatomyces, and Zygoascus are also identified in small quantities ( Chandler et al., 2015; Flores et al., 2022; Luis et al., 2019; Muturi et al., 2016).
** Fungal diversity associated with different stages of development of the species Aedes, Anopheles, Culex **
Mosquitoes host a diverse fungal community within their tissues, the composition of which varies according to developmental stage (larvae, pupae, adults) and origin of specimens (field or breeding) (Supplementary file 3. Table III).
At the larval stages
The fungal composition of larvae appears to vary depending on the mosquito species, the origin of the mosquito, and the larval habitat.
For example, while larvae of Ae. aegypti collected in the field (Lopé village) harbored three (3) phyla ( Ascomycota, Basidiomycota, Zycomycota) ( Zouache et al., 2022), only one phylum ( Ascomycota) with the species Candida parapsilosis was found at different stages of development (larvae, nymphs) in Ae. aegypti from a laboratory breeding program (Camerino) ( Bozic et al., 2017). Several yeasts belonging to two phyla ( Ascomycota, Basidiomycota), mainly of the genera Debaryomyces, Pichia, and Wickerhamomyces, and a filamentous fungus of the genus Exophiala ( Ascomycota) were found in Ae. albopictus larvae collected in the field (Community garden of Saint Priest) ( Malassigné et al., 2025). In addition, fungal species belonging to four (4) rare fungal phyla ( Chytridiomycota, Glomeromycota, Mucoromycota, Rozellomycota) were identified from the carcasses of Ae. albopictus larvae collected in the field (Manhattan, KS) ( Tawidian et al., 2021). Ae. fluviatilis larvae collected in the field (Municipality of Rio Preto da Eva (Amazonas) hosted a species of the genus Aspergillus belonging to the phylum Ascomycota ( da S Pereira et al., 2009).
In addition, in An. gambiae and An. stephensi species reared under laboratory conditions (Camerino), a yeast species, Candida parapsilosis, belonging to the Ascomycota phylum, was identified at different stages of development (larvae, pupae) ( Bozic et al., 2017). In addition, another yeast species, Wickerhamomyces anomalus, was found in An. gambiae larvae ( Cappelli et al., 2023), as well as at different stages of development (larvae, pupae) of An. stephensi kept in the laboratory (Camerino) ( Ricci et al., 2011a).
Respectively, one filamentous fungal genus ( Aspergillus) and three ( Penicillium, Gliocladium, Paecilomyces), all belonging to the phylum Ascomycota, were observed in larvae of two mosquito species, An. argyritarsis and An. darlingi, collected at the same site (Municipality of Rio Preto da Eva (Amazonas) ( da S Pereira et al., 2009).
Like other species, Cx. quinquefasciatus, bred in the laboratory (Camerino), also hosted the species Candida parapsilosis at different larval stages (larvae, pupae) ( Bozic et al., 2017). In addition to Candida parapsilosis, other species belonging to six genera comprising molds (Aspergillus, Cladosporium, Penicilium, Mucoromycotina) and yeasts ( Debaryomyces, Sterigmatomyces) were isolated from Cx. quinquefasciatus larvae kept under laboratory conditions (Argentina) ( Flores et al., 2022). In addition, filamentous fungal species ( Aspergillus flavus, Penicillium notatum, Rhizopus stolonifer) and yeast-like fungal species ( Candida parapsilosis, Candida pseudolambica, Rhodotorula mucilaginosa, Rhodosporidium diobovatum, Wickerhamomyces anomalus) were found respectively in larvae kept under laboratory conditions (Egypt) ( Galal et al., 2017) and larvae collected in the field (South Africa) ( Steyn et al., 2016). From the same collection site (South Africa) as the Cx. pipiens larvae, the Cx. theileri larvae hosted more fungal species belonging to nine genera ( Candida, Cryptococcus, Galactomyces, Hannaella, Meyerozyma, Pichia, Rhodosporidium, Rhodotorula, Trichosporon) ( Steyn et al., 2016).
Finally, fungal species such as Penicillium georgiense, Penicillium notatum, and Aspergillus flavus were identified in An. gambiae nymphs and Cx. pipiens eggs ( Galal et al., 2017; Nattoh et al., 2021a), highlighting the persistence of fungal species beyond the larval stage.
** In the adults stages **
Adult mosquitoes were found to harbor more diverse mycobiota. The major role of female mosquitoes in pathogens transmission justify the fact that several studies were primarily focused primarily on fungal diversity of these females mosquitoes.
Fungal genera including molds such as Aspergillus and Penicillium and a yeast such as Saccharomyces, all belonging to the phylum Ascomycota, have been identified in female Ae. aegypti mosquitoes bred in the laboratory ( Bishop-Lilly et al., 2010). As for Ae. albopictus, wild females harbored molds (genera Aspergillus, Aureobasidium, Cladosporium), yeasts (genera Candida, Hyphopichia) belonging to the phyla Ascomycota ( Bishop-Lilly et al., 2010), and only yeasts (genera Candida, Lachancea, Malassezia, Rhodotorula, Saccharomyces) belonging to the phyla ( Ascomycota, Basidiomycota) ( Hegde et al., 2024). In addition, a yeast-like fungus Wickerhamomyces anomalus, belonging to the phylum Ascomycota, has been found in male Ae. albopictus ( Cappelli et al., 2023).
Microsporidia MB, an obligate intracellular parasitic fungus belonging to the phylum Microsporidiomycota, has been identified in wild gravid and non-gravid females and in the carcasses of males An. arabiensis ( Herren et al., 2020; Nattoh et al., 2021b; Onchuru et al., 2024). In male and female An. stephensi mosquitoes from an insectary (Camerino), several yeast-like fungi, including the genera Candida, Pichia, Wickerhamomyces, Hanseniaspora, and Wallemia belonging to the phyla Ascomycota and Basidiomycota, were isolated ( Ricci et al., 2011a).
Culex mosquito species (particularly Cx. pipiens, Cx. quinquefasciatus, and Cx. fucoscephala) host a wide variety of fungi, including filamentous fungi (molds, phytopathogens) and yeast-like fungi. Several molds (genera Aspergillus, Cladosporium), yeasts ( Guehomyces, Malassezia, Saccharomyces), phytopathogens ( Cercosporella, Eurotium) and other fungi ( Chromocleïsta, Multiclavuta, Mycenella, Poria) have been found in wild female Cx. pipiens ( Chandler et al., 2015). Fungal genera consisting of a mold ( Aureobasidium), yeasts ( Candida, Cryptococcus, Pseudozyma), and a phytopathogen ( Sarocladium) were also identified in wild adults of Cx. pipiens ( Steyn et al., 2016). When female Cx. pipiens were kept under laboratory conditions, molds (genera Aspergillus, Penicillium, Rhizopus) and a yeast ( Candida) were found ( Galal et al., 2017). In addition, several yeast-like fungal genera (Candida, Malassezia, Saccharomyces) ( Hegde et al., 2024), a filamentous genus ( Cladosporium), an entomopathogenic genus ( Furia) ( Thongsripong et al., 2017), and another genus ( Pandora) ( Thongsripong et al., 2017) have been found in wild female Cx. quinquefasciatus. In one study, only yeast-like fungal species, including Candida oleophila, Lachancea thermotolerans, and Rhodotorula mucilaginosa, were isolated from adult Cx. quinquefasciatus maintained under laboratory conditions ( Hegde et al., 2024).
Finally, in one study, diet appears to influence fungal diversity in male and female Cx. quinquefasciatus kept under laboratory conditions ( Flores et al., 2022). After feeding on sugar, males and females harbored several identical fungal genera, including molds ( Aspergillus, Cladosporium), yeasts ( Candida, Rhodotorula, Sterigmatomyces), and a phytopathogen ( Didymella) ( Flores et al., 2022). Fungal genera such as Penicillium and Mucoromycotina were specifically isolated in sugar-fed males of Cx. quinquefasciatus ( Flores et al., 2022). Blood fed females were found blood to harbor mostly molds ( Anungitiomyces, Aspergillus, Cladosporium) and a phytopathogen (genus Puccinia) fungi ( Flores et al., 2022). In summary, the molds fungi such as Aspergillus and Cladosporium are found in both sugar-fed male Cx. q uinquefasciatus and blood- or sugar-fed females ( Flores et al., 2022).
In the digestive organs
Known for its important role in immunity and metabolism ( Muturi et al., 2016), the gut is the first place where pathogens are transported in infested mosquitoes ( Heu & Gendrin, 2018). It is one of the most studied organs and constitutes an important fungal microhabitat. A yeast-like fungus (genus Meyerozyma) and a mold ( Talaromyces) have been identified in the intestines ( Bozic et al., 2017) and midguts of wild female Ae. aegypti ( Angleró-Rodríguez et al., 2017), respectively Wickerhamomyces anomalus ( Ricci et al., 2011b) and Candida parapsilosis ( Bozic et al., 2017), two yeast species belonging to the phylum Ascomycota, were found in the intestines of adults of Ae. aegypti and Ae. albopictus, respectively, kept under laboratory breeding conditions. Similarly, this yeast species ( Wickerhamomyces anomalus) has been identified in the intestines of wild Ae. aegypti and Ae. albopictus ( Ricci et al., 2011b). A study showed that newly emerged female Ae. aegypti kept in laboratory and fed a diet with sucrose and blood, harbored several fungi species belonging to two genera including Candida and Pichia isolated from the intestines and intestinal diverticula ( Gusmão et al., 2010). Several fungal genera composed of molds ( Aureobasidium, Cladosporium, Penicillium, Trichoderma), a yeast ( Malassezia), and phytopathogens ( Golovinomyces, Lasiodiplodia, Leptosphaeria, Parastagonospora, Xylodon) were identified in the intestines and crops of Ae. albopictus mosquitoes kept in the laboratory ( Guégan et al., 2020a). Two species of yeast, Meyerozyma guilliermondii and Sporobolomyces cf. roseus, belonging to the phyla Ascomycota and Basidiomycota, respectively, have also been identified in the intestines of wild female Ae. albopictus ( Bozic et al., 2017). As for wild females of Ae. japonicus and Ae. triseriatus, several fungal genera, including yeasts ( Cryptoccoccus, Hanseniaspora, Hansenula, Meyerozyma, Wallemia, Wickerhamomyces, Yarrowia), molds ( Alternaria, Aspergillus, Cladosporium, Penicillium) and a phytopathogen ( Leptosphaerulina) have been identified in the midguts ( Muturi et al., 2016). A proliferation of the species Meyerozyma caribbica was observed in the midguts of both Ae. japonicus and Ae. triseriatus mosquitoes fed on infected blood.
In addition, the yeast species Wickerhamomyces anomalus has been identified in the intestines of adult An. gambiae and An. stephensi maintained in different laboratories ( Ricci et al., 2011b; Ricci et al., 2011b). Similarly, Candida parapsilosis has been identified in the intestines of adult An. gambiae and An. stephensi kept in the laboratory and also in the intestines of wild female An. gambiae ( Bozic et al., 2017). Other fungal species belonging to the genera ( Rhodotorula and Meyerozyma) ( Bozic et al., 2017) and ( Candida, Pichia, and Meyerozyma) ( Bozic et al., 2017; Ricci et al., 2011a) have been detected in the intestines of female An. gambiae and An. stephensi kept under laboratory conditions ( Bozic et al., 2017; Ricci et al., 2011a). In wild female An. gambiae, fungal genera including a yeast ( Hyphopichia) and phytopathogens ( Lichtheimia, Lepthosphaerulina, Epicoccum, Periconia) have been identified in the intestines ( Nattoh et al., 2021a). Penicillium chrysogenum, a species of mold, was also found in the midguts of wild adults of An. gambiae and An. stephensi ( Angleró-Rodríguez et al., 2016). Microsporidia MB, an obligate intracellular fungal species, has been detected in the abdomen of wild females of An. arabiensis, An. gambiae s.l., and An. gambiae s.s. ( Akorli et al., 2024), as well as in the midguts of male An. arabiensis kept in the insectarium ( Nattoh et al., 2021b).
A study identified also the yeast Meyerozyma guilliermondii, in the intestines of wild female Cx. quinquefasciatus, as well as another fungi species including Candida parapsilosis, in the intestines of wild adult Cx. quinquefasciatus and those kept under laboratory conditions ( Bozic et al., 2017).
In the reproductive organs
The analyse showed that mosquitoes harbor several yeast-like fungi species in their reproductive organs.
Two species of yeast, Candida parapsilosis and Wickerhamomyces anomalus, have been identified in the reproductive organs (male and female) of Ae. aegypti and Ae. albopictus kept under laboratory conditions ( Bozic et al., 2017; Ricci et al., 2011). In addition, yeasts of the genus Candida and Pichia were found in the ovaries of newly emerged female Ae. aegypti kept in the laboratory ( Gusmão et al., 2010), highlighting possible transmission via the reproductive organs.
Candida parapsilosis and Wickerhamomyces anomalus, two species of yeast, have been found in the reproductive organs (male and female) of An. gambiae and An. stephensi kept in various laboratories ( Bozic et al., 2017; Ricci et al., 2011). A yeast-like fungal genus ( Hyphopichia) and phytopathogens ( Leptosphaerulina, Lichtheimia, Epicoccum, Periconia) have also been isolated in the ovaries of wild female An. gambiae ( Nattoh et al., 2021a). In addition, the intracellular fungus Microsporidian MB has also been identified in the gonads of male An. arabiensis kept in the insectarium ( Nattoh et al., 2021b).
Finally, the species Candida parapsilosis has been identified in the reproductive organs (male and female) of wild Cx. quinquefasciatus and those kept under laboratory conditions ( Bozic et al., 2017).
** Vector control potential of mosquito fungal microbiota species **
Certain fungal species associated with mosquito microbiota show significant potential in vector control, either by inhibiting the development of larval stages or by negatively influencing the transmission of pathogens ( Table IV). For example, fungal species in the mosquito microbiota such as Microsporidia MB and Wickerhamomyces anomalus have demonstrated their effects by inhibiting the transmission of the Plasmodium parasite in An. arabiensis ( Herren et al., 2020) and by reducing vector competence in An. stephensi ( Cappelli et al., 2014; Cappelli et al., 2023; Valzano et al., 2016).
In addition, a filamentous fungus, Leptosphaerulina sp, isolated from the midgut of An. gambiae mosquitoes, delayed the development time of larvae when reintroduced by co-feeding from the larval stage except stage 1 (L1). Also, fungal species such as Candida albicans, Candida glabrata, and Candida pseudolambica isolated from Culex larvae ( Cx. pipiens, Cx. theileri) caused a reduction in survival rate followed by inhibition of pupation, a gradual reduction in larvae, and a slowdown in larval growth when reintroduced by co-feeding in Cx. pipiens larvae ( Steyn et al., 2016). In addition, other yeast species such as Martiniozyma asiatica and Torulaspora delbrueckii caused a delay in larval development, resulting in small adults when they were reintroduced through co-feeding in Ae. albopictus larvae ( Malassigné et al., 2025).
Discussion
General interpretation of the results
This systematic review showed that several studies on Aedes, Anopheles, and Culex mosquitoes microbiota fungal diversity have been published around the world. Most of these studies have been conducted in countries such as Italy, the United States, Brazil, France, Kenya, France, and Thailand. Few studies have been conducted in Argentina, South Asia, India, Vietnam, and a few African countries (South Africa, Burkina Faso, Egypt, Gabon, Ghana, Madagascar). In countries such as the United States, Brazil, European countries (France, Italy), and Kenya, most studies on mosquito microbiota are part of research for alternative and innovative tools for vector control. Research in this field of mosquito microbiota requires a certain medical entomology and microbiology expertise, research founds, and lab facilities that are not common in all countries. These factors could explain why mosquito fungal microbiota remains largely uncharacterized Africa and Asia despite the fact of the burden of vector-borne diseases in these countries.
During this systematic review, fungal genera, more than 41 (with more than 63 species), 19 (with 26 species), and 36 (with 54 species) belonging mainly to the phyla Ascomycota followed by Basidiomycota were detected in species of the genera Aedes, Anopheles, and Culex, respectively. In addition to the phyla Ascomycota and Basidiomycota, species of the genus Anopheles hosted a fungal genus ( Microsporidia) belonging to the phylum ( Microsporidiomycota). Species of the genus Aedes hosted more fungal genera, followed by species of Culex and species of Anopheles. This difference in fungal species among these vectors is partly due to their ecology. Although all mosquitoes share many biological traits, the preferred habitats are often specific to the different genera Aedes, Anopheles, and Culex. As cosmopolitan mosquito species, Aedes and Culex have preferred habitats that can be dark, stagnant water, such as puddles and water containers containing organic matter ( Ferede et al., 2018; Heu & Gendrin, 2018; Kahamba et al., 2020; Krol et al., 2024; Monteiro et al., 2019) that contribute to the proliferation of yeast-like fungi and certain molds. The different species of the genus Anopheles, on the other hand, have different aquatic habitats, which may be clear water exposed to sunlight, such as puddles and hoofprints left by animals, for some, and lakes, swamps, and rice fields for others ( Heu & Gendrin, 2018; WHO, 2022), resulting in a low presence of fungi.
Molecular biology-based approaches, whether or not combined with agar culture, and agar culture-based techniques have enabled the identification of different fungal species. The molecular biology-based technique enabled the identification of 46 fungal genera (more than 64 species), followed by 34 fungal genera (57 species) using the molecular biology technique combined with culture, and 6 fungal genera (10 species) using the culture-based technique alone. Due to their sensitivity and specificity, molecular biology-based methods (simple PCR, PCR followed by high-throughput sequencing or metagenomic sequencing) ( Flores et al., 2022; Guégan et al., 2020a; Hegde et al., 2024) offer the best performance in identifying fungal species. This technique allows the identification of non-cultivable species such as Microsporidia ( Akorli et al., 2024; Herren et al., 2020; Nattoh et al., 2021b; Onchuru et al., 2024), cultivable species, species present in low quantities in homogenates, and even species sensitive to environmental stress. In addition, molecular biology techniques combined with culture enable the identification of several fungal species, demonstrating that a large proportion of the fungal species in the mosquito microbiota are cultivable. The molecular biology method combined with culture appears to be effective because it ensures the viability of the identified fungal species ( Steyn et al., 2016). Few fungal species have been identified using the culture-dependent identification method alone.
In the larval stages, the Aedes, Anopheles, and Culex species harbored a fairly diverse fungal microbiota with 31 genera comprising 52 species belonging to eight phyla, mainly the phyla Ascomycota and Basidiomycota. Mostly yeast-like fungi (21 genera) with Candida, Meyerozyma, Pichia, Rhodotorula, and Wicherhamomyces, followed by molds (8 genera), including Penicillium and Aspergillus. Similarly, one genus of phytopathogenic fungi ( Gliocladium) and one genus of entomopathogenic fungi ( Paecilomyces) were associated with the larvae. These fungal communities associated with larval stages often varied between mosquito species and different larval environments (field/laboratory). Several studies have shown variation in fungal communities between different larval environments and between individuals coexisting in the same environment ( Shelomi, 2019; Tawidian et al., 2021). This diversity in the larval stages is thought to be due to the environment of the breeding sites. Female mosquitoes lay their eggs in breeding sites that vary according to the biotope and species ( Fontenille et al., 2017). After hatching, the eggs produce larvae and the mosquito larvae belonging to the Culicinae and Anophelinae subfamilies are aquatic and acquire part of their microbiota for feeding on organic particles contained in the water of their breeding sites, single-celled organisms (such as bacteria, fungi, and protozoa), and small invertebrates ( Fontenille et al., 2017; Hawkes & Hopkins, 2022; Heu & Gendrin, 2018). These organic particles contribute to the proliferation of fungi in mosquito larvae.
In addition, adults of the Aedes, Anopheles, and Culex species harbored a fungal microbiota that was as diverse in genus (33) belonging to six phyla, predominantly three phyla ( Ascomycota, Basidiomycota, Microsporidiomycota) with fewer species (45) than the larval stages. It appears that adult mosquitoes acquire part of their microbiota during emergence by absorbing water from larval habitats ( Malassigné et al., 2020; Steyn et al., 2016). However, as observed for bacteria, a significant reduction in fungal diversity is observed in newly emerged adults following the process of metamorphosis from nymph to adult ( Malassigné et al., 2020; Steyn et al., 2016). However, the fungal species consisted of yeasts (14 genera), phytopathogens (11 genera), molds (5 genera), entomopathogens (2 genera), and an obligate intracellular fungus that could vary depending on the origin of the mosquitoes (field/laboratory). Several fungal species, predominantly Candida, Aspergillus, Cladosporium, Penicillium, and Rhodotorula, were isolated from adults (males and females) kept under different breeding conditions in laboratories. The abundance of these fungal species in adults (males and females) could be linked to fungal recolonization after emergence, following reintroduction through feeding habits ( Flores et al., 2022), or through contaminants previously isolated in drinking water ( Novak et al., 2016). Also, as symbionts involved in nutrition in adult male Ae. albopictus ( Guégan et al., 2020a), Aspergillus and Cladosporium molds have been found in both sugar-fed male Cx. quinquefasciatus and blood- or sugar-fed females, suggesting a possible involvement in nutrition in adult (male and female) Cx. quinquefasciatus. Wild mosquitoes, on the other hand, have a greater fungal diversity than those kept in laboratory conditions. This fungal diversity in the microbiota is thought to be partly due to the nutritional behavior of wild mosquitoes. In addition to being hematophagous at one point in their life cycle, female mosquitoes of the Aedes, Culex, and Anopheles genera in the wild also feed on sugary juices (flower nectar, tree sap, fruit exudates) like male mosquitoes ( Fontenille et al., 2017; Hawkes & Hopkins, 2022; Heu & Gendrin, 2018). These mosquitoes prefer nectars rich in monosaccharides (glucose and fructose) and nectars rich in sucrose ( Guégan et al., 2022). Yeasts produced by the fermentation of sugar in nectars attract mosquitoes ( Luis et al., 2019), which also spread them throughout the environment. It turns out that filamentous fungi such as Cladosporium and Aspergillus are metabolically active in the digestion of fructose in male Ae. albopictus, and Malassezia yeast is metabolically active in the digestion of fructose in adult male and female Ae. albopictus ( Guégan et al., 2020b). Mosquitoes can also get infected through contact with fungi present in the environment, such as Microsporidia ( Nattoh et al., 2021b) and many other fungi.
In addition, digestive organs such as the intestines, crop, intestinal diverticulum, and mosquito species of the genera Aedes, Anopheles, and Culex harbor a diverse fungal community belonging to seven (7) phyla, with the phylum Ascomycota being the most prevalent, followed by Basidiomycota. These fungal species include yeasts, molds, phytopathogens, and an intracellular fungus. The presence of fungi (yeast, filamentous) in these digestive organs could be linked to nutrition. These organs play important roles in nutrition in general. In mosquitoes, the diverticulum is involved in storing sugar meals such as floral nectar before they slowly pass into the intestine to be digested ( Gusmão et al., 2007). As for the crop, it is involved in the selective storage of sugary solutions (nectar) before they are transferred to the mosquito's intestine ( Calkins et al., 2017; Guégan et al., 2020a), as well as the passage of protein-rich blood meals before they are transferred to the intestine in female mosquitoes. The intestine is involved in the digestion and absorption of sugar, which is used as a regular source of energy for mosquitoes, but also for the active intestinal microbiota, including fungi ( Malassigné et al., 2020).
Furthermore, certain species of fungi (yeasts, molds, phytopathogens) belonging to the phylum Ascomycota can colonize the male and female genital organs of certain mosquito species ( Ae. aegypti, Ae. albopictus, An. gambiae, An. stephensi, and Cx. quinquefasciatus). In addition, an obligate intracellular fungus belonging to the phylum Microsporidiomycota has been found in the male and female genital organs of An. arabiensis ( Nattoh et al., 2021b; Onchuru et al., 2024). The isolation of certain yeast species ( Wickerhamomyces anomalus, Candida parapsilosis) in the genital organs suggests a possibility of vertical transmission given their presence at all stages of development of certain mosquito species ( Bozic et al., 2017; Ricci et al., 2011; Ricci et al., 2011). Horizontal sexual transmission between adults is also possible with the Microsporidia MB species, given that it is isolated solely in the gonads ( Nattoh et al., 2021b). Other fungi found in the genitals, however, originated from the proliferation of environmental contaminants.
Finally, fungal species constitute a well resorce to support vector control ambition. They are able of inhibiting the development of the larval stages of certain species of Aedes, Anopheles, and Culex, or by inhibiting Plasmodium transmission or reducing the vectorial competence.
The inhibition of larval development may be due to competition between certain fungal species and the larvae for nutrients available in the water in the breeding sites, the production of toxins capable of inhibiting the larvae, and pollution of the water in the breeding sites.
As for inhibiting Plasmodium transmission or reducing the vector competence of Anopheles, one study has shown that the microbiota associated with mosquitoes can influence this by producing toxins and modulating the mosquito's immune system ( Malassigné et al., 2020). Indeed, the reduction in vector competence would be possible thanks to competition between the parasite and fungal species for nutrients available in the intestine or stomach, and the production of toxins that are often broad-spectrum antimicrobials (e.g., Wickerhamomyces anomalus) ( Cappelli et al., 2014). In addition, the reduction in vector competence may be due to an imbalance created by fungal species with microorganisms (bacteria and/or viruses and/or fungi) that promote the development of the parasite ( Plasmodium), or to the regulation of the immune system of mosquitoes ( Anopheles), leading to the activation of the main defense pathways, which reduces the survival and proliferation of the pathogen.
Limitations of the study
This study gathered and synthesized valuable information on Aedes, Anopheles and Culex mosquitoes fungi microbiota, but this contribution to the field do not exclude some has limitations of the study. First, articles were searched for in two databases (PubMed, Google Scholar) that are accessible free of charge without registration. Therefore, data on the fungal microbiota of species of the genera Aedes, Anopheles, and Culex remain insufficient. Second, for the creation of the geographical map, when the studies did not indicate the mosquito collection sites, we assigned the country of study ( Bozic et al., 2017; Cappelli et al., 2023).This geographical correction may be skewed by the absence of these mosquito species in these countries. Finally, the studies included have methodological limitations, with significant differences in sample collection, microbiological analysis, contamination that may originate from the environment, and the sometimes-small sample size.
In addition, the review process was limited by the exclusion of certain articles when the authors were not accessible. These missing data may introduce a selection bias. However, the exclusion of these articles does not impact the overall conclusion of our review. We selected only articles published in English. We may have omitted recent articles in other languages.
Prospects
** Implications for vector control practice **
Certain fungal species isolated from mosquito microbiota are able to affect larval stages of Anopheles, Aedes, and Culex mosquitoes and may help inhibit the development of Plasmodium pathogens. These fungal species can be used as a complement to chemical control methods, such as entomopathogenic fungi ( Beauveria, Metarhizium). In addition, the results identified a symbiotic yeast, Wickerhamomyces anomalus, an intracellular fungus like Microsporidia MB, which can block parasite transmission. Furthermore, given the fact that the fungal microbiota community in larval habitats has an impact on mosquito development, the destruction of organic matter and waste could contribute to reducing mosquito density. A good understanding of the ecological role of fungi in the mosquito microbiota will enable targeted use against specific mosquito species and reduce resistance.
** Implications for public health policy **
The results show that fungal microbiota can be integrated into national vector control programs. In integrated control, fungal symbionts can be used in the same way as insecticides and mosquito nets. However, the use of these symbionts in the environment requires support in the form of biosafety policies, impact assessment experts, and regulatory texts. Given the variation in fungal species depending on mosquito species, geographical locations, and ecological conditions, local regulations must be followed to ensure the proper use of fungal microbiota.
** Implications for future research **
The results obtained in this literature review could inspired future research direction. For example, it could be of interest to identifying all fungi in the mosquito microbiota that can further inhibit the development of larval stages or pathogens, to the interactions between mosquitoes, pathogens, and fungi and the associations that may lead to a greater antivector effect between fungi, bacteria, and viruses in the mosquito microbiota. In addition, the results could be of interest to develop tools for manipulating the fungal microbiota through biotechnology, paratransgenesis, and the introduction of competitive or genetically modified fungi, or to understand the interactions between mosquitoes, their fungal microbiota, and their environment.
Conclusion
This study provides an overview of the published literature on the Aedes, Anopheles, and Culex mosquitoes fungal microbiota and the prospects for their use as biological control tools against vector-borne diseases. Thus, most studies on the fungal microbiota of mosquitoes of the genera Aedes, Anopheles, and Culex have been conducted in Italy, the United States, Brazil, Kenya, France, and Thailand. Species of the genus Aedes harbored more fungal genera, followed by Culex and Anopheles. These species generally belonged to the phyla Ascomycota and Basidiomycota, with the phylum Microsporidiomycota specific to the genus Anopheles. The culture of homogenates on agar media allowed the isolation of certain fungal species, especially filamentous fungi. The cultures of homogenates, coupled or not with molecular biology techniques (PCR followed by high-throughput sequencing), allowed for better results in the identification of fungal species. Mostly yeasts of the genus ( Candida, Meyerozyma, Pichia, Rhodotorula, and Wicherhamomyces), molds ( Penicillium, Aspergillus), an entomopathogen ( Paecilomyces), and phytopathogens ( Gliocladium) were identified in mosquitoes at the larval stage. In the adult stages, yeasts of the genus ( Candida, Rhodotorula) and molds ( Cladosporium, Penicillium, Aspergillus) were found to be dominant in mosquitoes. Specifically, Microsporidia MB were found in some species of the genus Anopheles. In the digestive organs and certain genital organs, in addition to yeasts, molds, and phytopathogens found in species of the genus Aedes and Culex, a specific fungus, Microsporidia MB, was found specifically in certain species of Anopheles. Transovarian transmission and horizontal sexual transmission are thought to be possible specifically through yeasts ( Wickerhamomyces anomalus, Candida parapsilosis) and an intracellular fungus ( Microsporidia MB). Finally, certain fungal species have been found to infect the larval stages of certain species of Aedes, Anopheles, and Culex, while other fungal species ( Wickerhamomyces anomalus, Microsporidia MB) have been found to reduce the vector capacity of the Anopheles genus.
Registration and protocol
This study could not be registered in the PROSPERO International Prospective Register of Systematic Reviews.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akorli EA Andoh NE Egyirifa RK : Mosquito breeding water parameters are important determinants for Microsporidia MB in the aquatic stages of Anopheles species. Parasit Vectors. 2024;17(1): 509. 10.1186/s 13071-024-06596-9 39695866 PMC 11657153 · doi ↗ · pubmed ↗
- 2Angleró-Rodríguez YI Blumberg BJ Dong Y : A natural Anopheles-associated Penicillium chrysogenum enhances mosquito susceptibility to Plasmodium infection. Sci Rep. 2016;6: 34084. 10.1038/srep 34084 27678168 PMC 5039729 · doi ↗ · pubmed ↗
- 3Angleró-Rodríguez YI Talyuli OA Blumberg BJ : An Aedes aegypti-associated fungus increases susceptibility to dengue virus by modulating gut trypsin activity. e Life. 2017;6: e 28844. 10.7554/e Life.28844 29205153 PMC 5716662 · doi ↗ · pubmed ↗
- 4Bando H Okado K Guelbeogo WM : Intra-specific diversity of Serratia marcescens in Anopheles mosquito midgut defines Plasmodium transmission capacity. Sci Rep. 2013;3: 1641. 10.1038/srep 01641 23571408 PMC 3622076 · doi ↗ · pubmed ↗
- 5Barnard DR Xue RD Rotstein MA : Microsporidiosis (Microsporidia: Culicosporidae) alters blood-feeding responses and DEET repellency in Aedes aegypti (Diptera: Culicidae). J Med Entomol. 2007;44(6):1040–1046. 18047204 10.1603/0022-2585(2007)44[1040:mmcabr]2.0.co;2 · doi ↗ · pubmed ↗
- 6Batson J Dudas G Haas-Stapleton E : Single mosquito metatranscriptomics identifies vectors, emerging pathogens and reservoirs in one assay. e Life. 2021;10: e 68353. 10.7554/e Life.68353 33904402 PMC 8110308 · doi ↗ · pubmed ↗
- 7Bian G Joshi D Dong Y : Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science. 2013;340(6133):748–751. 10.1126/science.1236192 23661760 · doi ↗ · pubmed ↗
- 8Bilgo E Lovett B St Leger RJ : Native entomopathogenic Metarhizium spp. from Burkina Faso and their virulence against the malaria vector Anopheles coluzzii and non-target insects. Parasit Vectors. 2018;11(1): 209. 10.1186/s 13071-018-2796-6 29587876 PMC 5870526 · doi ↗ · pubmed ↗