Bioactivity-guided analysis of Moringa olifera fractionated extracts for potential medical application
Nancy G. Banoub, Masarra M. Sakr, Mahmoud M. Tawfick, Shaimaa Fayez, Mohamed M. S. Farag, Khaled M. Aboshanab

TL;DR
This study explores the bioactive properties of Moringa oleifera extracts and identifies their potential for cancer and viral treatments.
Contribution
The study provides a detailed bioactivity-guided analysis of Moringa oleifera extracts and their gene expression effects.
Findings
Hexane fraction showed strong cytotoxicity against cancer cells with low toxicity to normal cells.
Butanol fraction exhibited potent antiviral activity against HAV and HSV strains.
Ethyl acetate fraction had the highest antioxidant activity.
Abstract
Moringa oleifera, a nutrient-rich medicinal plant, has garnered growing scientific interest due to its diverse pharmacological properties. In this study, spectroscopic analysis, coupled with cytotoxic, antiviral, and antioxidant properties testing, was followed by gene expression analysis of various fractionated Moringa oleifera extracts to assess their potential biomedical applications. Ethanolic leaf extract was fractionated using solvents with increasing polarity (hexane, methylene chloride, ethyl acetate, and butanol). Cytotoxicity was tested against multiple cancer cell lines and normal cells, while antiviral activity was assessed against HAV ( Hepatitis A virus) and HSV-1/2 (Herpes simplex virus type 1/type 2). Antioxidant capacity was evaluated using DPPH assays. Gene expression of BAX and BCL-2 was analyzed by qRT-PCR to understand apoptotic pathways. Chemical profiling was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
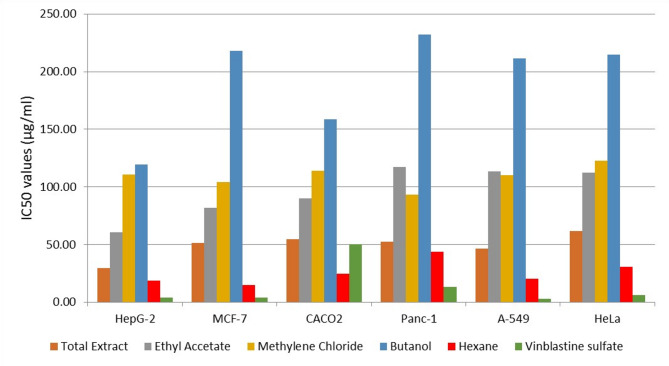 Figure 1
Figure 1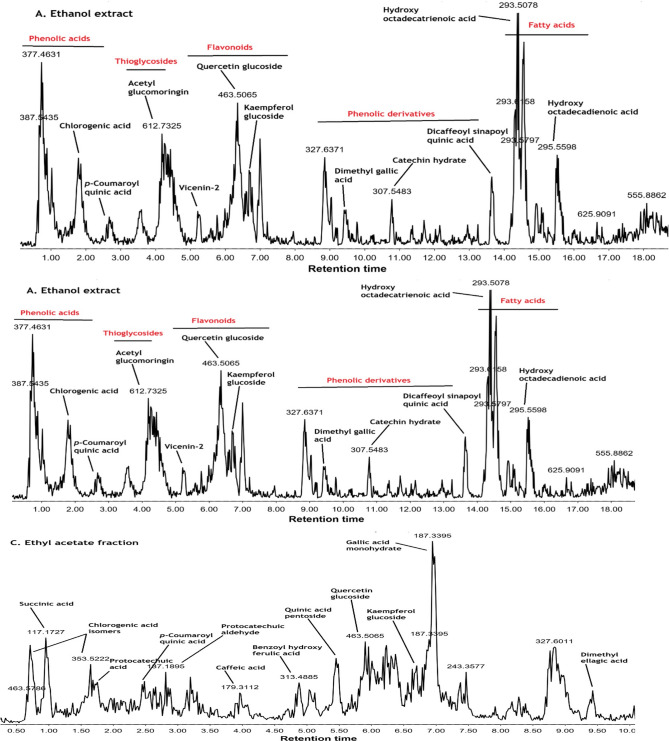 Figure 2
Figure 2- —Ain Shams University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMoringa oleifera research and applications · Metabolomics and Mass Spectrometry Studies · Seaweed-derived Bioactive Compounds
Introduction
Plants have long been considered effective medicinal agents due to their lower toxicity and fewer adverse effects than synthetic medications. Furthermore, phytochemical substances present in plants support a variety of complex cellular pathways^1,2^. Moringa oleifera, the “miracle tree”, is found across tropical and subtropical regions worldwide, even though Afghanistan, Bangladesh, India, and Pakistan are thought to be its original locations^3^. The Moringa family includes 13 species*,* of which Moringa oleifera is well known for its many purposes in medicine, nutrition, fertilizer, and biogas production^3^. According to reports, Moringa oleifera possesses several biological functions, including anti-inflammatory, anti-diabetic, hypotensive, and gastric ulcer-preventive effects^4^. It has also been demonstrated to control thyroid hormone levels and improve liver and kidney function**.** Additionally, Moringa oleifera leaves protect against liver damage, oxidative stress, cancer, and viral infections^5^. Fractions from the Moringa oleifera extract have been produced using various solvents, including ethanol, ethyl acetate, hexane, butanol, and methanol. These fractions have been observed to exhibit various properties, including antitumour, antioxidant, antiviral, immunomodulatory, antimicrobial, and radioprotective effects^6^. Moringa oleifera is known for its numerous bioactive compounds. The most often utilized components of the plant, the leaves, are notably rich in flavonoids, phenolic acids, vitamins, polyphenols, carotenoids, alkaloids, glucosinolates, isothiocyanates, tannins, and saponins, which are thought to be responsible for its diverse biological characteristics. Several in vitro and in vivo studies have confirmed these pharmacological properties^7^.
Moringa oleifera has shown antiviral properties against influenza and herpes simplex viruses by inhibiting viral replication and modulating immune responses^8^. It also exhibits anticancer effects, with research indicating that Moringa extracts can reduce tumor growth and stimulate apoptosis in numerous cancer cell lines^9^. Additionally, Moringa oleifera is a strong antioxidant that scavenges free radicals and reduces oxidative stress, thereby helping prevent chronic diseases^9^. Moreover, Moringa oleifera possesses immunomodulatory effects, enhancing immune function by promoting cytokine synthesis and strengthening the body’s defence mechanisms against infections and diseases^5^. Although previous studies have explored solvent-based extractions or examined the modulation of apoptotic markers such as BAX and BCL-2 in response to Moringa oleifera extracts, these investigations were often limited in scope, focusing on single bioactivities, fewer cell lines, or lacking detailed chemical characterization. The present study bridges these gaps by regarding how systematic fractionation using solvents of varying polarities can enrich specific metabolites and, consequently, modulate their biological efficacy. The novelty of this study lies in its comparative, bioactivity-guided approach. Rather than evaluating a single extract, we systematically fractionated the crude ethanolic extract using a sequence of increasing polarity: hexane, methylene chloride, ethyl acetate, and n-butanol. This allows for a precise mapping of the chemical constituents—identified via GC–MS and LC–MS/MS—against their specific cytotoxic and antiviral potencies. Therefore, this study aimed to isolate, extract, and fractionate ethanolic Moringa oleifera extract to identify bioactive compounds with potential in vitro biological activity. relevance. Moringa oleifera extract and various solvent fractions were examined for the antioxidant, cytotoxic, and antiviral properties. Furthermore, the antiviral activity of the most active fractions was assessed against specific immune cells. The study also assessed the expression levels of key apoptotic genes and identified and characterized the most active metabolites. The comparative evaluation of multiple solvent fractions derived from a single ethanolic extract, the distinct bioactivity profiles (cytotoxic, antiviral, antioxidant) of each fraction, and the association of these activities with dominant chemical classes, as revealed by LC–MS/MS and GC–MS analysis.
Materials and methods
Collection and processing of Moringa oleifera leaves
The Moringa oleifera plant material used in this study was collected from authenticated trees cultivated at the SEKEM Farm (Belbis, Sharkia Governorate, Egypt) in April 2023, Cairo, Egypt. The sample was received with an official Quality Control label documenting the sample name, number, and collection date, ensuring full traceability. The collection of plant material was established in compliance with the national guidelines. To further confirm the plant identity, the plant was kindly authenticated by Mrs Trease Labib, Senior Botanist at Mazhar Botanical Garden, Cairo, Egypt. A voucher specimen no. (PHG-P-MO-562) was deposited in the Herbarium of the Department of Pharmacognosy, Faculty of Pharmacy, Ain Shams University, Cairo, Egypt. Moringa oleifera leaves were extracted as previously described by Krishnamurthy et al.^10^. Approximately 900 g of fresh Moringa oleifera leaves were carefully washed under running tap water to get rid of dust and other debris. The leaves were then allowed to air-dry for two days at room temperature. The dried plant parts were coarsely ground in a grinder, yielding 218 g of powdered leaves.
Preparation of ethanolic Moringa oleifera extract and fractionation
The Moringa oleifera leaves powder was macerated with 1 L of 96% ethanol for 48 h at room temperature. The resultant mixture was filtered through filter paper, the filtrate was collected and evaporated under vacuum at 40 °C using a rotary evaporator, yielding 18 g of a dark green, oily extract. The extract was stored at 10 °C for further use^10^. The ethanolic extract was then subjected to sequential liquid–liquid fractionation using a series of solvents with increasing polarity: hexane, methylene chloride, ethyl acetate, and n-butanol. For each stage, the extract was partitioned with the respective solvent in a 1:1 (v/v) ratio, and the process was repeated three times to ensure exhaustive extraction of the bioactive constituents. Each resulting solvent fraction was evaporated under reduced pressure at 40°C and subsequently freeze-dried to obtain stable, dry residues. A rotary evaporator was used to dry the hexane, ethyl acetate, methylene chloride, and butanol fractions under vacuum at 40 °C before they were freeze-dried. The ethanolic Moringa oleifera extract and its solvent fractions were then utilized for experimental studies and referred to as tested samples^11^.
Cell lines and cell culture techniques
Normal human primary peripheral blood mononuclear cells (PBMCs) (ATCC® PCS-800011™) and normal animal Vero cells derived from the kidney of African green monkey (ATCC® CCL- 81™) were purchased from Vascera, Cairo, Egypt. The cancer cell lines A-549 (lung carcinoma), Panc-1 (pancreatic carcinoma), Caco-2 (intestinal carcinoma), HELA (cervical carcinoma), HepG-2 (human hepatocellular carcinoma), and MCF-7 (human breast carcinoma) were also acquired from Vascera, Cairo, Egypt. Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen, Carlsbad, CA, USA) containing 10% heat-inactivated fetal bovine serum (Hyclone, Marlborough, MA, USA), 1% L-glutamine (Sigma-Aldrish St. Louis, MO, USA), HEPES buffer and 50 µg/mL gentamicin (Grand Island, NY, USA) was used to maintain the MCF-7, HepG-2, A-549, Panc-1, Caco-2, and HELA cancer cell lines^12^. The Roswell Park Memorial Institute (RPMI) medium (Lonza, Germany) supplemented with 2% serum was used to cultivate Vero cells and PBMCs. In the assays performed in the current study, the cells were incubated at 37 °C in a humidified atmosphere with 5% CO_2_, and the cultures were subcultured twice weekly.
Cytotoxicity evaluation
The cytotoxicity of the tested samples was evaluated against PBMCs and Vero cells using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, as described by El Sayed et al.^13^, where absorbance was finally measured at 570 nm by a microplate reader (SunRise, TECAN, Inc., USA). Untreated cells were used as negative controls, while cells treated with cisplatin (Sigma-Aldrich, Darmstadt, Germany) served as positive controls^13^. To ensure reproducibility, all experiments were performed in triplicate. The cytotoxic effects of the tested samples were evaluated against several cancer cell lines, including MCF-7, HepG-2, Caco-2, A-549, and HeLa. Cytotoxicity was assessed using the crystal violet staining assay, as previously described by Feoktistova et al.^14^. Cells were incubated with the tested samples for 24 h at 37 °C in a humidified atmosphere containing 5% CO₂^14^. Cell viability was then measured by a colourimetric method, with absorbance of each well recorded at 570 nm using a microplate reader (TECAN, Inc.). Vinblastine sulfate-treated cells were used as positive controls, whereas untreated cells served as negative controls^14^. All experiments were performed in triplicate to ensure reproducibility.
Evaluation of the antiviral activity
Virus and cell culture
The antiviral activity of the tested samples was evaluated against Hepatitis A virus (HAV HM175 strain), Herpes simplex virus type 1 (HSV-1, GHSV-UL46 strain), and Herpes simplex virus type 2 (HSV). The viral strains were purchased from Nawah Scientific (Mokattam, Egypt) and propagated at the Regional Center for Mycology and Biotechnology (RCMB) et al.-Azhar University, Cairo, Egypt.
Virus propagation
Vero cells were seeded in cell culture flasks one day before the virus strain was propagated and assayed in confluent Vero cells^14^. Using the Spearman-Karber method, infectious viruses were quantified by calculating the 50% tissue culture infectious dose (TCID50) in eight wells per dilution, with 20 µL of inoculum per well^15^.
Antiviral activity evaluation
A cytopathic effect (CPE) inhibition assay, which measures the ability of the tested sample to inhibit virus-induced cytopathic effects in susceptible mammalian cells. A cytopathic effect inhibition assay was used to evaluate the antiviral activity of the tested samples. The assay was performed using the MTT method^16^. For each tested sample, Vero cell monolayers were added to 96-well plates at a density of 2 × 10^5^ cells/well and incubated for 24 h at 37 °C in a humidified atmosphere containing 5% CO₂. The monolayers were washed, and a standardized viral titer of 10^4^ TCID50 per well was used to challenge the cells (HAV, HSV-1, or HSV-2). The infected cells were then overlaid with RPMI medium containing 2% serum (100 µL) and various concentrations (0—1000 µg/mL) of the tested sample. The plates were incubated at 37 °C for 48 h. Infection controls (virus-infected cells without treatment) and untreated Vero cell controls (uninfected and untreated) were included as negative controls, while acyclovir was used as a positive control. After the incubation period, the MTT assay was used to assess cell viability^16^. The antiviral activity was calculated based on the ability of the tested sample to inhibit viral cytopathic effects. All experiments were performed in triplicate, and GraphPad Prism (GraphPad Software, San Diego, CA, USA) was used to determine the 50% inhibitory concentration (IC₅₀) and 50% cytotoxic concentration (CC₅₀). As described by Dogan et al.^17^, the selectivity index (SI) was calculated using the following formula: selectivity index (SI) = CC50/IC50^17^
Antioxidant activity assay
DPPH radical scavenging activity
To evaluate the antioxidant property of the tested samples, the 2,2-diphenyl1-1-picrylhydrazyl (DPPH) free radical scavenging assay was performed according to the method described by Lu et al*.*^18^. The examined samples were tested using a two-fold serial dilution ranging from 1000 µg/mL down to 0 µg/mL. Similarly, ascorbic acid, used as a positive control, was subjected to the same two-fold serial dilution to accurately determine its IC₅₀ value. The negative control consisted of the DPPH radical solution (0.004% w/v in methanol) without any antioxidant. All experiments were performed in triplicate. The % inhibition (PI) of the DPPH radical was calculated as previously reported^19^. The IC50, or sample concentration needed to block half of the free radicals, was calculated^12^.
Quantitative real-time PCR (qRT-PCR)
The qRT-PCR analysis was conducted to investigate the effect of ethanolic extract, ethyl acetate fraction, and hexane fractions on the expression of BCL-2 and BAX apoptotic genes in cancer cells (liver carcinoma, breast carcinoma, and pancreatic carcinoma cells)^20^. RNA extraction was performed according to the manufacturer’s instructions for the RNeasy Mini Kit from Qiagen (Catalogue no.74104). The PCR reaction was prepared using the Quantitect SYBR Green PCR Kit, as directed by the manufacturer, with a total reaction volume of 25 μL. Table 1 summarises the primer sequences used in this investigation.Table 1PCR oligonucleotide primers sequences.GenePrimer sequence (5’-3’)Expected Amplicon SizesReferenceGAPDH (Glyceraldehyde-3-Phosphate-Dehydrogenase)CTCTGATTTGGTCGTATTGGG150 bp^33^TGGAAGATGGTGATGGGATTBAX (BCL-2-Associated x gene)GGTTGTCGCCCTTTTCTA116bp^47^CGGAGGAAGTCCAATGTCBCL2 (B-Cell Lymphoma 2)GATGTGATGCCTCTGCGAAG128bpCATGCTGATGTCTCTGGAATCT
Cycling conditions for SYBR Green real-time PCR
After 30 min of reverse transcription at 50 °C, 94 °C was used for 15 min of initial denaturation. There were 40 cycles in the amplification cycle: 15 s of denaturation at 94 °C, 30 s of annealing (58 °C for GAPDH, 50 °C for BAX, and 64 °C for BCL-2), and 30 s of extension at 72 °C^20^. A dissociation curve analysis was performed in sequential steps: secondary denaturation at 94 °C for 1 min, annealing at the gene-specific temperature for 1 min, and final denaturation at 94 °C for 1 min.
Analysis of the SYBR Green RT-PCR
Amplification curves and cycle threshold (CT) values were calculated using the Stratagene MX3005P software. Gene expression levels were analyzed using the ^ΔΔ^CT method^20^. The relative fold change in gene expression was calculated using the formula 2^(-^ΔΔ^CT), with GAPDH serving as a housekeeping gene for normalization. All experiments were performed in triplicate, and the fold change was calculated using the average CT values. The ΔΔCT values were calculated as previously described^20^.
LC–MS/MS analysis of the ethanolic moringa oleifera extract, butanol fraction, and ethyl acetate fraction
The analysis of the ethanolic moringa oleifera extract, butanol fraction, and ethyl acetate fraction was performed using LC–ESI–MS/MS with a UPLC apparatus with a reversed-phase column ( ACQUITY UPLC BEH C18 column, 1.7 µm particle size, 2.1 × 50 mm column) coupled to a Xevo TQD triple quadrupole system (Waters Corporation, Milford, MA01757, USA) with an electroscopy ESI source (ESI voltage = 3.0 kV; nitrogen as sheath gas; capillary temperature 440 °C)^21^. Chromatographic separation was performed at a flow rate of 0.2 mL/min using gradient elution (A: H2O containing 0.1% formic acid, B: Acetonitrile containing 0.1% formic acid). Both positive and negative modes of mass spectra were acquired (scan range: m/z 100–1000) at a collision-induced dissociation (CID) energy of 20 eV^22^. The spectra were analyzed using the MassLynx 4.1 software^23^.
Gas chromatography/mass spectroscopy (GC–MS) analysis
Hexane fraction of Moringa oleifera extract was analyzed using a Shimadzu GC–MS-QP 2010 (Kyoto, Japan) with a Rxi-1MS fused bonded column ( 30m × 0.25 mm i.d., 0.25 × 0.25 μm film thickness, Restek, USA) fitted with a split-splittles injector^24^. The diluted samples (1% v/v) were injected in a split mode with a split ratio of 1:30. This ratio was specifically chosen to prevent column overloading and to ensure sharp, well-resolved peaks for both major and minor constituents in the extract. After 3 min at the initial column temperature of 50 °C, it was programmed to reach 300 °C at 5 °C/min; it remained at 300 °C for 10 min (isothermal). The injector’s temperature was set at 280 °C. 1.37mL/min was maintained as the helium carrier gas flow rate. The following parameters were used to record mass spectra: ion source temperature (220 °C), filament emission current (60 mA), and ionization voltage (70 eV), and the diluted samples (1% v/v) were injected as previously described^25^.
Statistical analysis
Unless otherwise noted, the mean ± standard deviation (SD) of at least three independent replicates is used to express descriptive results for all experiments. For statistical analysis, GraphPad Prism (GraphPad Software, San Diego, CA, USA) was used. Nonlinear regression analysis was used to determine the 50% inhibitory concentration (IC₅₀) values, which were used to evaluate cytotoxicity. The selectivity index (SI) was calculated from dose–response curves to evaluate antiviral activity. Fold changes were calculated using the ^ΔΔ^CT technique to evaluate gene expression changes. All experiments were conducted in triplicate to guarantee reproducibility, and the data were normalized to appropriate controls as described in the respective sections.
Results
Cytotoxic activity against PBMCs and Vero cells
Cytotoxic effects of the tested samples were evaluated against PBMCs and Vero cells. The IC50 values of PBMCs were determined as follows: ethanolic Moringa oleifera extract (624.31 ± 29.07 µg/mL), ethyl acetate fraction (77.76 ± 2.1 µg/mL), butanol fraction (126.92 ± 2.91 µg/mL), methylene chloride fraction (114.59 ± 3.47 µg/mL), and hexane fraction (93.27 ± 1.27 µg/mL). The IC50 values of Vero cells were determined as follows: ethanolic Moringa oleifera extract (647.4 ± 9.31µg/mL), ethyl acetate fraction (155.63 ± 2 µg/mL), butanol fraction (483.1 ± 9.2 µg/mL), methylene chloride fraction (327.96 ± 2.93 µg/mL), and hexane fraction (224.42 ± 5.18µg/mL). Cisplatin, utilized as a positive control, exhibited an IC50 value of 164.08 ± 0.78 µg/mL. These results indicated that all tested samples exhibited low cytotoxicity toward PBMCs and Vero cells. Among the solvent fractions, the ethyl acetate fraction showed the highest cytotoxic activity; however, this value remains well above the NCI’s high-cytotoxicity threshold (IC50 < 20 µg/ml), indicating a safe profile, while the ethanolic Moringa oleifera extract exhibited the lowest cytotoxicity.
Cytotoxic activity against cancer cell lines
As depicted in the dose–response curves (Figure S1), and the comparative viability bar charts (Fig. 1), all Moringa oleifera fractions exhibited a concentration-dependent reduction in cell viability. Notably, the hexane fraction and the total ethanolic extract demonstrated a remarkable safety profile, with the highest IC₅₀ values in normal PBMCs, resulting in superior Selectivity Index (SI) values exceeding 10.0 across all tested cancer lines (Table 2). The hexane fraction also maintained a favorable selectivity window (SI > 2), with its highest selectivity observed against breast cancer cells (SI = 6.30).In contrast, the ethyl acetate, methylene chloride, and butanol fractions showed moderate to weak cytotoxicity, with SI values generally near or below 2.0, indicating lower specificity for cancer cells. While the reference drug, Vinblastine Sulfate, displayed the highest absolute potency IC₅₀ for most cancer cell lines, its selectivity against several lines was comparable to or lower than that of the hexane fraction. These findings, supported by thedose-response curves in Figure S1, highlight the hexane fraction as the most selective and potent bioactive fraction derived from Moringa oleifera leaves.Fig. 1. Anticancer activities of the crude ethanolic Moringa oleifera extract and its fractions.Table 2. Cytotoxic activity (IC50) and selectivity indices (SI) of Moringa oleifera extracts and fractions against cancer cell lines compared to normal human PBMCs.Sample codePBMC IC50 (µg/mL)Cancer cell lineIC50 (µg/mL)SIEthanolic extract624.31 ± 29.07MCF-751.53 ± 2.0512.1HepG-229.68 ± 0.6321CACO254.82 ± 0.7811.4Panc-152.57 ± 2.1911.9A-54946.46 ± 1.7913.4HeLa61.90 ± 0.5310.1Ethyl Acetate fraction77.76 ± 2.1MCF-781.93 ± 3.980.9HepG-260.71 ± 0.421.3CACO289.88 ± 2.280.8Panc-1117.13 ± 2.520.6A-549113.51 ± 4.090.7HeLa112.32 ± 1.890.7Methylene chloride fraction114.59 ± 3.47 µg/mLMCF-7104.11 ± 1.911.1HepG-2110.87 ± 1.911CACO2113.80 ± 1.731Panc-193.00 ± 1.271.2A-549110.13 ± 4.771HeLa122.39 ± 0.970.9Butanol extract126.92 ± 2.91MCF-7218.11 ± 2.080.6HepG-2119.22 ± 2.121.1CACO2158.38 ± 5.900.8Panc-1231.92 ± 4.760.5A-549211.66 ± 10.290.6HeLa214.67 ± 4.180.6Hexane extract93.27 ± 1.27 µg/mLMCF-714.81 ± 0.326.3HepG-218.34 ± 0.765.1CACO224.78 ± 1.043.8Panc-143.82 ± 2.412.1A-54920.48 ± 0.104.6HeLa30.44 ± 0.493.1Vinblastine Sulfateof 164.08 ± 0.78 µg/mLMCF-73.78 ± 0.544.89HepG-23.68 ± 0.175.02CACO250.3 ± 6.100.36Panc-113.27 ± 0.941.39A-5492.84 ± 0.326.51HeLa6.14 ± 0.923.01Data are expressed as Mean ± SD (n = 3). SI (Selectivity Index) = IC50of normal PBMCs / IC50 of cancer cells. SI > 2 indicates high selective toxicity toward cancer cell.
Cytopathic effect of the tested samples against Viral Cell Lines
Results displayed in Table 3 revealed promising replication-inhibitory activity and showed that the butanol fraction demonstrated the most potent antiviral activity against all three viruses, with a high SI value confirming that the viral inhibition occurs at concentrations well below the cytotoxic threshold. These values indicate a favorable safety margin for this initial in vitro screening. The ethyl acetate fraction showed significant antiviral activity, particularly against HSV-1 and HSV-2, with moderate SI values. The hexane fraction demonstrated moderate antiviral activity against HSV-1 and HSV-2 but was inactive against HAV. The ethanolic extract and methylene chloride fraction showed weak or no antiviral activity against all tested viruses. As expected, acyclovir, the positive control, exhibited the most effective antiviral activity with the highest SI.Table 3. Antiviral activity of the tested samples.VirusSampleIC50 (µg/mL)CC50 (µg/mL)SIHAVEthanolic extractWeak270.33 ± 11.04InactiveEthyl Acetate fraction51.23 ± 0.96201.16 ± 4.383.923Methylene chloride fractionWeak419.04 ± 12.36InactiveButanol fraction24.42 ± 1.09176.2 ± 4.317.21Hexane fractionWeak161.98 ± 2.53InactiveAcyclovir14.75 ± 0.3297.39 ± 3.4720.16HSV-1Ethanolic extract93.88 ± 2.85270.33 ± 11.042.88Ethyl Acetate fraction14.75 ± 0.33201.16 ± 4.3813.63Methylene chloride fractionWeak419.04 ± 12.36InactiveButanol fraction13.23 ± 0.33176.2 ± 4.3113.32Hexane fraction29.74 ± 0.41161.98 ± 2.535.45Acyclovir3.07 ± 0.15138.54 ± 1.8545.12HSV-2Ethanolic extractWeak270.33 ± 11.04InactiveEthyl Acetate fraction25.41 ± 1.81201.16 ± 4.387.19Methylene chloride fractionWeak419.04 ± 12.36InactiveButanol fraction15.15 ± 0.28176.2 ± 4.3111.63Hexane fraction47.37 ± 1.12161.98 ± 2.533.41Acyclovir6.95 ± 0.36138.54 ± 1.8519.93CC50 is the half-maximal cytotoxic concentration; IC50 is the half-maximal inhibitory concentration; and SI is the selectivity index, calculated as CC50 divided by IC50.
Antioxidant activity
Results in Table 4 show that the ethyl acetate fraction exhibited the most potent free radical scavenging activity among all the tested samples, indicating the presence of a highly effective antioxidant. The butanol fraction showed moderate antioxidant activity, while the ethanolic extract and hexane fraction showed weaker effects. The methylene chloride fraction exhibited the least antioxidant activity.Table 4. The IC50 and standard deviation values of the tested samples in the DPPH.SampleIC50 (µg/mL) DPPH scavenging %Ethanolic extract109.55 ± 1.88Ethyl Acetate fraction14.66 ± 0.38Methylene chloride fraction459.39 ± 14.77Butanol fraction60.77 ± 0.75Hexane fraction229.28 ± 2.10Ascorbic acid10.21 ± 0.77
Effect of ethanolic Moringa oleifera extract, ethyl acetate fraction and hexane fraction on the expression of BCL-2 and BAX genes in Cancer Cells
The effect of ethanolic Moringa oleifera extract, ethyl acetate fraction, and hexane fraction (1000 μg/mL) on the expression of the apoptosis-related genes BCL-2 and BAX was evaluated in liver carcinoma, breast carcinoma, and pancreatic carcinoma cells after 24 h of treatment with the tested sample. Real-time PCR was used to analyze gene expression levels and was normalized to the reference gene GAPDH. Table 5 demonstrates a huge increase in BAX gene expression, accompanied by a huge reduction in BCL-2 expression compared to untreated cells. Representative qPCR amplification plots and melting curves for the target and reference genes are provided in the Supplementary Materials (Figure S2) to confirm the specificity and efficiency of the amplification process.Table 5. Effect of ethanolic Moringa oleifera extract, ethyl acetate fraction, and hexane fraction on mRNA expression of apoptosis markers (BCL-2 and BAX).SampleGAPDHBAX**BCL2CTCTFold changeCTFold changeLiver carcinoma20.4322.95–20.68–Liver carcinoma + ethanolic extract20.8120.169.000522.040.5070Liver carcinoma + ethyl acetate fraction19.7219.835.314720.290.8011Liver carcinoma + hexane fraction19.5418.4412.295021.950.2238Breast carcinoma19.2521.89–20.14–Breast carcinoma + ethanolic extract18.4717.5911.471620.870.3511Breast carcinoma + ethyl acetate fraction19.6219.626.233321.220.6113Breast carcinoma + hexane fraction18.5817.2119.111323.560.0587Pancreatic carcinoma20.1123.14–21.71–Pancreatic carcinoma + ethanolic extract19.2420.134.407621.680.5586Pancreatic carcinoma + ethyl acetate fraction20.1822.102.158522.090.8066Pancreatic carcinoma + hexane fraction20.5620.528.397723.270.4633
LC–MS/MS analysis of the ethanolic moringa oleifera extract, butanol fraction, and ethyl acetate fraction
Qualitative analysis of the bioactive ethanolic Moringa oleifera extract butanol and ethyl acetate fractions, which displayed promising antimicrobial activities, using ESI–MS/MS, led to the putative identification of more than 20 metabolites belonging to flavonoids, phenolic acids, and fatty acids (Table 6). Despite the enrichment of Moringa seeds with glucosinolates (thioglycosides), only acetyl glucomoringin (peak 4) in the leaves was detected, with a deprotonated molecular ion peak at m/z 612 and a prominent fragment ion at m/z 97 (HSO4). Identification of compounds was based on analyzing their MS2 fragments and comparing them with those reported in the literature. Compared to the ethanolic extract (Fig. 2), the butanol fraction was enriched in some phenolic acids, such as p- p-coumaroyl quinic acids and chlorogenic, along with some flavonoids, specifically vicenin-2 and kaempferol acetyl glycoside (Fig. 2). On the other hand, thioglycosides, such as acetylglucosamine, were less intense and lacked fatty acids. The ethyl acetate fraction exhibited diversity in phenolic acids, including p- p-coumaroyl quinic acids, chlorogenic acid, protocatechuic acid, caffeic acid, and a ferulic acid derivative (Fig. 2). Some flavonoids, such as quercetin and kaempferol glucosides, were particularly enriched in the ethyl acetate fraction. The ethyl acetate fraction showed the absence of thioglycosides and fatty acids. The results of the chemical profiling revealed that phenolic acids and flavonoids are the primary phytochemical classes contributing to the pronounced antimicrobial activity observed in the fractions, rather than fatty acids or thioglycosides.Table 6LC–MS/MS-based identification of the main metabolites in Moringa oleifera leaves crude ethanolic extract and its fractions (butanol and ethyl acetate).Peak not_R_ (min)[M–H]^–^[M + H]^+^MS/MSAnnotationChemical classButanol fractionEthyl acetate fractionReference(s)11.68153–123, 109, 108, 91, 81, 80Protocatechuic acidPhenolic acids– + ^29,41^21.81353–191, 179, 161, 135(neo)Chlorogenic acid + + + ^35^32.50337–191, 163, 136, 119, 111p–Coumaroylquinic acid + + + ^29,35^44.25612–275, 148, 97, 75, 72Acetyl–glucomoringinThioglycosides(Glucosinolates) + –^31^54.88313–161, 137, 121Benzoylhydroxy ferulic acidPhenolic acid– + ^36^65.24593–487, 353, 188Vicenin–2Flavonoid + –^35^75.45324–235Quinic acid pentosidePhenolic acid– + ^36^86.35463–300, 283, 151Quercetin–O–hexosideFlavonoids + + + ^43^96.70447–284, 255, 227, 223, 211, 174, 151Kaempferol–O–hexoside + + + ^35^106.77187–187, 169, 125, 123, 97, 57Gallic acid monohydratePhenolic derivative– + + + ^43^116.94533–285, 284, 283, 211, 185, 165Kaempferol–malonylglucosideFlavonoids + + + ^48^127.00–332332, 314, 286, 256, 191, 177, 148, 91, 86Pentahydroxy–methoxy–flavone + + + ^49^138.82301–255, 205, 179, 151, 149, 121, 107, 83Ellagic acid or quercetinPhenolic derivative– + ^50,51^148.85327–193, 183, 137, 97, 57, 43Unidentified–– + –159.44329–272, 249Dimethyl–O–ellagic acidPhenolic derivative– + ^52^169.79431–145Unidentified– + ––179.85–181181, 163, 145, 139, 135, 121, 107, 93, 91, 69TheobromineAlkaloid––^53^1810.7307*–185, 125, 121Catechin hydrateFlavonoid––^36^1911.3431–311, 283, 222, 215, 113Vitexin + –^31,54,55^2012.2609–300, 301Rutin + –^29,35^2113.6721–622, 560, 415, 144, 125, 89Di–O–caffeoyl–O–sinapoyl–quinic acidPhenolic derivative + + ^56^2213.9489–297, 284, 171Kaempferol acetyl glycosideFlavonoid + + ^29,31,57^2314.3293*–275, 265, 235, 211, 183, 181, 131, 121, 81Hydroxy octa–decatrienoic AcidHydroxy fatty acids––^31,36^2414.3409–161, 135, 133, 127, 85Unidentified– + ––2514.5–277185, 161, 149, 135, 121, 107, 93, 79, 69, 43Hydroxy hepta–decapentaenoic acidHydroxy fatty acid––^36^2615.3327–312Caffeoyl tartaric acid hexosidePhenolic derivative– + ^28^2715.5295–295, 195, 183, 113Hydroxy octa–decadienoic acidHydroxy fatty acid––^36^2816.3621–149, 89, 57Unidentified– + ––2918.1555–555, 255, 225, 206, 153SQMG 16:0(sulfoquinovosyl monoacylglycerol)Sulfolipids + –^58,59^3026.1–391273, 227, 209, 183, 167, 149, 111Diethylhexylphthalate (DEHP)Phthalates– + ^30^* Masses that were only detected in the ethanol extract, yet present in traces or neglected amounts in the butanol and ethyl acetate fractions.Based on the relative abundance of mass ions in the fractions, they were given symbols + + + (highly abundant), + + , + , and – (not present).Fig. 2LC–MS chromatogram of the (A) ethanol, (B) butanol, and (C) ethyl acetate extracts of Moringa oleifera leaves in negative mode.
Phenolic acids
Many phenolic acids were detected in the leaves of Moringa oleifera, including protocatechuic and chlorogenic acids, along with other quinic, ferulic, and caffeic acid derivatives. Protocatechuic acid (peak 1) displayed an [M-H]- peak at m/z 153 and a base peak fragment at m/z 109, which is formed due to decarboxylation (− 44 Da). (neo)Chlorogenic acid is a caffeoylquinic acid with a deprotonated ion peak at m/z 353, indicating a fragmentation transition typical of caffeic acid, such as m/z 179 [M- H-C7H10O5]-, which loses CO_2_ to form an ion at m/z 135. A base peak fragment at m/z 191 [M-H-C9H6O3]- is indicative of a deprotonated quinic acid moiety. Peak 3 was eluted at 2.5 min and showed [M-H]- ion at m/z 337 and a base peak fragment at m/z 163, indicative of 3-p-coumaroylquinoc acid based on the hierarchical scheme reported by Clifford et al.^26^. Compound 14 (tR = 9.44 min) displayed [M-H]-at m/z 329 and a fragment ion at m/z 272 [M-H-2CH3-CO]-, suggesting a dimethyl-O-ellagic acid. Peaks 20 and 25 were identified as dicaffeoyl sinapoyl quinic acid and caffeoyl tartaric acid hexoside, respectively, based on the similarity of their mass fragments to those previously reported in the literature^27,28^
Flavonoids
Phenolic acids and flavonoids are the two main classes identified in the ethanolic extract of Moringa oleifera, the butanol fraction, and the ethyl acetate fraction. Among the identified flavonoids are vicenin-2 (Apigenin 6,8-di-C-glucoside), quercetin-O-hexoside, kaempferol-O-hexoside, kaempferol malonyl glucoside, pentahydroxy methoxyflavone, catechin hydrate, vitexin, rutin, and kaempferol acetyl glycoside. Two apigenin-C-glucosides were detected, including vicenin-2, which was identified based on bibliographic data^29^, and vitexin (peak 18, tR = 11.3 min) with characteristic fragments at m/z 311 [M-H-120]- and m/z 283 attributed to the aglycone part. Two quercetin derivatives were detected, including compound 8 (tR = 6.35 min) with [M-H]- ion at m/z 463, showing a neutral loss of hexose (-162Da ) and rutin (peak 19, tR = 12.2 min) with a base peak fragment attributed to quercetin aglycone. Three kaempferol derivatives were identified including peak 9 (tR = 6.70 min) with base peak fragment attributed to the loss of hexose, peak 10 (tR = 6.94 min) with base peak fragment formed after the loss of malonylglucoside (-249 Da), and peak 21 (tR = 13.9 min) with base peak fragment of kaempferol aglycone formed after the loss of acetyl glucoside.
Hydroxy fatty acids
Three hydroxy fatty acids, eluting at the end of the chromatogram, were identified, including hydroxyoctadecatrienoic acid (tR = 14.3 min), hydroxyheptadecapentaenoic acid (tR = 14.5 min), and hydroxyoctadecadienoic acid (tR = 15.5 min). Their fragmentation is based on the successive loss of one or more water molecules and decarboxylation steps.
GC–MS analysis of the hexane fraction of Moringa oleifera
More than 40 metabolites (representing nearly 45.79% of the total GC–MS chromatogram) were tentatively identified in the hexane fraction of Moringa oleifera based on matching their retention indices with those reported in the NIST database and the human metabolome database. They are classified into linear, cyclic, and aromatic hydrocarbons, as well as lactones, fatty acid esters, phthalate esters, and steroids (Table 7). Abundant components, based on their peak area, are shown in Figure S3 and include hexahydrofarnesyl acetone, palmitic acid ethyl ester, 2- 2-hexanecan-1-ol, 3,7,11,15-tetramethyl acetate, linolenic acid methyl ester, 4,8,12,16-tetramethylheptadecan-4-olide,1-dehydro-10-gingerdione, strophanthidol, 24-norursa-3,12-diene, and betulinaldehyde.Table 7GC–MS analysis of the hexane extract of Moringa oleifera leaves.Peak not_R_ (min)Area %AnnotationBase m/zKI_rep_KI_exp_Chemical classSI (%)Molecular formula115.90.672,4-Dimethylundecane85.1511851194Linear hydrocarbons84C_13_H_28_216.80.181-Hexylcyclohexane83.1512241227Cyclic hydrocarbons80C_12_H_24_317.10.83Methyltetralin118.2012461235Cyclic hydrocarbons88C_11_H_14_418.10.515-Ethyl-5-methyldecane113.2012291269Linear hydrocarbons75C_13_H_28_518.70.25(3-Methylcyclopentyl)benzene104.1512951289Aromatic hydrocarbons80C_12_H_16_618.80.162,4-Dimethyldodecane85.1512851293Linear hydrocarbons80C_14_H_30_719.80.12n*-Heptyl cyclohexane83.151328133079C_13_H_26_820.10.13n-Heptylbenzene92.1513381339Aromatic hydrocarbons76C_13_H_20_922.30.35cis-Geranylacetone43.0514201419Oxygenated monoterpenes81C_13_H_22_O1023.60.53Dihydroactinidiolide111.1514731471Lactones88C_11_H_16_O_2_1124.00.112,4-di-t-Butylphenol191.2015121484Phenols91C_14_H_22_O1224.20.137-(1,3-Dimethylbuta-1,3-dienyl)-1,6,6-trimethyl-3,8-dioxatricyclo[5.1.0.0(2,4)]octane85.1514731495Cyclic ethers75C_15_H_22_O_2_1328.70.152-Phenylundecane105.1516921678Aromatic hydrocarbons94C_17_H_28_1429.30.166-Phenyldodecane91.101719170679C_18_H_30_1529.40.11(1-Butylheptyl)benzene91.101724171084C_17_H_28_1630.20.203-Phenylundecane91.101724174476C_17_H_28_1730.70.21Myristic acid, ethyl ester88.1017791768Fatty acid esters84C_16_H_32_O_2_1831.20.183-Methyl-butyric acid, 4-butyl-2-methyl-5-oxotetrahydrofuran-3-yl ester85.2018071790Lactones79C_14_H_24_O_4_1931.50.206-Phenyldodecane91.1018231804Aromatic hydrocarbons82C_18_H_30_2031.70.185-Phenyldodecane91.101823181186C_18_H_30_2131.91.82Hexahydrofarnesyl acetone43.0518231820Oxygenated sesquiterpenes75C_18_H_36_O2232.50.272-Methyl-7-octadecyne82.1518631849Linear hydrocarbons75C_19_H_36_2332.90.332-Methyl-7-octadecyne95.151863186774C_19_H_36_2433.20.172-Phenyltridecane105.1518941885Aromatic hydrocarbons92C_19_H_32_2533.50.99Palmitic acid, methyl ester87.1018781898Fatty acid esters83C_17_H_34_O_2_2633.60.44Di-sec-butyl phthalate149.1019081904Phthalate esters91C_16_H_22_O_4_2734.70.499-Hexadecenoic acid, ethyl ester96.1519551956Fatty acid esters72C_18_H_34_O_2_2834.92.04Palmitic acid, ethyl ester88.151978196882C_18_H_36_O_2_2936.50.14Hexadecanoic acid, 4-hydroxy, γ-lactone85.1521042049Lactones81C_16_H_30_O_2_3036.80.47Linolenic acid, methyl ester79.1020812063Fatty acid esters84C_19_H_32_O_2_3136.90.29(Z)-6-Octadecenoic acid, methyl ester96.152085206872C_19_H_36_O_2_3237.32.282-Hexadecen-1-ol, 3,7,11,15-tetramethyl acetate123.202168208979C_22_H_42_O_2_3337.50.20Stearic acid, methyl ester87.102077209881C_19_H_38_O_2_3438.00.951,16-Hexadecanediol81.1021302128Fatty alcohol73C_16_H_34_O_2_3538.11.68Linolenic acid, methyl ester79.1021012132Fatty acid esters82C_19_H_32_O_2_3639.20.293,7,11,15-Tetramethylhexadec-2-en-1-yl acetate123.202168219277C_22_H_42_O_2_3741.32.574,8,12,16-Tetramethylheptadecan-4-olide99.1522582311Lactones83C_21_H_40_O_2_3841.60.32n-Propyl linolenate79.123002324Fatty acid esters81C_21_H_36_O_2_3944.31.81Tetracosane85.224072488Saturated alkanes75C_24_H_50_4044.43.12Phthalic acid, di(6-methylhept-2-yl) ester149.124752494Phthalate esters90C_24_H_38_O_4_4148.80.21Octacosane85.228042785Saturated alkanes71C_28_H_58_4248.90.18Squalene81.12814279576C_30_H_50_4349.311.211-Dehydro-10-gingerdione43.128322816Phenolic53C_21_H_30_O_4_4450.40.661,1,6-trimethyl-3-methylene-2-(3,6,9,13-tetramethyl-6-ethenye-10,14-dimethylene-pentadec-4-enyl)cyclohexane137.128852895Hydrocarbons72C_33_H_56_4555.83.70Strophanthidol107.132383294Steroids73C_23_H_34_O_6_4656.32.1124-Norursa-3,12-diene218.23105332884C_29_H_46_4757.01.77Betulinaldehyde95.13628337778C_30_H_48_O_2_(%) Total identified compounds45.79%* Reported on another non-polar column.
Discussion
Moringa oleifera leaves have captivated the scientific community due to their potential for in vitro biological applications. properties. These include immune system modulation, thyroid hormone regulation, protection against gastric ulcers, anti-diabetic activity, hypotensive properties, anti-inflammatory, antibacterial, and anticancer activities, as well as the ability to improve liver and kidney function^4,5^. This study provides a comparative, solvent-dependent evaluation of Moringa oleifera leaf fractions derived from a single ethanolic extract, integrating biological screening with comprehensive chemical profiling. Rather than focusing on isolated bioactivities, the results demonstrate that each solvent fraction exhibits a distinct biological profile, including cytotoxic, antiviral, and antioxidant effects. Importantly, these fraction-specific activities correlate with the dominant phytochemical classes identified by LC–MS/MS and GC–MS analyses, highlighting the critical role of solvent polarity in shaping both chemical composition and biological performance. Collectively, these findings support the value of targeted fractionation as an exploratory strategy to elucidate the multifunctional bioactivity of complex plant extracts under in vitro conditions.
The cytotoxicity of the tested samples was evaluated at concentrations ranging from 0 to 1000 µg/mL against a panel of cancer cell lines: CA-549, MCF-7, HepG-2, Panc-1, Caco-2, and HELA. The superior potential in vitro biological action of the hexane fraction exhibited the highest cytotoxic potency, particularly against MCF-7 and HepG-2, with comparatively low toxicity against PBMCs (IC₅₀ = 93.27 µg/mL), this high potency can be attributed to the lipophilic nature of the hexane fraction, which, as revealed by GC–MS analysis, is enriched with fatty acids and sterols (e.g., β-sitosterol). Quantitatively, our hexane fraction demonstrated a potent IC₅₀ = 14.81 µg/mL against MCF-7 cells. This result shows superior efficacy compared to the study by Al-Asmari et al. (2015), who reported that the hexane extract of Moringa oleifera leaves exhibited an IC₅₀ = 230 µg/mL against the same MCF-7 cell line. This significant 11-fold increase in potency in our findings can be attributed to the enhanced recovery of specific lipophilic terpenoids and the geographic variation in the Egyptian chemotype, which appears to yield a more concentrated profile of cytotoxic metabolites compared to the samples from different agro-ecological zones reported in earlier studies^30^.
These non-polar compounds possess high membrane permeability, allowing them to effectively penetrate the lipid bilayer of cancer cells and interact with intracellular targets^31,32^. This selectivity mirrors earlier reports of Moringa’s anticancer effects. For instance, Pop et al.^9^ documented apoptosis induction in cancer cells via modulation of BAX/BCL-2, consistent with our qRT-PCR data showing BAX upregulation (e.g., 19.1-fold in breast carcinoma) and BCL-2 downregulation (0.06-fold) after hexane fraction treatment. Notably, while the modulation of BAX and BCL-2 provides strong evidence of apoptosis at the mRNA level, the absence of protein-level validation (e.g., Western blot) is a study limitation that should be addressed in future work. Likewise, Krishnamurthy et al. reported significant cytotoxicity of the hexane fraction of Moringa oleifera against breast and liver cancer cell lines, indicating that lipophilic metabolites play a central role in cytotoxicity^10^. The induction of apoptosis, evidenced in our study by the upregulation of pro-apoptotic BAX and downregulation of anti-apoptotic BCL-2, concurs with findings from Golestani Eimani et al.^33^, who reported similar apoptotic gene modulation following treatment with flavonoid-rich plant extracts. Notably, the efficacy of the hexane fraction against HepG-2 and MCF-7 cells aligns with Talib et al*.,* who attributed Moringa’s anticancer activity to glucosinolates and flavonoids^6^. However, our GC–MS analysis identified hexahydrofarnesyl acetone (11.21%) and phytol derivatives (e.g., 2-hexadecen-1-o1,3,7,11,15–tetramethylhexadecyl acetate) as major constituents, suggesting that terpenoids and fatty acid esters contribute to its potency—a divergence from seed-focused studies that emphasize glucosinolates. Geographic differences may explain this: Nigerian Moringa oleifera hexane fractions prioritized phthalates^34^, while Egyptian samples included lactones (4,8,12,16-tetramethylheptadecan-4-olide) and steroids (strophanthidol). This highlights the impact of agro-ecological conditions on metabolite profiles and bioactivity.
In comparison, the ethanolic Moringa oleifera extract showed moderate, balanced activity across all cell lines. In contrast, the butanol fraction exhibited the weakest cytotoxic effect, which is consistent with its relatively low levels of flavonoids and fatty acids, as determined by LC–MS/MS analysis. This indicates that phenolic acids and flavonoids, which are more abundant in the hexane and ethyl acetate fractions, are largely responsible for cytotoxicity^35–37^.
A crucial aspect of any potential lead compound is its safety profile. In this study, we evaluated the Selectivity Index (SI) to benchmark the toxicity of the fractions against normal human PBMCs. The hexane fraction displayed a high SI (> 2) for both MCF-7 and HepG2 lines, suggesting a favorable window of selectivity. This broad-spectrum selectivity suggests that the lipophilic compounds in the hexane fraction target common oncogenic mechanisms prevalent in diverse solid tumors. Furthermore, the ethanolic extract demonstrated remarkably high SI values (ranging from 10.1 to 21.0), further reinforcing the therapeutic potential of the whole leaf matrix.
The antiviral activities of the ethanolic Moringa oleifera extract and its solvent fractions were tested in vitro against Hepatitis A virus HAV, HSV-1, and HSV-2. The butanol fraction demonstrated the most potent antiviral activity against HAV (IC₅₀ = 24.42 µg/mL, SI = 7.21), HSV-1/2 (IC₅₀ = 13.23–15.15 µg/mL, SI = 11.63–13.32), and all three viruses with a high SI value. To assess the safety of utilizing a compound as an antiviral agent, the selectivity index (SI) is a commonly used parameter. SI determines the difference between the cytotoxic and antiviral activities. Higher SI values indicate that drugs are more effective and safer to use^46^. To eliminate the virus at a concentration that would not damage host cells, the ideal medication would destroy the virus at low concentrations and the cells at high concentrations^38,39^. These values surpass the tested samples and approach the efficacy of the positive control, acyclovir (SI for HSV-1 = 45.12). These findings are consistent with those of Giugliano et al., who observed that Moringa oleifera leaf extract significantly inhibited the replication of respiratory and HSV-1 viruses and modulatedhe host immune response^8^. Regarding antiviral activity, our butanol fraction IC₅₀ = 13.23 µg/mL against HSV-1 showed significantly higher potency compared to the findings of Lipipun et al.^40^, who reported that polar extracts of Moringa oleifera exhibited an IC₅₀ 100 µg/mL against HSV-1. This difference arises from the solvent-specific enrichment of polar flavonoids (e.g., vicenin-2) in our butanol fraction, which are less concentrated in the crude polar extracts used in previous studie. While the SI values for Moringa oleifera fractions are significant, they remain preliminary. Unlike the reference drug Acyclovir, which showed an SI for HSV-1 = 45.12, our fractions represent a complex mixture of metabolites. Therefore, these results should be interpreted as a primary in vitro screening. Future studies are required to determine whether the observed effects are virucidal (extracellular) or involve inhibition of intracellular replication.
LC–MS/MS revealed that the butanol fraction was enriched in flavonoids (e.g., vicenin-2, kaempferol acetyl glycoside) and phenolic acids (chlorogenic acid), which are known to inhibit viral enzymes and modify cellular redox states^31,32^. As suggested by Dogan et al. in similar models, these polyphenols may enhance host cell antiviral defences by modulating the IFN-β response or interfering with viral glycoproteins^17^. Significant antiviral activity was also demonstrated by the ethyl acetate fraction, specifically against HSV-1 (IC₅₀ = 14.75 µg/mL, selectivity index [SI] = 13.63). This confirms previous studies that found quercetin derivatives as inhibitors of herpes simplex virus polymerase and helicase-primase complexes^41^. The ethanolic Moringa oleifera extract and methylene chloride fraction showed weak to no antiviral activity, indicating that bioactivity is not only phenol-dependent but also fraction-specific. In contrast, Vergara-Jimenez et al*.* highlighted the importance of targeted fractionation, broadly attributing Moringa’s antiviral properties to its phenolic antioxidants^5^
Compounds known as antioxidants protect cells from the damage caused by unstable molecules such as free radicals. The ethyl acetate fraction showed the highest antioxidant activity (IC₅₀ = 14.66 µg/mL) based on DPPH radical scavenging. This is much higher than the reference antioxidant ascorbic acid (IC₅₀ = 10.21 µg/mL). This aligns with a previous study that showed the phenolic and flavonoid-rich fractions of Moringa oleifera leaf extract to have high antioxidant potential^42^. These antioxidants play a critical role in decreasing oxidative stress, a known contributor to viral replication and cancer cell proliferation^5^. The methylene chloride fraction’s low activity (IC₅₀ = 459.39 µg/mL) suggests that non-polar solvents extract fewer antioxidants, which aligns with previous findings^41^. Moreover, the LC–MS/MS analysis performed in the current study identified flavonoids, including quercetin-O-hexoside and kaempferol glucosides, in the ethyl acetate fraction. These compounds are known for their potent ROS-scavenging activities and synergy with other polyphenols^43,44^. On the other hand, since the hexane fraction lacks these hydrogen-donating groups, its antioxidant capacity remains limited.
Apoptotic Gene Modulation: Ethanolic Moringa oleifera extract, ethyl acetate, and hexane fractions extensively modulated BAX/BCL-2 expression in liver, breast, and pancreatic carcinoma cells. The hexane fraction induced the most substantial pro-apoptotic change, 12.29-fold BAX upregulation in liver carcinoma, while Bcl resulted in only 0.223-fold downregulation, likely due to its terpenoid and steroid components (e.g., betulinaldehyde, 24-norursa-3,12-diene). These results are supported by Golestani Eimani et al*.*^33^, who reported that plant-derived flavonoids and fatty acids increase mitochondrial apoptosis by modulating BAX/BCL-2.
Interestingly, the ethyl acetate fraction exhibited a strong pro-apoptotic gene expression profile, particularly in breast cancer cells, despite also showing moderate cytotoxicity. It contains abundant flavonoids, such as quercetin glycosides and kaempferol, which are known to induce apoptosis via ROS-mediated signalling pathways^9^.
The LC–MS/MS and GC–MS profiles, which revealed over 60 metabolites across fractions, show good correlation with the observed bioactivities. Phenolic acids and flavonoids, which have been shown to exhibit multifunctional bioactivity, including the induction of apoptosis, antioxidant properties, and antiviral effects, were abundant in the ethyl acetate fraction^9,45^. In contrast, the hexane fraction contained non-polar compounds, such as phytosterols, terpenoids, and hydroxy fatty acids, which contributed to its cytotoxicity but weak antiviral and antioxidant properties. These findings highlight that fraction polarity directly affects the potential in vitro biological activity profile of plant extracts, and that targeted extraction can optimise bioactivity for specific medical applications. Previous GC–MS of the hexane extract of Moringa oleifera from Palestine showed an abundance of fatty acids, especially stearic and palmitic acids^37^. Another study from India revealed the richness of the dichloromethane extract of Moringa oleifera in palmitic acid and cis-vaccenic acid^24^. In contrast, Jamiu et al*.*reported on the abundance of 2-heptanol, 6-methyl-1,2-benzenedicarboxylic acid, phthalic acid, cyclobutyl-heptyl ester, diethylphthalate, ethylisopropyl ester, trimethyl(4-tert.-butyl-phenoxy)silane, and 2,4,6-cycloheptatrien-1-one in the hexane and ethyl acetate fractions from Nigeria^34^. Differences in metabolites of the same plant species could be attributed to geographical distribution and agricultural conditions.
In summary, while the present findings highlight the in vitro potential of moringa oleifera fractions, particularly the hexane fraction’s pro-apoptotic activity, these results should be interpreted as exploratory. The transition from in vitro observations to clinical application involves significant physiological complexities. Therefore, further studies, including in vivo models and pharmacokinetic evaluations, are required to evaluate their efficacy and safety in complex biological systems.
Conclusion
Based on solvent-specific phytochemical clusters, this study confirms the cytotoxic, antioxidant, and antiviral characteristics of Moringa oleifera ethanolic extract and its fractions. Potent selective cytotoxicity was highest in the hexane fraction. The strongest activity, particularly against HepG-2 and MCF-7, while showing minimal toxicity to normal PBMCs, underscores its potential in vitro biological functions. safety and suggests tumor-selective mechanisms probably mediated by terpenoids and fatty acid esters. This effect was further supported by the upregulation of pro-apoptotic BAX and downregulation of anti-apoptotic BCL-2, indicating induction of programmed cell death. The butanol fraction displayed remarkable antiviral effects against HAV and HSV-1/2, with a high selectivity index, whereas the ethyl acetate fraction showed the strongest antioxidant capacity, likely due to its high content of phenolic acids. Our findings, supported by the identification of over 60 bioactive compounds using LC–MS/MS and GC–MS, provide a scientific basis for further development of these fractions either into targeted phytopharmaceuticals or broader therapeutic applications were a standardized enriched extract that combines several fractions can provide a multitarget therapy.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klimek-Szczykutowicz, M. et al. Moringa oleifera (drumstick tree)—nutraceutical, cosmetological and medicinal importance: a review. Front Pharmacol 15, (2024).10.3389/fphar.2024.1288382 PMC 1086962438370483 · doi ↗ · pubmed ↗
- 2Feoktistova, M., Geserick, P. & Leverkus, M. Crystal Violet Assay for Determining Viability of Cultured Cells. Cold Spring Harb Protoc 2016, pdb.prot 087379 (2016).10.1101/pdb.prot 08737927037069 · doi ↗ · pubmed ↗
- 3Eltokhy, M. A. et al. Exploring the nature of the antimicrobial metabolites produced by Paenibacillus ehimensis soil isolate mz 921932 using a metagenomic nanopore sequencing coupled with lc-mass analysis. Antibiotics 11, (2022).10.3390/antibiotics 11010012 PMC 877306535052889 · doi ↗ · pubmed ↗
- 4Jamiu, A. M. et al. Nutritional Values and Effects of Selected Solvents on the Phytochemical Properties of Moringa Oleifera Leaves Obtained from Sokoto, Nigeria. in 2024 International Conference on Science, Engineering and Business for Driving Sustainable Development Goals (SEB 4SDG) 1–7 (IEEE, 2024). 10.1109/SEB 4SDG 60871.2024.10630192.