Biochemical, Molecular and Behavioral Changes Induced by High-fat and High-sugar Diets: A Systematic Review of Non-clinical Studies
Camila Bach, Julia Vicentin Souza, Cláudia Sirlene Oliveira, Rosiane Guetter Mello, Daniele Maria-Ferreira

TL;DR
This review explores how high-fat and high-sugar diets affect the body and brain, leading to harmful biochemical, molecular, and behavioral changes.
Contribution
The study systematically reviews non-clinical evidence linking high-fat/high-sugar diets to neural and physiological changes.
Findings
High-fat and high-sugar diets induce physiological changes in the peripheral and central nervous systems.
These diets are associated with changes in gut microbiota, glucose metabolism, and bile acid signaling.
Adverse effects include neuronal and structural adaptations, suggesting a need for further research on prevention strategies.
Abstract
Diets that promote excessive caloric intake, particularly those high in fat and/or sugar, can cause harmful changes in the human body, including the brain. Excess consumption of fat and sugar may impair neuronal function and have both short- and long-term adverse effects. This systematic review examined the influence of high-fat and/or high-sugar dietary patterns on biochemical, molecular, and behavioral changes (PROSPERO protocol number: CRD42024526471). A systematic search was conducted in PubMed, EMBASE, and Scopus, and twenty-eight articles were included for data extraction. The studies indicate that obesogenic and energy-dense dietary patterns induce physiological changes in both the peripheral and central nervous systems, leading to neuronal, functional, and structural adaptations accompanied by biochemical, molecular, and behavioral changes. These adaptations appear to be…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
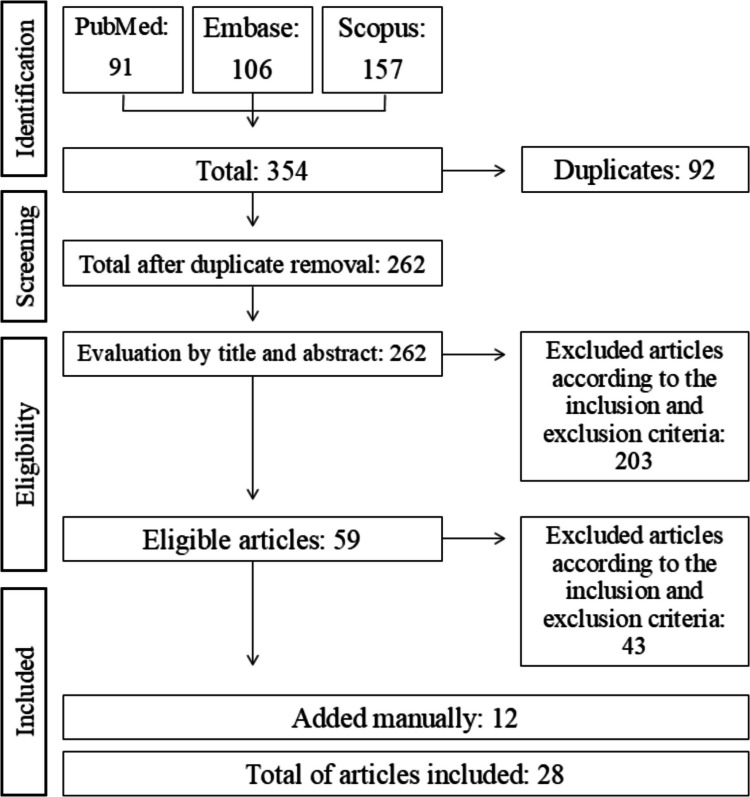 Figure 1
Figure 1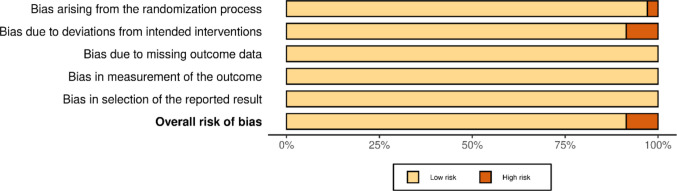 Figure 2
Figure 2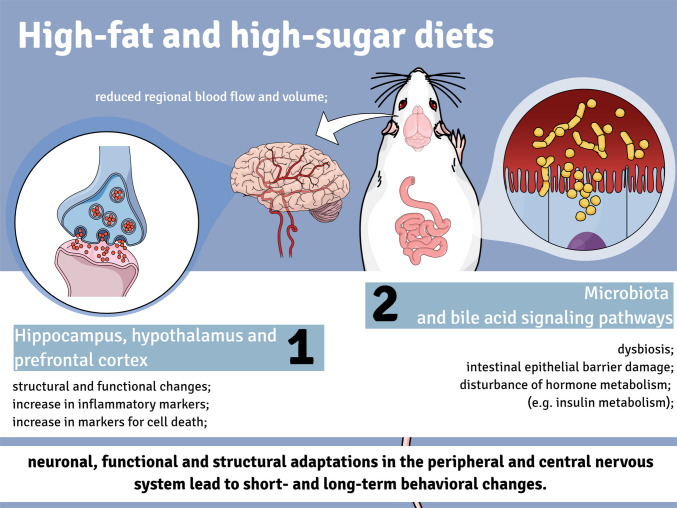 Figure 3
Figure 3- —Faculdades Pequeno Príncipe
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiet and metabolism studies · Regulation of Appetite and Obesity · Diet, Metabolism, and Disease
Introduction
The brain is the most complex organ of the human body and accounts for about 2% of the total body weight. It has a considerable and constant energy requirement and consumes about 20% of the total energy from nutrients [1–3]. An adequate intake of macro- and micronutrients is essential for efficient neuronal function, synaptic activity, and the synthesis of neurotransmitters and protein components [4, 5].
A growing body of neuroscientific evidence shows that adult mammal brains can undergo structural and functional remodeling in response to learning or experience [6, 7]. This process enables adaptive plasticity in mammalian brains, supporting learning, memory formation, and recovery of neural circuitry after injury, as demonstrated in experimental animal models [8]. The neurophysiological processes associated with these events range from rapid adjustments in synaptic strength, which are transient in nature, to long-term structural changes [9]. Important contributing factors are the formation of new synaptic connections, new neurons, new blood vessels, and non-neuronal glial cells in the brain [10]. The extent of neuronal remodeling that is triggered in response to learning or experience depends in part on factors that influence brain activity and the predisposition to trigger that activity. These factors include the quality of sleep, physical activity, and diet, which are important pillars of mental health due to their effects on the structure and function of the brain [11].
In fact, it has long been recognized that regular physical activity combined with adequate sleep and a good diet is essential for a healthy lifestyle and the prevention of various health problems [11]. Diet, in particular, is a recognized lifestyle factor that can influence brain structure and function and has the potential to alter predisposition to neuronal remodeling [4, 5, 12]. In 2018, a cross-sectional study showed that consumption of raw fruits and vegetables predicted a reduction in depressive symptoms and an increase in positive mood and life satisfaction [13]. In 2020, another cross-sectional study confirmed that the consumption of raw fruits and vegetables in addition to adequate sleep and physical activity improved depressive symptoms and well-being in young adults [11]. Conversely, recent evidence suggests that a diet high in sugar and saturated fat can significantly affect key elements of both neuronal and behavioral plasticity [14, 15], supporting the notion that dietary changes can have deleterious effects on the brain [16].
Despite the wealth of information found in the literature, the role of a high-fat and/or high-sugar diet on biochemical, molecular, and behavioral changes in non-clinical studies is still not fully understood and has not been systematically organized. Therefore, the aim of this review is to systematically present and critically evaluate recent advances in scientific literature regarding the effects of a high-fat and/or high-sugar diet on these parameters. This review provides a comprehensive discussion of the animal models used in these studies, as well as an in-depth analysis of the study designs, behavioral outcomes, and associated biochemical and molecular changes.
Methods
The present study followed the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guideline for systematic reviews and meta-analysis [17] and its protocol was registered in the International Prospective Register of Systematic Reviews (PROSPERO), under the number CRD42024526471.
Research Question
To formulate the guiding research question, the acronym PICOS [18] was used, where “P” denotes the problem, “I” the intervention, “C” the comparison, “O” the outcome, and “S” the study. The following strategy was created for this study: P: rodents exposed to different diets (high sugar and/or high fat),I: different diets (high sugar and/or high fat); C: animals fed standard rodent chow; O: alterations in biochemical, molecular, or behavioral markers; S: Animal studies.
Search Strategy
A comprehensive literature search was carried out in the PubMed, EMBASE, and Scopus databases. The articles included in this review cover the period between 2010 and 2023. The search terms considered in this review included “neuroplasticity" and “diet”. The search approach was adapted for each database as needed.
Inclusion and Exclusion Criteria
The search focused on original non-clinical studies related to the influence of diet on rodents’ biochemical and molecular alterations in the brain, as well as potential behavioral alterations. It was limited to one publication period (2010 to 2023) and articles published in English. Studies in experimental rodent models (rats and mice) exposed to different diet types (high-fat, high-fat/high-sugar, cafeteria, or high-sugar diets) were included. Only studies with primary (i.e., biochemical and/or molecular assays) and, if possible, secondary (e.g. behavior assays) results were included. Clinical studies, conferences, book chapters, reviews, and studies conducted before 2010 were not included. Studies with animals with comorbidities, with animal species other than rodents, in silico, and in vitro studies were not included.
Study Selection and Data Extraction
After the database search, the studies were retrieved to Rayyan (https://www.rayyan.ai/) and duplicates were removed. Title and abstracts were independently selected regarding the eligibility criteria by two authors (CB and JVS). The evaluation of the full texts was performed independently by two authors (CB and JVS). The data (diet type, animal strain, study design, behavioral, biochemical and molecular alterations, and main conclusions) were collected independently by two authors (DM-F and CSO) and organized in an Excel spreadsheet. All discrepancies were resolved by senior authors (DM-F and RGM) at all stages.
Assessment of the Risk of Bias
All studies included in this review were assessed for methodological quality using the RoB SYRCLE tool [19], considering the risk of bias (RoB) for animal studies. The assessment was conducted by two independent authors (CB and JVS) and included observation of the following characteristics, i) randomized housing, ii) description of allocation, iii) incomplete outcome data, iv) selective outcomes, and v) other sources of bias: statistical analysis and conflict of interest statement. For each analysis, there were three possible ratings (“low risk”, “high risk” or “uncertain risk”). If the information was unclear, the rating “no information” was selected. Articles with a high risk of “other sources of bias” were rated as high risk overall.
Results
Characteristics of the Systematically Selected Studies
Following a literature search, we identified 354 studies, and 92 duplicates were removed, totaling 262 studies for title and abstract review. After applying predefined inclusion and exclusion criteria, 59 studies examining the effects of various dietary interventions on biochemical, molecular, and behavioral outcomes were retrieved for full-text assessment. After detailed evaluation, 16 studies met all eligibility criteria, while 43 were excluded for not fully complying with the inclusion criteria upon full-text review. Additionally, 12 relevant articles were identified through manual searches of reference lists and complementary sources. Consequently, a total of 28 studies were included and qualitatively analyzed in this systematic review (Fig. 1). From those 28 studies, 11% evaluated the effects of a cafeteria diet, 22% examined the high-fat and high-sugar diet, 63% examined the effects of a high-fat diet and 7% assessed the effect of a high-sugar diet on biochemical, molecular, and/or behavioral alterations in rodents. It is worth noting that one study assessed more than one type of diet.Fig. 1. Flowchart of studies covered by the search strategy used to produce this review
Assessment of the Risk of Bias
The results regarding the risk of bias are presented in Fig. 2. The vast majority of studies showed a low risk of bias.Fig. 2. Assessment of methodological quality and risk of bias using the Rob SYRCLE tool for animal studies. The percentage of articles found in each category is shown in color and in bars, with green for low-risk studies, red for high-risk studies, and blue for studies that did not report the information sought
Main Findings
The main characteristics and findings of the included studies are described in Table 1. Most studies (63%) used a high-fat diet, to investigate the effects of dietary imbalance on biochemical, molecular, and behavioral changes. The diets offered to the animals during the experimental protocols varied considerably in composition across studies. When specified, diets were mainly composed of different fat sources (including saturated fat (e.g., lard) and polyunsaturated fats such as sunflower oil), carbohydrates (e.g., corn starch, sucrose, and fructose), protein sources (e.g., casein), fiber, and cholesterol. However, several studies did not provide detailed information on the specific type or source of macronutrients. In some experimental models, authors used commercially available human foods to induce a high-calorie intake, including white chocolate, orange-flavored soda, sausages, cookies (e.g., pizza-flavored crackers, chocolate chip cookies, strawberry-flavored cookies, Monte Carlo, Scotch Fingers, and Oreos), cakes (e.g., chocolate mud pie, jam roll, and lamingtons), processed protein products (e.g., party cakes, dim sims, and dog roll), or chow supplemented with bacon mortadella. Table 1. Main characteristics and findings of the included studiesDietStrain/Age/NStudy DesignMain ConclusionsReferenceHigh-fat diet (HFD; D12331, 5.56 kcal/g): 58% fat, 25.5% carbohydrate, and 16.4% proteinStandard control diet (CD; D12328; 4.07 kcal/g): 10.5% fat, 73.1% carbohydrate, and 16.4% proteinDamsC57BL/6N mice/4 weeks-old/~ 26OffspringC57BL/6N mice/10 and 50 days-old/~ 208F1 offspring were born to dams fed either a CD or an HFD for 70 days before mating and throughout pregnancy (~ 21 days). After weaning, all F1 offspring were fed standard rodent chow for approximately 50 daysMaternal consumption of an HFD disrupted neuroendocrine regulations in male offspring, while inflammatory-redox balance and emotionality are impaired in female offspring. Did not report the maternal or F1 offspring weight[20]High-fat diet (HFD; Research Diets Inc.): 60% kcal from fatStandard control diet (CD; Research Diets Inc.): 10% kcal from fatDamsCD-1 IGS mice/N.I./~ 16OffspringCD-1 IGS mice/21 and 112 days-old/~ 96F1 offspring were born to dams fed either a CD or an HFD for 42 days before mating and throughout pregnancy and lactation (~ 42 days); after weaning, offspring either were analyzed immediately or were maintained on standard rodent chow for approximately 90 daysMaternal HFD intake causes anxiety- and depressive-like behaviors in F1 offspring, as well as alters the expression of genes involved in the neuroplasticity process. HFD caused a 20% body weight increase compared to CD in dams and F1 offspring[21]High-fat diet (HFD): 25.7% fat (4.5% palmitic acid, 1.99% stearic acid, 0.12% palmitoleic acid, 6.86% oleic acid, 2.58% linoleic acid, 0.25% α-linolenic acid, and 0.19% arachidonic acid), 19.5% protein, 41.3% carbohydrate, 3.5% fiberStandard control diet (CD): 5.3% fat [corn oil], 21.2% protein, 57.4% carbohydrate, and 4.6% fiberDamsSprague–Dawley rats/44 weeks-old/16OffspringSprague–Dawley rats/N.I./~ 128 (F1); ~ 176 (F2); ~ 464 rats (F3)F1, F2, or F3 offspring were derived from dams fed either a CD or an HFD for approximately 50 days; offspring were subsequently maintained on standard rodent chow for ~ 50 days, except for a subgroup of F3 offspring from chow-fed dams that received an HFD for ~ 50 daysCentral insulin resistance appears to be responsible for the transmission of maternal HFD-induced brain vulnerability and cognition impairment to the next generations, through the posttranslational modifications of glutamate receptors in the hippocampus. Maternal weight was not reported. Maternal HFD consumption did not alter F1, F2, and F3 offspring weight[22]High-fat diet (HFD): 60% kcal from fat, 21% kcal from carbohydrates, and 19% kcal from proteinsStandard control diet (CD): 13% kcal from fat, 67% kcal from carbohydrates, and 20% kcal from proteinsMale C57BL/6 mice/4 weeks-old/42Mice were assigned to groups based on diet type and duration: CD for 7 days, HFD for 7 days, CD for 14 days, HFD for 14 days, or HFD for 7 days followed by CD for 7 daysHFD reduces dendritic tree complexity of DCX + cells and this effect was correlated with reduced BDNF expression. HFD animals showed increased caloric intake; however, body weight did not differ from CD animals[23]High-fat diet (HFD; saturated fat): 9 kj/g (~ 47.6% kcal) from fat (39.2% lard, 45.1% monounsaturated fatty acids, and 11.2% sunflower oil), 6.9 kj/g from carbohydrates, and 3 kj/g from proteinsHigh-fat diet (HFD; polyunsaturated fat): 8.5 kj/g (~ 47.2% kcal) from fat (10.1% lard, 22.8% monounsaturated fatty acids, and 58.3% sunflower oil), 6.9 kj/g from carbohydrates, and 3 kj/g from proteinsStandard control diet (CD): 2.7 kj/g (~ 20.9% kcal) from fat, 7.2 kj/g from carbohydrates, and 3 kj/g from proteinsMale Sprague–Dawley rats/N.I./36Animals were allocated to groups and fed for 13 days with either CD, HFD-saturated fatty acids, or HFD-polyunsaturated fatty acidsHFD-saturated fatty acids alters the hypothalamic marker of inflammation and the exploratory/memory behavior. Body weight did not differ among groups; however, fat-derived energy intake was higher in both HFD groups compared to CD[24]High-fat diet (HFD; TP23300): 60% from fat (lard/soybean oil mixture; 10:1), 20.6% carbohydrate, and 19.4% proteinStandard control diet (CD; TP23303): 70.6% carbohydrate, 10% fat, and 19.4% proteinMale C57BL/6 mice/4 weeks-old/~ 46Animals were assigned to groups and fed either CD or an HFD for 28 daysHFD intake effects in adolescence on emotion and neuroplasticity may be attributed to the microglial engulfment of nascent neurons. Body weight was not reported[25]High-fat diet (HFD; TP23300): 60% from fat (lard/soybean oil mixture; 10:1), 20.6% carbohydrate, and 19.4% proteinStandard control diet (CD; TP23303): 70.6% carbohydrate, 10% fat, and 19.4% proteinMale C57BL/6 mice/4 weeks-old/~ 40Animals were allocated to groups and fed either CD or an HFD for 35 daysHFD consumption induces microglial dysfunction and prefrontal neuroplasticity deficits associated with mood disorders in adolescent mice. HFD animals showed increased caloric intake and body weight compared to CD animals[15]High-fat diet (HFD): 60% from fatStandard control diet (CD): N.IMale C57BL/6N mice/8 weeks-old/~ 20Animals were allocated to groups and fed either CD or an HFD for 77 daysHFD consumption did not alter the markers of neuroplasticity in the amygdala. HFD animals showed increased body weight compared to CD animals[26]High-fat diet (HFD): 60% from fatStandard control diet (CD): N.IMale C57BL/6 mice/8 weeks-old/N.IAnimals were allocated to groups and fed either CD or an HFD for 84 daysHFD intake disturbs the function of hippocampal astrocytes and induces depression-like behaviors, which could be related to alterations in glutamate transporters. HFD animals showed increased body weight compared to CD animals[27]High-fat diet (HFD): 60% from fat, 20% carbohydrate, and 20% proteinStandard control diet (CD): 62.9% carbohydrate, 12.5% fat, and 24.6% proteinMale Sprague–Dawley rats/7 weeks-old/~ 75Animals were allocated to groups and fed either CD or an HFD for 112 daysHFD intake causes alteration in endoplasmic reticulum stress proteins and apoptosis, which could be related to the neuroplasticity alterations. HFD animals showed increased body weight compared to CD animals[28]High-fat diet (HFD): 60% from fat (lard)Standard control diet (CD): 10% from fatMale C57BL/6 mice/36 weeks-old/32Animals were allocated to groups and fed either CD or an HFD for 140 daysHFD intake impairs immature neurons in the hippocampus. HFD animals showed increased body weight compared to CD animals [29] High-fat diet (HFD): 60% from fatStandard control diet (CD): N.IMale C57BL/6 J mice/N.I./60Animals were allocated to groups and fed either CD or an HFD for 140 daysHFD intake reduces PPAR levels and could be related to neuroinflammation with secondary degeneration of dopaminergic neurons. HFD animals showed increased body weight compared to CD animals[30]High-fat diet (HFD): 60% from fatStandard control diet (CD): 10% from fatMale C57BL/6 mice/4 and 8 weeks-old/~ 50Animals were allocated to groups and fed either CD or an HFD for 168 daysHFD intake effects on neurobehavior and hippocampal neuroplasticity were more pronounced in the young animals. HFD animals showed increased body weight compared to CD animals[31]High-fat diet (HFD, p1400f): 45 to 60% from fatStandard control diet (CD): N.IMale C57BL/6 mice/N.I./N.IAnimals were allocated to groups and fed either CD or an HFD for 210 daysHFD intake alters hippocampal neuroplasticity via GLP-1R/BDNF/TrkB signaling. Body weight was not reported[32]High-fat diet (HFD): 60% from fatStandard control diet (CD): 10% from fatMale C57BL/6 mice/4 and 8 weeks-old/~ 50Animals were allocated to groups and fed either CD or an HFD for 252 daysHFD intake effects on neurobehavior and hippocampal neuroplasticity were more pronounced in the young animals. HFD animals showed increased body weight compared to CD animals[33]High-fat diet (HFD; TP23300): 60% from fat (lard/soybean oil mixture; 10:1), 20.6% carbohydrate, and 19.4% proteinStandard control diet (CD; TP23303): 70.6% carbohydrate, 10% fat, and 19.4% proteinMale C57BL/6 mice/8 weeks-old/48Animals were allocated to groups and fed either CD or an HFD for 168 or 252 daysHFD consumption alteration on behavior and hippocampal neuroplasticity appears to be attributable to alteration of microglial status, which is closely associated with the cellular lipid content. HFD animals showed increased body weight compared to CD animals[34]High-fat diet (HFD): 60% from fat (lard)Standard control diet (CD): N.IMale C57BL/6 mice/~ 9 weeks-old/30Animals were allocated to groups and fed either CD or an HFD for 504 daysHFD intake alters IGF-IRβ/PI3K/Akt/Gsk-3β and mTOR signaling pathways. HFD animals showed increased body weight compared to CD animals[35]High-fat high-sugar diet (HFHS): 20% fat (lard), 39.6% sucrose, and 19.4% proteinStandard control diet (CD): N.IMale Sprague–Dawley rats/~ 4 weeks-old/32Animals were assigned to groups and fed either CD for 28 days or CD for 28 days with additional access to a HFHS diet for 2 h/dayHFHS diet intake caused physiological changes to the brain, particularly expression of mRNA associated with reward and neuroplasticity, which could explain the behavioral alterations. HFHS animals showed increased caloric intake and body weight compared to CD animals [36] High-fat high-sugar diet (HFHS): 20% fat (lard), 39.6% sucrose, and 19.4% proteinStandard control diet (CD): 65% carbohydrates, 12% fat, and 23% proteinMale Sprague–Dawley rats/~ 4 weeks-old/24Animals were assigned to groups and fed either CD for 28 days or CD for 28 days with additional access to a HFHS diet for 2 h/dayHFHS intake induces prefrontal cortex dysfunction and subsequent behavioral deficits. Neither body weight nor energy intake differed among the animals[37]High-fat high-sugar diet (HFHS): 46% fructose and 20% lardStandard control diet (CD): 5% cellulose, 20% casein, 25% maize starch, 25% potato starch, 16% maltodextrin, and 4% soybean oilMale Wistar rats/8 weeks-old/60Animals were allocated to groups and fed either CD or an HFHS for 84 daysHFHS intake altered several inflammatory and neuroplasticity markers. Neither body weight nor fat weight differed among the animals[38]High-fat high-sugar diet (HFHS): 29% fat, 34% sucrose, and 1.25% cholesterol combined with glucose/fructose-supplemented drinking water (42 g/L; 55%/45% w/w)Standard control diet (CD): 6.2% fat, 44% carbohydrate, and 18% proteinMale C57BL/6 mice/3 weeks-old/~ 10Animals were allocated to groups and fed either CD or an HFHS for 203 daysHFHS intake caused neuroplasticity alteration through IL-17 signaling. Body weight was not reported[39]High-fat high-sugar diet (HFHS): 21.2% fat, 34% sucrose, and 0.2% cholesterolStandard control diet (CD): 5.2% fat, 12% sucrose, and 0.01% cholesterolC57BL/6 mice/3 weeks-old/~ 12Animals were allocated to groups and fed either CD or an HFHS for 252 daysHFHS diet intake disturbed the bile acid signaling pathways, which seems to be related to neuroplasticity impairment. Body weight was not reported[40]High-fat high-sugar diet (HFHS): 21.2% fat, 34% sucrose, and 0.2% cholesterolStandard control diet (CD): 5.2% fat, 12% sucrose, and 0.01% cholesterolMale C57BL/6 mice/3 weeks-old/~ 12Animals were allocated to groups and fed either CD or an HFHS for 287 daysHFHS diet intake disturbed the bile acid signaling pathways, which seems to be related to neuroplasticity impairment. HFHS animals showed increased body weight compared to CD animals[41]Cafeteria diet (CAF): CD supplemented with a variety of palatable human foods, including cakes, biscuits, and savoury protein-rich items (e.g., party pies, dim sims, dog roll), and 10% sucrose solution in addition to waterStandard control diet (CD): N.IMale Sprague–Dawley rats/N.I./20Animals were allocated to groups and fed either CD or a CAF for 25 daysCAF diet intake disturbs inflammatory and neuroplasticity pathways. CAF diet increased caloric intake and body weight compared to CD [42] Cafeteria diet (CAF): 32% from fat, 57% from carbohydrates, and 10% from protein; CD supplemented daily with a variety of palatable high-fat, high-sugar human foods (e.g., meat pies, cakes, biscuits)Standard control diet (CD): 12% from fat, 23% protein, and 65% carbohydrateMale Sprague–Dawley rats/6 weeks-old/32Animals were allocated to groups and fed either CD or a CAF for 56 daysCAF diet intake altered hippocampal plasticity and caused contextual fear memory deficits. CAF diet increased body weight compared to CD[43]Cafeteria diet (CAF): 42% carbohydrates, 16% protein, and 42% lipids; CD supplemented ultra-processed, highly palatable foods (e.g., bacon mortadella, sweet biscuits, chocolate biscuits, pizza-flavored crackers, white chocolate, sausages, and sugar-sweetened soda)Standard control diet (CD): 63% carbohydrates, 26% protein, and 11% lipidsMale Wistar ratsAge = 12 weeks-oldN = 14Animals were allocated to groups and fed either CD or a CAF for 140 daysCAF diet intake altered the marker of neuroplasticity. Body weight was not reported[44]High-sugar diet (HS): 2.69 kj/g from fat, 4.12 kj/g from complex carbohydrates, 4.2 kj/g from simple carbohydrates (sucrose), and 2.98 kj/g from proteinsStandard control diet (CD): 2.7 kj/g from fat, 5.11 kj/g from complex carbohydrates, 2.05 kj/g from simple carbohydrates, and 3 kj/g from proteinsMale Sprague–Dawley rats/N.I./24Animals were allocated to groups and fed either CD or an HS for 13 daysHS consumption did not alter the expression of the marker of neuroplasticity evaluated, but altered the hypothalamic marker of inflammation and the exploratory/memory behaviors. Body weight did not differ among groups; however, sucrose-derived energy intake was higher in both HS group compared to CD[24]High-sugar diet (HS): 1.5 g/100 g from fat (sunflower oil), 20.4 g/100 g from carbohydrates (fructose), and 9.2 g/100 g from proteins (casein)Standard control diet (CD): 1.5 g/100 g from fat (sunflower oil), 20.4 g/100 g from carbohydrates (cornstarch), and 9.2 g/100 g from proteins (casein)Male Wistar ratsAge = ~ 3 weeks-oldN = ~ 32Animals were allocated into three groups and fed either CD for 42 days, HS for 21 days, or HS for 21 days followed by CD for an additional 21 daysSeveral of the alterations in the frontal córtex (BDNF, CML, CEL, acetylcholinesterase activity, dysregulation of neurotransmitter levels) persisted after switching to the control diet, thus pointing to adolescence as a critical phase. HS did not alter body weight nor intake energy compared to CD[45]
The time during which the animals were exposed to the diets ranged from a few days (e.g., 7 days) to weeks, months, or even years (e.g., 504 days). Mice or rats, male or female animals of different ages (from 3 to 64 weeks old) were used. In addition, some studies investigated whether the offspring of mothers exposed to a high-fat and/or high-sugar diet showed behavioral or neuronal changes. The animal strains used also varied widely. The studies included C57BL/6 and C57BL/6N mice, CD-1 IGS mice, and Sprague–Dawley and Wistar rats.
The behavioral changes induced by a high-calorie diet, as reported in the systematically selected studies, are presented in Table 2. While a few studies found no changes in locomotor activity and memory, most studies indicated that a high-calorie diet can cause various cognitive and other changes. Almost all studies described anxiety- and depression-like behaviors in both young and adult, male and female animals following acute or chronic exposure to high-calorie diets. Other effects included decreased exploratory behavior, social interaction and social motivation and recognition; decreased learning and memory and decreased cognitive flexibility; increased passive stress coping; cognitive impairment, and trace fear conditioning. In addition, the offspring of mothers exposed to a high-fat and/or high-sugar diet exhibited anxious and/or depression-like behaviors. The mothers also showed increased passive stress coping, spent more time away from the nest, and were less likely to nest-build. Table 2. Behavioral changes induced by a high-calorie diet, as reported in the systematically selected studiesDietType of AlterationAlterationReferenceHFDBehavioral↓ exploratory behavior in female offspring↑ active coping strategy in female and male offspring[20]HFDBehavioral and SocialDams↑ time spent away from the nest↓ the frequency of nestbuilding↑ passive stress copingF1 offspring↑ anxiety- and depressive-like behaviors↑ passive stress coping↓ social interaction[21]HFDBehavioral and Cognitive↓ learning and memory in F1, F2, and F3↓ learning and memory in F3 offspring fed with an HFD[22]HFDBehavioral↓ exploratory activity[24]HFDBehavioral↑ anxiety- and depressive-like behaviors[25]HFDBehavioral↑ anxiety- and depressive-like behaviors[15]HFDBehavioral↓ locomotor activity↑ depression-like behaviors[27]HFDBehavioral and Cognitive↑ cognitive Impairment and anxiety-like behavior↓ locomotor activity[30]HFDBehavioral and Cognitive↓ cognitive flexibility↑ anxiety and depression-like behavior[31]HFDBehavioral↑ depression-like behavior[32]HFDBehavioral↑ anxiety and depression-like behavior[33]HFDBehavioral↑ anxiety and depression-like behavior[34]HFDCognitive↓ spatial memory and learning[35]HFHSSocial↓ social motivation and recognition [46] HFHSSocial↓ social recognition[37]HFHSBehavioral↑ distance traveled, and time spent in the center[39]CAFN/ANo alterations were observed in the memory and locomotor activities[47]CAFBehavioral↓ trace fear conditioning[43]HSBehavioral↓ exploratory activity[24]HFD high-fat diet, HFHS high-fat high-sugar diet, CAF cafeteria diet, HS high-sugar diet
Data was extracted from all eligible studies. For clarity and synthesis, Table 2 highlights behavioral outcomes with reported diet-related alterations. Behavioral tests with no significant differences between groups were not individually included in the table.
Table 3 shows the main biochemical and molecular alterations caused by a high-calorie diet, as reported in the systematically selected studies. In terms of biochemical and molecular changes, studies have investigated a variety of markers that indicate acute and chronic changes in neuronal remodeling and consequently confirm the above findings on cognitive decline and behavior. The hippocampus was the most frequently examined structure in the studies included in this systematic review. Changes described include increases and decreases in the expression of various molecules such as BDNF and inflammatory markers, including TNF-α, IL-1β, and IL-6. Changes were also described in the hypothalamus, prefrontal cortex, substantia nigra, striatum, and microglia. Some authors did not isolate specific structures for dosages but also showed changes in the general structure of the brain, and others provided results on plasma changes, such as changes in corticosterone levels. Table 3. Biochemical and molecular alterations caused by a high-calorie diet, as reported in the systematically selected studiesDietTissue/OrganBiochemical and Molecular ChangesReferenceHFDPlasma; Hippocampus↓ basal plasma corticosterone levels in male offspring↓ BDNF, NRF-2, and Keap-1 expression in the hippocampus only in the female offspring↑ CD68 expression in male and female offspring↑ Ucp-2 expression in female offspring[20]HFDWhole brain; Prefrontal cortexDams↑ brain TLR4Pups↑ prefrontal cortex GLUN2C and ZIF-268 expression in 21-day-old female offspring↓ prefrontal cortex SYP expression 112-days-old offspring[21]HFDHippocampus↓ cornu ammonis CA 3-CA1 hippocampal long-term potentiation (LTP) in the F1, F2, and F3↓ CA 3-CA1 hippocampal LTP in the F3 offspring fed with a HFD↓ hippocampal CA1 dendritic spine density in F3 offspring and F3 offspring fed with an HFD[22]HFDHippocampus↓ dendritic branches and dendritic intersections in the DCX-positive neurons in the hippocampus↓ BDNF in the dorsal hippocampus[23]HFDHypothalamus↓ hypothalamic NfkBia expression[24]HFDHippocampus↓ Ki67- and DCX- positive cells in the hippocampus↓ dendritic branches, dendritic length, and the number of dendritic intersections in the DCX-positive neurons in the hippocampus↑ hippocampal Ibal- and CD68-positive cells↑ microglial engulfment of DCX positive material in the hippocampus[25]HFDPrefrontal cortex↓ dendritic branches, dendritic length, and the number of dendritic intersections in the medial prefrontal cortex↓ PSD95 density in the medial prefrontal cortex↑ microglial phagocytosis of the PSD95 in the medial prefrontal cortex[15]HFDAmygdalaNo alteration in TrkB, BDNF, SYT-1, SYT-4, and SNAP-25 protein levels in the amygdala[26]HFDHippocampus↑ activation of astrocytes (GFAP protein) in the hippocampus↓ intersections and branches in GFAP-positive astrocytes in the hippocampus↓ astrocytic neuroplasticity-related proteins (GLAST, GLT-1, and Cx-43) in the hippocampus[27]HFDPrefrontal cortex↑ FATP1, GRP78, p-PERK/PERK, p-elF2α/elF2α, caspase 12, CHOP, and Bax/Bcl-2 in the prefrontal cortex↓ BDNF and SYN in the prefrontal cortex[28]HFDHippocampus↓ dendritic branches in the DCX-positive neurons in the hippocampus [48] HFDNigrostriatal system (Substantia nigra + Striatum); Mesolimbic system↓ dopaminergic neurons in the substantia nigra↓ dendritic spine density in the substantia nigra and striatum↑ Iba-1 and GFAP in the substantia nigra and striatum↓ PPAR in the dopaminergic neurons of the substantia nigra and ventral tegmental area[30]HFDHippocampus↓ dendritic branching, branch length, complexity of neuronal dendritic trees, and spine density in the hippocampus of 4 weeks initial age of treatment↑ CD68-positive cells in the hippocampus of both ages evaluated↑ microglial engulfment of PSD95^+^ puncta at 4 weeks after the initial age of treatment↑ p-IR in hippocampal microglia 4 weeks after the initial age of treatment[31]HFDSerum; Hippocampus↓ serum 5-HT↑ serum corticosterone, IL-1β, and IL-6↓ hippocampal BDNF, p-ERK/ERK, p-TrkB, p-AMPK, p-CREB/CREB, and GLP-1R[32]HFDHippocampus↑ hippocampal insulin↓ hippocampal pY-IR/IR in 4 weeks initial age of treatment↓ hippocampal DCX positive cells in 4 weeks initial age of treatment↓ hippocampal DCX and BDNF levels in 4 weeks initial age of treatment[33]HFDHippocampus↓ hippocampal dendritic length, dendritic intersections, and dendritic spine density↑ hippocampal CD68-positive cells↑ microglia engulfment of PSD95 and lipid droplets in the hippocampus[34]HFDHippocampus↑ hippocampal TNF-α, COX-2, Caspase-3, Iba-1, GFAP, p-JNK, AKT, mTOR, p-Tau, Beta-Amyloid↓ hippocampal NeuN, Ki-67, p-CREB, BDNF, IGF-1R, JNK, PI3K, p-PI3K, p-AKT, GSK3β, p-mTOR, GLUT4, Tau, SAP 102[35]HFHSHippocampus; Prefrontal cortex↓ BDNF, MaoA, COMT IL-6, NLRP3 expression in the prefrontal cortex↓ MaoA expression in the hippocampus [46] HFHSPrefrontal cortex↓ the total number of parvalbumin-positive interneurons and perineuronal nets in the medial prefrontal cortex↑ FosB/ΔFosB in the prefrontal cortex[37]HFHSSerum; Hypothalamus; Hippocampus↑ plasmatic IL-6 concentration↓ hypothalamic IL-1R1, mineralocorticoid receptors, corticotropin-releasing hormone, NGF, and SYP expression↑ hippocampal IL-1ra, glucocorticoid receptor expression[38]HFHSHippocampus; Whole brain homogenate/isolated microglia↓ long-term potentiation (LTP) at Schaffer collateral–CA1 synapses↓ BDNF and PSD-95 protein in the brain↑ RoRyt, IL-17A, IL-22, IL-1β, IL-6, TNF-α, MCP-1, CCL-5, CCL-17, CCL-20, NOS-2, F4/80, CX3CR1, KCA3.1, KV1.3 expression in the brain[39]HFHSHippocampus; Frontal cortex; Whole brain↓ long-term potentiation (LTP) at Schaffer collateral–CA1 synapses↑ hippocampal IL-1β, IL-6, NOS2, CH25H, SAA1, TNF-α, CCL5, CCL17, CCL20, ApoE, KCA3.1, KV1.3, and KIR2.1 expression↑ CD11b, pERK1/2, IL-6, IL-1beta, and TNF-α levels in the brain↓ GPBAR1, BDNF, and PSD-9 levels in the brain↓ BDNF, GPBAR1, PC1/3, NOS1, DIO2, GLP1R, PYY, FXR, CYP46A1, NOS3, SHP, CYP39A1, and CYP27A1 expression in the brain[40]HFHSHippocampus; Frontal cortex and ventral posteromedial thalamic nucleus; Whole brain homogenate/isolated microglia↓ long-term potentiation (LTP) at Schaffer collateral–CA1 synapses↑ IL-1β, IL-6, TNF-α, FOXp3, SAA1, CCL17, CCL20, ICAM1, KCA3.1, and KV1.3 expression in the brain and isolated microglia↓ claudin1 expression in the brain↓ Occludin, RARβ, CYP26A1, and ALDH1A1 expression in isolated microglia↓ TGR5, GLP1R, PYY, FGF21, and BDNF expression in the brain and isolated microglia↑ DCA and TCA levels in the brain↓ βMCA, CDCA, UDCA, TLCA, and T-α-β-MCA levels in the brain↑ HMGCR, SQLE, CH25H, CYP39A1, CYP7A1 expression in the brain↓ CYP27A1, CYP7B1, CYP46A1, FXR, SHP expression in the brain↑ CH25H, CYP7A1 expression in isolated microglia↓ HMGCR, SQLE, CYP27A1, CYP7B1, CYP46A1, CYP39A1, FXR, SHP expression in isolated microglia[41]CAFHippocampus; Perirhinal cortex↑ Homer1a, IGF1, Syn1, BDNF, CREB1, mTOR, GFAP, Ptgs2, 5HT1A, mGLUR5, Grin2b, MAPK8, MAPK10, Nr3c1, GLUT3, JAK2, and NOD2 expression in the hippocampus↓ IKBKB and 5HT2c expression in the hippocampus↑ BDNF expression in the perirhinal cortex↓ MCP-1 expression in the perirhinal cortex[47]CAFHippocampus↓ hippocampal Reelin expression[43]CAFHippocampus↓ hippocampal synaptophysin levels[44]HSHypothalamus↓ hypothalamic NfkBia expression[24]HSFrontal Cortex↑ Glut-5 and GFAP expression in HS feedings rats↑ fructose, uric acid, TNF-α, Hpt, N-Tyr, TrkA, and acetylcholine levels in HS feedings rats↓ Trkβ, dopamine, tyrosine, and tyramine levels in HS feedings rats↓ Synaptophysin, synaptotagmin I, and PSD-95 expression in HS feedings rats↑ AChE and MAO activity in HS feedings rats↑ CML and CEL levels in HS and HS + rodent chow recover feeding rats↑ pErk1/Erk1 and pErk2/Erka ratio in HS and HS + rodent chow recover feeding rats↓ BDNF and pCREB levels in HS and HS + rodent chow recover feeding rats↑ glutamate and GABA levels in HS + rodent chow recover feeding rats[45]HFD high-fat diet, HFHS high-fat high-sugar diet, CAF cafeteria diet, HS high-sugar diet
Discussion
The concept of a healthy diet has evolved significantly in recent years, shifting from a primary focus on caloric adequacy and macronutrient balance to a broader emphasis on dietary quality and overall eating patterns. Epidemiological and non-clinical studies have shown that adequate nutrition, combined with physical activity and other lifestyle factors, plays a central role in promoting general health and well-being [49]. At the same time, research across multiple disciplines has consistently found that high consumption of Western-style, energy-dense, and ultra-processed foods is strongly associated with overweight, obesity, and an increased risk of metabolic and cardiovascular diseases, including diabetes [50].
The detrimental effects of consuming foods high in saturated fat, sodium, sugar, and easily assimilated carbohydrates appear to be due not only to their very low nutritional value but also to their tendency to promote non-homeostatic behaviors characterized by repetitive and excessive consumption of foods, which can alter gut-brain communication and the links between taste perception and nutrient response [51, 52]. If these changes persist, the previously established behavior can cause several important problems, such as a change in the homeostatic environment of the gut, intestinal inflammation, and hormonal problems that contribute to neuronal adaptations in the central nervous system, leading to more serious issues, including cognitive decline [53–55].
In fact, in this systematic review, we have described some studies that confirm the above. Most studies that have examined the negative effects of a high-fat, high-sugar diet or a cafeteria diet have primarily documented decreased performance on tasks that depend on good hippocampal and prefrontal cortex function, and interestingly, these findings have often been associated with cognitive decline, anxiety, and depression-like states. However, the mechanisms behind all the changes described are only partially understood. Some authors speculate that a high-calorie diet could affect the brain directly, while others put forward the idea that this may occur through an indirect effect via the periphery [54, 56–58].
A high-fat diet can alter cerebral vascular homeostasis by damaging the endothelium of cerebral microvessels, impairing the absorption of nutrients and the excretion of toxic substances, and leading to an increase in inflammatory markers in the brain [59]. A persistent state of inflammation in certain brain regions, including the hippocampus, hypothalamus, and prefrontal cortex, contributes to acute and chronic structural and functional changes and can lead to decreased regional blood flow and volume, activation of microglia, increased gene expression of proinflammatory brain mediators, including IL-1β and TNF-α [60], and increased markers of cell death [60–62]. Ultimately, it can lead to a reduction in specific biomarkers such as BDNF [14, 63, 64] and result in acute and long-term changes associated with significant alterations in brain reward circuitry, contributing to the development of anxiety and depression-like behaviors in animal models [65, 66]. Metabolic abnormalities such as decreased expression of glucose transporters and alterations in insulin metabolism can also cause neuronal changes at the morphological and physiological level, reflected in a reduction in short- and long-term potentiation response and a decrease in specific markers of synaptic neuronal plasticity [59, 67]. In addition, excessive consumption of certain nutrients, such as salt, can disrupt peripheral hormone metabolism, impair bile acid signaling pathways, alter the composition of the gut microbiota, and disrupt the protective barrier of the intestinal epithelium [67, 68]. These peripheral changes appear to exacerbate the negative effects of a high-calorie diet and influence the development of central nervous system problems via the gut-brain axis [39, 41].
Interestingly, some authors have investigated whether different times and different moments of exposure to a high-calorie diet can cause changes in the brain and behavior; or even whether exposure in utero can lead to altered phenotypes in the offspring and the risk of long-term morbidity. Long time spans [23, 26, 27, 69], but also very short periods of high-fat diets or cafeteria diets can impair cognitive function and inflammatory and neuroplasticity pathways in the brain [30, 35, 70]. Mechanistically, these early cognitive impairments appear to be driven by converging inflammatory, vascular, and neuroplasticity-related pathways triggered by exposure to high-fat diets**.** Using a high-fat diet providing 60% of total energy from fat, Kao et al. [30] demonstrate that HFD-induced neuroinflammation in nigrostriatal and mesolimbic circuits, marked by microglial and astroglial activation, reduced PPAR signaling, dopaminergic neuron loss, and decreased dendritic spine density is associated with cognitive deficits and anxiety-like behavior. Similarly, De Paiva et al. [35] show that an HFD (60% fat derived from lard) is sufficient to induce hippocampal inflammation, activation of stress and neurodegenerative pathways, and suppression of neurotrophic and synaptic plasticity signaling, culminating in impaired learning and memory. Extending these findings beyond diet exposure alone, Feng et al. [70] identify obesity-induced blood–brain barrier dysfunction as a critical upstream mechanism that facilitates peripheral inflammatory signaling into the brain and amplifies neuroinflammation and cognitive decline.
While some studies indicate that the effects are more pronounced in young animals [15, 25, 31, 33], others describe that the adverse effects do not depend on age and can extend over generations [34]. Offspring of mothers exposed to a high-fat and/or high-sugar diet also show significant changes, including metabolic syndrome, cardiovascular and cerebrovascular problems, and behavioral changes [20–22].
Recent research has also suggested that overconsumption of hyperpalatable diets could be associated with the development of low-grade inflammation [71]. Although research on this topic is still limited, preliminary evidence suggests that overconsumption of such foods, possibly due to their non-nutritive components, may contribute to chronic inflammation and gut dysbiosis, which have previously been associated with a pro-inflammatory state [72]. Homeostatic balance and the interplay of immune and metabolic responses are disrupted, which can lead to chronic metabolic inflammation [73]. A diet high in salt or sugar contributes to the infiltration of immune cells and the subsequent release of pro-inflammatory cytokines in tissues [74]. This inflammatory reaction can, for example, prevent glucose uptake and alter lipid metabolism [75]. These metabolic changes can easily contribute to glucose intolerance, excessive weight gain, metabolic inflexibility and ultimately diabetes and cardiovascular disease in susceptible individuals [76, 77]. For example, insulin resistance often results from prolonged exposure to inflammatory biomarkers, which often culminate in the development of diabetes [78]. In addition, low-grade inflammation is associated with atheroprogression and may promote the progression of various cancers through increased cell proliferation, decreased apoptosis and increased angiogenesis and metastasis [79–82].
The present study provides an overview of the effects of different types of caloric diets on biochemical, molecular and behavioral changes and the associated consequences, but may be limited by the lack of some information. Some studies examined only female or male animals, and the articles did not consider sex-specific effects. Although some studies have examined animals at different ages and even the effects on the offspring of mothers exposed to a high-calorie diet, no protocols have been developed that reverse the diet, which is an important limitation. Future studies aimed at filling these gaps are important.
Conclusion
Our review suggests that a high-calorie diet induces several changes at the peripheral and central nervous system level, leading to neuronal, functional and structural adaptations that may contribute to behavioral changes in the short and long term. These effects could be related to changes in the gut microbiota, glucose and insulin metabolism, and alterations in bile acid signaling pathways. Finally, certain aspects remain unexplored, such as the reversibility of damage caused by high-calorie diets. This underscores the need for future research aimed at mitigating the harmful effects of such diets on the brain. Additionally, studies investigating the different types and sources of high-calorie diets across various species could be crucial for translating these findings to humans.
Supplementary Information
Below is the link to the electronic supplementary material. ESM 1(DOCX 31.3 KB)