Advances in the use of morphogenic regulators and peptide regenerating factors for boosting plant transformation and genome editing
V. Mohan Murali Achary, Easter D. Syombua, Simranjit Kaur, Sri Cindhuri Katamreddy, Danni Zou, Sarah J. Hearne, Anindya Bandyopadhyay

TL;DR
This paper reviews recent advances in using morphogenic regulators and peptides to improve plant transformation and genome editing for better crop development.
Contribution
The paper introduces novel strategies using morphogenic regulators and peptide factors to enhance plant regeneration and genome editing efficiency.
Findings
Co-expression of GRF and GIF facilitates regeneration in recalcitrant plant varieties.
Peptide factors like REF1 and transcription regulators from WOX, DOF, AP2/ERF families improve transformation efficiency.
Applications across monocots, dicots, and recalcitrant species show promise for genotype-flexible breeding.
Abstract
Plant regeneration and transformation remain significant bottlenecks towards the genetic improvement of most crop species by either genome editing or transgenic approaches. Recent research has therefore, transitioned from manual optimization of culture media and components to the use of morphogenic regulators (MRs) and novel peptide regeneration factors that can reprogram somatic cell fate to a totipotent state. For instance, the co-expression of TRs such as GROWTH-REGULATING FACTORs (GRF), and GRF-INTERACTING FACTOR (GIF) have been shown to facilitate regeneration of transgenic plants from recalcitrant varieties. Genotype dependence and low regenerabilty have also constrained the adoption of precision breeding tools such as Cas9, Cas12a, Cas13, base and prime editors for trait improvement in some species and genotypes. This review first explores the status of plant transformation and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
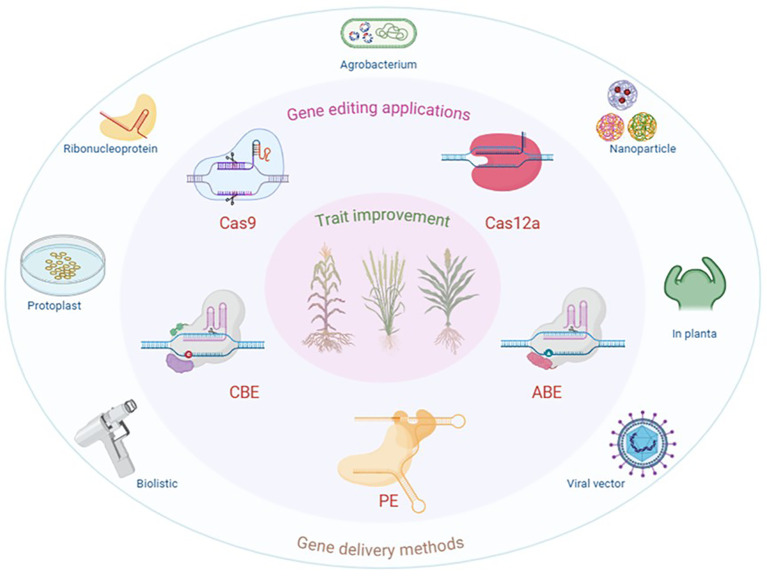 Figure 1
Figure 1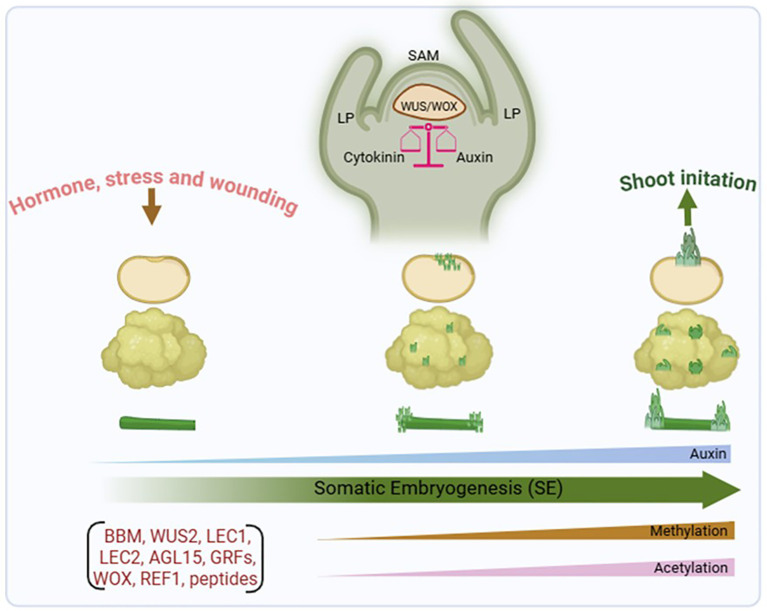 Figure 2
Figure 2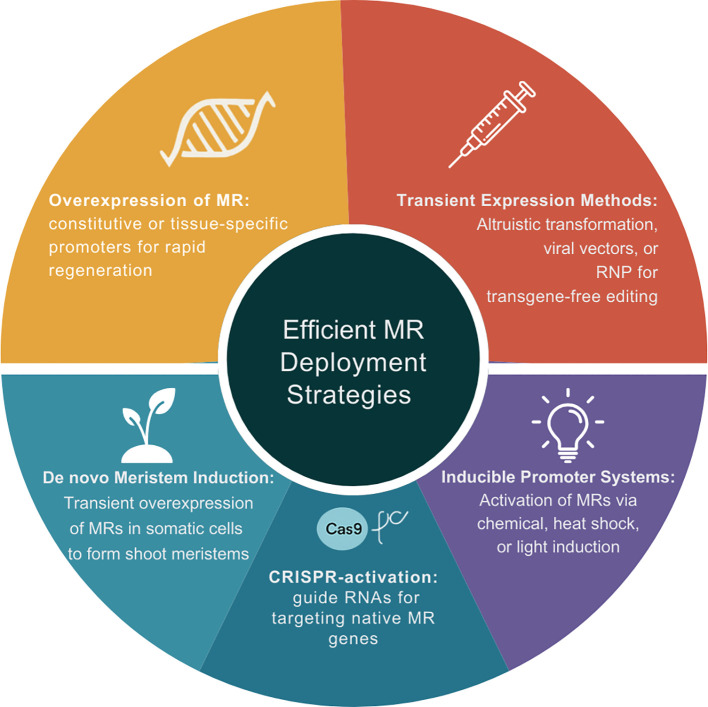 Figure 3
Figure 3| Strategy | Morphogenic regulators | Crop(s) | Key advantages | Limitations/considerations | Promoter(s) | References |
|---|---|---|---|---|---|---|
| Constitutive expression | Induced spontaneous somatic embryo formation, ectopic shoot development, enhanced cell proliferation, and hormone-free regeneration of explants. | Pleiotropic effects: neoplastic growth, dwarfism, altered leaf/flower morphology, delayed flowering, increased anthocyanins, wax deficiency, poor root growth, compromised vegetative development, and reduced male/female fertility. | Double-enhanced 35S + Alfalfa mosaic virus (AMV) translational enhancer; Sunflower UbB1 polyubiquitin | ( | ||
|
| Significantly improves regeneration efficiency | miRNA-resistant VvGRF4-GIF1 and TaGRF4-GIF1 variants caused abnormalities due to deregulated expression | CaMV 35S | ( | ||
|
|
| Dose-dependent developmental outcomes: high expression promotes somatic embryogenesis, while low expression induces organogenesis; also activates LAFL gene expression. | Not specified | CaMV 35S | ( | |
|
| Promotes somatic embryogenesis and enhances regeneration efficiency | Not specified | CaMV 35S | ( | ||
|
| Induces somatic embryo formation and promotes embryo development | Delayed flowering, reduced seed set, enlarged floral organs, elongated cotyledons, post-germination somatic embryos, thick/short hypocotyls, and abnormal roots | Double CaMV 35S promoter with AMV translational enhancer | ( | ||
|
| Involved in somatic embryogenesis | Not specified | CaMV 35S | ( | ||
| Spontaneous somatic embryo formation and enhanced shoot regeneration | Pleiotropic effects: short roots, altered leaf morphology, delayed flowering | Cauliflower mosaic virus 35S | ( | |||
|
|
| Promotes hormone-free transition to embryonic state | Abnormal cotyledon development, serial initiation of multiple somatic embryos (meta-embryos) | CaMV 35S | ( | |
| Tobacco ( | Induces organogenesis; somatic embryogenesis upon cytokinin addition | Severe pleiotropic effects including dwarfism, loss of apical dominance, callus formation, leaf rumpling, floral abnormalities, and sterility | 35S | ( | ||
|
|
| Promote vegetative-to-embryonic transition and somatic embryo formation | Not specified | CaMV 35S | ( | |
|
|
| Strong enhancement of somatic embryo production (400x) and callus formation | Requires specific developmental stage and hormonal conditions for full embryogenic effect | CaMV 35S | ( | |
|
| Capsicum chinense | Induced formation of globular structures suggesting embryogenesis in a highly recalcitrant species | No complete regeneration or transformation system established; early-stage embryogenesis only | CaMV 35S | ( | |
|
| WUS and STM synergistically trigger organogenesis from non-meristematic tissue | Did not restore full meristem self-maintenance or correct phyllotaxy; Arabidopsis-specific | CaMV 35S | ( | ||
|
| Promotes embryogenesis and organogenesis without exogenous hormones; induces somatic embryos from non-embryogenic tissues | Overexpression of a repressor form (GhWUS-SRDX) blocked embryogenic callus formation in cotton, highlighting dosage and regulation sensitivity | CaMV 35S | ( | ||
|
|
| 3× embryogenic explants; induced organogenesis in embryo-like tissues | No plant regeneration in recalcitrant variety under standard protocols; requires optimization of hormones | CaMV 35S | ( | |
|
| 47.75% increase in embryogenic callus formation in a recalcitrant cultivar | Potential pleiotropic effects; evaluated only in callus stage | CaMV 35S | ( | ||
|
| Wheat | 7.8× transformation frequency in elite cv. Fielder, Marker-free selection, Applicable to recalcitrant genotypes, Shoots visible in 7–10 days vs. 3–4 weeks control | Yield-related tradeoff: exhibited a 23.9 % reduction in number of grains per spike | Maize Ubiquitin | ( | |
|
| Triticale | Transformation in previously non-transformable genotypes, Faster shoot emergence | Limited data across cultivars | Maize Ubiquitin | ( | |
|
| Rice | Faster regeneration, Shoots in 6–10 days vs. 15–20 days control, Increased number of transgenic plants | Not validated in japonica/indica breadth | Maize Ubiquitin | ( | |
| Citrus | Improved shoot regeneration from epicotyls, Shoots in 12–14 days vs. 3–4 weeks control, First report of chimeric GRF-GIF effect in dicots | Chimera version not identical to wheat construct | CaMV 35S | ( | ||
|
| Sugar beet | Enabled transformation of recalcitrant varieties; increased transformation efficiency | None reported | 2×35S | ( | |
|
| Sunflower | Improved transgenic shoot formation | None reported | 2×35S | ( | |
|
| Soybean | Improved transgenic shoot formation | None reported | PcUbi4-2 | ( | |
|
| Canola | Promoted callus production | None reported | PcUbi4-2 | ( | |
|
| Maize | Increased transformation efficiency ~3× | None reported | BdEF1 | ( | |
|
|
| Involved in somatic embryogenesis and somatic seedling growth | None reported | CaMV 35S | ( | |
| Induction of shoot regeneration and transformation efficiency | Not specified | CaMV 35S | ( | |||
|
| Bok choy and cabbage | Improved shoot regeneration and transformation | Some morphological alterations; viable seed production | CaMV 35S | ( | |
|
| Sweet pepper | Promoted formation of transgenic callus and somatic embryos | None reported | CaMV 35S | ( | |
|
| Induction of | may impair regeneration | 35S and pER8 | ( | ||
| Up to 2.12X increase in regeneration efficiency | None reported | maize ubiquitin | ( | |||
|
| Zea mays (B104) | 1.75X improvement in transformation | None reported | maize ubiquitin | ( | |
|
| 1.31–3.44× improvement in transformation | None reported | Rice callus-specific promoter (CSPpro) | ( | ||
|
| 6 to 12X increase in transformation efficiency | None reported | Native gene promoters PRP and SlWIND1 | ( | ||
|
| 2 to 5X increase in transformation efficiency | None reported | Native gene promoters PRP and SlWIND1 |
| ||
|
| 4X increase in transformation efficiency | None reported | Native gene promoters PRP and SlWIND1 |
| ||
|
| 4X increase in transformation efficiency | None reported | Native gene promoters PRP and SlWIND1 |
| ||
|
| Triticum aestivum (Fielder, Chinese Spring) | Significant improvement in transformation | Not reported | ZmUbi | ( | |
|
|
| Involved in somatic embryogenesis | Not specified | 35S | ( | |
|
|
| Expressed during early somatic embryogenesis | Not specified | 35S and XVE | ( | |
|
|
| Expressed during early somatic embryogenesis | Not specified | 35S and XVE | ( | |
|
|
| Change in number and quality of cotyledon embryos | Not specified | 35S | ( | |
|
|
| Promote somatic embryogenesis and organogenesis | Not specified | 35S | ( | |
|
|
| Improves transformation efficiency | Not specified |
| ( | |
|
| High transformation efficiency (>20%) | No obvious negative effects on plant growth |
| ( | ||
| Watermelon | High transformation efficiency (67.27%), | None reported | CaMV 35S | ( | ||
|
| Cannabis (Hemp) | Improved regeneration efficiency | Genotype-specific (DMG278 had highest regeneration) | CaMV 35S | ( | |
|
| Enhance regeneration efficiency | No obvious developmental defects | ZmUBI and pPLTP | ( | ||
|
| Enhance regeneration and transformation efficiencies | Abnormal development and maturation of vegetative shoots and leaves | CaMV p35S | ( | ||
|
|
| Regulates and enhances somatic embryogenesis | Not specified | CaMV 35S | ( | |
|
|
| Strong promotion of somatic embryogenesis | Not specified | CaMV 35S | ( | |
|
|
| Promotes somatic embryogenesis | Not specified | CaMV 35S | ( | |
|
|
| Promotes embryo development | Mutant embryos from all lines had abnormal suspensors | CaMV 35S | ( | |
|
|
| Induces somatic embryogenesis from asexual cells | Suspensor abnormalities in | CaMV 35S | ( | |
|
|
| Enables somatic embryo formation in cotyledon explants | Not specified | CaMV 35S | ( | |
|
|
| Promotes somatic embryogenesis | Mutants had leafy cotyledons with trichomes on the adaxial surface; defects in suspensor morphology during early embryogenesis; defective seeds (desiccation-intolerant) which had to be rescued before seed maturation. | CaMV 35S | ( | |
|
|
| Promotes somatic embryogenesis | No pleiotropy: T2 transgenic lines developed into fertile plants | 35S | ( | |
|
|
| Transit cells | Not specified | 35S | ( | |
|
|
| Induces zygotic and somatic embryogenesis | Embryogenesis in the | ( | ||
|
|
| Expressed in zygotic/somatic embryogenesis | Twisted seeds or a partially developed root apex with normal cotyledons | 35SCaMV | ( | |
|
|
| 3- to 4-fold increase in efficiency for somatic embryogenesis | No altered plant phenotype | 35S | ( | |
|
|
| Induces callus without exogenous hormones | May require optimization for shoot regeneration | 35S and chemical-inducible XVE | ( | |
|
| Maize (B104) | 3.5–6.5× improved transformation frequency; efficient CRISPR editing; no impact on plant development | Maize (B104) | ZmUbi | ( | |
|
| Promotes wound-induced | Did not validate in monocots | 35S, XVE, ProWIND1 | ( | ||
|
| Induces callus without exogenous auxin | Validated only in | CaMV 35S; XVE | ( |
| Strategy | Mechanism | Promoter(s) | Delivery method | Morphogenic regulators | Crop(s) | Key advantages | Limitations/considerations | References |
|---|---|---|---|---|---|---|---|---|
| Tissue-specific expression | MRs driven by egg cell-specific promoter | EC1 | Enables haploid induction and regulates endosperm proliferation and cellularization. | No phenotypic abnormalities or pleiotropic effects reported | ( | |||
| MRs under egg cell-specific promoter | PsASGR | Biolistic particle delivery |
| Induction of parthenogenesis | No adverse developmental effects; main limitation was technical, I.e., ensuring effective expression for inducing parthenogenesis. | ( | ||
| Seed specific | Oleosinpro: WUS 2 | Particle bombardment and |
| Involved in somatic embryogenesis and stimulating somatic embryogenesis | Chimeric and often necrotic callus which regenerated only non-transformed plants. Aberrant phenotypes, including thick, short roots | ( | ||
| Tissue-specific BBM expression | Zm-PLTPpro:: BBM and Nospro:: WUS 2 | Rapid somatic embryo formation, reduced pleiotropic effects on T0 plants | Germination problems in T1 seed (Due to WUS 2) | ( | ||||
| Tissue-specific BBM expression, Auxin inducible WUS expression | PLTPpro : HvBBM, Axig1pro:HvWUS | Hordeum vulgare (Barley) | 3× improvement in transformation efficiency; functional transcriptomic support linking auxin, meristem genes, and epigenetic regulators to SE | Requires immature embryos; genotype dependency may persist; overexpression risks need optimization | ( | |||
| Inducible expression | Heat shock induced DNA excision | CaMV 35S + HSP18.2 promoter (FRT/FLP excision system) |
|
| Expressed during the early stages of somatic embryogenesis | Normal plants with no pleiotropic effects after T-DNA excision | ( | |
| Drought inducible DNA excision | (1) RAB17M: CRE lox P sites; (2) NOS At : WUS2; (3) UBI M:BBM; (4) UBI M: GFP |
|
| Expressed during the early stages of somatic embryogenesis | Before excision, ectopic expression manifesting as secondary clusters of somatic embryos, folded leaves, and very short plantlets with thick abnormal root structures. Normal regeneration after excision | ( | ||
| Drought inducible DNA excision | (i) Zm-Rab17pro::Cre, (ii) NOSpro:: WUS 2, (iii) Zm-Ubipro:: BBM, and (iv) Zm-Ubipro::GFP |
|
| Change the number and quality of embryos in cotyledon | Not observed | ( | ||
| Desiccation-induced excision | Ubi pro:Bbm, nos pro: WUS 2 |
|
| Promote somatic embryogenesis and organogenesis | Not observed after excision | ( | ||
| Heat-induced excision | Zm-Hsp17.7pro:cre, Axig1pro: WUS 2 and Pltppro : Zm- BBM |
|
| Improve the transformation efficiency | None reported | ( | ||
| Chemical inducible promoter | maize IN2 promoter; WUS | Particle gun delivery |
|
| Expressed during the early stages of somatic embryogenesis | Not specified | ( | |
| GR inducible BBM | GR fusion gene, 35S::BBM |
|
| Expresses at specific stages of somatic embryos (torpedo and cotyledonary) | Reduced pleiotrophic effects | ( | ||
| DEX-inducible | GR fusion gene |
|
|
| 2–3.5-fold increase in shoot regeneration on hormone-free medium; high-quality embryogenic callus formed under DEX | Variation in lengths of hypocotyls and primary roots | ( | |
| GR-based inducible system | GR fusion |
|
|
| Morphologically normal mutant plants by inducer withdrawal | ( | ||
| GR-based inducible system | GR fusion |
|
|
| Induces somatic embryogenesis in leaves | ( | ||
| Auxin inducible WUS | Zm-PLTPpro:: BBM and Zm-Axig1pro:: WUS 2 |
| No deleterious pleiotropic effects | ( | ||||
| Chemical inducible promoter | XVE (estradiol-inducible) | Stable transformation via floral dip |
|
| Induces high-frequency somatic embryo formation without auxin or 2, 4-D; direct germination into fertile plants | Requires chemical inducer (estradiol); studied primarily in | ( | |
| Chemical inducible promoter | XVE (estradiol-inducible) |
|
| Enables regeneration from root tip; applicable to recalcitrant species | Requires β-estradiol; shoot formation from root segments needs cytokinin | ( | ||
| Estradiol-inducible overexpression of morphogenic regulators | Estradiol-inducible (XVE) |
|
| Promote regeneration from leaf segments and free cells | Not specified | ( | ||
| Estradiol-inducible overexpression of morphogenic regulators | Estradiol-inducible (XVE) | Demonstrated transgene expression throughout somatic seedlings | Developmental disruption of embryos; inhibited germination; no SE induction despite high expression | ( | ||||
| Jasmonate-induced |
|
| Enabled PGR-free callogenesis and somatic embryogenesis in leaf and root explants | Effectiveness depends on jasmonate responsiveness in tissue | ( | |||
| Altruistic transformation | Herbicide (GOI); No selection for MRs | PLTP:: WUS 2, with 3× viral enhancer::UBI:: BBM |
| BBM and WUS supplied in trans; only the target gene cassette was integrated | ( | |||
| GOI marker only | Pltp and 3×Enh–UBI for WUS 2/ BBM | Biolistics |
| Transient MR expression boosts genome editing without MR integration | ( | |||
| Marker for GOI; No marker for MRs | Pltp and 3×Enh–UBI for WUS 2/ BBM | cobombardment and | Co-delivered but not selected; transient expression sufficient for regeneration | ( | ||||
| Dual-vector transformation with differential selection | Pltp and 3×Enh–UBI for WUS 2/ BBM |
| MRs carried on a separate T-DNA with a different selection marker (hygromycin), while the gene of interest was selected with phosphinothricin (Bialaphos); resulting transformants lacked the MR construct | ( | ||||
| Altruistic delivery of viral and conventional T-DNA vectors | OsEf1a: | Foxtail Mosaic Virus (FoMV) and | Embryo formation improved with altruistic GRF4-GIF1 delivery (61.3%) compared to regular methods (43–50%). Using viral vectors with GRF4-GIF1 or BBM / WUS 2 boosted this further to over 75% | FoMV expression is transient, with viral transcripts disappearing within two weeks post-inoculation | ( | |||
| Excision system + Altruistic transformation | CRE/loxP-based removal of morphogenes and marker after regeneration | PLTP (WUS 2), ZmUbi (BBM), GLB1 (CRE), 35S (CRE) |
| Reduced transformation time by ~50%- Enabled transformation of recalcitrant lines- Editing of | Requires co-infection strategy; transgene-free lines must be screened; not all genotypes may respond equally well | ( |
| Strategy | Mechanism | Promoter(s) | Delivery method | Morphogenic regulators | Crop(s) | Key advantages | Limitations/considerations | References |
|---|---|---|---|---|---|---|---|---|
| Viral vector delivery | ||||||||
| Transient MR expression | FoMV replication signals (native viral promoters) | Viral vector (FoMV) via rub-inoculation | Sorghum bicolor | High embryogenic callus formation (79–75%) without genomic integration | Transient expression only; stability of long-term expression unknown | ( | ||
| Transient MR expression | FoMV native promoters | Viral vector (FoMV) | Multiple Poaceae (e.g., maize, sorghum) | Broad applicability via leaf transformation; avoids | Need follow-up on fertility and field performance |
| ||
| Altruistic delivery of viral and conventional T-DNA vectors | OsEf1a: | Foxtail Mosaic Virus (FoMV) and | maize Zm | Embryo formation improved with altruistic GRF4-GIF1 delivery (61.3%) compared to regular methods (43–50%). Using viral vectors with GRF4-GIF1 or BBM / WUS 2 boosted this further to over 75% | FoMV expression is transient, with viral transcripts disappearing within two weeks post-inoculation | ( | ||
| In planta delivery of MRs | Ubiquitin / constitutive T-DNA DR + CRISPR cassette | Tomato, Potato, Grape, other dicots | Tissue culture bypass; regenerated edited plants transmitted to next generation | Potential somatic mosaicism; limited monocot examples |
| |||
| CaMV 35S |
| Snapdragon | Promoted callus and shoot regeneration at wound sites | Some morphological alterations; stable inheritance confirmed |
| |||
| CaMV 35S |
| Tomato | Highest transformation efficiency among tested MRs | Not reported |
| |||
| CaMV 35S |
| Snapdragon | Improved transformation efficiency | Not specified |
| |||
| CaMV 35S |
| Snapdragon | Increased transformation potential | Not specified |
| |||
| CaMV 35S |
| Snapdragon | Promoted shoot regeneration at wound sites | Morphological alterations; pleiotropic effects |
| |||
| Synthetic activator (ARR12, WUS driven) | Arabidopsis | Precise control over shoot regeneration | Synthetic system requires optimization |
| ||||
| CRISPR-Combo | ||||||||
| Co-delivery of morphogenic factors and CRISPR reagents in one T-DNA cassette, for dCas9-based activation of | U6 (for sgRNA), 35S/Ubi (for Cas9) | Genome editing (e.g., | Combines DSB or base editing and gene activation in one construct; enables speed breeding and hormone-free regeneration | Requires precise sgRNA engineering; efficiency varies between constructs/crops | ( | |||
- —Bill and Melinda Gates Foundation10.13039/100000865
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Plant Molecular Biology Research · Plant tissue culture and regeneration
Introduction
1
The development of new crop varieties with enhanced traits is a crucial strategy for addressing the increasing demand for food and feed amidst climate change, biodiversity loss, declining arable land, and increased incidence of agricultural pests and diseases (Muluneh, 2021). Since marketing of the first transgenic crop in 1994 and most recently, the gene editing technology, biotech has significantly enhanced food security and catalyzed economic advancement by facilitating gene modification to incorporate desirable traits (Tripathi et al., 2022). Notably, traditional methods for in vitro transformation hinge on delivery of DNA or genome-editing reagents into explants, followed by extensive tissue culture to regenerate whole plants. These processes are time-consuming, highly genotype-dependent, and prone to somaclonal variation, thus quite inefficient (Maher et al., 2020). For example, although standard methods for Agro-mediated transformation have worked in several model crops, and select monocots and dicots, they’re only applicable in a narrow range of cultivars. Besides, they often demand laborious media optimization and stringent selection regimes to obtain transformed events. The second most-common transformation system, particle bombardment, often causes chromosomal rearrangements, cell damage and often integrates in multiple copies, and has the pre-requisite of a regenerable target tissue. On the other side, PEG-based DNA delivery to protoplasts has only been demonstrated in a few crop species (Brandizzi et al., 2025; Zhong et al., 2024). The effective development of stably transformed plants, therefore, remains a substantial bottleneck in most agriculturally important crops (Lee and Wang, 2023).
Recent studies have uncovered a distinct set of peptide regenerating factors and plant morphoregulatory genes (MRs) that actively facilitate plant regeneration. Further research demonstrated that the ectopic expression or targeted regulation of these genes improves the efficiency of plant transformation by unlocking the regenerative ability of somatic cells in previously unresponsive tissues (Debernardi et al., 2020; Hoerster et al., 2020; Jorge et al., 2021; Lowe et al., 2018; Maren et al., 2022; Xu et al., 2022; Yang et al., 2024; Lee and Wang, 2023). Characterization of the molecular mechanism of MRs has shown that they govern the biosynthetic and signaling pathways of auxins and cytokinins, thereby regulate plant regeneration processes (Gordon-Kamm et al., 2019).
According to (Maren et al., 2022), MRs encode transcription factors or signaling molecules that control cellular differentiation, embryogenesis, organogenesis, meristem identity and the overall plant regeneration process (Maren et al., 2022). For instance, the landmark study by (Lowe et al., 2016) found that the constitutive overexpression of the MRs BABY BOOM (BBM, an AP2/ERF-family transcription factor) and WUSCHEL2 (WUS2, a homeodomain transcription factor) improves maize transformation efficiencies. The MRs enables transformation of maize multiple inbred lines that were previously considered recalcitrant, with up to twenty-fold increase in shoot regeneration and highly reduced genotype specificity. The researchers reported that co-expression of BBM and WUS keeps transformed cells in a dedifferentiated, embryogenic state and promotes rapid somatic embryo formation, effectively bypassing a prolonged callus phase. This MR combination also expands the range of ‘regenerable’ and ‘transformable’ explants, including maize mature seeds (Lowe et al., 2016) and leaf bases of rice, pearl millet, maize and sorghum (Wang et al., 2023). Instead of solely relying on synthetic hormone regimes, plant transformation systems based on MRs benefit from intrinsic embryogenesis pathways by globally resetting hormonal and developmental cues in the cell. For instance, WUS and BBM upregulate LEAFY COTYLEDON1/2 (LEC1/2) and AGL15, which are master embryogenesis genes that modulate cytokinin and auxin pathways (Horstman et al., 2017). These insights indicate that an optimal combination of MRs can switch the balance from a wound response that typically results in cell death to a regeneration response to form somatic embryos and/or shoots. While BBM and WUS remain the core MRs, several other transcription factors and peptide regenerating factors, such as regeneration factor 1 (REF1), GRF-GIF chimeras, WIND1, and the WOX and PLT5 gene families have been shown to exert synergistic effects on plant regeneration (Debernardi et al., 2020; Lee and Wang, 2023; Maher et al., 2020). By illuminating the molecular underpinnings of plant regeneration and cellular totipotency, our overarching objective is to advance precision breeding technologies through plant genetic transformation from diverse vantage points.
This review provides a comprehensive overview of how MRs and peptide factors are expediting plant transformation and gene editing. We first summarize the prevailing plant transformation techniques and recent advances in genome editing to set the context of the challenges these innovations aim to solve. In Section 3, we define morphogenic regulators and detail the mechanism of action and key examples then Section 4 discusses practical strategies for deploying these regulators in transformation and editing workflows. Sections 8 and 9 have addressed the challenges and limitations of these approaches and highlight recent advances and future prospects, including emerging peptide regeneration factors like REF1 and novel tools like inducible CRISPR activation of endogenous development genes.
Advancement in gene editing technology in basic and translational research
2
Engineered sequence-specific nucleases (SSNs), which are designed to identify and modify specific regions of a genome and introduce in vivo DNA breaks, are the main tool used in genome editing. For precise plant genome engineering, three key genome editing technologies have been employed: zinc finger nuclease (ZFN), transcription activator-like effector nuclease (TALEN), and clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated protein 9 nuclease (Cas9). ZFNs and TALENs, which define target specificity through protein–DNA interactions, are costly and challenging to design. In contrast, CRISPR systems utilize RNA–DNA interactions to direct DNA targeting and cleavage, and are simple, highly efficient, and cost-effective. In recent years, CRISPR systems have become the most popular genome editing technology and are used widely across a variety of plant species (Zhu et al., 2020; Hassan et al., 2021). Since the CRISPR/Cas9 and CRISPR/Cpf1 systems offer simplicity of manipulation and targeted mutagenesis, they have been widely used in different fields. The various CRISPR reagents and new advancements that have gradually expanded the usefulness and efficiency of genome editing techniques in plants are covered in this section.
The CRISPR-Cas type-II nucleases have become widely used in genome editing research since they were discovered in Streptococcus pyogenes (SpCas9). For targeting DNA, it forms a complex with a single guide RNA (sgRNA), and for DNA recognition The Cas9-sgRNA complex attaches to the target sequence after Cas9 recognizes its PAM sequence, creating a double strand DNA break (DSB) at the target location by recognition domain (RuvC) and the nuclease domains (HNH) of the Cas9 protein. Staphylococcus aureus Cas9 (SaCa9) is a prominent and naturally occurring variation of these. High target specificity and low off-target Cas variants have been used for plant genome editing including eSpCas9 (Zhang et al., 2019a), SpCas9-HF1 (Zhang et al., 2019a; Xu et al., 2019), HypaCas9 (Xu et al., 2019), eHF1-Cas9 (Liang et al., 2018), eHypa-Cas9 (Liang et al., 2018), HiFi Cas9 (Banakar et al., 2020), SaCas9 (Wolter et al., 2018) and xCas9 (Wang et al., 2019);. Similarly, alternate PAM has been developed and used for genome editing in various crop species including SpCas9-VQR (Hu et al., 2016), SpCas9-EQR (Yamamoto et al., 2019), SpCas9-VRER (Hu et al., 2016) and St1Cas9 (Steinert et al., 2015). Flexible PAM variants including SpCas9-NG (Qin et al., 2020), SaCas9-KKH (Qin et al., 2019), ScCas9 (Wang et al., 2020a), XNG-Cas9 (Niu et al., 2020), SpRY (Li et al., 2021a), SpCas9-NRRH (Li et al., 2021a), SpCas9-NRCH (Li et al., 2021a) and SpCas9-NRTH (Li et al., 2021a).
Cas12 nucleases are the second most used Cas proteins in plants belonging to class 2 type V CRISPR system consists of two main components, crRNA and Cpf1 nuclease. Cpf1 only needs one short CRISPR RNA (crRNA, about 42 nt), is a low-molecular-weight multifunctional effector protein that is not only involved in targeted DNA cleavage, but also in the processing of pre-crRNA (Fonfara et al., 2016). Various natural variants of Cas12 have been utilized in genome editing studies, including AsCas12a (Malzahn et al., 2019; Bernabé-Orts et al., 2019), LbCas12a (Malzahn et al., 2019; Schindele and Puchta, 2020), FnCas12a (Zhang et al., 2021a), AacCas12b (Ming et al., 2020), AaCas12b (Ming et al., 2020), BthCas12b (Ming et al., 2020), BhCas12b v4 (Wu et al., 2020a), BvCas12b (Wu et al., 2020a), Lb5Cas12a (Zhang et al., 2021a), BsCas12a (Zhang et al., 2021a), and Mb2Cas12a (Zhang et al., 2021a). Engineered variants of Cas12a, including LbCas12a-RR (Li et al., 2018), bCas12a-RVR (Li et al., 2018), and FnCas12a-RVR (Zhong et al., 2018), have been investigated in plants to broaden the PAM recognition sequence. Additionally, the temperature tolerance variant of Cas12a has also been demonstrated in plants, namely enLbCas12a (Schindele and Puchta, 2020), ttLbCas12a (Huang et al., 2021) and AacCas12b (Ming et al., 2020).
The base editor can facilitate precise base changes without introducing double stranded breaks (DSBs) or the requirement of additional donor DNA templates. The base editing system comprises two variants: cytosine base editor (CBE) and adenine base editor (ABE). These base editors use a combination of a base-modifying enzyme (deaminase) with a catalytically inactive Cas9 (dCas9) or nickase (nCas9) capable of only cleaving one strand (Figure 1). The nCas9 is typically used in modern base editors due to its superior performance over dCas9-based base editors. The CBE comprises the Cas9 nickase (D10A) conjugated with a cytidine deaminase (APOBEC1) and an inhibitor of uracil DNA glycosylase (UGI), facilitating the direct conversion of C:G to T:A at DNA target site, upstream of the PAM region (Komor et al., 2017). The ABE consists of nCas9 (D10A) coupled to adenosine deaminases, enabling the direct substitution of A:T with G:C by base pairing (Gaudelli et al., 2017). A dual base editor has been reported that effectively modifies C:G to T:A and A:T to G:C simultaneously (Li et al., 2020). CGBEs (C to G base editors) represent a novel category of base editors that can facilitate the transversion of C:G to G:C within a DNA molecule (Chen et al., 2021; Kurt et al., 2021; Zhao et al., 2021). The BE3 is the most efficient and has been extensively utilized in various plant species (Mishra et al., 2020; Gürel et al., 2019). Recently, PmCDA1-CBE_V04 and A3A/Y130FCBE_V04 exhibited high editing activity and specificity while minimizing indel byproducts (Ren et al., 2021). Further, TadA8e and TadA9 are ABEs recommended for converting A/T to G/C with extensive sequence compatibility and enhanced performance (Ren et al., 2021; Yan et al., 2021).
Applications of genome editing technology for trait improvement in plants. Methods for delivering CRISPR–Cas into plant cells include Agrobacterium-mediated genetic transformation to produce genome-edited plants. Other approaches involve the use of DNA plasmids or preassembled ribonucleoprotein complexes for protoplast transfection, viral vectors that carry gRNAs and Cas nucleases in either wild-type or Cas9-expressing plants, the biolistic method for introducing editing components into plant cells, and nanoparticles for mediating the delivery of DNA constructs for CRISPR–Cas expression into plant cells. Created in BioRender (2026). https://BioRender.com/dd7rsiq.
Currently, six out of the twelve possible base alterations can be accomplished using the existing base editors. In contrast, the prime editing system facilitates all twelve categories of base substitutions, with specific and accurate insertions and deletions, in addition to base transversions. The “prime editing tool” consists of an nCas9 (H840A) fusion enzyme coupled with a reverse transcriptase (rt) guided by a prime editor guide RNA (pegRNA). PegRNA consists of two primary components: a 20 nucleotide sgRNA sequence and a reverse transcription template that features a primer binding site (PBS) containing the desired mutation information. Several versions of prime editors have been developed over the years, including Prime Editor 1 (Anzalone et al., 2019), Prime Editor 2 (Anzalone et al., 2019), Prime Editor 3 (Xu et al., 2022), Prime Editor 4max (Vu et al., 2024), Prime Editor 6c (Cao et al., 2024), pH-ePPE (Zong et al., 2022), pH-nCas9-PPE-V2 (Lin et al., 2020) and ePPEplus (Ni et al., 2023). Several different crop species have successfully been edited by prime editing, including rice (Lin et al., 2020; Xu et al., 2020; Cao et al., 2024), maize (Qiao et al., 2023), wheat (Ni et al., 2023; Lin et al., 2021), potato (Perroud et al., 2022), and tomato (Vu et al., 2024). As advances in the field continue, genome editing is gaining advantages for an increasing number of crops and traits. Crop genome modification using CRISPR technology requires several crucial components, including the availability of genome sequences, a validated target gene, an effective DNA delivery system, suitable expression systems, and an improved tissue culture regeneration system.
Method of delivery of editing components into plant tissue
3
Across multiple kingdoms of life, CRISPR/Cas genome editing has been proven to be a powerful platform for editing the genome. The difficulties in delivering CRISPR/Cas editing tools to plant cells and in recovering plants with transmissible variations bring significant technological barriers for the CRISPR/Cas utilization in plants. The introduction of genome editing reagents into intact plant tissues has been achieved mainly through Agrobacterium and particle bombardment. However, the growing trend of alternative innovative approaches, such as Agrobacterium-mediated viral-based replicons, direct introduction of genome editing components as ribonucleoprotein complexes, and nanoparticles, is gaining significant attention. The next section illustrates these broad topics with specific examples. Agrobacterium-mediated plant transformation has been continuously improved since its discovery (Barton et al., 1983). Additionally, other alternative methods have been discussed for delivering editing components to the plant cell for efficient genome editing.
Enhanced Agrobacterium- mediated gene delivery using virulent plasmids
3.1
The development of T-DNA binary vector systems (Hoekema et al., 1983) and addition of several non-oncogenic Agrobacterium strains such as LBA4404 (Ooms et al., 1981), AGL1 (Lazo et al., 1991), and EHA strains 101 and 105 (Hood et al., 1993) greatly revolutionized the use of Agrobacterium in plant biotechnology. However, alternative methods are still needed for plants that are recalcitrant to Agrobacterium transformation. Strategies such as altering the expression of virulence genes and adding copies of various Vir genes to create ‘supervirulent’ strains and ‘superbinary vectors’ (pSB1) have, therefore, increased transformation frequencies in many plants (Komari, 1990; Hiei et al., 1994; Ishida et al., 1996). The ternary pVir system, described by Anand et al. (2018), represents a simplified and improved second-generation superbinary vector. Specifically, it was developed through modifications and corrections of the original vector (pSB1), which further enhanced transformation frequency (Anand et al., 2018). Additionally, pSB1 was further modified to incorporate an improved gentamicin selectable agent to minimize the spontaneous development of resistance to tetracycline. The resulting pVir system was not only smaller and more virulent than pSB1 but also failed to confer spontaneous to tetracycline resistance (Anand et al., 2018). Such modifications have streamlined and enhanced genetic transformation in elite maize and sorghum varieties that were historically recalcitrant to Agrobacterium-mediated transformation (Anand et al., 2018; Che et al., 2018). This resulted in improved transformation frequencies (31.1%) and single-copy integrations compared to pSB1 (13.7%). Indeed, transferring the vir genes to a separate plasmid drastically reduced complexity and further streamlined the T-DNA vector construction. Furthermore, these systems have been used successfully with genome editing tools for editing purposes in various crop systems, leading to high editing efficiency (Wang et al., 2023). Helper plasmids that contain additional copies of key Agrobacterium virulence genes have been effectively utilized in various crops to enhance transformation efficiency (Anand et al., 2018; Zhang et al., 2019b; Kang et al., 2022a; Jeong et al., 2024; Aliu et al., 2024; Vandeputte et al., 2024). Additionally, the Type III Secretion System and its effector proteins from Pseudomonas syringae to supress plant immune response (Raman et al., 2022), as well as centrifugation (Ye et al., 2023) and sonication (Zhong et al., 2024) methods, have been used to enhance Agrobacterium-mediated transformation.
Biolistic-mediated gene delivery
3.2
The biolistic method is an alternative approach employed by researchers to introduce gene editing tools into plant systems that have historically proven resistant to Agrobacterium-mediated transformation. Once inside the cell, the microcarriers (tungsten or gold or silicon) coated with cargo biomolecules can transform not only the nucleus but also plastids and mitochondria, either by integrating the cargo into the genome or remaining as extrachromosomal material (Sanford, 2000). This system has many advantages, including the ability to target different tissue types and deliver large or multiple genome editing components on separate plasmids. However, the success of biolistic experiments is critically dependent on numerous parameters, including the acceleration of particles, target tissue types, microparticles size, and DNA to particles ratio, all of which must be carefully optimized to achieve efficient and precise genome editing. Biolistic delivery has been successfully employed for genome modification in a wide range of plant species, including wheat (Liang et al., 2017; Hamada et al., 2018) maize (Ainley et al., 2013a; Svitashev et al., 2015, Svitashev et al., 2016; Shi et al., 2017), rice (Sun et al., 2016) and soybean (Jacobs et al., 2015; Li et al., 2015). Recently, the flow guiding barrel (FGB) biolistic delivery method improved transient transfection efficiency by 22 times, CRISPR-Cas9 ribonucleoprotein editing efficiency by 4.5 times in onion epidermis, and viral infection efficiency by 17 times in maize seedlings. Furthermore, utilizing B104 immature embryos boosts stable transformation frequency in maize by more than tenfold and doubles the efficacy of CRISPR-Cas12a-mediated genome editing in wheat meristem tissue (Thorpe et al., 2025). The main disadvantages of this method include limited control over the distribution and penetration of microparticles, cell lethality, chromosome damage, and complex and multicopy vector DNA integration.
Virus vector-mediated gene delivery
3.3
Viruses are obligate parasites that unequivocally complete their genome replication within the host plant cells, and they frequently transmit between cells. Plant viruses-based replicons are being increasingly used as an alternative to Agrobacterium in the delivery of genome editing tools into plants, due to their ability to introduce the editing tools into plants at high levels of efficiency. The common viral components which have been replaced with editing reagents include the removal of viral coat proteins (Cody et al., 2017), movement proteins (Baltes et al., 2014), or vector-assisted transmission protein (Liu et al., 2023a). In the meantime, it will be difficult to employ such vectors for systemic or heritable modifications in situ as partially breaking down the virus genome may stop cell to cell movement. Tobacco rattle virus (TRV) possesses a broad host range and can invade to infect germline cells, making it one of the most frequently used viral vectorsand used in gene editing by multiple research groups in various plant species, including tobacco (Ellison et al., 2020) and Arabidopsis (Ghoshal et al., 2020; Nagalakshmi et al., 2022; Liu et al., 2022). Barley stripe mosaic virus (BYSMV), a positive strand RNA virus that infects many gramineae species, has been used to deliver editing reagents in various crops, including wheat (Hu et al., 2019; Li et al., 2021b; Wang et al., 2022a), maize (Hu et al., 2019), and barley (Tamilselvan-Nattar-Amutha et al., 2023). Similarly, Barley yellow striate mosaic virus (BYSMV) vector was used to deliver Cas9, and guide RNA in tobacco (Gao et al., 2019). In a similar vein, various positive RNA viruses have been utilized for genome editing, such as foxtail mosaic virus (FoMV) (Beernink et al., 2022; Mei et al., 2019), cotton leaf crumple virus (CLCrV) (Lei et al., 2022), tomato mosaic virus (ToMV) (Kaya et al., 2017), beet necrotic yellow vein virus (BNYVV) (Jiang et al., 2019), tobacco mosaic virus (Cody et al., 2017), and potato virus X (PVX) (Uranga et al., 2021).
Ribonucleoproteins -mediated gene delivery
3.4
Agrobacterium-mediated transformation or particle bombardment introduces CRISPR vector reagents into plant cells and tissues, facing regulatory obstacles due to random integration. Segregation eliminates transgenes, but it takes time and effort, making it untrainable for trees and vegetative plants. Genomic DNA is continuously exposed to CRISPR reagents during editing, inducing off-target mutations and chimeric mutants (Hashimoto et al., 2016). Plasmid-based CRISPR genome editing requires species-specific promoters, terminators, and codon modifications. Polyploid organisms may need several gRNAs, making CRISPR vector systems challenging. To address the challenges, the Cas protein and gRNAs can be preassembled into ribonucleoproteins (RNPs) for plant delivery to generate transgene-free events with reduced off-target effects (Kim et al., 2017; Liang et al., 2017). The efficiency of delivery plays a significant role in the performance of RNP-mediated editing. RNPs have previously been introduced into plants using four different techniques: lipofection, particle bombardment, electroporation, and cell transfection mediated by polyethylene glycol (PEG). Lipofection is regarded as a simple and affordable cell transfection technique (Liu et al., 2020). Another popular method for delivering RNPs into plant tissues and cells is particle bombardment. The RNP-coated particles can target explants that are commonly used for plant regeneration, such as leaf discs and embryos, with or without selection, and the explants can then be used for regeneration. In maize (Svitashev et al., 2016), wheat (Liang et al., 2017), and rice (Banakar et al., 2020), RNP-mediated genome editing through particle bombardment has been effectively demonstrated with editing efficiency less than 10% without any selection. Plasmids encoding a selective marker gene can be co-transformed into plants with RNPs to increase editing efficiency (Svitashev et al., 2016; Banakar et al., 2020). The most popular RNP-mediated delivery method is polyethylene glycol (PEG)-mediated cell transfection, which necessitates the pre-removal of the cell wall by pectinase and cellulase enzymes. The method allows for rapid evaluation of Cas systems in plant cells, and editing efficiencies can be determined between one and three days after transfection. PEG-mediated genome editing using protoplast assays have been used for many plant species including tobacco (Woo et al., 2015; Kim et al., 2017), maize (Sant’Ana et al., 2020), wheat (Liang et al., 2017), potato (Andersson et al., 2018; González et al., 2020), petunia (Yu et al., 2020), soybean (Kim and Choi, 2020), cabbage (Murovec et al., 2018), banana (Wu et al., 2020b) and rice, Arabidopsis and lettuce (Woo et al., 2015). To date, researchers have successfully edited entire plants from protoplasts in a variety of species, including rice (Barrera et al., 2025) and maize (Svitashev et al., 2016), potato (Moon et al., 2022) and wheat (Poddar et al., 2023). The rapid development of CRISPR and other biotechnologies makes RNP-mediated genetic engineering a promising platform for crop enhancement, broadening its applications in agriculture, food production, and research.
Challenges associated with plant transformation and regeneration
3.5
The success of discovery and translational research relies on the efficiency of plant transformation, which involves three critical steps: selection of appropriate explants, adoption of effective gene delivery systems, and regeneration of the complete plant from transformed cells (Lee and Wang, 2023; Syombua et al., 2021). Additional challenges encompass the restricted availability of explants, inadequate gene delivery systems, and the accessibility of transformation protocols to a limited number of species with restricted genotypes. Resistance to genetic transformation may arise from inadequate gene delivery to regenerative target cells and the limited regeneration capacity of explant material (Bidabadi and Jain, 2020; Long et al., 2022). It is often difficult for small laboratories to adopt established protocols as they require experienced technical personnel and the availability of high-quality explants (immature embryo) round the year. The outcome of plant transformation is largely dependent on cellular totipotency, which refers to the capacity of differentiated cells to dedifferentiate, attain embryonic stem cell characteristics, and regenerate into full-grown plants (Maren et al., 2022). Unlike some plant cells, not all plant cells are capable of undergoing transformation. For example, most monocot transformations utilize embryos or embryogenic calli as explants, whereas dicots use young leaves, leaf nodes, embryonic axes, shoot meristems, and leaf bases as explants (Chen et al., 2022a; Bélanger et al., 2024). The regeneration capacity of various explants exhibits significant variability among different genotypes, resulting in the regeneration of only a limited number of genotypes, which often do not include commercially and cultivated varieties, employing existing tissue culture protocols (Kausch et al., 2021; Long et al., 2022). The following sections summarize recent applications of MRs in transformation systems, as well as newly identified plant regeneration enhancers, signal peptides, and their potential role in improving transformation efficiency.
Key morphogenic regulators in plant transformation, regeneration and gene editing applications
4
Plant morphogenesis is a complex biological process that determines plant form and structure, encompassing growth, cell differentiation, and communication, and is influenced by biochemical signaling and genetic factors (Marconi and Wabnik, 2021). Foliage development is a great example of plant morphogenesis. The shoot apical meristem (SAM) produces leaves through shoot bud, which is controlled by phytohormones. Auxins and cytokinins have a major influence on the process that produces plant shoots and leaves. While cytokinin is essential for sustaining meristematic activity and cell proliferation, auxins stimulate organ initiation and patterning (Azizi et al., 2015; Kean-Galeno et al., 2024). Auxin plays a vital role in the development of the shoot apical meristem and its gradients, formed by transport and local biosynthesis, affect the spatial organization of the SAM, leading to the formation of new organs such as leaves at specified sites (Demesa-Arevalo et al., 2024). The development and function of the SAM are regulated by a complex interplay of hormones and developmental genes or regulators. Cytokinins are vital for maintaining the stem cell population in the shoot apical meristem (SAM) and promoting cell division. They work with auxin to regulate meristem activity and influence the expression of genes like WUSCHEL (WUS), essential for stem cell maintenance. Cytokinin signaling gradients, shaped by epidermal expression and transport, contribute to SAM spatial organization. The complex interaction between auxin and cytokinin affects cell fate, growth, and patterning, while gibberellins enhance cell division and elongation, contributing to plant height (Kean-Galeno et al., 2024; Demesa-Arevalo et al., 2024). Synthetic plant hormones can be used to precisely manipulate in vitro somatic embryogenesis processes, and somatic embryo development (Fehér, 2015). This model system serves as an excellent platform for investigating early plant development, pinpointing essential genes associated with embryogenesis, and identifying gene regulatory networks (Rose and Nolan, 2006; Horstman et al., 2017). During embryogenesis, somatic cells undergo multiple stages, including dedifferentiation, totipotency, differentiation, and embryo maturation, and these processes reprogramme gene expression both globally and locally in response to external cues and cellular morphogenic signals. In the past two decades, advancements in genomic technologies have facilitated the identification of essential genes that regulate somatic embryogenesis called morphoregulators (MRs) and morphogenic genes (BBM, WUS2, LEC1, LEC2, AGL15, GRFs, WOX, REF1, etc) (Figure 2). These MRs have been applied to recalcitrant genotypes, resulting in favorable outcomes for genotype-flexible transformation into a wide variety of crops (Gordon-Kamm et al., 2019; Maren et al., 2022; Chen et al., 2022a; Lee and Wang, 2023).
The diagram illustrates the process of somatic embryogenesis in plants. Number of factors involved for embryogenesis process, including transcription factors, epigenetic changes (DNA/histone methylation and histone acetylation), and hormones. The WUS/WOX is crucial for spanning hormonal balance in the shoot meristem region and assists in maintaining the shoot apical meristem (SAM) and embryonic development process. LP (Leaf primordia), BBM (BABY BOOM), WUS2 (WUSCHEL2), LEC (LEAFY COTYLEDON), AGL15 (AGAMOUS-Like 15), GRF1 (GROWTH-REGULATING FACTOR1), WOX (WUSCHEL-related homeobox) and REF1 (Regeneration Factor 1). Created in BioRender. (2026) https://BioRender.com/0vowex7.
WUSCHEL
4.1
WUSCHEL (WUS) genes are homeodomain TFs with key roles in development and regulate the maintenance of the stem cell population in the SAM, as reported in Arabidopsis (Zuo et al., 2002; Jha et al., 2020). WOX are another class of WUS-related homeobox TFs that have similar developmental roles, including stem cell identity maintenance and embryo development (Van Der Graaff et al., 2009; Wang et al., 2022b; Fambrini et al., 2022). Due to their regulatory roles in axis determination and stem cell maintenance, WUS/WOX genes are particularly useful in plant transformation systems, specifically in promoting meristem and somatic embryo development. WUS overexpression enhances somatic embryogenesis in various dicot species, including Medicago (Tvorogova et al., 2019), tobacco (Rashid et al., 2007), white spruce (Klimaszewska et al., 2010) and coffee (Arroyo-Herrera et al., 2008). Expression of ZmWUS2 promotes somatic embryogenesis in recalcitrant maize varieties (Lowe et al., 2016; Zuo et al., 2002). However, high expression levels of the WUS2 gene resulted in a friable callus and pleiotropic effects during tissue culture plant development (McFarland et al., 2023; Gordon-Kamm et al., 2019). To address the issue, the WUS2 gene was employed under the weak maize auxin-inducible (Axig1) promoter in conjunction with BBM1 under strong expression, enabling effective transformation without detrimental effects on plant development (Lowe et al., 2018). The aforementioned issue was addressed through a non-integrating ZmWUS2 based transformation system known as altruistic transformation. This method has been demonstrated in maize (Hoerster et al., 2020) and Sorghum bicolor (Che et al., 2022; Aregawi et al., 2022; Nelson-Vasilchik et al., 2022), utilizing the non-cell-autonomous movement of WUS protein via plasmodesmata to promote cell proliferation in adjacent cells (Fuchs and Lohmann, 2020). Additionally, the use of inducible site-specific recombinase CRE-loxP for the excision of morphogenetic genes (WUS2 and/or BBM) after transformation was employed to produce fertile T0 plants to tackle the aforementioned problem (Wang et al., 2020b). Further developmentally regulated promoters (Ole, Glb1, End2, and Ltp2) were found to facilitate the excision of morphogenetic genes in early embryo development. The strategy resulted in 25–100% of excised events (Wang et al., 2020b).
ZmWOX2a is another morphoregulator that does not induce pleiotropic effects. Overexpression of this gene promotes the development of somatic embryos and the formation of embryogenic calli, making it an interesting candidate for the application of more versatile maize transformation methods using immature embryos (McFarland et al., 2023). Similarly, wheat TaWOX5 surmounts genotype dependence and enhances transformation efficiency in wheat (60% across 21 varieties), rye, barley, and maize (Wang et al., 2020b). The Hairy Root-to-Shoot Conversion (HRC) method has been employed to generate shoots from hairy roots in cotton and citrus (Cui et al., 2020; Ramasamy et al., 2023). The combination of HRC and MRs approaches was used in apple, using the MdWOX5 gene under an estradiol-inducible promoter to achieve a high frequency of shoot regeneration and develop transgenic plants from initial leaf explants. Furthermore, the combination of ZmWUS2 and IPT resulted in a 29-40% increase of shoot induction frequency using HRC methods in radish (Yi et al., 2024).
In light of the importance of the WUS and WOX gene families, it is crucial to identify more suitable gene family members for use in enhancing plant regeneration and transformation. A phylogenetic analysis of WOX/WUS was conducted using the maximum likelihood method, which predicted the orthologs of these genes. These orthologs can be utilized individually or in combination, making them most suitable for transformation systems aimed at enhancing plant regeneration potential in recalcitrant crops (Youngstrom et al., 2025). Studies revealed that WUS plays a crucial role in the regulation of shoot apical meristem (SAM) maintenance, WOX2 is essential for embryonic development, and WOX5/7 regulates the maintenance of root stem cell populations (Haecker et al., 2004; Sarkar et al., 2007; Zhang et al., 2017a). Research indicates that AtWOX8 or AtWOX9 members can rescue wus null mutants (Dolzblasz et al., 2016). Additionally, WOX8/9 has been shown to induce somatic embryogenesis in tobacco and Medicago plants (Kyo et al., 2018; Tvorogova et al., 2019; Yakovleva et al., 2024). These investigations indicate that a controlled and limited expression of WUS orthologs can be an effective strategy for successfully inducing somatic embryogenesis while avoiding pleiotropic effects. WOX2 and WOX5 orthologues, on the other hand, can easily be used in the transformation of several monocot and dicot plants in a straightforward manner. Consequently, WUS orthologs may be more effectively employed through well-regulated expression methods, such as inducible promoters or excision post-somatic embryogenesis, instead of depending on robust constitutive promoters. In contrast, employing WOX2 or WOX5 members would be more straightforward, since they do not require the removal of MRs for normal plant development.
BABY BOOM
4.2
BABY BOOM (BBM) gene, a member of the AP2 (APETALA2)/ERF (ETHYLENE RESPONSIVE ELEMENT BINDING FACTOR) superfamily of transcription factors, has been gained attention in plant transformation research from its first time discovered in Brassica napus which is regulator of plant cell totipotency and produce somatic embryos in turn produce plants (Boutilier et al., 2002). This process involves somatic embryogenesis and organogenesis through the auxin signaling pathway. It has been demonstrated that ectopic expression of BBM induces somatic embryogenesis without the need for exogenous plant growth regulators or stress conditions (Boutilier et al., 2002; Horstman et al., 2017), but hormone addition improves somatic embryogenesis process (Boutilier et al., 2002; Lee and Wang, 2023). Research indicates that elevated expression of BBM, which is involved in embryo development, can occasionally result in pleiotropic effects that interfere with normal plant development (Lee and Wang, 2023; Maren et al., 2022; Chen et al., 2022a). BBM orthologues have been utilized in various crop systems to enhance somatic embryogenesis and transformation efficiency in multiple species, including Rosa canina (Yang et al., 2014a), Theobroma cacao (Florez et al., 2015), Capsicum annuum (Heidmann et al., 2011), Coffea arabica (Silva et al., 2015), Sorghum bicolor (Lowe et al., 2016; Mookkan et al., 2017), Saccharum officinarum (Lowe et al., 2016), Oryza sativa (Lowe et al., 2016) and Musa acuminata (Shivani et al., 2017). Considering the fact that WUS and BBM individually promote varying degrees of somatic embryogenesis, it has been recommended to combine both MRs in order to increase the rate of genotype-independent transformation of maize using BBM and WUS2 genes (Lowe et al., 2016). The Lowe et al. group has developed a strategy that expresses BBM under the maize phospholipid transferase protein promoter (PLTP) and WUS2 under the maize auxin-inducible gene 1 promoter (Axig1), which promotes high transformation efficiency in several elite maize varieties (Lowe et al., 2018). Monocot transformation primarily utilizes immature embryo explants. Generally, monocot transformation uses immature embryo explants. Having year-round access to immature embryo explants makes the transformation process difficult. Since leaf bases maintain embryonic cell mass, Wang et al. (2023) developed a leaf-based transformation system using the combination of ZmBBM and ZmWUS2 expression system and established the system and successfully generated plants in different crops species including teff, maize, sorghum, foxtail millet, pearl millet, rye, barley, rice. Additionally, the ZmBBM1 and ZmWUS2 expression systems have been used to develop highly efficient transformants in hexaploid wheat (Johnson et al., 2023). Based on the results of these studies, it can be concluded that the combination of growth regulators is capable of promoting high efficiency transformation and further overcoming the barriers to genetic transformation in numerous elite crops, which have previously been difficult to transform.
GRF-GIF
4.3
Plant growth-regulating factors (GRFs) play an important role in regulating plant growth, including the development of leaves (Horiguchi et al., 2005; Kim and Lee, 2006; Omidbakhshfard et al., 2015). GRFs physically interact with GRF-interacting factors (GIFs), a group of transcriptional coactivators, forming a complex that recruits chromatin remodeling complexes (SWI2/SNF2) to facilitate chromatin opening and enhance transcription, thereby promoting cell division and leaf development (Vercruyssen et al., 2014; Debernardi et al., 2014, Debernardi et al., 2020). The expression of various GRFs in plants is regulated by the microRNA miR396. GRFs exhibit higher expression levels in actively growing tissues, such as roots and shoots, particularly in regions of cell proliferation (Kim et al., 2003; Rodriguez et al., 2010). In land plants, there are typically 8–20 GRF members present in their genomes, which is highly conserved (Omidbakhshfard et al., 2015). Research indicates that GRF5 facilitates organogenesis in various dicotyledonous plant species (Kong et al., 2020). A strong expression of the GRF5-mediated transformation and regeneration system has been developed in multiple dicot and monocot plant species, showing improved regeneration efficiencies in soybean, canola, sunflower, sugar beet, and maize (Kong et al., 2020). GRFs constitute a multi-gene family within each crop genome and exhibit diverse functions. Arabidopsis GRF5 overexpression significantly improved sugar beet transformation, however sugar beet GRF5 showed no significant improvement. Furthermore, the overexpression of AtGRF5 did not exhibit pleiotropic effects in canola and sugar beets; however, it was occasionally linked to abnormal root development in soybeans (Kong et al., 2020). Studies indicates that the overexpression of GRF5–GIF1 fusion product boosts both organogenesis and embryogenesis, demonstrating greater efficiency in plant regeneration and transformation compared to the overexpression of AtGRF5 alone (Debernardi et al., 2020; Kong et al., 2020).
Study conducted by Debernardi and coworkers, developed a wheat genome editing construct using a miR396 resistant variant of wheat GRF4 in combination with GIF1 under strong expression of maize ubiquitin promoters. The TaGRF4-TaGIF1 chimera enhanced regeneration and transformation efficiency by approximately 7.8-fold when utilizing wheat immature embryos as explants (Debernardi et al., 2020). Additionally, the study showed that chimeric TaGRF4-TaGIF1 constructs improved regeneration efficiency in Triticum durum, Triticum aestivum, triticale, and rice. Interestingly, Debernardi et al. (2020) found that TaGRF4-TaGIF1 chimeras enhanced shoot regeneration without including cytokinin in their media for shoot induction. In contrast to BBM or WUS-mediated transformation, the overexpression of GRF orthologs does not exhibit pleiotropic effects and results in the generation of fertile transformed plants (Debernardi et al., 2020; Kong et al., 2020; Qiu et al., 2022). In recent studies, the Agrobacterium ternary vector system was combined with GRF-GIF chimeras, which resulted in an additive effect in the transformation of sorghum and maize (Li et al., 2024; Vandeputte et al., 2024). The TaGRF4-OsGIF1 chimera construct in sorghum, regulated by the Arabidopsis UBQ10 promoter, was combined with the pVS1-VIR2 ternary helper plasmid (Zhang et al., 2019b), resulting in a 7.71-fold increase in transformation efficiency (Li et al., 2024). It has been shown that using the pVS1-VIR2 ternary helper in maize B104 transformations with immature embryos increases the transformation frequency from 2.3 to 8.1% (3.5-fold), and adding ZmGRF1-ZmGIF1 chimeras increases the transformation frequency by another 3.5-fold from 13.6 to 47.4% (Vandeputte et al., 2024). The GRF4-GIF1 fusion demonstrated functionality in dicots as well. GRF4-GIF1 chimeras showed about a five-fold increase in regeneration rate in citrus or grape (Debernardi et al., 2020). Besides the GRF4-GIF1 chimera, other GRF-GIF fusion proteins, such as GmGRF3-GmGIF1, have been effectively utilized to improve transformation in soybean (Zhao et al., 2025), grape (VvGRF4-VvGIF1), and tomato (SlGRF8-SlGIF4) and SlGRF12-SlGIF4 chimeras in lettuce (Bull et al., 2023). The studies indicate that crop-specific combinations of GRF-GIF represent a promising approach to enhance regeneration efficiency in various monocot and dicot species.
Other class of morphoregulators
4.4
A family of LEAFY COTYLEDONs genes called LEC1, LEC2, and FUS3 (FUSCA3) function as transcription factors crucial for embryogenesis and maturation processes (Chen and Du, 2022). LEC transcription factors are involved in embryonic development, seed maturation, and stalk growth in Arabidopsis thaliana. LECs and several other transcription factors establish a complex network that regulates numerous facets of plant growth and development. LEAFY COTYLEDONs (LEC1, LEC2, and FUSCA3) are the key genes in plant embryonic development (Chen and Du, 2022; Guo et al., 2022). During embryonic morphogenesis, the LEAFY COTYLEDONs determine the fate of stalk cells and control cotyledon characteristics (Chen and Du, 2022). LEAFY COTYLEDONs (LEC1, LEC2, and FUSCA3) are essential genes in plant embryonic development that interact to activate target genes and facilitate the development process in plants (Santos-Mendoza et al., 2008; Chen and Du, 2022). LEC1 promotes embryonic development and activates maturation and storage genes, as well as lipid accumulation in vegetative organs when expressed ectopically (Mu et al., 2008; Manan et al., 2023; Zhang et al., 2024a). Previous study indicates that 35S:PaLEC1-GFP transgenic lines induce various types of somatic embryonic structures and tissue clusters containing multiple shoot apical meristems (Chen et al., 2019). Citrus sinensis LEC1 overexpression in sweet orange resulted in the formation of embryo-like structures in the typically recalcitrant epicotyls after one month, and subsequently, after an additional two months on elongation medium, shoots with aberrant leaves were produced (Zhu et al., 2014). Moreover, the overexpression of PaHAP3A, a member of the LEC1/L1L family, did not lead to the formation of ectopic somatic embryos in vegetative tissues (Uddenberg et al., 2016). Transgenic lines introducing 35S::AtLEC2 into lec2–1 and lec2–5 mutant lines of Arabidopsis, leading to the formation of ectopic somatic embryos capable of germination and plant production (Stone et al., 2001). LEC2 appears to promote more complete somatic embryo formation than LEC1, L1L, or FUS3. This aligns with evidence suggesting that LEC2 operates upstream of and activates both LEC1 and FUS3. However, the resultant plant phenotypes were abnormal, and the authors did not address fertility (Santos Mendoza et al., 2005). Consequently, the existing fragmented research on the role of LECs and their molecular mechanisms in linking embryonic and vegetative growth periods with the reproductive stage requires increased attention in future studies.
SOMATIC EMBRYOGENESIS RECEPTOR KINASE (SERKs) belongs to the leucine-rich repeat receptor-like kinase (LRR-RLK) family, play vital roles in morphological development, stress and defense responses, and signal transduction (Arimarsetiowati et al., 2024). The SERK gene is pivotal in embryo formation from single somatic cells. SERK genes were first linked to somatic embryogenesis, with later studies showing their role in somatic embryogenesis and organogenesis across various species including sunflower (Thomas et al., 2004), Ocotea (Santa-Catarina et al., 2004), citrus (Shimada et al., 2005), grapevine (Schellenbaum et al., 2008), potato (Sharma et al., 2008), wheat (Singla et al., 2008) banana (Huang et al., 2010). In vanilla, enhanced SERK expression is linked to embryogenesis (Ramírez-Mosqueda et al., 2018). SERK1 overexpression of the plant gene results in enhanced formation of somatic in Arabidopsis without changes in plant phenotype (Hecht et al., 2001). The overexpression of the Coffea canephora SERK1 gene during in-vitro somatic embryogenesis significantly boosts the production of somatic embryos, increasing it by twofold (Pérez-Pascual et al., 2018). AGL15, a component of the SERK1 protein complex, demonstrated a response similar to that of SERK1 (Karlova et al., 2006). Overexpression of the AGL15 gene enhanced secondary somatic embryo formation in Arabidopsis (Harding et al., 2003), boosted somatic embryo production in soybean (Thakare et al., 2008), and further improved embryogenic callus formation in cotton (Yang et al., 2014b). Additional investigation into the biological roles of the SERK gene in crops, along with its validation, would improve the development of effective regeneration systems, thereby enhancing understanding and application of transformation techniques.
MRs have the ability to induce the development of axillary meristems along with the development of shoots and roots. Several basic helix-loop-helix transcription factors are involved in the regulation of axillary meristems. It has been shown that the LAX PALICLE1 of wheat (TaLAX1) and rice (OsLAX1) plays a significant role in the formation of axillary meristems and grain production (Gallavotti et al., 2004; Oikawa and Kyozuka, 2009). There was a considerable increase in regeneration frequency when the orthologs of TaLAX1 were overexpressed in both maize and soybean, suggesting that TaLAX1 and its orthologs could be manipulated to improve transformation in both monocots and dicots (Yang et al., 2024). By overexpressing TaLAX1a in wheat varieties, it was found that florets per spikelet increased, regeneration improved, and gene editing frequencies increased as well (Yu et al., 2024). Based on the results of these studies, it appears that overexpression of TaLAX1 results in enhanced gene expression of TaGRFs and TaGIF1, which ultimately improve regeneration and transformation in wheat, as observed by Debernardi et al. (2020).
KNOTTED1 (KN1) is a homeodomain transcription factor that plays a crucial role in the maintenance of shoot stem cells in plants (Kitagawa et al., 2022). Tobacco plants that overexpression ZmKN1 constitutively can dramatically enhance shoot organogenesis compared to control ones (Luo et al., 2006). Tobacco plants overexpressing NTH15 and NTH20 exhibited pleiotropic phenotypes but also formed ectopic shoot meristems on the leaves to some extent. Nonetheless, the other members exhibited significant abnormalities in tobacco (Nishimura et al., 2000). A KNOTTED1-like homeobox (KNOX) gene, OSH1, is induced by cytokinin, leading to the regeneration of shoots from calli mass in rice (Naruse et al., 2018).
AGAMOUS-LIKE15 (AGL15), which is a MADS-domain protein, is a crucial player both in Arabidopsis and soybean in the development of the somatic embryo (Zheng et al., 2013). It has been demonstrated that ectopic constitutive expression of AGL15 promotes somatic embryogenesis in Arabidopsis, soybean, and cotton (Harding et al., 2003; Thakare et al., 2008). In contrast, mutants of AGL15 and its close relative AGL18 significantly impede somatic embryogenesis in Arabidopsis (Zheng and Perry, 2014). In soybeans and Arabidopsis, AGL15 and AGL18 also promote somatic embryonic development when expressed ectopically (Zheng and Perry, 2014; Paul et al., 2021).
Utilization of small peptides for plant regeneration
4.5
Plants use diverse regeneration strategies after injury, primarily by regenerating their shoot or root apical meristem following localized damage. This enables the formation of new organs or entire plants from small tissue fragments or individual cells through de novo organogenesis or somatic embryogenesis. De novo organogenesis reactivates cellular proliferation at wound sites that aids regeneration (Yang et al., 2024). WOUND-INDUCED DEDIFFERENTIATION (WIND) AP2/ERF transcription factors, including WIND1 and its homologs WIND2, WIND3, and WIND4, are key regulators of wound-induced cellular reprogramming. These factors promote dynamic transcriptional changes and regulate multiple physiological responses following wounding. WIND1 facilitates callus formation and shoot organogenesis through the direct activation of a shoot meristem regulator (Iwase et al., 2011, Iwase et al., 2017). Ectopic expression of AtWIND1 in B. napus increased shoot regeneration rate by 24- to 47-fold from segments of hypocotyl (Iwase et al., 2015). WIND1 is involved in plant regeneration and functions downstream of the REF1-PORK1 module. The transgenic overexpression of ZmWIND1 led to a 3–4 fold increase in callus induction in maize (Jiang et al., 2024a). Additionally, ESR1 functions downstream of WIND1 and ESR1 does not provide feedback to cytokinin-responsive genes (Iwase et al., 2011). Ectopic expression of ESR1 or its paralog ESR2 alone can induce shoot organogenesis (Ikeuchi et al., 2019) In Arabidopsis, ESR1 expression increased shoot regeneration efficiency by 18.2% via the activation of CUC1, WUS, and STM (Iwase et al., 2021). Researchers found that stable transgenic tomatoes and peppers expressing PLT1/WOX5/WIND1 can boost regeneration efficiency without external hormones (Wittmer et al., 2025). Further research on WIND could help identify genes directly targeted by WIND1, shedding light on how WIND1-mediated signaling controls the transcription of key regulators in regeneration and uncovering a critical transcriptional mechanism linking the wound response to organ regeneration in plants.
Systemin and its precursor gene PROSYSTEMIN (PRS) mainly regulate systemic defense responses. CRISPR-Cas9-mediated knockout mutants of PRS and the systemin lacked systemic defense responses (Yang et al., 2024). Yang and coworkers show that reducing the tomato Pep (SIPep) precursor gene or its receptor gene eliminates regeneration capacity in wound-induced callus formation and shoot regeneration. In contrast, overexpression enhances regeneration capacity. Exogenous application of the SlPep peptide also significantly boosts regeneration capacity. Subsequently, SlPep was named REGENERATION FACTOR1 (REF1) a plant-derived polypeptide that functions as a localized wound signal molecule, playing a pivotal role in tissue regeneration. The REF1 peptide has been demonstrated to boost regeneration and transformation efficiencies in maize, soybean and wheat, suggesting its potential to significantly improve the recalcitrant crop regeneration and transformation processes (Yang et al., 2024). REF1 signaling activates SlWIND1, additionally, SlWIND1 amplifies the REF1 signal by promoting the transcription of the REF1 precursor gene. Thus, REF1 functions as a local wound signal that supports plant regeneration. REF1 activates downstream MAPK cascades through strong binding to its receptor ORTHOLOG RECEPTOR-LIKE KINASE 1 (PORK1), resulting in the expression of SlWIND1. SlWIND1 facilitates callus development and organ regeneration through its interaction with wound-responsive elements in the proline-rich protein (PRP) promoter, thereby establishing a “REF1-PORK1-WIND1-PRP” positive feedback loop. This loop enhances regeneration signals and preserves equilibrium in cells (Yang et al., 2024). A dose-dependent effect was noted with the exogenous application of the REF1 peptide, which enhanced callus formation and shoot regeneration in prp and pork1 mutants, while also addressing regeneration defects in these mutants (Yang et al., 2024; Schaller, 2024). Recent studies indicate that exogenous supplementation of 10 nmol CaREF1 (10 nmol/L) enhanced regeneration by 2.1-fold and increased shooting frequency by 4.4-fold, resulting in 100% editing efficiency in CRISPR/Cas9 integrated pepper plants (Wang et al., 2025). The new generation approach in REF1 application offers a robust platform for molecular breeding, especially in recalcitrant crops like soybean, wheat, and maize and enhances genetic transformation efficiency by 4–5 times, addressing the transformation bottleneck in these economically important species for food security (Trösch, 2024). The identification of REF1 offers a viable approach to augment the regeneration potential of recalcitrant crops, consequently enhancing their transformation efficiency.
Small signaling peptides are vital for plant growth, development, and environmental responsiveness, with the CLAVATA3/EMBRYO SURROUNDING REGION-RELATED (CLE) family being one of the most studied (Kang et al., 2022b). De novo shoot regeneration involves forming new shoots from explants lacking pre-existing shoot meristems, requiring coordination among signaling molecules and transcription factors like CLE peptides and WUSCHEL (WUS). CLE peptides regulate stem cell homeostasis, vascular differentiation, and root cap development, and are promising targets for improving genetic transformation and genome editing efficiency (Kang et al., 2022b; Niu et al., 2025). CLE genes encode precursor proteins with an N-terminal signal sequence for secretion, a variable central region, and a conserved C-terminal CLE domain, often modified post-translationally to form functional polypeptides (Ji et al., 2025). These genes are crucial for stem cell regulation (Kang et al., 2022b). The expression of CLE varies during the phases of shoot regeneration (Wang et al., 2022c).The transcription of CLE1-CLE7 genes is markedly increased by callus- or shoot-inducing medium (Kang et al., 2022b). CLE1-CLE7 (Kang et al., 2022b) and CLE9/10 (Glazunova et al., 2025) peptides negatively impact shoot regeneration. While cle1–7 septuple mutant exhibits enhanced regeneration capacity and an increased number of adventitious shoots formation (Kang et al., 2022b). Mutations in the single cle locus have minimal impact on de novo shoot regeneration, while both cle4 and cle7 mutants demonstrate enhanced de novo shoot formation (Kang et al., 2022b). The overexpression of CLE4/CLE7 significantly inhibits regeneration (Kang et al., 2022b). Exogenous administration of synthetic CLE peptides (CLE1–7) exhibits a dose-dependent reduction of de novo shoot regeneration. Recent research suggests that CLE9/10 peptides suppress de novo shoot regeneration but do not influence callus formation (Glazunova et al., 2025). The overexpression of CLE41/42 promotes axillary bud formation, while the external application of either tracheary element differentiation inhibitory factor (TDIF) or CLE42 peptide to the wild type similarly induces an increase in bud emergence (Yu et al., 2025). CLE peptides facilitate signaling via CLV1 and BAM1 receptors, establishing a CLE-WUS regulatory axis that inhibits WUS expression and restricts adventitious shoot formation (Zhang et al., 2024b). Inhibition of CLE peptides alleviates this suppression, thereby improving regeneration and the efficiency of genetic transformation. CLE signaling modules, typically consisting of CLE peptides, receptor-like kinases (RLKs), and WUSCHEL-related HOMEOBOX (WOX) transcription factors, are key regulators of stem cell homeostasis in various plant meristems, including shoot apical, root apical, axillary, vascular, and nodule meristems (Olsson et al., 2019; Fletcher, 2020). Mutants of clv1 and bam1 exhibit enhanced shoot regeneration abilities and demonstrate insensitivity to the inhibitory effects of CLE1-CLE7 peptides on shoot regeneration in arabidopsis. This confirms the CLE1-CLE7 signal negatively regulates de novo shoot regeneration via CLAVATA1 (CLV1) and BARELY ANY MERISTEM1 (BAM1) receptors (Glazunova et al., 2025). Furthermore, the CLE-CLV1/BAM1 module suppresses the expression of WUSCHEL, a critical regulator regeneration, thereby regulating the possibility for shoot formation (Shpak and Uzair, 2025).
Methods to discover novel regeneration morphoregulators
4.6
RNA-seq, ATAC-seq, and CUT&Tag are effective methodologies employed to examine the interactions among multiple levels of cellular information within cells. RNA-seq offers insights into gene expression, while ATAC-seq identifies accessible regions of chromatin, suggesting possible regulatory elements. CUT&Tag technology demonstrates chromatin accessibility while targeting specific proteins or histone modifications (Ouyang et al., 2021). Incorporating these methods helps researchers understand gene regulation and cellular function. In wheat, RNA-seq, ATAC-seq, and CUT&Tag have been used to identify candidate MRs to understand transcriptional regulatory networks that regulate embryonic callus formation and organogenesis (Liu et al., 2023b). This study revealed 446 core TFs involved in regeneration, as well as transcriptomic and epigenetic changes during callus maturation and shoot regeneration. As a result of the comparison of these 446 core TFs between Arabidopsis and wheat transcriptomes, TaDOF3.4 and TaDOF5.6 were identified as candidate MRs for wheat varieties with high and low regeneration genotypes (Liu et al., 2023b). In subsequent experiments, overexpression of the TaDOF5.6 or TaDOF3.4 genes under the maize Ubi promoter significantly improved wheat transformation efficiency (Fielder and Jimai22), which displayed a normal plant phenotype and seed production rate, indicating their potential use to boost wheat transformation (Liu et al., 2023b). Further, according to a recent study, 53 plant MRs have homologs in a wide variety of species, and their expression increases during embryonic development, embryogenic callus formation, and somatic embryogenesis (Jiang et al., 2024a), including ZmBBM, TaWOX5a, TaDOF5.6, which have been previously shown to improve transformation efficiency (Liu et al., 2023b; Lowe et al., 2016, Lowe et al., 2018; Wang et al., 2023, Wang et al., 2022b). Based on the results of this study, a novel morphoregulator of the ERF/AP2 TF family member is identified as maize WIND-INDUCED DIFFERENTIATION 1 (ZmWIND1) (Jiang et al., 2024a). It has been reported that the Arabidopsis ortholog of ZmWIND1 (AtWIND1) promotes callus formation and organogenesis through direct activation of ENHANCER OF SHOOT REGENERATION1 (ESR1) (Iwase et al., 2017). ZmWIND1 is negatively regulated by ZmSAUR15, which inhibits embryonic callus induction in maize. It was found that the loss of ZmSAUR15 function was associated with an increase in ZmWIND1 expression, which was associated with an increase in embryonic callus formation (Wang et al., 2022d). When ZmWIND1 was strongly overexpressed, regeneration and transformation frequencies were increased by approximately two to three-fold in two maize varieties (Jiang et al., 2024a).
Strategies for using morphogenic regulators in transformation and gene editing
5
While the identification and characterization of developmental regulators that could efficiently enhance the transformation efficiency of recalcitrant species was a significant breakthrough, the greatest bottleneck was their efficient deployment in a way that harness the benefits but reduces the associated tradeoffs. For instance, numerous studies reported that the constitutive overexpression of MRs resulted in detrimental effects and developmental abnormalities (Gordon-Kamm et al., 2019). As such, researchers have come up with the strategies outlined below for delivering, expressing and regulating MRs, including constitutive vs. tissue-specific promoters, transient expression methods, inducible systems, and RNA-based delivery (Aesaert et al., 2022; Wang et al., 2020a) (Figure 3).
Deployment strategies for morphoregulatory (MR) genes to enhance plant transformation and genome editing. The illustration summarizes five approaches used to maximize regeneration efficiency while minimizing pleiotropic effects associated with morphogenic gene expression. These include: (i) constitutive or tissue-specific promoters for spatial control of MR expression; (ii) transient expression methods such as altruistic transformation or direct delivery; (iii) inducible promoter systems triggered by chemical, heat, or light stimuli; (iv) RNA-based delivery systems utilizing CRISPR activation of endogenous MR genes; and (v) de novo meristem induction through transient MR expression in somatic cells. Image generated on Canva (https://www.canva.com/design/DAGqZjdHQMY/8sZdpSBeaoTT-frOpwBhLQ/edit).
Overexpression of MRs under constitutive promoters and spatial/temporal control via tissue-specific promoters and excision
5.1
The most robust, simplest and widely applicable strategy involves inclusion of the MRs expression cassette in the T-DNA of plant transformation vectors and expressing them under strong constitute promoters such as the maize Ubiquitin1 (ZmUbi) or the cauliflower mosaic virus (CaMV) 35S promoter. This strategy offers the advantage of simplicity of delivery and immediate robust expression since every plant tissue expresses constitutive promoters (Gordon-Kamm et al., 2019). For instance, Lowe et al. (2016) demonstrated enhanced recovery of transgenic plants, particularly in recalcitrant or marginally transformable maize, rice, sorghum, and sugarcane varieties, following constitutive expression of WUS and BBM under the maize ubiquitin promoter. Even more interesting is that the procedure enabled direct Agrobacterium-mediated transformation of mature seed-derived embryo axis or leaf segments, without an intervening callus or meristem culture step. However, most of the regenerated plants had undesired pleiotropic effects ranging from altered lateral growth to sterility, since the MRs remained active beyond the regeneration stage (Jiang et al., 2024b) (Table 1).
To mitigate these challenges, researchers have resorted to expressing the MRs only during early growth stages during which they are necessary then restricting their expression in later reproductive stages or entirely eliminating them. One such strategy is the use of tissue specific promoters; for instance, Lowe et al. (2018) regenerated perfectly normal and fertile transgenic maize plants by driving BBM under the maize Phospholipid Transfer Protein (PLTP) promoter and WUS under the auxin-inducible Axig1 promoter. Notably, PLTP is only active in dividing embryogenic cells such as in an immature embryo callus while Axig1 is only active under high auxin levels such as during the callus phase (Chu et al., 2019; Lowe et al., 2018).
Another strategy for spatial/temporal control via promoters involves the use of developmentally regulated promoters such as Oleosin, Glb1, End2, and Ltp2 to drive Cre (inducible site-specific recombinase) enabled excision of morphogenic genes in early embryo development. In the study by Wang et al. (2020a), this strategy produced excised events at a rate of 25–100%. The same study also reported a different strategy utilizing an excision-activated selectable marker that produced excised events at a rate of 53–68%. Similarly (Mookkan et al., 2017), demonstrated desiccation induction of RAB17_M_: CRE expression to remove the WUS2 and BBM transcriptional genes from the transgenic embryogenic callus before transfer to the regeneration medium. The ideal promoter drives enough expression during the regeneration window, but little to no expression in later plant development (Table 2).
Transient expression of MRs for genetic transformation and genome editing
5.2
Ensuring that MRs do not express stably in the host genome offers another popular and effective way of sidestepping their lasting drawbacks. As such, the MRs only act during the early transformation stages but are absent in the transgenic plant. Some of the methods proven to aid in this transient set up include altruistic transformation, use of viral vectors, in planta transformation, plasmid-based transient expression, and direct delivery as a protein.
Among these, co-culture with a morphogene vector that doesn’t integrate (altruistic transformation) stands as a favorite in monocots. This involves co-delivering the construct that harbors the gene of interest (GOI) with another that carries a morphogenic construct but has a different selectable marker. Previously, a method using a non-integrating WUS2 gene approach recovered fertile T0 plants free of morphogenic genes (Hoerster et al., 2020; Syombua et al., 2024). The studies demonstrated that co-infecting with two Agrobacterium strains, one with a WUS2 expression cassette, and the other with a combination of both selectable and visual marker cassettes, produced transformed T0 plants that contained only a single copy of the selectable marker T-DNA, without the integration of WUS2. However, it is important to optimize the process to determine the optimal ratio of the two Agrobacterium strains that enables modest recovery of selectable marker-containing T0 plants that do not contain the morphogene (Kang et al., 2023).
Use of MRs for de novo meristem induction
5.3
While conventional transformation methods target pre-existing shoot apical meristems (SAMs) and axillary buds (Zhang et al., 2017b), recent breakthroughs have demonstrated that MRs can transiently induce the formation of new meristematic tissues directly from somatic cells (Table 3). First demonstrated by (Maher et al., 2020), this novel strategy involves co-delivering MRs, in this case WUSCHEL2 (WUS2), SHOOT MERISTEMLESS (STM), and the cytokinin biosynthesis gene ISOPENTENYL TRANSFERASE (IPT), along with genome editing reagents into somatic cells. The transient expression of these factors in Nicotiana benthamiana promoted the formation of new meristems at the infection sites and genome-edited shoots were subsequently regenerated. Notably, these edits were stably transmitted to the next generation (Maher et al., 2020).
The broad applicability of this method has since been demonstrated though its successful application in tomato, potato, grape, two brassica cabbage varieties (Brassica rapa var. Bok choy and Pei‐Tsai) and citrus, among other crops (Kelly et al., 2025; Lian et al., 2022). Lian et al. showed that PLETHORA5 (PLT5), WOUND INDUCED DEDIFFERENTIATION 1 (WIND1), and WUS significantly enhanced the efficiency of in planta transformation. The study further reported that PLT5 induces callus formation and shoot regeneration at wound sites on aerial tissues, enables stable transgene inheritance in tomato and cabbage progeny and facilitates in vitro somatic embryogenesis in sweet pepper (Lian et al., 2022). A more recent report is the IPGEC (in planta genome editing in citrus) system (Kelly et al., 2025), an in planta approach for genome editing in soil‐grown citrus plants via direct transformation of young seedlings. The Agrobacterium-based editing system was designed to transiently co‐deliver three gene modules, including (i) MRs (ii) Cas9 and sgRNAs targeting the carotenoid biosynthetic gene PHYTOENE DESATURASE (PDS), and (iii) accessory genes to enhance T-DNA delivery. Notably, the integrated system significantly improved de novo shoot induction and regeneration efficiency of albino edited tissue (Kelly et al., 2025).
In the agroinfiltration-based Fast-TrACC method (Cody et al., 2023), Agrobacterium is briefly applied to intact seedlings or explants to deliver MRs and CRISPR, then the bacteria are washed off. Following a transient co-culture, the plant tissue forms edited shoots, hence bypassing laborious tissue culture processes. Such In planta transformation systems and associated de novo meristem induction approaches offers a powerful alternative to tissue culture dependent regeneration and facilitates genome engineering even in recalcitrant species (Zhang et al., 2017c).
Use of inducible promoter systems to fine-tune expression
5.4
Inducible expression systems that remain off until the availability of a specific signal allow temporal control over the trade-off between high-level transgene expression and its pleiotropic effects on plant growth. For example, chemical, heat shock, or light inducers can be activated at specific developmental stages to initiate regeneration and then switched off to allow normal plant development (Jiang et al., 2024b). This strategy minimizes off-target effects and developmental defects. In maize, Wang et al. (2020a) utilized heat shock promoters to drive Cre recombinase expression in transgenic calli; a brief heat pulse was then applied during early embryo development to excise the WUS/BBM genes from the T-DNA. This allowed for regeneration of fertile plants free of the morphogenic transgenes (Wang et al., 2020a).
The XVE chemical-inducible system is another common inducible promoter system; it uses a chimeric transcription factor activated in the presence of the estrogen analog β-estradiol. Upon activation, it drives GOI expression under the LexA operator promoter. In cotton, for instance, placing GhLBD18 (LATERAL ORGAN BOUNDARIES DOMAIN 18) transcription factor under the XVE inducible promoter (pER8) ensured that callus proliferation only occurred when estradiol was added to the medium. Similarly, WUS could be placed under a dexamethasone-inducible promoter then apply dex only for a short duration to spark regeneration (Omelina et al., 2022; Xinying et al., 2024). Another approach is light-inducible systems, such as using a plant usable light-switch elements (PULSE) or a UVresponsive promoter to allow spatial control by illuminating only part of a tissue to induce local organ formation (Ochoa-Fernandez et al., 2020).
Transient activation of MRs via CRISPR and RNA guides
5.5
Rather than introducing DNA cassettes encoding MRs, RNA-based approaches deliver synthetic RNA molecules to influence morphogenic pathways. Thus, this RNA-guided approach does not introduce any new coding sequences but achieves MR overexpression by turbocharging the plant’s existing morphogenic genes (Table 3). For instance, a recent preprint (Butler et al., 2025) developed an inducible CRISPR activation system where multiplexed guide RNAs were used to turn on native morphogenic genes in planta. Instead of adding an extra WUS gene, the system employs a dCas9 activator and guide RNA to turn on the plant’s endogenous WUS gene. Notably, the study reports regeneration of fluorescent shoots from both conventional and viral vectors, providing evidence on the broad delivery strategies of RNA-based MRs for improving plant transformation (Butler et al., 2025). Similarly, a previous study used a chemically inducible CRISPR activation tool to activate endogenous BBM and LEC2, and achieved enhanced regeneration efficiencies in alfalfa, woodland strawberry, and sheepgrass (Zhang et al., 2024c).
A CRISPR-Combo approach was recently shown to aid the editing of one gene and simultaneous activation of another in the same cell using a dual gRNA system (Pan et al., 2022b). The system incorporates catalytically active Cas9 and transcriptional activation domains to allow one gRNA to direct editing of the GOI and the other upregulates gene transcription by targeting the promoter region of a different gene. Pan et al. first used CRISPR-Combo to induce early flowering in Arabidopsis by editing trait genes and simultaneously activating the flowering gene FT. The approach was then extended to rice and poplar, where editing of a GOI and simultaneous transient activation of the endogenous WOX11 resulted in higher regeneration efficiencies. Since the activation component depends solely on gRNAs and no transgene integration, it mitigated the risk of pleiotropic effects and enabled the recovery of edited, transgene-free plants in subsequent generations (Pan et al., 2022b).
Challenges and limitations
6
Developmental defects: The constitutive expression of some morphogenes causes aberrant phenotypes, such as organ malformation, fasciation, and sterility (Lowe et al., 2016). As such, the need to silence the genes’ activity post regeneration is imperative. Current mitigation strategies include the use of inducible promoters such as estrogen or heat shock inducible and excision mechanisms such as Cre/lox from bacteriophage P1, Flp/frt from Saccharomyces cerevisiae, R/RS from Zygosaccharomyces rouxii, and Gin/gix from bacteriophage (Gordon-Kamm et al., 2019; Wang et al., 2020b). Although no apparent reproductive or morphological aberrations were observed in regenerated maize with excised WUS/BBM, there is need for downstream phenotypic and genetic analysis of progeny for subtle anomalies. Could transient morphogene expression have caused epigenetic reprogramming, stress induced mutations, or hidden somaclonal variation? Besides, current excision mechanisms do not guarantee complete removal of MRs and the efficacy differs across genotypes and species (Hoerster et al., 2020).
Licensing and Intellectual property (IP) constraints: The usage of key morphogenic regulators such as BBM/WUS, GRF/GIF and excision constructs are patented by major agbiotech companies or institutions (Gordon-Kamm et al., 2014; Jorge et al., 2021; Stern and Hynes, 2015). Although these technologies are free for use by academic institutions under research use exemptions, entities using them to develop seed systems or for commercialization need to obtain appropriate licensing and freedom to operate agreements. Notably, such systems could constraint accessibility for researchers in low resource settings and breeding programs in the public sector (Van Overwalle et al., 2006).
Species and genotype dependency of response to MRs: Plant response to MRs varies across species and genotypes, and some species require tailored MRs combinations (Wang et al., 2025). Besides, morphogenic regulators alone do not solve issues like recalcitrance to Agrobacterium infection or extreme sensitivity of tissues to wounding, hence parallel fine tuning of other parameters like Agrobacterium strain, explant type, media composition and culture conditions are needed for each case (Duan et al., 2022; Wang et al., 2023).
Regulatory hurdles and public acceptance issues: Heterologous usage of MR genes, especially in vegetatively propagated crops where transgene segregation is challenging, could add an additional layer of regulatory scrutiny during commercial deployment. Such concerns could also further complicate public acceptance of such products.
Recent advances and future prospects
7
The field of plant transformation is currently in a dynamic period thanks to the advent of morphogenic genes and breakthroughs in developmental biology. Among these is the recent discovery of the peptide regenerating factor REF1, as it provides a completely novel mode for enhancing plant regeneration. Unlike conventional MRs that are typically incorporated in a gene construct then transformed into the plant, REF1 is introduced into cells by externally treating plant tissue with the peptide signal (Yang et al., 2024). Notably, REF1’s broad effectiveness across species suggests it could become a standard supplement in transformation protocols to boost shoot formation. Besides, its discovery suggests there is potential to identify or engineer other peptides with similar effects (Youngstrom et al., 2025).
In a recent study, Zhang et al. (2024c) used an inducible CRISPR activation libraries and multiplexed guide RNAs to upregulate gene combinations in Arabidopsis. The study subsequently identified pairs with synergistic effects on regeneration. This high-throughput approach offers unprecedented ways to systematically pinpoint which gene sets promote best regeneration in recalcitrant species. As such, we can now apply CRISPR activation to map out regeneration gene networks for different crops.
The possibility of using MR for in planta Transformation will offer the long-awaited transition from tissue culture-based plant engineering systems. Tissue culture-free transformation systems have great potential in addressing genotype-dependency challenges, shortening transformation timeline, and improving operational efficiency by significantly reducing personnel and supply costs (Zhong et al., 2025). Recent advances inch toward that: for example (Maher et al., 2020), demonstrated the induction of de novo meristems directly from somatic tissues, bypassing the need for traditional callus-based regeneration. In Arabidopsis, co-delivery of WUS/STM enabled shoot formation from leaf and stem tissues, significantly accelerating transformation workflows (Maher et al., 2020). Similar strategies have been extended to woody species like citrus, where transient expression of AtWUS enabled editing of the PDS gene via the in planta IPGEC approach (Kelly et al., 2025). In another study, soil-grown tomato plants and snapdragon (Antirrhinum majus) were successfully transformed by inducing meristematic outgrowths on wound sites using MRs (Lian et al., 2022).
Integration with speed breeding and haploidy: Morphogenic regulators could accelerate doubled haploid production – an area critical for crop breeding. For instance, maize haploid inducer lines could potentially express a morphogenic factor to cause haploid embryo-like structures to germinate more readily or undergo genome doubling in tissue culture. Similarly, in species where making haploids is tough, adding BBM might coax unfertilized ovules to develop (some BBM variants cause parthenogenesis in the absence of fertilization, which is being looked at for synthetic apomixis). So there’s interplay with creating homozygous lines faster.
Regulations surrounding genome editing
8
CRISPR–Cas technology faces challenges such as reagent delivery, precision editing with minimal off-target effects, and the production of transgene-free plants. The integration of CRISPR components like Cas, guide RNA, and selectable markers into the host genome raises regulatory issues. However, advancements in gene-editing, DNA-independent RNP delivery, grafting-mediated editing, nanoparticles, and viral delivery are facilitating transgene-free crop improvement and aligning with biosafety regulations. Further, the transgenic-free delivery method and regeneration protocols enhance efficiency and minimize cytotoxicity, paving the way for broader applications of this DNA-free technology. Regulatory frameworks for genome-edited crops differ worldwide, affecting commercialization, as some countries accept these technologies while others are still debating their legal status (Ramakrishnan et al., 2025). In most countries, genome editing products are regulated according to product- and process-based guidelines (Fernández Ríos et al., 2025). Guidelines based on processes emphasize recombinant DNA technology, while product-based rules evaluate organisms based on their attributes, regardless of the method of development. DNA delivery methods such as Agrobacterium-mediated transformation and particle bombardment can lead to random transgene integration in plant genomes, potentially disrupting essential genes and causing variability in transgene expression. Consequently, such genetically modified organisms (GMOs) are subject to regulatory scrutiny. To mitigate these issues, DNA-free genome editing technologies have been developed, particularly through the use of pre-assembled ribonucleoproteins (RNPs) comprising Cas proteins and synthesized guide RNAs. This method ensures that no recombinant DNA is involved, rendering the edited plants transgene-free. RNPs demonstrate advantages in precision and transient activity within plant cells, allowing targeted genomic editing while minimizing off-target effects and reducing risks of mosaicism and toxicity, as they are degraded shortly after delivery (Ramakrishnan et al., 2025; Maia et al., 2024). The regeneration of whole plants from edited protoplasts remains a significant challenge for many species employing RNA-mediated genome editing. To address this, alternative strategies such as morphogenic induction have been proposed. These approaches involve the co-expression of morphoregulator alongside gene editing cassettes, enabling the formation of gene-edited shoots and ensuring the heritable transmission of modified DNA to subsequent generations. Current research emphasizes enhancing the delivery, selection, and regeneration of CRISPR reagents across diverse plant species, thereby broadening the application of CRISPR technology in crop improvement and agricultural innovation. The existing regulatory framework for the commercialization of genome editing products is regulated by both product- and process-oriented criteria. The Agrobacterium-mediated introduction of CRISPR vector reagents into plant cells and tissues faces regulatory obstacles due to random integration, and the subsequent segregation to remove transgenes is a laborious operation, rendering it unfeasible for trees and vegetative plants. Furthermore, the persistent exposure of genomic DNA to CRISPR agents throughout the editing procedure may lead to off-target mutations and the emergence of chimera mutants (Hashimoto et al., 2016). Additionally, polyploid organisms frequently necessitate numerous gRNAs, hence complicating the transformation of CRISPR vector systems. In light of the aforementioned facts, plant virus-based replicons and ribonucleoprotein-mediated gene delivery, along with integration of the morphogenic regulator-based improved transformation system, are increasingly utilized as alternatives to Agrobacterium for the introduction of genome editing tools into plants, aiming to enhance crop improvement while minimizing the time and labor associated with the current regulatory framework.
Conclusion and future directions
9
Morphogenic regulators and peptide factors are reshaping the landscape of plant transformation and gene editing. What was once an art of coaxing recalcitrant tissues to regenerate is becoming a predictable, engineerable process. By tapping into the plant’s own developmental control circuits through genes like WUS, BBM, LEC, GRF-GIF, and signals like REF1 peptide, researchers can dramatically improve the efficiency and speed of obtaining transgenic or edited plants across a wide range of species. The successes in major crops like maize and wheat have broken the dogma that some elite genotypes are untransformable. Furthermore, the convergence of morphogenic regulators with cutting-edge genome editing and synthetic biology heralds a new era for plant science and agriculture. The ultimate vision is to establish a robust and streamlined plant transformation system that will allow us to move from a genetic concept to edited plant in a season for any crop of interest. Nonetheless, realization of the full potential of these advances will require continued innovation. For instance, more refined control systems will be needed to ensure MRs do their job and then exit fully, without trace. Inducible and tissue-specific systems are a great start, and the trend will be to make these ever more precise. Considering the rapid progress to date, however, this is a highly attainable goal. The integration of RNA-seq, ATAC-seq, and CUT&Tag techniques enables researchers to elucidate gene regulation and cellular function, facilitating the identification of candidate MRs for understanding transcriptional regulatory networks that govern embryonic callus formation and organogenesis (Ouyang et al., 2021; Liu et al., 2023b). These approaches uncover novel transcription factors and peptides implicated in regeneration, along with transcriptome and epigenetic alterations during callus maturation and shoot regeneration. The integration of artificial intelligence and machine learning, combined with the functional validation of newly identified morphogenic factors, enhances the understanding of somatic embryogenesis, de novo regeneration, and addresses challenges associated with plant regeneration, thereby significantly improving the transformation efficiency of numerous economically important crops. Many technology stakeholders are set to benefit from these advances, including scientists being able to accelerate functional genomics research and breeders tailoring crops to the needs of consumers and changing climates. The society is also set to benefit as we adapt our food system to meet global challenges. By expediting plant transformation and gene editing, morphogenic regulators are not just tools of convenience, but also catalysts helping us to engineer crops for a sustainable future.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aesaert S. Impens L. Coussens G. Van Lerberge E. Vanderhaeghen R. Desmet L. . (2022). Optimized transformation and gene editing of the B 104 public maize inbred by improved tissue culture and use of morphogenic regulators. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.883847, PMID: 35528934 PMC 9072829 · doi ↗ · pubmed ↗
- 2Ainley W. M. Sastry-Dent L. Welter M. E. Murray M. G. Zeitler B. Amora R. . (2013 a). Trait stacking via targeted genome editing. Plant Biotechnol. J. 11, 1126–1134. doi: 10.1111/pbi.12107, PMID: 23953646 · doi ↗ · pubmed ↗
- 3Alemanno L. Devic M. Niemenak N. Sanier C. Guilleminot J. Rio M. . (2008). Characterization of leafy cotyledon 1-like during embryogenesis in Theobroma cacao L. Planta 227, 853–866. doi: 10.1007/s 00425-007-0662-4, PMID: 18094994 · doi ↗ · pubmed ↗
- 4Aliu E. Ji Q. Wlazlo A. Grosic S. Azanu M. K. Wang K. . (2024). Enhancing Agrobacterium-mediated plant transformation efficiency through improved ternary vector systems and auxotrophic strains. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1429353, PMID: 39109064 PMC 11300283 · doi ↗ · pubmed ↗
- 5Anand A. Bass S. H. Wu E. Wang N. Mc Bride K. E. Annaluru N. . (2018). An improved ternary vector system for Agrobacterium-mediated rapid maize transformation. Plant Mol. Biol. 97, 187–200. doi: 10.1007/s 11103-018-0732-y, PMID: 29687284 PMC 5945794 · doi ↗ · pubmed ↗
- 6Andersson M. Turesson H. Olsson N. Fält A. Ohlsson P. Gonzalez M. N. . (2018). Genome editing in potato via CRISPR-Cas 9 ribonucleoprotein delivery. Physiol. Plantar. 164, 378–384. doi: 10.1111/ppl.12731, PMID: 29572864 · doi ↗ · pubmed ↗
- 7Anzalone A. V. Randolph P. B. Davis J. R. Sousa A. A. Koblan L. W. Levy J. M. . (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 576, 149–157. doi: 10.1038/s 41586-019-1711-4, PMID: 31634902 PMC 6907074 · doi ↗ · pubmed ↗
- 8Aregawi K. Shen J. Pierroz G. Sharma M. K. Dahlberg J. Owiti J. . (2022). Morphogene-assisted transformation of Sorghum bicolor allows more efficient genome editing. Plant Biotechnol. J. 20, 748–760. doi: 10.1111/pbi.13754, PMID: 34837319 PMC 8989502 · doi ↗ · pubmed ↗