Promotion of plant growth by endophyte Bacillus amyloliquefaciens N3 through modulation of auxin translocation under nitrate-limited conditions
Huan Luo, Yong Hu, Hong Shen, Xiaohui Zhang, Nannan Li

TL;DR
A soil bacterium helps plants grow better when nitrate is low by influencing how a plant hormone is transported.
Contribution
The study reveals a novel mechanism by which an endophytic bacterium promotes plant growth under low-nitrate conditions via auxin translocation modulation.
Findings
Bacillus amyloliquefaciens N3 promotes lateral root formation and plant growth under low nitrate.
The bacterium increases auxin accumulation via upregulated PIN gene expression.
Auxin transport and NRT1.1 are essential for the observed growth promotion.
Abstract
Numerous rhizobacteria have been isolated and could be utilized in an environmentally friendly way to improve crop yield by promoting plant growth. However, the underling mechanism is not well understood. In this study, we demonstrated that Bacillus amyloliquefaciens N3 isolated from purple soil promoted the lateral root formation and plant growth under nitrate-limited conditions. B. amyloliquefaciens N3 increased the local auxin accumulation in the lateral root tip, which may be attributed to the upregulated expression of PIN-FORMED (PIN) genes. Auxin polar transportation inhibition by NPA neutralized the promotion effects on lateral root density. Genetic analysis with mutants auxin response regulator 7 (arf7), arf9, and nitrated transporter 1.1 (nrt1.1) revealed that the promotion of plant growth by B. amyloliquefaciens N3 depends on both auxin-responsive ARF-mediated transcriptional…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
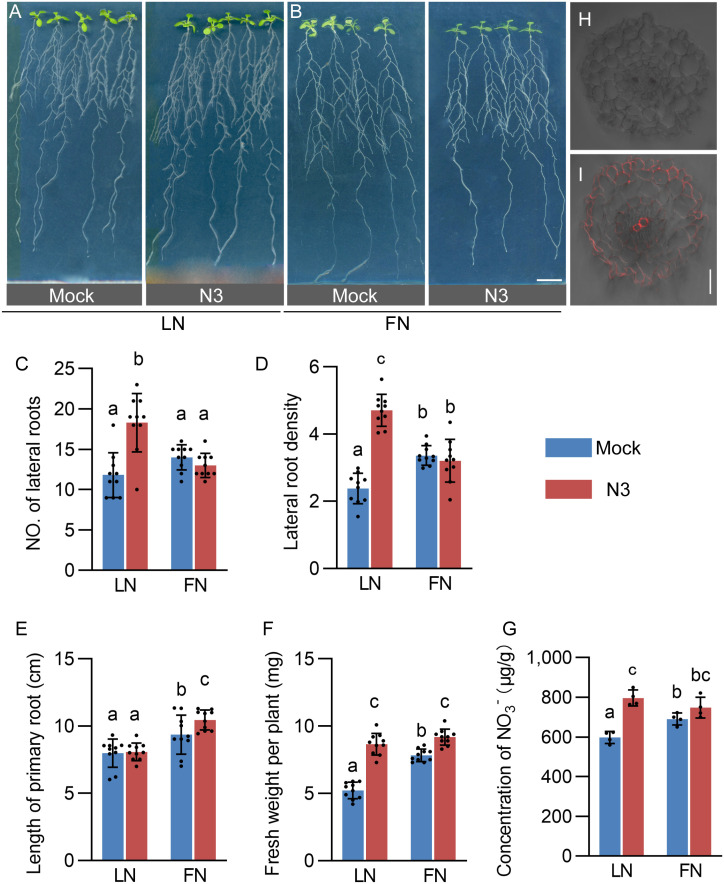 Figure 1
Figure 1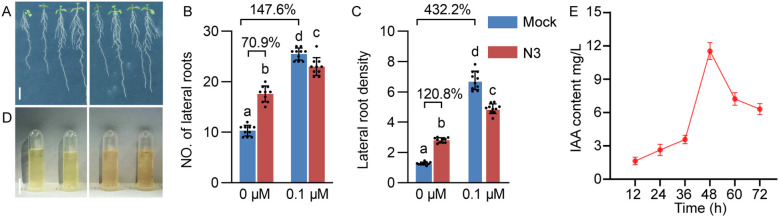 Figure 2
Figure 2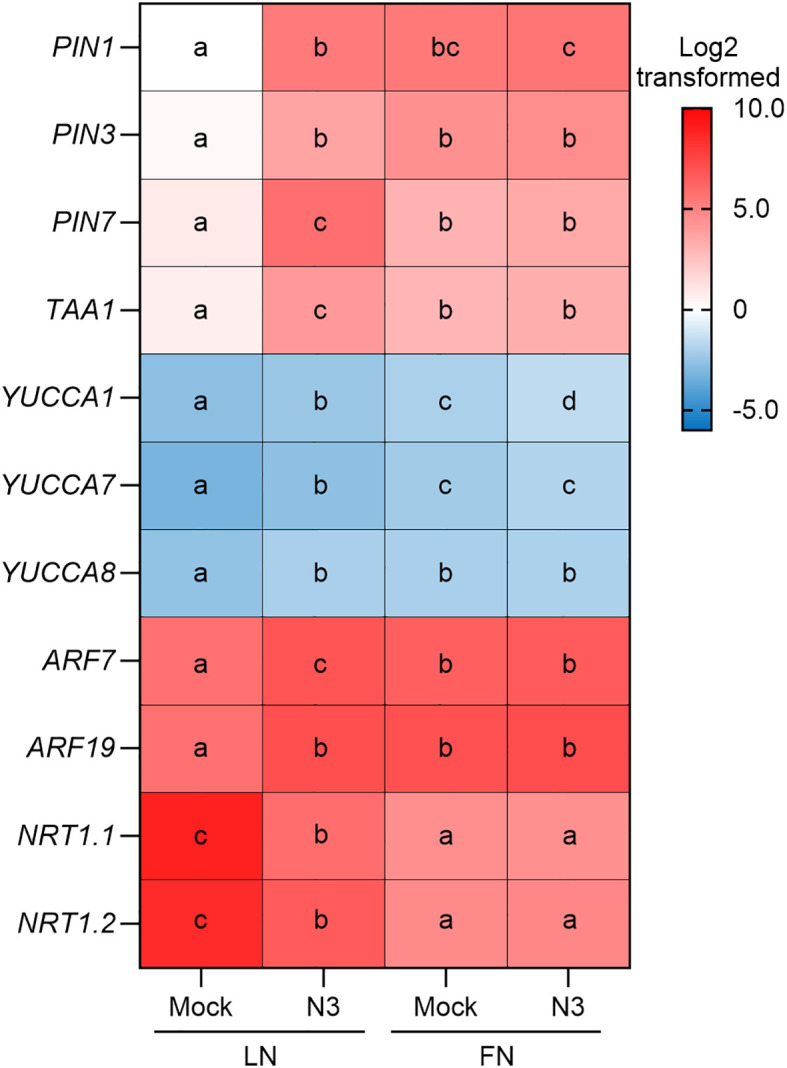 Figure 3
Figure 3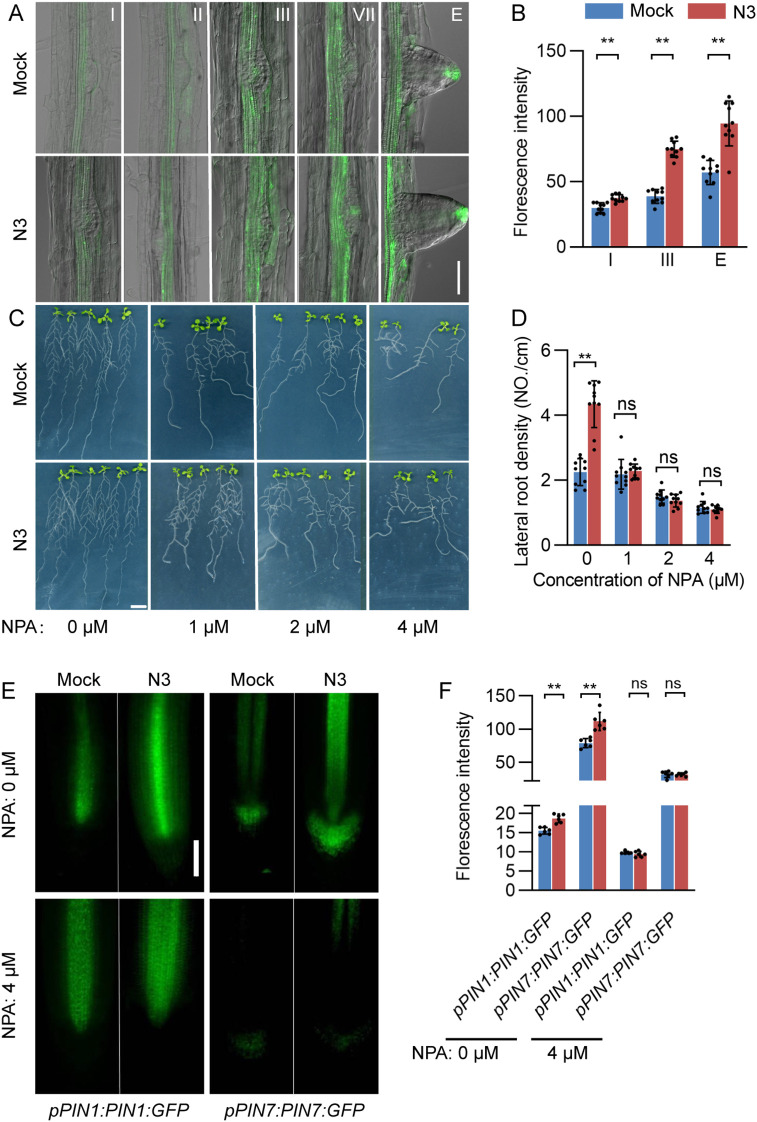 Figure 4
Figure 4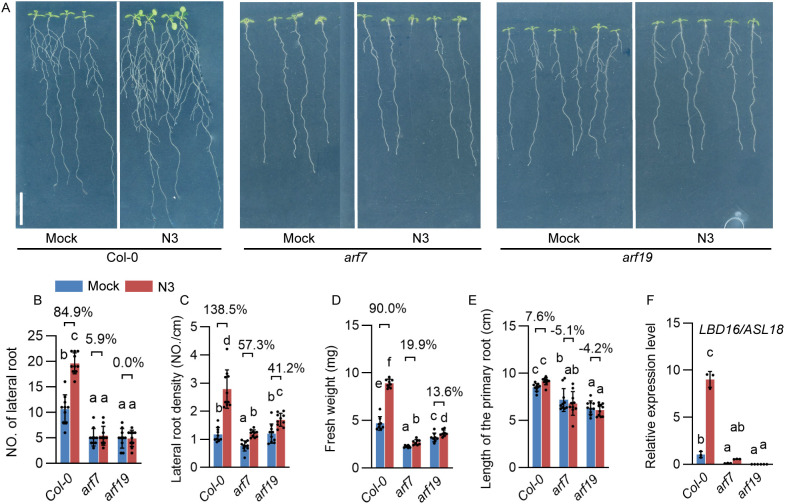 Figure 5
Figure 5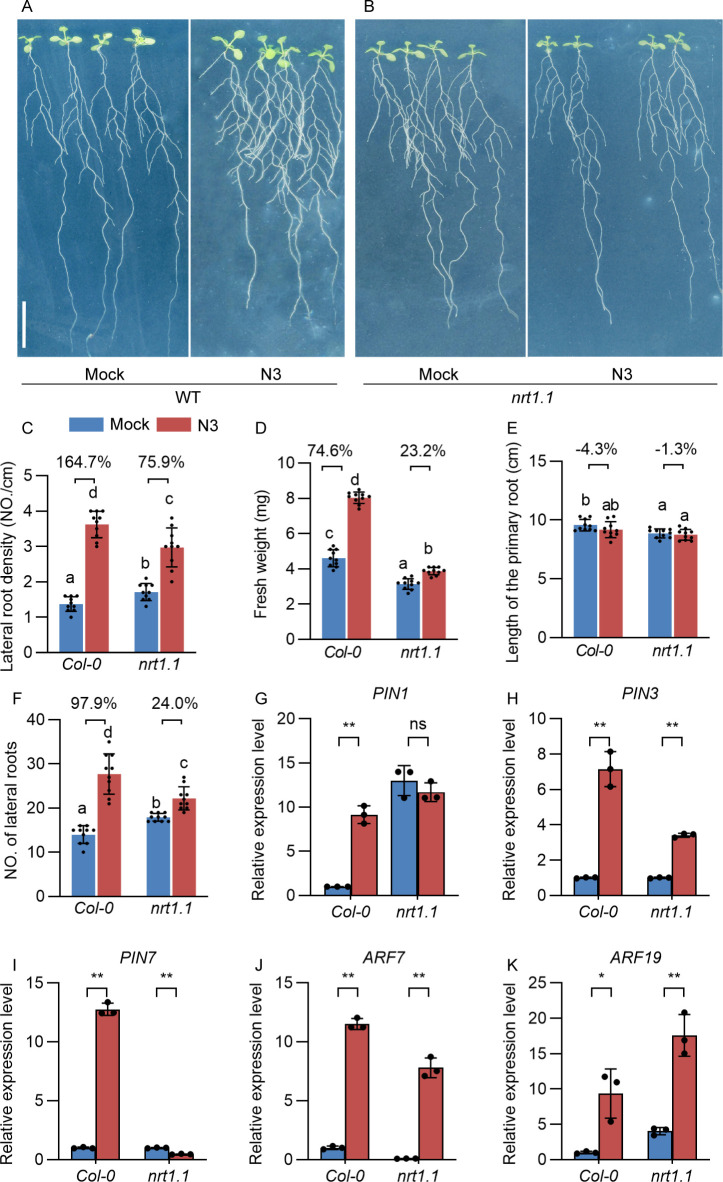 Figure 6
Figure 6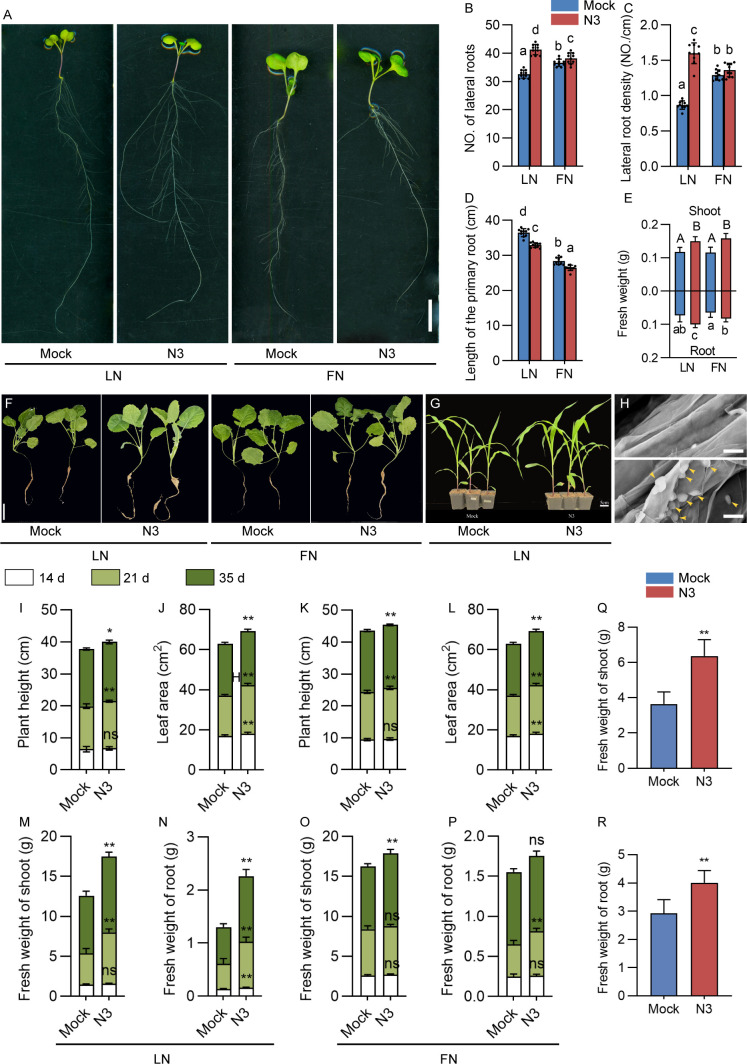 Figure 7
Figure 7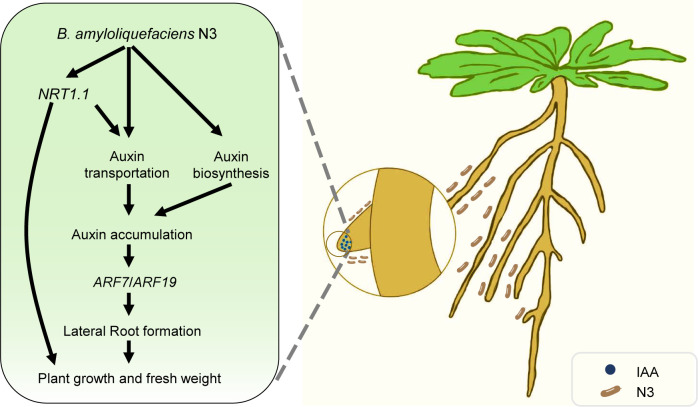 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Molecular Biology Research · Legume Nitrogen Fixing Symbiosis
Introduction
1
Plants have a high degree of plasticity in shaping their root architecture to optimize nutrient absorption. Lateral roots are important to forage for nutrients due to their ability to increase the uptake area of a root system (Pélissier et al., 2021). The NITRATE TRANSPORTER1.1 (NRT1.1) that functions in nitrate sensing and auxin transport plays a complex role in blocking lateral root primordia development (Pélissier et al., 2021). Under low nitrate conditions, NRT1.1 acts as an auxin transporter and facilitates basipetal transport of auxin away from the lateral root tip. This prevents the local auxin accumulation necessary for growth, thereby inhibiting lateral root elongation (Krouk et al., 2010). Conversely, under high nitrate conditions, NRT1.1 protein is downregulated in lateral root primordia, allowing auxin to accumulate (Bouguyon et al., 2016). Despite the regulation by NRT1.1, auxin accumulation in lateral root primordia is also affected by PIN1-, PIN3-, and PIN7-mediated auxin transportation (Lewis et al., 2011; Marhavý et al., 2014; Wang et al., 2019). This localized auxin buildup accelerates the degradation of AUXIN/INDOLE-3-ACETIC ACID proteins, thereby derepressing AUXIN RESPONSE FACTOR (ARF) proteins, such as ARF7 and ARF19 function as transcriptional activators. Once derepressed, ARF7 and ARF19 initiate the transcriptional programs required for lateral root primordium development and subsequent emergence (Okushima et al., 2007; Zhang et al., 2023).
Bacteria that colonize the roots of plants are essential contributors of plant growth. The genus Bacillus is one of the predominant bacterial genera found in soil and have been identified as plant growth-promoting rhizobacteria that improve plant performance through various mechanisms (Saxena et al., 2020). The Bacillus subtilis strain GB03 promotes cell expansion in Arabidopsis through volatile organic compound-dependent regulation of auxin homeostasis (Zhang et al., 2007). Another strain Bacillus methylotrophicus M4–96 isolated from the maize (Zea mays) rhizosphere increases growth and auxin content in Arabidopsis thaliana (Pérez-Flores et al., 2017). The most recent study revealed that the volatile compounds of Bacillus amyloliquefaciens SQR9 induced lateral root formation by regulating auxin biosynthesis (Li et al., 2021). In addition, Bacillus spp. can also help plants to tolerate abiotic stress caused by environmental factors such as drought, salinity, heat, and cold (Tsotetsi et al., 2022). The mechanisms associated with enhancing stress tolerance involve triggering biological and physiological processes such as ROS detoxification mechanisms, osmoprotection, stomatal regulation, membrane stability, xylem hydraulic conductance, root zone water, and nutrient availability and metal chelation (Mahapatra et al., 2022). While these studies shed light on the mechanisms of Bacillus-mediated plant growth promotion, a crucial gap remains: evaluating its effectiveness under conditions of nutrient stress, especially the nitrate-limited conditions. In this study, we explored the effects of strain Bacillus amyloliquefaciens N3 on lateral root development by utilizing molecular genetics approaches and validated its effects in different crop plants in soil conditions. Our finding expanded the understanding of beneficial endophytic bacterial application potential in nitrate-limited environments.
Materials and methods
2
Plant material and growth conditions
2.1
The Arabidopsis thaliana materials included the wild-type accession Col-0, the mutants nrt1.1 (SALK_097431), arf7 (SALK_027626C), and arf19 (SALK_009879C), and the transgenic lines pDR5:GFP, pPIN1:PIN1:GFP, and pPIN7:PIN7:GFP, which were kindly provided by Prof. Ruifu Zhang (Nanjing Agricultural University). Brassica napus cultivar Zhongshuang 11 (ZS11) and Zea mays inbred line B73 were also used.
After 2 days of vernalization at 4°C in the dark, Arabidopsis thaliana seeds were immediately subjected to surface sterilization. Seeds were treated with 75% (v/v) ethanol for 1 min, followed by immersion in 10% (v/v) NaClO solution for 7 min, and then rinsed thoroughly five times with sterile distilled water. For Brassica napus ZS11 and Zea mays inbred line B73, seeds were sterilized using 75% (v/v) ethanol for 3 min, followed by 10% (v/v) NaClO solution for 5 min, and subsequently rinsed five times with sterile distilled water. The Arabidopsis and ZS11 seeds were grown in agar plates containing MS Base Salts (without nitrate) with addition of 1.9 or 0.19 g/L KNO_3_ in square petri plates (12×12 cm or 25×25 cm). Plates were then placed in the dark for 2 days to allow germination. Seeds with a similar germination state were chosen for inoculation experiments in plates. The 7-day-old ZS11 seedlings and germinated B73 seeds were used in the inoculation experiments in soil conditions.
For inoculation assays in plates, B. amyloliquefaciens N3 was cultured in Luria–Bertani (LB) liquid medium at 28°C with shaking at 180 rpm for approximately 16 h and then harvested by centrifugation at 5,000 × g for 10 min at room temperature, washed twice with sterile water, and resuspended in sterilized water to an optical density of OD_600_ = 0.1 ± 0.02. The inoculation medium was prepared by mixing the bacterial suspension with the indicated medium at a ratio of 1:10 (v/v). For mock treatment, sterile water prepared using the same procedure but without B. amyloliquefaciens N3 was mixed with the indicated medium at the same ratio. All subsequent growth conditions were identical between mock-treated and inoculated plants. Sterilized seeds with a comparable germination state were transferred to the inoculation medium and grown in a growth chamber at 22°C under a 16-h light/8-h dark photoperiod with a light intensity of approximately 200 μmol m^−2^ s^−1^.
For the inoculation experiments in soil conditions, plants were grown in the same batch of potting soil (total nitrogen, 1.36 g/kg^−1^; alkali-hydrolyzable nitrogen, 56 mg/kg^−1^, pH value, 6.42). Soil temperature was maintained according to the chamber settings, and soil texture was identical across all treatments. The 7-day-old ZS11 seedlings and germinated B73 seeds were transplanted into soil. Plants were grown in a growth chamber under the same conditions as described above. The bacterial inoculum was prepared as described above and resuspended with sterilized water to an optical density of OD_600_ = 0.1 ± 0.02. A 15-mL bacterial inoculum was inoculated into the root zone every week. Accordingly, the mock treatment was inoculated with 15 mL of sterilized water.
Phenotype evaluation and data analysis
2.2
For the phenotypes of plate inoculation experiments, seedlings inoculated for 12 days were scanned with an Epson Perfection V850 Pro scanner and further analyzed with ImageJ software for analysis of root phenotype. The total biomass of each seedling was measured on an analytical balance. For the phenotypes of soil inoculation experiments, at 14, 21, and 35 days after first inoculated treatment, seedlings were harvested for fresh weight evaluation.
Analysis of contents of indole-3-acetic acid and nitrate
2.3
Salkowski’s test was used to detect the presence of indole-3-acetic acid (IAA) or its precursors in the supernatant of bacterial culture as described before (Rahman et al., 2010; Gang et al., 2019). A salicylic acid-concentrated sulfuric acid colorimetry method (Qian et al., 2017) was used to determine the nitrate content in mock treatment and B. amyloliquefaciens N3-inoculated Arabidopsis plants.
N-1-naphthylphthalamic acid treatment and fluorescence microscopy
2.4
N-1-Naphthylphthalamic acid (NPA) was dissolved in dimethyl sulfoxide (DMSO) to make a 50-mM solution and supplied at different doses (1, 2, and 4 μM) to the plant growth medium. Arabidopsis plants grown in petri plates were placed in a growth chamber for 12 days to detect the phenotype of roots. For confocal microscopy, mock or B. amyloliquefaciens N3-inoculated DR5:GFP seedlings were mounted in distilled water on microscope slides. A Zeiss 780 laser-scanning microscope was used for fluorescence imaging of the Arabidopsis lateral roots at early stages of development as previously described (Malamy and Benfey, 1997). These stages were classified as the following: stage I, initiation of the lateral root primordium; stage II, periclinal division of the lateral root primordium into two distinct cell layers; stage III, further periclinal division of the outer layer, resulting in a three-layered primordium; stage IV, formation of a four-layered primordium; stages V–VII, expansion of the lateral root primordium through the parental cortex until it is poised to emerge from the primary root surface; and stage E, the emergence of lateral root. Chromophores were excited using a 488-nm argon laser, and fluorescence was detected at 500–550 nm. More than eight independent seedlings were analyzed for each genotype, and treatment representative images were selected for figure construction. In pPIN1:PIN1:GFP and pPIN7:PIN7:GFP transgenic lines, fluorescence signals were captured under identical microscope settings. Quantification was performed using ImageJ software. Measurements were performed on lateral root tip region, and the mean fluorescence intensity of each individual sample was calculated after subtraction of background fluorescence.
RNA extraction and quantitative real-time PCR analysis
2.5
The root materials collected after 12 days of co-cultivation were ground in liquid nitrogen, and the total RNA was extracted with three biological replicates. The cDNA synthesis was generated from total RNA using the PrimeScript™ RT reagent Kit (TaKaRa). Quantitative real-time (RT) PCR (q-PCR) was performed on a StepOnePlus™ Real-Time PCR System (Applied Biosystems) using a GoTaq qPCR Master Mix (Promega) and specific primers (Supplementary Table 1). AtACTIN2 (AT3G18780) was used as the reference gene. The results were obtained from three biological replicates, and –ΔΔCt was taken for every sample as the relative expression levels.
Construction of mCherry-tagged B. amyloliquefaciens N3 strain and detection of its colonization in plants
2.6
The pUT-mini-Tn5-Kna-Tac-mCherry-SacB plasmid was transformed into B. amyloliquefaciens N3 by conjugation using a filter-mating method, which generates an mCherry-tagged strain B. amyloliquefaciens N3-mCherry. The 8-day-old Arabidopsis seedlings subjected to limited nitrate treatment were inoculated with B. amyloliquefaciens N3-mCherry strain in plates as described above. After 2 days, roots were carefully washed with sterile water three to four times and fixed with 1.5% agarose and then cut into 80 pieces with a vibrating microtome to obtain 80-μm sections. These sections were analyzed with Zeiss laser confocal microscope. At least 10 seedlings were treated, and three independent biological repeated experiments were carried out.
To detect B. amyloliquefaciens N3 colonization in the roots of Brassica napus ZS11, the B. amyloliquefaciens N3 strain was inoculated with 7-day-old ZS11 seedlings grown on plates, as described above. After 2 days of inoculation, root samples were collected and examined using scanning electron microscopy (JSM-5610, Phenom, Holland) at an accelerating voltage of 10 kV to observe bacterial colonization.
Results
3
B. amyloliquefaciens N3 promotes lateral root formation and plant growth under nitrate-limited conditions
3.1
In our previous study, B. amyloliquefaciens N3 (hereafter referred to as N3) was isolated from purple soil, a specific type of soil found predominantly in regions like the Sichuan basin in China (Shen et al., 2018). N3 has been found to significantly promote the growth of Ginger (Zingiber officinale Roscoe) (Shen et al., 2018). In this study, we first confirmed the promotion effect on the growth of Arabidopsis thaliana. As expected, N3 increased the lateral root number, lateral root density, and total biomass but did not affect the primary root length per plant (Figures 1A–F). These effects were found more obvious under limited nitrate (LN, 0.19 g/L KNO_3_) than that in full-nitrate (FN, 1.9 g/L KNO_3_) conditions. In order to explore whether N3 can increase the absorption of nitrate by Arabidopsis and promote its growth and root development under LN treatment, the nitrate content in the plants was measured. The results showed that the nitrate content in the N3-inoculated Arabidopsis plants was significantly higher than that of the control group, but only under LN conditions (Figure 1G). The mCherry-tagged N3 line was constructed to trace the exact colonization region in the root. Strong signals were observed in the apoplastic spaces of epidermal and xylem tissues, weak fluorescence signals were also detected in the apoplast of the pericycle layer (Figures 1H, I). These results suggest that N3 is an endophytic bacterium.
Promoted growth of Arabidopsis seedlings by Bacillus amyloliquefaciens N3. (A, B)Arabidopsis Col-0 seedlings co-cultivated with bacterial B. amyloliquefaciens N3 (OD600 = 0.1 ± 0.02) for 12 days in limited nitrate (LN) and full nitrate (FN) conditions. Scale bar, 1 cm. (C-G) Quantification of number of lateral root per plant (C), lateral root density (D), length of the primary root (PR) (E), total biomass per plant (F), and total NO3- concentration of seedling after 12 days of co-cultivation (G). Data are shown as mean ± standard deviation (SD), n = 15; different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test. (H, I) Fluorescence observation of the B. amyloliquefaciens N3 as the control (H) and mCherry-tagged B. amyloliquefaciens N3 (I) in the cross-cut section of Arabidopsis roots.
The increase of lateral root number and density by N3 mimics the effects of exogenous auxin treatment, which motivated us to inspect the role of auxin in this process. We first compared the effects of N3 with the synthetic auxin, 1-naphthaleneacetic acid (NAA), on the growth of Arabidopsis roots. Treatment of N3 and NAA both increased the lateral root number and density, but more pronounced effects were observed in NAA treatment compared with N3 inoculation (Figures 2A–C). However, treatment of 0.1 μM NAA decreased the total biomass of Arabidopsis plants. Moreover, Salkowski’s test revealed that N3 exhibits IAA production potential and produces a maximum of IAA at 48 h post of culturing (Figures 2D–E). Taken together, these results showed that auxin may play an important role in shaping the root architecture by N3.
The effect of NAA treatment and B. amyloliquefaciens N3 inoculation on the root development of Arabidopsis. (A) The water-treated control (0 μM NAA, left) and 0.1 μM NAA (right) Arabidopsis plants. (B, C) Number of lateral roots (B) and lateral root density (C) of control and NAA-treated plants with or without inoculation of B. amyloliquefaciens N3; n ≥ 9. Percentages in the bar charts show the relative changes between the indicated bars; different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test. (D) Supernatant of control (left) and B. amyloliquefaciens N3 culture (right) treated with Salkowski’s reagent. (E) Quantitative analysis of IAA content in supernatant of B. amyloliquefaciens N3 culture; data are shown as mean ± SD, n=3. Scale bar, 1 cm in A and D.
Upregulation of auxin-related genes after inoculation of N3
3.2
To further dissect the involvement of auxin in the regulation of lateral root formation, we compared the expression levels of several auxin-related genes between N3-inoculated plants and control. Interestingly, under LN conditions, pronounced elevations of expression levels of PIN1, PIN3, and PIN7 involving auxin transportation were detected when plants were inoculated with N3 (Figure 3). A similar pattern of expression changes was observed in the flavin monooxygenase genes, YUCCA1, YUCCA7, YUCCA8, and TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 (TAA1) which regulate auxin biosynthesis, as well as in the two Auxin Response Factor (ARF) genes mediating the auxin-responsive regulation (Figure 3). These results suggest that N3 treatment transcriptionally altered the auxin biosynthesis, transport, and auxin-responsive ARF signaling in Arabidopsis plant.
Expression analysis of gene involved in auxin-related genes and nitrate transporter genes. Expression levels of indicated genes in mock and B. amyloliquefaciens N3-inoculated 12-day-old plants under both LN and FN conditions. Data are shown as the Log2-transformed mean values of three replicates. Different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test.
N3-mediated lateral root development depends on auxin transport and ARF function
3.3
The markedly increased expression of PIN genes inspires us to explore their roles in shaping lateral root architecture by N3. Local auxin distribution in the lateral root or its primordium was detected by using the pDR5:GFP lines. Significant enhancement of auxin signal was observed in the xylem and lateral root tip of N3-treated root compared to that of the mock plants at stages III to E (Figures 4A, B). A series of concentrations from 1 to 4 μM of NPA, a polar auxin transport inhibitor, were applied to block the auxin polar transportation in the N3-inoculated plants. In the NPA-free treatment, N3 treatment significantly increased the lateral root density (Figures 4C, D). The lateral root density decreased gradually with increases in NPA concentration. Most importantly, the NPA treatment totally abolished the promotion effects of N3 on the lateral root density regardless of the NPA concentrations. To further validate this, GFP-tagged PIN1 and PIN7 transgenic lines (pPIN1:PIN1:GFP and pPIN7:PIN7:GFP) were used to analyze the auxin transport protein accumulation upon NPA treatment (Figures 4E, F). As expected, PIN1 and PIN7 protein accumulation was enhanced by N3 inoculation. However, these effects were compromised when treated with 4 μM of NPA (Figures 4E, F). Thus, the promotion of lateral root development by N3 inoculation depends on the auxin polar transportation pathway.
*Auxin accumulation in the lateral root tip and its effects on lateral root development. (A) Expression patterns of pDR5:GFP during primordium development after 7 days of co-cultivation with B. amyloliquefaciens N3 detected with confocal laser scanning microscope; Scale bar, 50 μm. (B) Quantification of pDR5::GFP signal in the lateral root tips; n ≥ 9; **, p < 0.01, t-test. (C) Phenotype of NPA-treated plants with mock and B. amyloliquefaciens N3 treatment. (D) Lateral root density of NPA-treated plants with mock and B. amyloliquefaciens N3 treatment. Data are shown as mean ± SD, n ≥ 9, **, p < 0.01, ns, not significant, t-test. (E, F) GFP protein accumulation analysis of pPIN1:PIN1:GFP and pPIN7:PIN7:GFP signal (E) and quantification (F); bar, 2 cm. Data are shown as mean ± SD, n = 6, *p < 0.01, ns, not significant, t-test.
Despite the upregulation of PIN genes, the expression levels of ARF7 and ARF19 were enhanced in the N3-inoculated plants. To analyze whether they mediated the interaction between N3 and plants, the phenotypes were compared between inoculated Col-0 and arf mutants under LN and FN conditions (Figure 5, S1). In the Col-0 plants, inoculation of N3 increased lateral root number by 84.9% compared to the control under LN conditions. However, this effect is totally neutralized in arf7 or arf9 backgrounds (Figures 5A, B). For the phenotype of lateral root density, lower percentages of increasement were observed in arf7 (57.3%) and arf19 (41.2%) than that in Col-0 (138.5%) when inoculated with N3 (Figure 5C). A similar pattern was observed for fresh weight: N3 increased fresh weight by 90.0% in the Col-0, but only by 19.9% and 13.6% in arf7 and arf19, respectively (Figure 5D). Moreover, we further performed expression analysis of LOB DOMAIN-CONTAINING PROTEIN 16 (LBD16)/ASYMMETRIC LEAVES2-LIKE 18 (ASL18) to validate the involvement of the ARF/LDB signaling pathway. Enhanced expression of LDB16/ASL18 was detected in N3-inoculated Col-0 plants. These effects were weakened in the arf7 and abolished in arf19 (Figure 5F). Taken together, our results demonstrated the promotion effect of N3 on the lateral root architecture depending on the auxin transport and auxin-responsive ARF signaling.
Effects of B. amyloliquefaciens N3 on growth and root development in auxin response factor (arf) mutants. (A) Phenotype of roots of Col-0, arf7, and arf19 mutants with 12 days of mock or B. amyloliquefaciens N3 inoculation treatments, scale bar, 1 cm. (B-E) Number of lateral roots (B), lateral root density (C), fresh weight (D), and length of primary root (E) after inoculation indicated in (A); data are shown as mean ± SD, n ≥ 8. Percentages in the bar charts shows the relative changes between the indicated bars; different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test. (F) Expression analysis of LOB DOMAIN-CONTAINING PROTEIN 16 (LBD16)/ASYMMETRIC LEAVES2-LIKE 18 (ASL18) in Col-0, arf7, and arf19 mutants. Data are shown as the mean ± SD of three replicates. Different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test.
NRT1.1 is crucial for enhancement of biomass by N3
3.4
The transcript level analysis also showed that several nitrate transporter genes were downregulated by N3 treatment under LN conditions (Figure 3). Moreover, NRT1.1 was a key gene that is responsible for the root development at LN conditions (Krouk et al., 2010; Pélissier et al., 2021). To analyze its role in the N3-mediated plant growth promotion, we used the nrt1.1 mutant in the inoculation experiment. Interestingly, the promotion of lateral root density was less effective in the nrt1.1 mutant (75.9%) compared to that of Col-0 (164.7%). Moreover, less effective promotion effects on fresh weight were observed in nrt1.1 (23.2%) mutant compared with that in Col-0 (74.6%) (Figures 6A–F, Supplementary Figure 2). These results indicated the significant but partial role of NRT1.1 in the regulation of fresh weight during N3 inoculation. Because our results mentioned above demonstrated that N3-mediated regulation of lateral root formation totally relied on the auxin transport and auxin-responsive ARF-mediated pathways, we speculated that NRT1.1 may act upstream of the auxin-related genes. To validate this hypothesis, we compared the expression of PIN and ARF genes between the inoculated and control groups in different backgrounds. As expected, the expression differences of PIN1, PIN3, and PIN7 between N3-inoculated and mock plants were relatively smaller in nrt1.1 than that in Col-0 (Figures 6G–K). More specifically, expression levels of PIN1 in nrt1.1 showed no difference between mock and inoculated plants, which were distinct from the significant differences found in Col-0 plants. Therefore, the influence on auxin transportation by N3 is partially dependent on NRT1.1.
*Effects of B. amyloliquefaciens N3 on growth and root development in nitrate transporter 1.1 (nrt1.1) mutant. (A, B) Phenotype of roots of Col-0 and nrt1.1 mutant with 12 days of mock or B. amyloliquefaciens N3 inoculation treatments. (C-F) Lateral root density (C), fresh weight (D), length of the primary root (E) and the number of lateral roots (F); data are shown as mean ± SD, n ≥ 9. Percentages in the bar charts shows the relative changes between the indicated bars; different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test, n ≥ 9. (G-K) Expression levels in Col-0 and nrt1.1 mutant under mock or B. amyloliquefaciens N3 inoculation treatments. Data are shown as the mean ± SD of three replicates. *p < 0.05; *p < 0.01, t-test.
N3 effectively promotes the growth of rapeseed and corn
3.5
To evaluate the promotion effect of N3 on the crop plants, inoculation experiments were performed on dicot crop rapeseed and monocot crop corn (Figures 7A–R). In the plate experiments, N3 treatment significantly increased the lateral root number and density under LN conditions (Figures 7A–E). Significant increases of fresh weight of root and shoot were detected in N3-inoculated plants under both LN and FN conditions compared to the mock. The effects of N3 in the soil cultivation conditions were also evaluated (Figures 7F, G). Colonization of N3 in root of rapeseed was also validated with scanning electron microscopy (Figure 7H). For rapeseed, increases in plant height, leaf area, and fresh weight of shoot and root were detected after different days of inoculation (Figures 7I–P). These promotion effects were more obvious under LN compared with FN conditions, especially for the fresh weight traits. For the monocot corn, both shoot and root fresh weights were significantly increased by inoculation of N3 (Figures 7G, Q, R). These results suggested the N3 bacteria show consistent and stable promotion effects on different crop plants under soil conditions.
*The promotion effect of B. amyloliquefaciens N3 on crop plants. (A) Promotion effect of B. amyloliquefaciens N3 on ZS11 under LN or FN conditions in the plate experiment. (B-E) Number of lateral root (B), lateral root density (C), length of primary root (D), and fresh weight (E); data are shown as mean ± SD, n ≥ 9. Different letters indicate the significant difference at p < 0.05, one-way ANOVA, Duncan’s test, n ≥ 9. (F) Seedling phenotypes of soil-grown ZS11 under LN or FN conditions with mock or B. amyloliquefaciens N3 inoculation; bar, 5 cm. (G) Phenotype of soil-grown Zay maize with mock or B. amyloliquefaciens N3 treatments under LN conditions; bar, 5 cm. (H) Colonization analysis of B. amyloliquefaciens N3 in root of Brassica napus ZS11 by using scanning electron microscope. Bar, 5 μm. Yellow arrows indicate the B. amyloliquefaciens N3 bacteria. (I-P) Plant height (I, K), leaf area (J, L), and fresh weight of shoot (M, O) and root (N, P) with mock or B. amyloliquefaciens N3 treatments under LN or FN conditions. Data are shown as the mean ± SD, n ≥ 9; *p < 0.05; **p < 0.01, ns, not significant, t-test. (Q, R) Fresh weight of corn shoot (Q) and root (R) with mock or B. amyloliquefaciens N3 treatments under LN conditions. Data are shown as the mean ± SD, n = 10; *, p < 0.01, t-test.
Discussion
4
In the model plant Arabidopsis thaliana, several genera of beneficial bacteria such as Bacillus, Pseudomonas, and Phyllobacteria strains can change the root system through different mechanisms including affecting plant hormone homeostasis or signal transduction (Kudoyarova et al., 2019; Dahmani et al., 2020; Jiménez-Vázquez et al., 2020). However, under the limited-nitrate conditions, the mechanism of promoting lateral root growth by beneficial bacteria is less investigated. In this study, N3 significantly increased the number of lateral roots, lateral root density, and nitrate content in Arabidopsis under nitrate-limited conditions. These promotion effects were very weak under FN conditions (Figure 1, Supplementary Figure 3). Our previous study has shown that N3 has N_2_-fixing activity (Shen et al., 2018). Thus, to understand the mechanisms behind plant promotion, it is important to determine whether it is caused by changes in auxin transport pathways or by enhanced N_2_ fixation. This study showed that the inoculation of N3 suppressed the expression of NRT1.1 (Figure 3) and promoted the lateral root development (Figures 1C, D). This is different from the effects of direct application of external nitrate, which induces the expression of nitrate transporter genes and elicits lateral root elongation (Mantelin and Touraine, 2004). Therefore, changes in auxin transportation by N3 are more likely be the reason for enhancement of the root system which increased nitrate uptake (Figure 1G) and enhanced biomass accumulation (Figure 1F, Figures 7E, O–R).
NRT1.1 was reported as a repressor of lateral root growth at low nitrate availability by promoting basipetal auxin transport (Krouk et al., 2010). In the present study, NRT1.1 expression was transcriptionally suppressed upon N3 inoculation (Figure 3), and mutation of nrt1.1 partially attenuated the N3-induced increase in lateral root density and fresh weight (Figures 6C, D). These results suggest that suppression of NRT1.1 contributes, at least in part, to the promotive effect of N3 on lateral root development (Figure 8). In addition to NRT1.1, we observed enhanced accumulation of PIN1 and PIN7 proteins following N3 treatment (Figures 4E, F), indicating that PIN-mediated auxin transport is also strengthened. Thus, N3 appears to modulate auxin distribution through coordinated regulation of multiple transport components. It is noteworthy that mutation arf7 or arf19 did not completely compromise the promotion effects of N3 (Figures 5C, D), which suggested the existence of additional mechanisms that operate beyond ARF7/ARF19-dependent signaling. Given that NRT1.1 functions as an auxin transporter under limited nitrate conditions (Krouk et al., 2010), altered NRT1.1-dependent auxin transport may also contribute to the auxin accumulation in the lateral root tip. Furthermore, N3 inoculation significantly induced the expression of several auxin biosynthesis genes including YUCCA1, YUCCA7, YUCCA8, and TAA1 (Figure 3). This is consistent with previous findings that constitutive overexpression of YUCCA1 in the yuc1D mutant elevates IAA levels and enhances lateral root formation (Mashiguchi et al., 2011). Therefore, increased auxin biosynthesis triggered by N3 may represent another important mechanism underlying N3-induced lateral root initiation. Notably, the reduced promotive effect observed under exogenous NAA treatment may be explained by the elevated auxin levels provided by NAA, which could partially saturate auxin signaling and thereby attenuate the additional contribution of N3. Taken together, these findings indicate that N3 promotes lateral root development through coordinated modulation of auxin biosynthesis, auxin transport, and auxin-responsive ARF signaling, thereby reshaping local auxin homeostasis to favor lateral root formation (Figure 8).
Model for B. amyloliquefaciens N3 promoting the development of lateral roots and plant growth. Under LN conditions, B. amyloliquefaciens N3 promoted the accumulation of auxin in the lateral root tips through affecting the auxin transportation mediated by PIN-FORMED and NRT1.1 proteins. B. amyloliquefaciens N3 also enhances the auxin biosynthesis by elevation of expression levels of YUCCA genes. The accumulated auxin enhanced the development of lateral roots through the ARF7 and ARF19 genes, which may promote the nutrient availability and plant growth.
Our results showed that the promotion functions of N3 was more pronounced under LN conditions (Figures 1A–G) compared with FN conditions (Supplementary Figure 2-S3). It correlated with the expression pattern that enhanced the expression of auxin-related genes, which was only observed under LN conditions (Figure 3). Considering that the promotion effect of bacteria was largely affected by the root exudation compounds (Jansson et al., 2023), we speculated that limited nitrate conditions may induce a specific compound secretion which may change the metabolism or growth state and enhance the effects of N3 in the root. On the contrary, metabolites seriated by the bacteria also influence the development and growth of host or nearby plants. The 6-pentyl-pyrone secreted by Trichoderma atroviride has been reported as a regulator of Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning (Garnica-Vergara et al., 2016). A similar mechanism may also exist in N3-mediated effects, which need further investigation in the future.
Although mCherry-tagged N3 was predominantly localized in the apoplastic spaces of epidermal and xylem cells, weak fluorescence signals were also detected in the apoplast of the pericycle layer (Figures 1H, I). As lateral roots originate from pericycle cells adjacent to the xylem pole (Pélissier et al., 2021), the presence of N3 in proximity to this cell layer suggests a potential spatial association between bacterial colonization and sites of lateral root initiation. While colonization was not strictly confined to the pericycle, this spatial proximity may facilitate local modulation of developmental signaling.
Under low nitrogen conditions, the growth-promoting effect of N3 exhibited a clear temporal pattern. Although no significant increase in plant height or shoot fresh weight was observed at 14 days after inoculation under LN conditions, leaf area and root fresh weight were already significantly enhanced at this early stage (Figures 7J, L, N, P). In contrast, significant increases in shoot biomass became evident at 21 and 35 days. This sequential response suggests that early stimulation of root growth may improve nutrient acquisition capacity, which subsequently supports aboveground biomass accumulation. Such a time-dependent enhancement is consistent with the notion that root system modulation represents an initial adaptive strategy under nitrogen limitation (Jia and von Wirén, 2020), ultimately leading to improved whole-plant performance. These findings further support the idea that N3-mediated regulation of root development plays a central role in promoting plant growth under LN conditions.
Although volatile compounds of Bacillus amyloliquefaciens SQR9 promote lateral root development (Li et al., 2021), our findings demonstrated that N3 is an endophyte, because it is detected in the apoplast and the vascular bundles (Figures 1H, I). N3 promotes nitrogen fixing and nitrate uptake into roots under LN conditions. Despite the growth promotion effects on Arabidopsis, our results have also revealed the consistent effects on both dicot crop Brassica napus and monocot crop Zea mays (Figure 7). Thus, the endophyte N3 may have great potential in application as microbial fertilizer.
In conclusion, our study demonstrates that N3 promotes lateral root development and enhances plant growth under low nitrogen conditions. Mechanistically, N3-mediated growth promotion depends on auxin transport and ARF function and is accompanied by transcriptional changes in auxin biosynthesis-related genes. Soil-based experiments further confirm that N3 improves plant performance in an agronomically relevant context. Together, these findings provide new insights into the interaction between beneficial rhizobacteria and auxin-regulated root development under nitrogen limitation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bouguyon E. Perrine-Walker F. Pervent M. Rochette J. Cuesta C. Benkova E. . (2016). Nitrate controls root development through posttranscriptional regulation of the NRT 1.1/NPF 6.3 transporter/sensor. Plant Physiol. 172, 1237–1248. doi: 10.1104/pp.16.01047, PMID: 27543115 PMC 5047109 · doi ↗ · pubmed ↗
- 2Dahmani M. A. Desrut A. Moumen B. Verdon J. Mermouri L. Kacem M. . (2020). Unearthing the Plant Growth-Promoting Traits of Bacillus megaterium Rm Bm 31, an Endophytic Bacterium Isolated From Root Nodules of Retama monosperma. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00124, PMID: 32174934 PMC 7055178 · doi ↗ · pubmed ↗
- 3Gang S. Sharma S. Saraf M. Buck M. Schumacher J. (2019). Analysis of indole-3-acetic acid (IAA) production in Klebsiella by LC-MS/MS and the Salkowski method. Bio-protocol 9, e 3230–e 3230. doi: 10.21769/Bio Protoc.3230, PMID: 33655016 PMC 7854044 · doi ↗ · pubmed ↗
- 4Garnica-Vergara A. Barrera-Ortiz S. Muñoz-Parra E. Raya-González J. Méndez-Bravo A. Macías-Rodríguez L. . (2016). The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning. New Phytol. 209, 1496–1512. doi: 10.1111/nph.13725, PMID: 26568541 · doi ↗ · pubmed ↗
- 5Jansson J. K. Mc Clure R. Egbert R. G. (2023). Soil microbiome engineering for sustainability in a changing environment. Nat. Biotechnol. 41, 1716–1728. doi: 10.1038/s 41587-023-01932-3, PMID: 37903921 · doi ↗ · pubmed ↗
- 6Jia Z. von Wirén N. (2020). Signaling pathways underlying nitrogen-dependent changes in root system architecture: from model to crop species. J. Exp. Bot. 71, 4393–4404. doi: 10.1093/jxb/eraa 033, PMID: 31970412 PMC 7382383 · doi ↗ · pubmed ↗
- 7Jiménez-Vázquez K. R. García-Cárdenas E. Barrera-Ortiz S. Ortiz-Castro R. Ruiz-Herrera L. F. Ramos-Acosta B. P. . (2020). The plant beneficial rhizobacterium Achromobacter sp. 5B 1 influences root development through auxin signaling and redistribution. Plant J. 103, 1639–1654. doi: 10.1111/tpj.14853, PMID: 32445404 · doi ↗ · pubmed ↗
- 8Krouk G. Lacombe B. Bielach A. Perrine-Walker F. Malinska K. Mounier E. . (2010). Nitrate-regulated auxin transport by NRT 1. 1 defines a mechanism for nutrient sensing in plants. Dev. Cell 18, 927–937. doi: 10.1016/j.devcel.2010.05.008, PMID: 20627075 · doi ↗ · pubmed ↗