Oral-heart axis from pregnancy and postpartum: maternal oral microbiota relates with cardiac reverse remodeling
Juliana Morais, Maria João Azevedo, Ana Filipa Ferreira, Bernd W. Brandt, Egija Zaura, Mark J. Buijs, Adelino F. Leite-Moreira, Carla Ramalho, António Barros, Inês Falcão Pires, Benedita Sampaio-Maia

TL;DR
This study shows that a mother's oral bacteria during pregnancy are linked to how well her heart recovers after giving birth.
Contribution
The study identifies specific oral bacteria associated with postpartum cardiac reverse remodeling.
Findings
Porphyromonas during pregnancy predicts less favorable heart recovery after delivery.
Higher levels of certain oral bacteria after birth correlate with better heart recovery.
Microbiota-cardiac associations were confirmed independently of diet.
Abstract
Emerging evidence links the oral microbiota to cardiovascular health, but this relationship remains poorly understood during pregnancy. Here, we investigated associations between the oral microbiota, cardiovascular physiology, and diet during and after pregnancy. Salivary microbiota from 65 women in the 3rd trimester and 6 months postpartum were analyzed by 16S rRNA gene sequencing. Cardiovascular function was assessed via echocardiography, endothelial function by EndoPATTM, nitric oxide levels by plasma nitrate/nitrite levels, and diet through the Food Frequency Questionnaire. Left ventricular end-diastolic volume (LVEDV) was negatively associated with nitrite-reducing bacteria, namely, Prevotella, at both time points. Cluster analysis identified two reverse-remodeling profiles, one with poorer remodeling, greater postpartum weight retention and higher pregnancy microbial richness…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
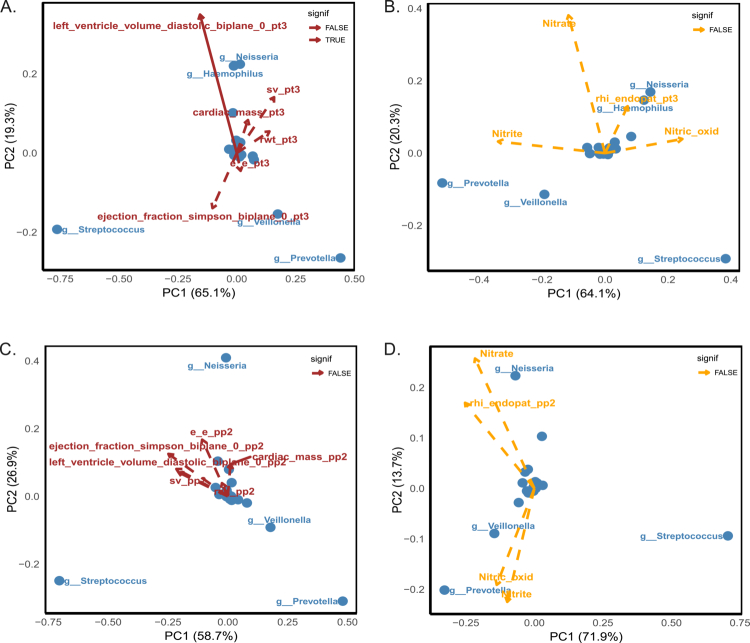 Figure 1
Figure 1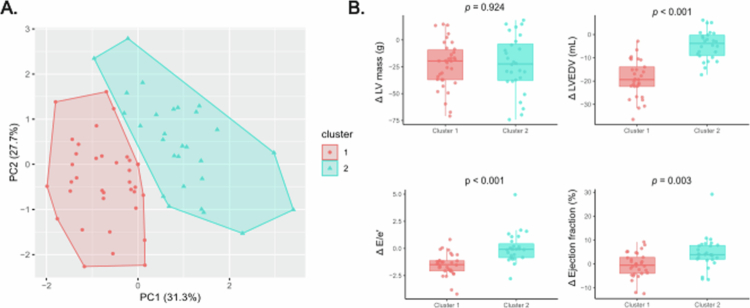 Figure 2
Figure 2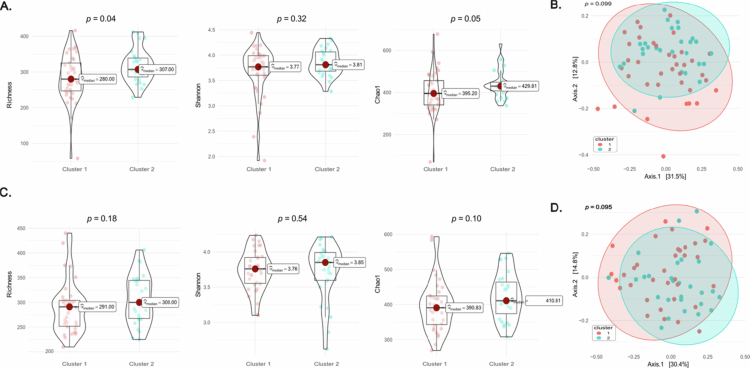 Figure 3
Figure 3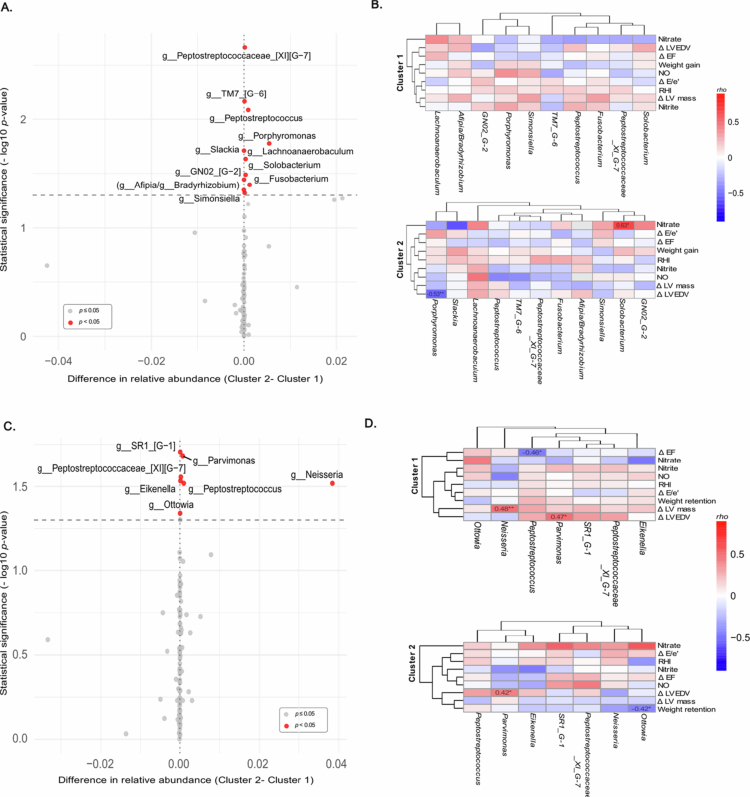 Figure 4
Figure 4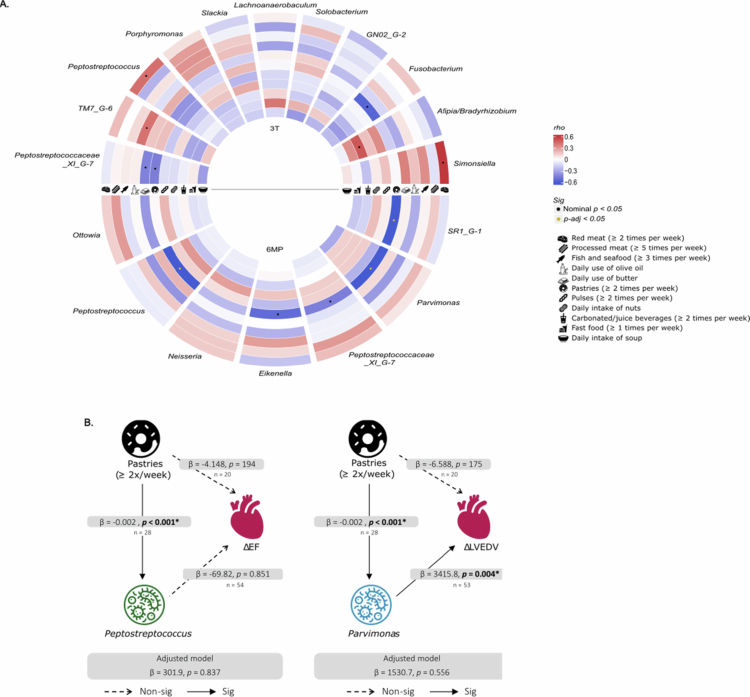 Figure 5
Figure 5| Maternal clinical data ( | |
|---|---|
| Maternal age (years), median [IQR] | 35 [32; 37] |
| Education level, | |
| Basic | 1 (1.5) |
| High school | 20 (30.8) |
| Bachelor’s degree | 27 (41.5) |
| Master’s degree | 13 (20.0) |
| Doctoral degree | 4 (6.2) |
| Smoking habits, | |
| Yes | 5 (7.7) |
| No | 60 (92.3) |
| Pregestational BMI (kg.m−2), median [IQR] | 23.1 [21.5; 28.3] |
| Pregestational BMI category, | |
| Underweight | 1 (1.5) |
| Normal weight | 39 (60.0) |
| Overweight | 14 (21.5) |
| Obesity | 11 (17) |
| Pregestational diabetes (T2D), | 1 (1.5) |
| Gestational diabetes, | 8 (12.5) |
| Chronic hypertension, | 14 (21.5) |
| Gestational hypertension, | 1 (1.5) |
| Gestational weight gain (kg), median [IQR] | 10.1 [7.00; 13.0] |
| Postpartum weight recovery at 6MP (kg), median [IQR] | 2.14 [-1.00; 6.00] |
| Gestational age at birth (weeks), median [IQR] | 39 [38; 39] |
- —European Society of Clinical Microbiology and Infectious Diseases
- —European Regional Development Fund (ERDF)
- —Biocodex
- —Foundation for Science and Technology FCT
- —Dutch Research Council (NWO)
- —Foundation for Science and Technology FCT
- —the Portuguese Foundation for Science and Technology (FCT)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOral microbiology and periodontitis research · Infective Endocarditis Diagnosis and Management · Streptococcal Infections and Treatments
Introduction
Over the course of gestation, profound physiologic changes occur in the woman's body, significantly affecting the endocrine, metabolic, immune, and microbial systems [1,2]. In the early period of pregnancy, an increase in systemic vasodilation and a higher cardiac output are observed [3]. These alterations persist throughout pregnancy, causing hypertrophy and dilation of the heart due to blood volume expansion, which normalises during the postpartum period through a process known as cardiac reverse remodelling [4].
Changes also occur in the oral cavity of pregnant women. An increase in hormones (oestrogen and progesterone) is associated with gingival inflammation and, consequently, periodontal diseases [5]. The maternal oral microbiota, which encompasses various oral niches [6], has gathered attention because of its potential impact on pregnancy outcomes. As reviewed by Saadaoui and colleagues (2021), a dysbiotic oral environment has been associated with miscarriage, preeclampsia, low birth weight, and preterm birth [7].
Despite physical distance, emerging evidence shows associations between oral dysbiosis and cardiovascular diseases [8]. Two possible mechanisms have been described to explain how the oral microbiota could contribute to systemic diseases. The first mechanism involves the direct entry of bacteria into the systemic circulation through an existing breach in the oral epithelium [9], which may be inflamed and ulcerated during pregnancy [10]. A meta-analysis of 63 studies concluded that 23 oral commensal bacterial species were present in atherosclerotic plaques, either individually or in co-existence [11]. Of these, 5 were unique to coronary plaques, while the remaining 18 were found in non-cardiac tissues [11]. The second mechanism involves the translocation of bacterial byproducts, toxins, and related pro-inflammatory cytokines that promote chronic low-grade inflammation in other tissues or affect cardiovascular physiology [12]. One of these products is the result of the microbial nitrate‒nitrite‒nitric oxide (NO) pathway. In saliva, nitrate-reducing bacteria convert nitrate to nitrite. In the stomach, owing to its acidic pH, nitrite is reduced to NO [13]. NO is a key molecule with vasoactive and endothelial protective effects associated with reduced blood pressure, ischaemia‒reperfusion injury and atherosclerosis [13].
In this study, we aimed to investigate the association between the salivary microbiota and cardiovascular health, specifically cardiac reverse remodelling and endothelial function, across the transition from late pregnancy to postpartum, and to assess whether maternal diet influences this oral-heart axis.
Materials and methods
Cohort design and ethical considerations
In this nested study within the longitudinal, prospective observational cohort PERIMYR/OralBioBorn, women were evaluated during the 3^rd^ trimester (3 T, 30‒35 gestational weeks) (n = 65) and 6 months postpartum (6MP, n = 63). Pregnant women were recruited at the Unidade Local de Saúde de São João (ULS São João), and the Unidade Local de Saúde de Matosinhos (ULSM). The exclusion criteria were age under 18 years, pre-existing cardiomyopathy, renal disease, chronic obstructive airway disease, active systemic infection, genetic syndromes, or type 1 diabetes Mellitus. The Ethical Committees of ULS São João and ULSM approved the study in 2018, with reference numbers 294/2018 and 86/CE/JAS, respectively. The study was conducted in accordance with the principles of the Declaration of Helsinki, and all eligible participants provided written informed consent to be included in this study.
Clinical data collection
Detailed clinical data were collected through extensive questionnaires that included information about pregestational chronic disease, obstetric outcomes (gestational hypertension, gestational diabetes and weight gain), maternal smoking and oral healthcare habits.
Maternal dietary habits assessment
A subgroup of 23 and 26 women completed the 86-item validated Food Frequency Questionnaire (FFQ) to assess their dietary intake during pregnancy and postpartum, respectively. The response options followed a 9-point frequency scale, ranging from ‘never’ to ‘≥6 times per day’. Data from the FFQ were used to explore the consumption of specific key food groups based on the Mediterranean Diet Adherence Screener [14]. The key food groups and respective intake thresholds studied included: red meat (≥2 servings/week); processed meat (≥5 servings/week); fish and seafood (≥3 servings/week); daily use of olive oil; daily use of butter; pastries (≥2 servings/week); pulses (≥2 servings/week); daily intake of nuts; carbonated/juice beverages (≥2 servings/week); fast food (≥1 servings/week); and daily intake of soup. These categories were classified as ‘YES’ or ‘NO’ based on whether the threshold was met.
Echocardiography and endothelial function assessment
Echocardiographic assessment of cardiac function and structure was performed using a conventional transthoracic echocardiography performed using a 3 MHz phased-array probe (ACUSON SC2000 PRIME) by a single operator. Left ventricular (LV) mass (g) was estimated using the following formula: LV mass = 0.8 × {1.04 × [(LVIDd + PWTd + IVSd)³–(LVIDd)³]} + 0.6, where LVIDd = LV internal diameter at end-diastole, PWTdP = posterior wall thickness in diastole and IVSd = interventricular septal thickness in diastole. The biplane Simpson method was used to estimate the left ventricular end-diastolic volume (LVEDV), stroke volume and ejection fraction (EF). Additionally, E/e’ was calculated using the ratio of E mitral inflow velocity to the average early diastolic septal velocity with the lateral mitral annular velocity.
Endothelial function was assessed by measuring endothelium-mediated changes in vascular tone using two-finger probes on each index finger. This was performed with the non-invasive Peripheral Arterial Tone (PAT) signal technology system EndoPAT^TM^ 2000 (Itamar Medical, Israel). In brief, participants were placed in the supine position during the 15-minute assessment. Following a 5-minute rest period, a blood pressure cuff was inflated to one arm to a supra-systolic level (at least 60 mmHg above the systolic levels, typically between 200 and 300 mmHg) and maintained for 5 min to induce occlusion. After the cuff was released, reactive hyperaemia was monitored for an additional five minutes. The reactive hyperaemia index (RHI) was automatically calculated by the EndoPAT^TM^ software using the following formula: (C/D) ÷ (A/B), adjusted by a baseline correction factor. Endothelial dysfunction was considered when RHI was below 1.67 [15].
Blood sample collection and indirect measurement of nitric oxide
Blood samples were collected and centrifuged for 15 min at 3857 × g at 4 °C. Nitric oxide production was estimated by measuring the relative levels of nitrate and nitrite in plasma (Invitrogen, Thermo Fisher). Briefly, to quantify nitrite concentrations, 25 μL of plasma was diluted 1:4 and analysed using the Griess reaction, a colorimetric assay. For nitrate determination, the remaining nitrate in the sample was enzymatically reduced to nitrite using nitrate reductase. The resulting total nitrite (the pre-existing nitrite plus nitrate that was converted) was then measured again with the Griess assay. The nitrate concentration was calculated by subtracting the initial nitrite value from the total nitrite value. The sum of both nitrite and nitrate concentrations represents the estimated total NO production.
Clusters identification
To explore cardiac reverse remodelling profiles, we performed cluster analysis on echocardiographic changes (Δ) between 3T and 6MP, including ΔLV mass, ΔLVEDV, ΔEF and ΔE/e’. Among the 65 women included in the study, echocardiographic data at 6MP were available for 63, as two participants were lost to follow-up, and these missing values were imputed using the variable-specific mean. The optimal number of clusters was first estimated using the elbow method applied to the PCA scores. Based on this analysis, a two-cluster solution was selected. We then performed hierarchical clustering analysis on principal components (HCPC; FactoMinerR package, v. 2.11 [16]).
Saliva collection
Participants were asked to collect unstimulated saliva 1 h after a meal or toothbrushing by passively drooling into a 50 mL sterile tube until 5 mL was collected. All samples were placed on ice until they were divided into 1 mL aliquots, after which they were stored at −80 °C until further analysis.
Microbial analysis and data processing
Bacterial DNA extraction and quantification were described previously [17]. In brief, DNA was isolated from 200 μL of each saliva samples using the MagMini DNA isolation kit (LGC Genomics Ltd., UK). Quantification of the extracted DNA was carried out via qPCR using universal primers targeting the 16S rRNA gene. Sequencing was conducted on the MiSeq system (Illumina, San Diego, CA, USA) at TNO (Zeist, The Netherlands). The paired-end reads obtained from the MiSeq platform were merged and quality-filtered, and contaminant PhiX sequences were removed. High-quality reads were denoised using the UNOISE algorithm (USEARCH v10.0.240), producing zero-radius operational taxonomic units (zOTUs). Representative zOTU sequences were taxonomically assigned using the Human Oral Microbiome Database [18] as previously described [19]. Microbiome data analysis was performed using R (version 4.4.3 [20]). Based on the distribution of reads across samples and to retain sufficient depth while minimising sample loss, the samples were randomly subsampled to an equal depth of 4903 using the rarefy_even_depth function (phyloseq v. 1.50.0. [21]). Alpha diversity was calculated on the rarefied dataset (4903 counts per sample) at the zOTU level using the estimate_richness function from the phyloseq package. For beta diversity and taxonomic composition analyses, counts were normalised to relative abundances using Total Sum Scaling (TSS). No Centred Log Ratio (CLR) transformation was applied, as recent evidence suggests that CLR may retain sequencing depth-related signals that can affect ordination-based analyses [22]. Instead, samples were rarefied to a common sequencing depth to control for uneven sequencing effort.
Statistical analysis
Clinical descriptive statistics were performed by SPSS version 30 (IBM SPSS Statistics, Chicago, IL, USA). Continuous variables are expressed as medians with interquartile ranges, and categorical variables are expressed as absolute values and relative frequencies. Clinical and microbial differences between clusters at each timepoint were evaluated using non-parametric tests (Wilcoxon rank-sum test for continuous variables and Fisher's exact test for categorical variables). Statistical significance was considered if p < 0.05.
To explore associations between the salivary microbiota composition and echocardiographic/vascular variables, biplots of relative abundances at the genus level were generated for each timepoint. The environmental fitting of variables onto the PCA ordination was conducted with the envfit function (999 permutations, vegan R package v. 2.6.10. [23]). Genera showing strong associations in biplots (based on the envfit results) were subsequently included in linear regression analyses to quantify their associations with selected echocardiographic parameters. Multivariable linear regression models were fitted with echocardiographic measurements as dependent variables, adjusting for clinical covariates previously identified as key determinants of cardiac structure and function [24], namely, previous hypertension, pregestational obesity, smoking habits and weight change (weight gain up to 3T or weight retention at 6MP, depending on the analysis timepoint). This model is referred to as the ‘clinical model’ hereafter.
Alpha-diversity indices, namely, the Shannon, Chao1, and observed richness, were estimated on a rarefied dataset using the estimate_richness function (phyloseq v. 1.50.0 [21]). and compared between cardiac reverse remodelling clusters using the Wilcoxon rank-sum test. For alpha-diversity indices showing significant differences between clusters, sensitivity analyses were performed using linear regression models with cluster membership as the main predictor, adjusting for maternal age and pre-pregnancy body mass index (BMI).
For each timepoint, the beta diversity was assessed using Bray–Curtis distances computed from zOTU-level relative abundance data (distance, method = ‘bray’ from the phyloseq package), and principal coordinate analysis (PCoA) was performed with the ordination function. Differences in microbial community composition between clusters were tested by permutational multivariate analysis of variance (PERMANOVA) with 9999 permutations, as implemented in the adonis2 function (vegan package), after verifying the homogeneity of multivariate dispersions (betadisper).
Comparisons of genus-level relative abundance between clusters were performed using Wilcoxon rank-sum tests with the Benjamini‒Hochberg false discovery rate (FDR) correction method. For visualisation purposes, volcano plots were generated to illustrate the magnitude of differences (Cluster 2–Cluster 1, based on mean relative abundance) against the strength of the association (−log10 p-value). In these plots, unadjusted p-values were used to better display overall trends, whereas formal statistical interpretation relies exclusively on FDR-adjusted values. To explore potential relationships between bacterial genera and clinical variables, Spearman correlation coefficients were computed using corr.test function (psych package, v. 2.5.6. [25]). Correlation matrices were visualised as clustered heatmaps with the pheatmap package v 1.0.13. [26]). Associations between genus-level taxa that were significantly different between clusters and dietary variables were explored using Spearman’s correlations. Further regression analysis was performed to examine the interplay between diet, oral microbiota, and echocardiographic parameters. During data analysis, artificial intelligence-based tools (ChatGPT, OpenAI and GPT-5.2) were used to assist in draughting and debugging R scripts. All analyses were reviewed, validated and interpreted by the authors.
Results
Participants characterisation
A total of 65 pregnant women were recruited and followed from April 2019 to January 2022. The participating women had a median age of 35 years [IQR: 32; 37] at the moment of the first visit (3T) and a median pregestational BMI of 23.1 [IQR: 21.5; 28.3] kg.m^−2^. Most of the participants (67%) were highly educated and held a bachelor's degree (41.5%), master’s degree (20%) or doctoral degree (6%), and 7.7% smoked during the 3T of pregnancy. Maternal sociodemographic and clinical data are reported in Table 1.
Regarding maternal oral care, at 3 T of pregnancy, most women brushed their teeth 2 times a day (61.5%) and used a toothbrush complement, such as an elixir, dental tape, or water jet (83.1%) (Table S1). In the year before the first visit (3T), 21.5% of participants never went to the dentist, 32.3% and 20.0% went once or twice, respectively, primarily for dental cleaning (Table S1).
Among the participants who completed the study and provided saliva samples at both visits (3T and 6MP), only 23 filled the FFQ at the 3rd trimester and 28 – at 6 months postpartum. With these data, no differences were found in the consumption of the food items explored (Table S2).
Regarding the cardiovascular characteristics, we observed significant differences between the gestational period and the postpartum period (Table S3). Echocardiographic assessment revealed a significant reduction in relative wall thickness (0.35 [0.31; 0.40] vs 0.31 [0.28; 0.36], p < 0.001), LV mass (32.9 [28.0; 38.2] vs 26.7 [23.0; 30.2], p < 0.001), and LVEDV (24.5 [22.7; 26.8] vs 21.8 [19.7; 23.8], p < 0.001). While the stroke volume and ejection fraction remained unchanged, the E/E’ ratio decreased (6.76 [5.76; 7.89] vs 5.70 [5.22; 6.51], p = 0.001) (Table S3).
The endothelial function was assessed through the RHI, which did not significantly change (1.53 [1.39; 1.73] vs 1.50 [1.32; 2.18], p = 0.676) (Table S3). After categorising participants in two groups regarding the RHI cut-offcutoff of 1.67, an indicator of endothelial dysfunction, the prevalence of endothelial dysfunction decreased from 71.7% at 3T to 60.0% at 6MP. However, this difference did not reach statistical significance (p = 0.254). Despite no differences observed in nitrate levels between the two timepoints, there was a significant reduction in nitrate-derived estimates of NO production (454.1 [383.9; 658.5] vs 330.8 [256.0; 460.4], p = 0.007) and nitrite levels (383.99 [331.6; 466.8] vs 285.4 [236.7; 369.8, p < 0.0019 (Table S3).
Association between oral microbiota and cardiovascular structure and function
To explore the relationship between the oral microbiota and cardiovascular parameters, we generated biplots illustrating the distribution of bacterial genera in relation to echocardiographic and endothelial parameters at 3T and 6MP (Figure 1). During pregnancy, Prevotella and Veillonella were positioned in the opposite direction as the vector for LVEDV (left_ventricle_volume_diastolic_biplane_pt3) (Figure 1A). On the other hand, Haemophilus and Neisseria were also located in the same direction as the LVEDV and closer to the vector of stroke volume (sv_pt3) (Figure 1A), RHI (rhi_endopat_pt3), and plasma nitrate (nitrate) and indirectly measured NO (Nitric_oxid) (Figure 1B). In the postpartum period, despite Prevotella and Veillonella remaining positioned in the quadrant opposite to the LVEDV vector, the cardiac parameters are short and centrally located, indicating a limited contribution to the variance explained by PC1–PC2, whereas the dispersion of oral genera appears to account for most of the visible separation among participants (Figure 1C). With respect to the vascular markers, Neisseria was associated with the direction of the RHI and nitrate vectors, suggesting a positive association, although this pattern did not reach statistical significance (Figure 1D).
Principal component analysis (PCA) biplots illustrating the relationship between bacterial genera and cardiovascular outcomes during pregnancy (A and B) and six months postpartum (C and D). Panels A and C depict associations with echocardiographic parameters (red vectors), whereas panels B and D show endothelial function and nitric oxide-related biomarkers (yellow vectors). The blue dots represent individual bacterial genera. The direction and length of the vectors reflect the direction and strength of the correlations. The solid arrows indicate statistically significant associations (p < 0.05), while dashed arrows denote non-significant trends.
Based on the PCA results, in which only the LVEDV showed meaningful correlations with bacterial genera (Figure 1A), this variable was selected for further statistical modelling. Linear regression models were constructed to explore associations between LVEDV and the five genera that were most prominent in the PCA biplots (Neisseria, Haemophilus, Veillonella, Prevotella, and Streptococcus) (Table S4). During pregnancy, in both the unadjusted model (β = –34.3, p = 0.026) and the clinical model adjusted for arterial hypertension, obesity, smoking habits, and weight gain until the 3 T visit (see methods section), the relative abundance of Prevotella (β = −40.3, p = 0.004) remained significantly associated with LVEDV (Table S4). At 6MP, Prevotella remained significantly associated with the LVEDV only in the adjusted clinical model (β = −26.8, p = 0.025) (Table S4). No significant associations were observed for Haemophilus, Veillonella, or Streptococcus in either model (Table S4).
Cluster of participants based on cardiac reverse remodelling profile
Given the variability observed in echocardiographic parameters at 6MP, we stratified participants into clusters reflecting different cardiac reverse remodelling trajectories, with 2 identified clusters (Figure 2A). Cluster 1 (n = 37) comprised women with a more favourable cardiac reverse remodelling profile, showing greater recovery in three of the assessed parameters, namely, the LVEDV, E/e’, and ejection fraction (Figure 2B). In contrast, Cluster 2 (n = 28) had incomplete or less efficient postpartum reverse remodelling, characterised by significantly smaller changes in LVEDV, E/e’, and ejection fraction, despite a similar reduction in LV mass compared with Cluster 1 (Figure 2B).
Clustering of participants according to dynamic changes (Δ) in echocardiographic parameters between 3T and 6MP. (A) Principal component analysis (PCA) plot showing the projection of the HCPC-defined clusters, with Cluster 1 (red) and Cluster 2 (blue). (B) Boxplots of the echocardiographic Δ-variables used to define the clusters, including the ΔLV mass, ΔLVEDV, ΔE/e’ and Δ Ejection fraction.
When comparing maternal characteristics between clusters, women in Cluster 2 had significantly greater weight retention during the first 6 months postpartum than those in Cluster 1 (Table S5). No significant differences were observed for age, pre-existing obesity, diabetes, hypertension, gestational complications, and gestational weight gain up to T3 (Table S5). Subsequent microbiota analyses were conducted according to these clusters to explore potential microbial signatures of divergent recovery patterns.
Alpha- and beta-diversity of the oral microbiota in the cardiac reverse remodelling clusters
At 3T, Cluster 2 showed significantly higher community richness (p = 0.04, Figure 3A). However, in sensitivity analyses adjusting for maternal age and pre-pregnancy BMI, this difference was attenuated and did not remain statistically significant (adjusted p = 0.074). Other alpha-diversity indices, including the Shannon and Chao1 diversity did not differ between the clusters (Figure 3A). Similarly, beta-diversity analysis based on Bray‒Curtis distances revealed no separation between clusters at 3T (PERMANOVA p = 0.099, Figure 3B), and no differences were observed in any alpha-diversity index (Figure 3C), nor in beta diversity among clusters (PERMANOVA p = 0.095, Figure 3D).
Comparison of oral microbiota diversity indices between the two clusters of cardiac reverse remodelling at 3T (A and B) and at 6MP (C and D). Alpha-diversity indices (richness, Shannon diversity index, and Chao1) are shown as violin plots (A and C). Beta diversity was measured by Bray‒Curtis distances using principal coordinate analysis (PCoA) (B and D).
Taxonomical changes of the oral microbiota in the cardiac reverse remodelling clusters and network with cardiovascular outcomes
The overall taxonomic distribution was broadly similar between the two clusters and across time. At the phylum level, Bacillota dominated in both groups during pregnancy (~54–56%), followed by Bacteroidota (17%) and Pseudomonadota (12–14%) (Table S6). At 6MP, the relative abundance of the Absconditabacteria phyla was significantly higher (0.016% vs 0.049%, p-adj = 0.04), as well as Fusobacteriota with a significant nominal difference (3.10% vs 3.87%, p = 0.030) (Table S6). At the genus level, Streptococcus was consistently the most prevalent genus during pregnancy (~39–40%), followed by Prevotella (13–14%) and Veillonella (8–10%) (Table S7). After delivery, we observed an increase in the relative abundance of Porphyromonas (1.203% vs 2.293%, p = 0.001) and SR1_G-1 (0.016% vs 0.049%, p = 0.04) (Table S7).
Although between-group comparisons identified several nominal differences in the relative abundance, many of these genera were present at an extremely low abundance and should therefore be interpreted with caution. During 3T, women in Cluster 2 exhibited a higher relative abundance of Peptostreptococcaceae_[XI][G-7] (0.023% vs 0.005%, p = 0.002), TM7_[G-6] (0.023% vs 0.011%, p = 0.007), Peptostreptococcus (0.244% vs 0.152%, p = 0.008), Porphyromonas (1.512% vs 0.969%, p = 0.017), Slackia (0.000% vs 0.001%, p = 0.019), Lachnoanaerobaculum (0.220% vs 0.185, p = 0.023), Solobacterium (0.087% vs 0.052%, p = 0.032), GN02_[G-2] (0.000% vs 0.002%, p = 0.036), Fusobacterium (1.411% vs 1.290%, p = 0.040) and Simonsiella (0.016% vs 0.002%, p = 0.045) compared to Cluster 1 (Figure 4A; Table S8). Cluster 1 showed higher relative abundance of taxa assigned to the genera Afipia or Bradyrhizobium (which could not be distinguished via 16S rRNA analysis) (0.003 vs 0.001, p = 0.044) (Figure 4A; Table S8).
*Taxonomix changes between clusters at 3T (A and B) and at 6MP (C and D) and association with clinical variables. Volcano plots showing genus-level differences in relative abundance between the two cardiac remodelling clusters at 3T (A) and at 6M (C). Heatmaps of Spearman correlations between the differentially abundant bacterial genera and echocardiographic and clinical variables at 3T (B) and at 6M (D). The colours represent correlation coefficients (red = positive, blue = negative), with significance indicated in the boxes (p < 0.05).
Genera that showed significant differences in relative abundance between clusters were further explored for correlation with Δ-echocardiographic parameters, endothelial biomarkers, and gestational weight gain or postpartum weight retention (Figure 4B). No associations were observed between the levels of the mentioned genera and clinical variables in Cluster 1. On the other side, in Cluster 2, we observed a strong positive correlation between Solobacterium and nitrate levels (rho = 0.63, p = 0.036), as well as a negative association between Porphyromonas and ΔLVEDV (rho = −0.53, p = 0.005). After adjustment for the clinical model (adjusted for arterial hypertension, obesity, smoking habits and weight gain until 3T), the correlation observed between Solobacterium and nitrate levels was not confirmed (ẞ = 20.4, p = 0.728), indicating that this association was not robust. In contrast, Porphyromonas remained independently associated with a lower ΔLVEDV (ẞ = −280.1, p = 0.003). The adjusted model explained 38% of the variance in ΔLVEDV (adjusted R^2^ = 0.38).
At 6MP, Cluster 2 also showed nominal significantly higher proportion of SR1 (0.054% vs 0.045%, p = 0.020), Parvimonas (0.155% vs 0.083%, p = 0.021), Peptostreptococcaceae_[XI][G-7] (0.034% vs 0.009%, p = 0.028), Eikenella (0.028% vs 0.014%, p = 0.029), Neisseria (10.6% vs 6.73%, p = 0.030), Peptostreptococcus (0.271% vs 0.172%, p = 0.030) and Ottowia (0.011% vs 0.004%, p = 0.046) (Figure 4C; Table S9). Among participants of Cluster 1, higher levels of Neisseria and Parvimonas were significantly correlated with ΔLVmass (rho = 0.48, p = 0.006) and ΔLVEDV (rho = 0.47, p = 0.013), respectively. After adjustment for the clinical model (adjusted for arterial hypertension, obesity, smoking habits and weight retention until 6MP visit), both genera remained associated with the ΔLV mass (Neisseria: ẞ = 181.5, p = 0.023 and Parvimonas: ẞ = 6537.2, p = 0.046). A negative correlation was observed between Peptostreptococcus and ΔEF (rho = -0.46, p = 0.017) and confirmed by the clinical model (ẞ = -999.6, p = 0.013) (Figure 4D).
As observed in Cluster 1, in Cluster 2, it was also observed that Parviromonas correlated with ΔLVEDV (rho = 0.42, p = 0.033), but this association was lost after adjusting for the clinical model (ẞ = 1121.8, p = 0.203). In addition, in Cluster 2, a negative correlation was observed between Ottowia and weight retention (rho = −0.42, p = 0.026).
Interaction between diet-oral microbiota and cardiovascular parameters
We further explored whether bacterial genera showing nominal differences between clusters were associated with dietary habits (Figure 5). In the third trimester, several nominal correlations were found between diet and bacterial genera. Simonsiella was positively correlated with red meat intake (≥2 times per week; rho = 0.50, p = 0.016). higher consumption of pastries during the 6MP was negatively associated with several genera after FDR correction, including adjusted p-values, namely, SR1_G-1 (rho = −0.72, p-adj = 0.002), Parvimonas (rho = −0.69, p-adj = 0.003) and Peptostreptococcus (rho = −0.61, p-adj = 0.016) (Figure 5A). To evaluate whether pastry intake accounted for the association between Peptostreptococcus and ΔEF, and between Parvimonas and ΔLVEDV, we fitted regression models adjusted for pastry consumption. After adjustment, neither association remained statistically significant, indicating that these relationships were not independent of pastry intake (Figure 5B).
Associations between diet and the oral microbiota. (A) Heatmap showing Spearman correlations between bacterial genera that showed nominal differences and maternal intake of selected food groups during pregnancy and postpartum. (B) Regression models exploring the relationships among ΔEF or ΔLVEDV (dependent variables) and Peptostreptococcus or Parvimonas, respectively, adjusted for pastry consumption.
Discussion
In this study, we characterised maternal cardiovascular remodelling from late pregnancy to 6 months postpartum and investigated its associations with the salivary microbial composition and dietary habits as part of a larger project in which the cardiovascular [24] and oral microbiota [19] components were independently explored. As expected, we observed a postpartum normalisation of cardiac structure and diastolic function to pre-gestational values, including reductions in relative wall thickness, LV mass, LVEDV and E/e’. Interestingly, our initial analysis revealed that LVEDV was negatively associated with nitrate-reducing oral bacteria, particularly Prevotella, at both time points. We then identified two distinct cardiovascular remodelling profiles through cluster analysis. One subgroup of women exhibited less efficient postpartum cardiac recovery and greater weight retention, and was characterised by higher microbial richness in unadjusted analyses during pregnancy and a sustained abundance of inflammation-associated genera such as Porphyromonas. In contrast, health-associated taxa, including Neisseria and Parvimonas are associated with more favourable remodelling trajectories.
There is limited published data available that links cardiac structure and function to changes in the oral microbiota [8]. In our cohort, we observed that the relative abundance of Prevotella, a Gram-negative bacterial genus, was inversely associated with the LVEDV at 3T and at 6MP even after adjustment for the clinical model. LVEDV is defined as the telediastolic volume and is a structural marker of cardiac remodelling. While in pregnancy, a higher LVEDV typically reflects a better capacity of the left ventricle to accommodate increased blood volume, in the postpartum period, the LVEDV is expected to decrease to pregestational volumes [27]. Previous studies have shown associations between a higher proportion of bacteria from the genus Prevotella and hypertension [28] and the risk of cardiovascular disease [29], by triggering pro-inflammatory mechanisms. Specifically, during pregnancy, increased progesterone is associated with an increase in Prevotella spp., many of which are associated with periodontal diseases [30]. In addition, Prevotella is listed as one of the nitrate-reducing bacteria capable of influencing the NO production and thereby enhancing vasodilation [31].
Beyond their ability to activate inflammatory pathways and induce NO production, several Prevotella strains metabolise sulphur-containing amino acids, producing hydrogen sulphide (H_2_S) [32,33]. H_2_S has been shown to increase NO bioavailability through synergetic signalling interactions [34]. As NO is a potent endothelial vasodilator, higher NO levels reduce systemic vascular resistance and, consequently, lower afterload, which may contribute to a smaller LVEDV. Given this, we hypothesise that lower Prevotella abundance results in lower NO production (directly and indirectly via H_2_S), leading to less vasodilation and, therefore, higher LVDEV.
In line with the physiological cardiovascular adaptations known to occur during and after pregnancy [24,35], the echocardiographic findings of this study demonstrate a pattern of postpartum cardiac remodelling and functional recovery, as evidenced by significant reductions in relative wall thickness, LV mass, LVEDV and E/e’ at 6MP. These changes suggest a reversal of volume and pressure overload, typically in late gestation, corresponding to the normalisation of hemodynamic demands postpartum. However, we found two different clusters regarding the cardiac reverse remodelling profile, with Cluster 2 showing less favourable cardiac reverse remodelling.
During pregnancy, Cluster 2 showed a tendency toward higher community richness with increased levels of inflammatory bacterial genera, namely, Peptostreptococcus [36], Porphyromonas [37], Fusobacterium [38] and Solobacterium [39]. Among these genera*, Porphyromonas* remained associated with a lower ΔLVEDV after adjustment for the clinical model, suggesting its potential role as a predictor of poorer cardiac reverse remodelling. Although an additional 20 species exist within this genus, the available literature has focused mainly on the systemic effect of Porphyromonas gingivalis [37]. This bacterium is highly prevalent in the human oral cavity and possesses several virulence features, including the presence of gingipains and lipopolysaccharides (LPS) in its outer membrane, which can activate the Toll-like receptors TLR2 and TLR4 and thereby trigger inflammation [40]. As reviewed by Zhang et al., the inflammatory state induced by Porphyromonas spp. contributes to endothelial oxidative stress, upregulating inducible nitric oxide synthase (iNOS) and downregulating endothelial nitric oxide synthase (eNOS) [40]. Thus, a reduction in NO-mediated vasodilation leads to increased afterload, which may contribute to increased LVEDV, as observed in our cohort. Supporting this mechanism, a recent study in mice treated with P. gingivalis reported elevated systemic pro-inflammatory markers, a reduced ejection fraction, increased LV dimensions on echocardiography, and increased myocardial levels of matrix metalloproteinases [41].
During the postpartum period, Cluster 2 no longer presented increased alpha diversity but continued to show increased abundances of Peptostreptococcaceae_[XI][G-7] and Peptostreptococcus compared to Cluster 1, as well as increased levels of Parvimonas, Eikenella, Neisseria and Ottowia. In our study, we showed a positive correlation between Parvimonas and the change in LVEDV in both clusters, suggesting that a higher abundance of this genus may be linked to more favourable reverse remodelling. However, in Cluster 2, this association was lost after adjustment for the clinical model. The genus Parvimonas has only three species identified to date (https://lpsn.dsmz.de/search?word=parvimonas), with P. micra being the most common [42]. P. micra is not only associated with oral infections but also with endocarditis [43] because of its high proteolytic activity and H_2_S toxicity, as reviewed by Shimizu [44]. More studies are needed to confirm the association between Parvimonas and greater LVEDV.
In our study, we observed that the Ottowia genus was associated with greater postpartum weight retention in Cluster 2, which showed higher weight retention compared with Cluster 1. Although Ottowia is not a highly abundant or frequently reported genus in oral microbiota studies, it is significantly enriched in the oral microbiota of healthy individuals when compared with the myocardial infarction group [45].
Cluster 1, characterised by a more favourable cardiac reverse remodelling profile, showed a higher abundance of Neisseria than Cluster 2. Interestingly, within this cluster, higher levels of Neisseria were positively associated with greater recovery of the LV mass during the postpartum period. Certain species of Neisseria have been linked to endothelial activation [46], which may contribute to subtle cardiovascular adaptations. Growing evidence supports the role of the oral microbiota in modulating systemic vascular tone via the NO pathway. As Neisseria is a well-recognised nitrate-reducing bacterium [47], enhanced activity of the enterosalivary nitrate‒nitrite‒NO pathway and thereby the bioavailability of NO. This could be one plausible mechanism that relates higher levels of Neisseria to improved LV mass recovery. It was also observed that higher levels of Peptostreptococcus were also associated with a smaller change in the ejection fraction, suggesting that the presence of this bacterial genus may impair cardiac reserve remodelling. Peptostreptococcus genus is a Gram-positive anaerobic coccus that can act as a commensal organism but also as an opportunistic pathogen, which is typically linked to oral infection and, although rare, with infective endocarditis [48]. However, there is evidence that Peptostreptococcus spp. can induce an inflammatory state capable of affecting cardiac function [49], no direct mechanism has yet been identified to explain its influence on the ejection fraction.
Noteworthy findings confirm the impact of pastries on the oral microbiota, showing that this effect is evident in pregnant women during pregnancy and becomes particularly significant after birth. In general, we found that participants who frequently consumed pastries during pregnancy had lower levels of Peptostreptococcaceae_[XI][G-7] and Fusobacterium. After birth, pastries promoted lower levels of Peptostreptococcaceae_[XI][G-7], Parvimonas, Peptostreptococcus, Eikenella and SR1_G-1. These findings seem counterintuitive because they suggest that the consumption of sugar and refined carbohydrates could have a potential protective effect against certain pathogenic bacteria, as these genera are often associated with inflammation [50]. The shift in bacterial populations might not necessarily confer a health benefit but rather indicate a microbiome adjustment to dietary patterns that are generally considered less healthy, such as frequent pastry consumption. A previous study that included 8,173 adolescents showed that high salivary glucose, which is associated with the intake of pastries, reduced the overall bacterial load [51]. The authors propose a possible hypothesis for this phenomenon: the increase in salivary glucose caused by hyperglycaemia status promotes higher levels of bacteria that synthesise acidic metabolites that, owing to the lower levels of pH in the oral cavity, promote a decrease in salivary bacterial numbers with a change in bacterial composition [51].
Despite the valuable and fresh insights provided by this study on the oral-cardiovascular axis, several limitations must be acknowledged. First, the sample size was small, as was the number of saliva and blood samples collected at each timepoint, which has limited the robustness of the analyses. This constraint is likely related to the fact that the cohort was assembled during the COVID-19 pandemic. As a result, the FFQ was administered exclusively through an online form, which significantly reduced the response rate and limited the possibility of conducting paired analyses, since not all participants responded at both timepoints. In addition, owing to pandemic-related restrictions, oral clinical examinations could not be performed in all participants, with full oral assessments obtained in only 17 of the 65 participants. Additionally, saliva samples were not collected under standardised conditions (e.g. at the same time of day), thereby not respecting the circadian variations in the oral microbiota [52]. Another limitation is the method we used to sequence the oral microbiota, which does not allow for conclusions regarding species-level or microbial functions. Finally, while NO is a highly reactive and short-lived molecule, its quantification through its stable precursors (nitrite and nitrate) provides an indirect, though generally reliable, estimation of total NO production.
Conclusions
This longitudinal study highlights a bidirectional relationship between the oral microbiota and maternal cardiovascular physiology during the transition from late pregnancy to the postpartum period. Notably, women exhibiting less favourable cardiac reverse remodelling postpartum, characterised by smaller reductions in LVEDV and greater weight retention, had a higher tendency toward microbial richness during pregnancy, along with a persistent overrepresentation of inflammation-associated genera such as Porphyromonas. This pattern suggests that a pro-inflammatory oral microbiota may be linked to delayed cardiovascular recovery. In contrast, genera such as Neisseria, which is known for nitrate-reducing activity and potential vascular benefits, and Parvimonas, which emerged in association with greater improvements in cardiac parameters, were more abundant in women with healthier cardiovascular trajectories. While no direct causal link between diet, microbiota, and cardiac remodelling could be established, our findings point to specific microbial signatures that may reflect cardiovascular adaptation across this critical period. These insights support the concept of an oral-heart axis in maternal health, open promising avenues for novel, non-pharmacological strategies to enhance cardiovascular recovery, and underscore the need for further studies to elucidate underlying mechanisms and confirm clinical relevance.
Supplementary Material
Oral_heart_axis_supplementary material.xlsxOral_heart_axis_supplementary material.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chandra M, Paray A. Natural physiological changes during pregnancy. Yale J Biol Med. 2024;97:85–92. doi: 10.59249/JTIV 413838559455 PMC 10964813 · doi ↗ · pubmed ↗
- 2Zakaria Z, Al-Rumaihi S, Al-Absi R, et al. Physiological changes and interactions between microbiome and the host during pregnancy. Front Cell Infect Microbiol. 2022;12. doi: 10.3389/fcimb.2022.824925 PMC 889966835265534 · doi ↗ · pubmed ↗
- 3Sanghavi M, Rutherford J. Cardiovascular physiology of pregnancy. Circulation. 2014;130:1003–1008. doi: 10.1161/CIRCULATIONAHA.114.00902925223771 · doi ↗ · pubmed ↗
- 4Johnson M, von Klemperer K. Cardiovascular changes in normal pregnancy. Heart disease and pregnancy. Cambridge University Press; 2016. p. 19–28. doi: 10.1017/CBO 9781316156063.005 · doi ↗
- 5Cornejo Ulloa P, Krom B, van der Veen M. Sex steroid hormones as a balancing factor in oral host microbiome interactions. Front Cell Infect Microbiol. 2021;11. doi: 10.3389/fcimb.2021.714229 PMC 851181134660339 · doi ↗ · pubmed ↗
- 6Li X, Liu Y, Yang X, et al. The oral microbiota: community composition, influencing factors, pathogenesis, and interventions. Front Microbiol. 2022;13. doi: 10.3389/fmicb.2022.895537 PMC 910067635572634 · doi ↗ · pubmed ↗
- 7Saadaoui M, Singh P, Al Khodor S. Oral microbiome and pregnancy: a bidirectional relationship. J Reprod Immunol. 2021;145:103293. doi: 10.1016/j.jri.2021.10329333676065 · doi ↗ · pubmed ↗
- 8Li Y, Zhu M, Liu Y, et al. The oral microbiota and cardiometabolic health: a comprehensive review and emerging insights. Front Immunol. 2022;13. doi: 10.3389/fimmu.2022.1010368 PMC 971628836466857 · doi ↗ · pubmed ↗