Males have a greater mite burden than females, and size does not matter: species- and sex-specific infestation patterns of mites (Uropodina) on burying beetles (Nicrophorus spp.)
Daria Bajerlein, Piotr Zduniak, Aleksandra Wyszyńska, Edward Baraniak, Marek Przewoźny, Tomasz Grzegorczyk, Arkadiusz Urbański

TL;DR
Male burying beetles carry more mites than females, and larger beetles don't necessarily carry more mites, showing complex patterns in mite dispersal.
Contribution
Reveals species- and sex-specific mite infestation patterns on burying beetles, challenging the assumption that body size influences mite burden.
Findings
Nicrophorus vespillo is most frequently infested with U. nova mites.
Male beetles have higher mite prevalence and intensity than females.
Body size does not predict mite infestation levels.
Abstract
Phoretic mites and their carriers represent a dynamic system shaped by ecological and evolutionary processes. In highly specific phoresy, which involves long-term or permanent associations, profound consequences for phoretics, including cospeciation or the transition to phoretic parasitism, can occur. Mites within the complex of cryptic species of Uroobovella nova are carried exclusively on burying beetles (Nicrophorus spp.). Nevertheless, compared with the Poecilochirus mite-Nicrophorus system, this type of interaction remains poorly understood. In this study, we investigated whether different species of burying beetles play the same role in the local dispersal of U. nova deutonymphs. To achieve this, we compared the infestation patterns of deutonymphs among field-collected beetle species, while accounting for sex and body size. Our results revealed species-specific patterns in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
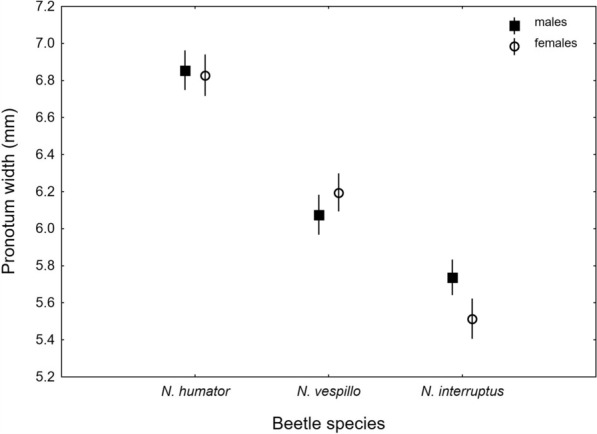 Figure 1
Figure 1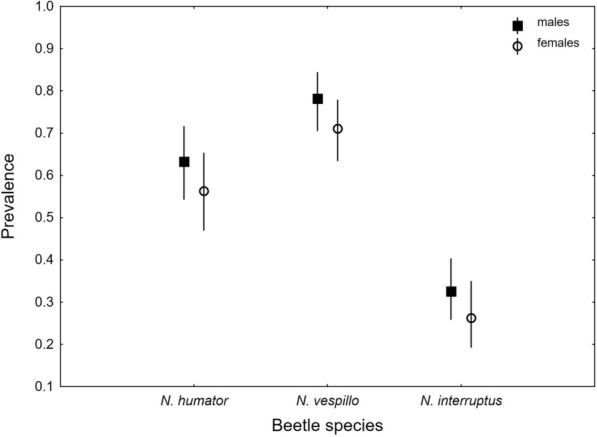 Figure 2
Figure 2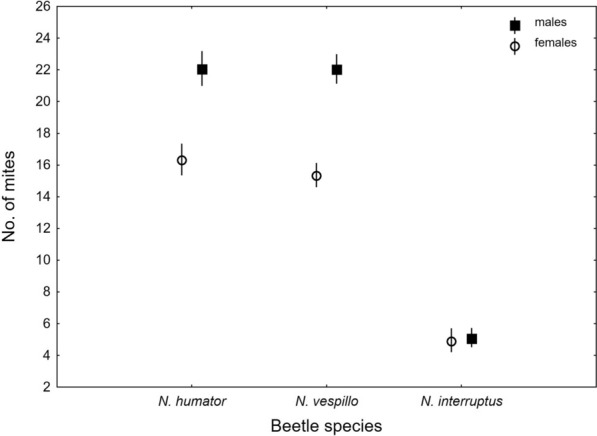 Figure 3
Figure 3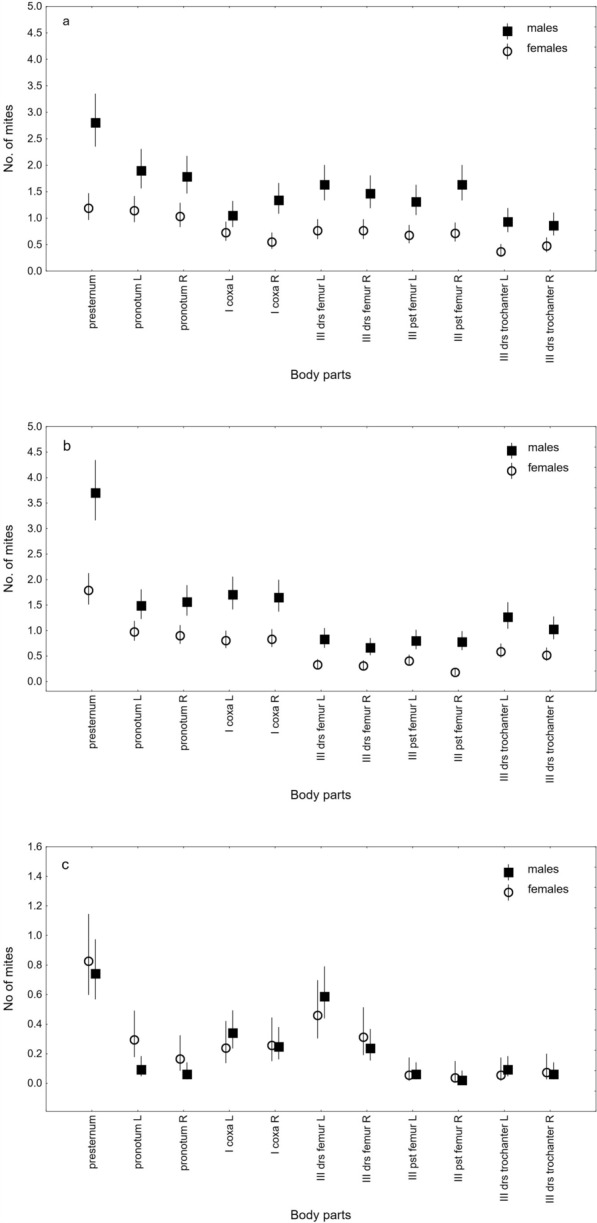 Figure 4
Figure 4 Figure 5
Figure 5- —the National Forest Holding "The State Forests", Poland
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStudy of Mite Species · Bird parasitology and diseases · Insect-Plant Interactions and Control
Background
Phoresy is a temporary symbiosis in which a smaller animal (the phoretic or phoront) attaches to a larger animal (the carrier or host) for dispersal [1]. This behaviour is common among minute, low-mobile arthropods such as mites, which exploit ephemeral and patchily distributed resources [2]. Although the primary function of phoresy is dispersal, facilitating colonization of habitats suitable for further development, it also helps avoid overcrowding and inbreeding and reduces competition [3]. Phoresy has traditionally been regarded as a commensal interaction in which the phoront benefits from being carried without harming its host. However, increasing attention has recently been devoted to the complex nature of this relationship, which may shift along a continuum from commensalism to mutualism or parasitism, depending on the relative costs and benefits experienced by both interacting partners [4].
Phoretic interactions can range from low specificity, where a phoront uses a variety of carriers, to high specificity, where the phoront is restricted to one or a few closely related carrier species [5]. Highly specific phoresy, which involves long-term or permanent associations, can have profound consequences for phoretics, including synchronization with carrier life cycles, cospeciation, or even a transition to phoretic parasitism [1, 6, 7]. A well-studied model of highly specific phoresy is the association between mites of the genus Poecilochirus Canestrini and Canestrini, 1882 (Parasitidae) and burying beetles (Nicrophorus spp.), in which both sexes cooperate in the utilization of small vertebrate carrion as a resource for feeding and breeding [e.g., 7–13]. Deutonymphs of Poecilochirus attach to adult Nicrophorus to access their brood chambers and reproduce alongside their carriers. Depending on the ecological context, these mites may either positively or negatively influence the reproductive success of their hosts [4, 10, 11, 14].
Burying beetles also interact with mites, representing a complex of cryptic species within Uroobovella nova (Parasitiformes: Uropodina) (hereafter referred to as Uroobovella nova mites) [2, 15–17]. Unlike most species of Uropodina, which utilize a broad spectrum of carriers, including representatives of several coleopteran families, U. nova deutonymphs are transported exclusively by burying beetles [18]. Like Poecilochirus carabi, U. nova was initially considered a generalist with a broad range of burying beetle hosts. However, Knee et al. [16] demonstrated that U. nova comprises at least five cryptic species associated with different Nicrophorus species. The true nature of the phoretic associations between U. nova mites and their hosts remains poorly understood.
A fundamental step in understanding any phoretic interaction is to determine the carrier range of the phoronts. This involves identifying the carriers and assessing the level of mite infestation on each carrier to distinguish primary carriers from incidental carriers. Beetles of the genus Nicrophorus are distributed worldwide and comprise approximately 65 species [19–21]. Eleven species of Nicrophorus have been reported from Europe [22], including eight from Poland [23, 24]. In a previous study [17], we presented findings on the symbiosis between U. nova mites and the common European burying beetle Nicrophorus vespilloides Herbst, 1783. Most notably, we reported a high mite prevalence of approximately 90%, with a mean infestation intensity of 24.5 deutonymphs per beetle. We also observed strong mite specificity in the selection of attachment sites, a positive but weak relationship between the number of carried deutonymphs and beetle body size, and a slight deutonymph preference for female beetles [17].
In this study, we further examined the Uroobovella nova–Nicrophorus system, focusing on three additional, less abundant burying beetle species recorded during our research: Nicrophorus vespillo (Linnaeus, 1758), Nicrophorus humator (Gleditsch, 1767), and Nicrophorus interruptus Stephens, 1830. Among European Nicrophorus beetles, N. vespilloides and N. vespillo are the most frequently reported carriers of U. nova mites [2, 15–17, 25, 26]. Deutonymphs have been recorded less commonly in other burying beetle species, such as N. humator [2, 15]. The primary objective of this study was to assess whether N. vespillo, N. humator, and N. interruptus play as important a role as carriers of U. nova mites as N. vespilloides does. To address this, we quantified phoretic deutonymph infestations by analysing both the prevalence and intensity of infestations across beetle species, as well as the potential effects of beetle sex and body size. Additionally, we examined whether the distribution patterns of mites on the studied Nicrophorus species corresponded to those previously reported for N. vespilloides.
Materials and methods
Fieldwork
Burying beetles were obtained as bycatch in 2018 and 2019 from traps originally deployed for saproxylic beetles in the Niepołomice Forest—a large woodland area located near Kraków, southern Poland (49°59′–50°07′ N, 20°13′–20°28′ E; covering approximately 110 km^2^). The trapping method used was the IBL-2 flight interception trap, which consisted of a triangular screen and a funnel attached to a container filled with ethylene glycol, which served as a preservative. Immersion of beetles in ethylene glycol prevented the movement of attached mites between hosts, thereby allowing examination of infestation patterns.
Beetles with attached mites were collected during four sampling periods: 30 May–12 July 2018, 13 July–18 September 2018, 18 May–2 July 2019, and 3 July–5 September 2019. For the analysis, we used 880 beetles, including 275 specimens of N. humator, 315 of N. interruptus, and 290 of N. vespillo, all collected between 13 July and 18 September 2018. The number of beetles collected during the other sampling periods was too low and was thus excluded from the analysis. The numbers of beetles collected during each sampling period are provided in Supplementary Table 1.
Ecology of the studied beetles
Nicrophorus humator, N. interruptus, and N. vespillo are Palearctic species distributed from Europe and North Africa eastwards to Siberia and northwestern China [27]. Among Polish burying beetles, N. vespillo and N. humator are, along with N. vespilloides, the most widespread species, whereas N. interruptus occurs more frequently in southern Poland [24, 28].
Nicrophorus vespillo is active from March to October, with activity peaks occurring either in early spring or mid-summer, depending on geographic location [22, 29–33]. This species typically reproduces up to three times per year, with the last breeding period occurring in August [31]. Nicrophorus vespillo is active from noon to midnight, with peak activity in the afternoon [31]. It prefers open habitats such as meadows, fields, wetlands, and sandy grasslands [29, 30, 34–36] but is also occasionally found at forest edges [32]. Individuals most frequently appear on carcasses around the fifth day of exposure and are attracted equally to both smaller (mice) and larger (rats) carrion [32].
Nicrophorus humator is active from April to October, with two activity peaks in April–May and August–October [29, 30, 32, 33]. It is considered a typical forest species [22, 29, 30, 32, 36], although it is also found in nonforest habitats, including field margins, forest edges, and sandy grasslands [29, 30, 32]. The species breeds up to three times per year, with beetles observed in October representing young individuals from the last generation [31]. Its activity extends from the afternoon until midnight, peaking after dusk [31]. Nicrophorus humator is most frequently recorded on small carcasses, such as those of mice [32, 37].
In Europe, N. interruptus occurs from March to October, with activity peaking in summer [22, 33, 38]. It is regarded as a primarily forest-dwelling species, but it also inhabits open and semiopen areas, including extensively used wetlands and fields [22, 39, 40]. According to Jakubec and Růžička [27], N. interruptus is a eurytopic species that occurs in both forested and open landscapes. Its diurnal activity is predominantly crepuscular [22]. The preferences of Nicrophorus interruptus for carcass size remain poorly understood.
Analysis of mite-beetle interactions
All collected beetle specimens were examined for phoretic deutonymphs of U. nova mites and their pedicels. The pedicel is a temporary, stalk-like structure with two extended termini [41–43]. One terminus attaches to the deutonymph’s anal region, whereas the other attaches to the carrier’s body surface. After deutonymph detachment, the pedicel typically remains on the carrier as evidence of the mite’s prior presence.
For each beetle, the number of deutonymphs and pedicels without deutonymphs, as well as their locations on the host’s body, were recorded. Phoretic deutonymphs were identified via the Uropodina identification key by Karg [44]. Burying beetles were identified according to keys for the identification of burying beetles by Mroczkowski [45], Jałoszyński [46] and UK Beetle recording website [47] and sexed by genital dissection [48]. Beetle body size was determined by measuring pronotum width, defined as the distance between the two most lateral points, following Jarrett et al. [49].
Data analysis
The differences in body size between beetles were calculated with the Factorial ANOVA, where SPECIES, SEX, and SPECIESSEX were the factors. Furthermore, we analysed the influence of beetles’ SPECIES, SEX, and SIZE as well as the interaction effect of SPECIESSEX on deutonymph prevalence (the proportion of infested beetles) via a Generalized Linear Model (GLM) with a binomial distribution and logit link function, where prevalence (binary variable) was the dependent variable and SPECIES (categorical variable), SEX (binary variable), and SIZE (continuous variable) were the factors. Moreover, we analysed the impacts of beetles’ SPECIES, SEX and SIZE, as well as the interaction effects of SPECIESSEX on the intensity of deutonymph infestation, e.g., the mean number of deutonymphs per infested beetle, via a GLM with a Poisson distribution and a log link function, where the intensity of mite infestation was the dependent variable and SPECIES, SEX, SIZE, and SPECIESSEX were the factors. Finally, we examined the distribution of mites on beetles, considering the most infested parts of their bodies [PART], taking into account their left or right side (a total of eleven body parts), SPECIES, SEX, and body SIZE via a Generalized Linear Mixed Model (GLMM) with a Poisson distribution and log link function, where the number of mites was the dependent variable; SPECIES, SEX, PART, body SIZE, and SPECIESSEXPART were fixed factors; and the beetle ID was a random factor. All calculations were performed via IBM SPSS Statistics for Windows [50]. Throughout the text, the mean values are presented with 95% confidence limits (CLs).
Significant differences in body size were determined based on pronotum width among the collected SPECIES. Nicrophorus humator was the largest, N. interruptus smallest, and N. vespillo of intermediate size (Fig. 1, Supplementary Table 2). The effect of sex was insignificant, but we found a significant interaction of SPECIES and SEX, with larger males in N. interruptus, and no intersexual differences in the other species (Fig. 1, Supplementary Table 2).Fig. 1. Body size expressed as pronotum width in Nicrophorus humator, N. vespillo, and N. interruptus, including their sex; mean values are presented with 95% confidence limits
Results
Deutonymph prevalence
The infestation status of the beetles varied between SPECIES, with N. vespillo being the most frequently infested and N. interruptus the least frequently infested (Tables 1, 2, Fig. 2). The effect of SEX was also significant, with a relatively high deutonymph prevalence reported in males (Tables 1, 2, Fig. 2). The deutonymph prevalence did not depend on beetles’ SIZE, and the interaction effect of SPECIES and SEX was also insignificant (Tables 1, 2, Fig. 2). Table 1. Summary of Generalized Linear Model (GLM) analysis of factors influencing the prevalence of deutonymphs from the Uroobovella nova complex of cryptic species on Nicrophorus humator,* N*. vespillo, and* N*.* interruptusFactorWald χ^2^dfpINTERCEPT0.59510.441SPECIES101.8882< 0.001SEX4.76510.029SPECIESSEX0.05820.971BODY SIZE0.24210.623Table 2Prevalence and intensity of infestation of deutonymphs within the Uroobovella nova complex of cryptic species of mites phoretic on burying beetlesBeetle speciesNKP (%)95% CLI ± SDMin–MaxNicrophorus humator**(Gleditsch)27516359.2753.21–65.1319.43 ± 27.531–147Females1337556.3947.53–64.9715.96 ± 26.311–138Males1428861.9753.45–69.9822.39 ± 28.341–147Nicrophorus vespillo** (Linnaeus)29021674.4869.06–79.4018.58 ± 27.741–192Females15210871.0563.15–78.1115.28 ± 18.951–105Males13810878.2670.44–84.8321.88 ± 34.141–192 Nicrophorus interruptus** Stephens3159630.4825.44–35.884.88 ± 5.261–28Females1383726.8119.63–35.014.78 ± 5.191–28Males1775933.3326.44–40.804.93 ± 5.351–26N, The number of collected beetles; K, The number of infested beetles; P, Prevalence; I, The intensity of mite infestation; CL, Confidence limits; SD, Standard deviation; Min–Max, The minimal and maximal number of carried mitesFig. 2Prevalence of deutonymphs within the Uroobovella nova complex of cryptic species carried by Nicrophorus humator, N. vespillo, and N. interruptus in relation to sex; mean values are presented with 95% confidence limits
Intensity of deutonymph infestation
The intensity of deutonymph infestation differed significantly among SPECIES, being the highest on N. vespillo and N. humator and the lowest on N. interruptus (Tables 2, 3, Fig. 3). The number of mites was positively related to beetles’ SEX, with males carrying a higher mite load (Tables 2, 3, Fig. 3). The interaction effect between SPECIES and SEX was also significant; males of N. vespillo and N. humator carried more mites, whereas no sex-related differences were detected in N. interruptus (Tables 2, 3, Fig. 3). The SIZE of the beetle did not affect the intensity of the deutonymph infestation (Tables 2, 3). Table 3. Summary of the Generalized Linear Model (GLM) analysis of factors influencing the intensity of infestation of deutonymphs from the Uroobovella nova complex of cryptic species on Nicrophorus humator, N. vespillo, and* N. interruptusFactorWald χ^2^dfpINTERCEPT424.4401< 0.001SPECIES677.2532< 0.001SEX42.8791< 0.001SPECIESSEX10.71320.005BODY SIZE2.35610.125Fig. 3Intensity of infestation of deutonymphs within the Uroobovella nova complex of cryptic species carried by Nicrophorus humator, N. vespillo, and N. interruptus in relation to sex; mean values are presented with 95% confidence limits
Localization of deutonymphs on the beetles’ bodies
The analysis of the most heavily infested beetles’ body PARTS (n = 11), considering both the left and right sides of the body as well as SPECIES, SEX, body PARTS, body SIZE, and the interaction effect of SPECIESSEXPARTS, revealed differences in deutonymph location on their carriers (Table 4, Fig. 4). Table 4. Summary of the Generalized Linear Mixed Model (GLMM) analysis of factors influencing the attachment patterns of deutonymphs from the Uroobovella nova complex of cryptic species on Nicrophorus humator, N. vespillo, and* N. interruptusFactorFdf1df2pSPECIES215.79325114< 0.001SEX47.71915114< 0.001BODY PARTS64.425105114< 0.001BODY SIZE2.034151140.154SPECIESSEX*BODY PARTS9.922525114< 0.001MODEL36.170665114< 0.001Fig. 4Mean numbers of deutonymphs per infested beetle within the Uroobovella nova complex of cryptic species attached to body parts of Nicrophorus humator (a), N. vespillo (b), and N. interruptus (c). Legend: L, R–the left and right body sides, presternum–the prothorax presternum, I–forelegs, III–hindlegs, pronotum–ventral parts of pronotum lateral margins, coxa–the ventral side of the coxae, drs–the dorsal side, pst–the posterior side; the means are presented with 95% confidence limits
The effect of SPECIES was significant, with greater numbers of deutonymphs recorded on particular body parts in N. humator and N. vespillo than in N. interruptus (Table 4, Fig. 4). In N. humator, the distribution of mites was relatively uniform, with smaller differences between the numbers of deutonymphs attached to the presternum, the lateral sides of the ventral pronotum, and the coxae of the forelegs. In contrast, in N. vespillo, the difference between the intensity of mite infestations on the presternum and other body parts was more pronounced. In N. interruptus, deutonymphs, in addition to their distinct preference for the presternum, also attach particularly frequently to the left femora of hindlegs (Table 4, Figs. 4, 5). SEX significantly affected mite distribution, with males exhibiting more heavily infested body parts than females. The most pronounced intersexual differences in the intensity of deutonymph infestation were observed within the presternum (Table 4, Figs. 4, 5). The effect of body PARTS was significant, with more deutonymphs attached to anterior regions such as the presternum, lateral sides of the ventral pronotum, and coxae of the forelegs than to posterior body parts, particularly the femora and trochanters of the hindlegs. Deutonymphs were evenly distributed between the left and right sides of the host body, except for the femora of the hindlegs in N. interruptus (Table 4, Fig. 4). The interaction effect of SPECIES, SEX, and PARTS was also significant, with males of N. humator and N. vespillo carrying more deutonymphs on the anterior body regions of the hosts. In N. interruptus, differences in mite distribution were less consistent and similar between the sexes. Moreover, in females of N. humator and N. vespillo, deutonymphs were distributed more evenly across the examined body regions. In contrast, in males, a more distinct preference for the presternum was observed (Table 4, Figs. 4, 5). The effect of body SIZE on mite localization was not significant (Table 4).Fig. 5. Burying beetles with attached deutonymphs and pedicels of the Uroobovella nova complex of cryptic species. Legend: a Localization of mites on the ventral side of Nicrophorus vespillo; b Localization of mites on the ventral side of N. humator; c Mites attached to the prothorax presternum and foreleg coxae of N. vespillo; d Mites attached to the prothorax presternum and foreleg coxae of N. interruptus; e Mites attached to the prothorax presternum, pronotum lateral margins, and foreleg coxae of N. humator; f, g Pedicels and deutonymphs attached to the dorsal femora and trochanters of hindlegs in N. vespillo (f) and N. interruptus (g). I–prothorax presternum; II–pronotum lateral margin, III–foreleg coxa, IV–posterior part of the femur of the hindleg, V–dorsal part of the femur of the hindleg, VI–dorsal part of the trochanter of the hindleg. Scale bars: 2 mm (a, d, e); 5 mm (b); 1 mm (c, f, g)
Discussion
In this work, we present novel and significant findings on the highly specific, phoretic interactions between U. nova mites and burying beetles (Nicrophorus spp.). Nicrophorus vespilloides—previously investigated by us [17]—exhibits the highest level of infestation, followed by N. vespillo and N. humator. In contrast, N. interruptus has the lowest infestation rate and has not yet been reported as a carrier of U. nova mites.
There may be several reasons for the variation in U. nova mite infestations among the studied beetles. First, the question arises whether all burying beetles studied carry the same mite species within the analysed mite complex. Knee et al. [16] reported that in Europe, N. vespilloides and N. vespillo carry Uroobovella sp. 3. In turn, Athias-Binche et al. [2] reported that deutonymphs collected from N. vespillo and N. vespilloides are smaller than those collected from N. humator, which may indicate that they are separate morphotypes. Research conducted by Kočárek [31] demonstrated that Horn’s index, which measures niche overlap, remains consistently high for N. vespilloides and N. vespillo throughout their entire seasonal activity. Owing to differences in diurnal activity and habitat preference, Horn’s index is very low in spring between N. vespillo and N. humator as well as between N. vespilloides and N. humator but increases significantly during summer and autumn. A high degree of niche overlap between N. vespilloides and N. vespillo may facilitate possible host switching in U. nova mites, in contrast to the lower niche overlap observed between these two species and N. humator. Different Nicrophorus species may compete for the same carcass [51, 52], creating opportunities for interspecific switching between hosts in phoretic deutonymphs. These events could promote gene flow among mite populations and hinder speciation. However, host switching on carcasses seems unlikely, as, according to previous observations [41], deutonymphs of Uropodina detach only in suitable microhabitats, where their development occurs. Attachment via a pedicel further limits movement between hosts. Thus, host switching probably occurs mainly when a breeding chamber is used by another Nicrophorus species, allowing deutonymphs of the next generation to disperse with the new host’s offspring. Most likely, the deutonymphs examined in our study represent a single species, Uroobovella sp. 3, distinguished by Knee et al. [16], with the possible occurrence of host races associated with N. humator and perhaps N. interruptus. Therefore, it is likely that the observed variation in deutonymph infestation among the examined beetles results not from their different representative species within the studied complex but rather from interspecific differences in the biology of their carriers, particularly in their breeding behaviour. Unfortunately, these behaviours have been poorly studied in N. humator and N. interruptus, making it difficult to draw any firm conclusions at this stage. The susceptibility of the carrier to infestation by Uropodina mites may also be shaped by other host-dependent factors, such as seasonal activity and population dynamics, duration of exposure to mites, morphology of the host body surface, and host behaviors related to grooming [for a review, see 18]. Infestation is also influenced by mite-dependent factors, such as their seasonal activity, population dynamics, body size, and method of attachment [for a review, see 18].
In the studied Uroobovella–Nicrophorus system, carrier body size does not appear to be a factor driving mite prevalence or load. Although our previous study demonstrated that the body size of N. vespilloides positively affects the intensity of deutonymph infestation, the effect was weak and only marginally significant. Nicrophorus vespilloides, despite being the smallest among the studied burying beetle species (mean pronotum width of 4.92 mm [17]), was more heavily infested than the much larger N. humator examined in this study. In this context, our results align with previous findings suggesting that body size does not play a consistently significant role in determining host suitability for symbionts [e.g., 53–55].
The effect of beetle sex on prevalence was significant, with males being more frequently selected as carriers than females. In N. vespillo and N. humator, males carried a higher mite load, whereas in N. interruptus, no effect of sex on the number of carried deutonymphs was observed. For N. vespilloides, we recorded a slightly greater mite load in females (8.1% more mites than in males) [17]. These sex-specific infestation patterns are likely related to differences in breeding biology between sexes across the studied species. Cooperation between males and females in the preparation of brood chambers and in parental care has been well documented in N. vespilloides [56, 57] and N. vespillo [58] but not in the other Nicrophorus species examined. Therefore, further research is needed to explore the breeding behaviour of burying beetles in more detail.
Although the general pattern of U. nova mite distribution was similar among the Nicrophorus species examined, with more individuals occurring in the anterior body regions, particularly the prothorax presternum, regardless of host body size (consistent with our previous findings for N. vespilloides), our results revealed species- and sex-specific differences in the intensity of deutonymph infestation across the studied body parts. In N. humator, deutonymphs were distributed more evenly across all body parts. In contrast, in N. vespillo, differences observed between the number of mites attached to the presternum and those on other body parts were more distinct. In N. interruptus, the overall number of deutonymphs was very low and evenly distributed across the body, except for the dorsal side of the femora of the left hindlegs. In N. vespillo and N. humator, pronounced intersexual differences in deutonymph distribution were recorded, but these differences were not detected in N. interruptus. In the previously studied N. vespilloides, no effect of sex on the distribution of mites was found, and the coxae of the first pair of legs were, after the presternum, the second most heavily colonized body part [17]. The observed variation in deutonymph distribution patterns most likely results from interspecific and intersexual differences in host biology, which remain poorly understood for the species studied here. The spatial distribution of Uropodina mites on the host likely affects their dispersal success, with mites possibly attaching to areas that lower the risk of detachment and do not hinder host movement [e.g., 59–61].
Our study provides new and intriguing insights into the highly specific symbiosis between U. nova and burying beetles, complementing previous research on this interaction. Future studies should address local genetic differentiation within the U. nova species complex to clarify its taxonomic status. In addition, laboratory experiments are needed to observe the biology of U. nova mites within the breeding chambers of burying beetles, their impact on their hosts, and their interactions with Poecilochirus mites, thereby contributing to a better understanding of how phoretic symbiosis is shaped in mites coevolving with their hosts in carrion-associated systems.
Conclusions
We showed that at the local scale, U. nova mites use different burying beetle species for dispersal and may exhibit host-specific infestation patterns. These findings suggest that individual burying beetle species may play different roles in the dispersal of deutonymphs. The previously studied N. vespilloides is the primary carrier of U. nova deutonymphs, followed by N. vespillo and N. humator analysed in the present study, whereas N. interruptus plays a considerably less significant role. We also observed sex-specific infestation patterns, which appeared more consistent in terms of prevalence but more variable in terms of the intensity of mite infestation. Interestingly, variation in host body size was not a significant predictor of Uroobovella nova infestation. This finding demonstrates that the simple correlation “larger hosts carry more symbionts” does not hold universally in ecology. Although more mites are attached to the anterior than to the posterior parts of the beetle body in all the examined species, species- and sex-specific patterns in the distribution of deutonymphs were evident. Multiple factors likely influence differences in infestations among burying beetle species; in our view, beetle species- and sex-specific breeding behaviours may play a key role in shaping the infestation patterns of U. nova mites.
Supplementary Information
Additional file1 (DOCX 24 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1https://www.gbif.org
- 2Jałoszyński P. https://entomo.pl/artykuly/abcjaloszynski/index.php
- 3https://coleoptera.org.uk