Inositol phosphates: small molecules with a BIG impact on maize embryo development
Catherine P Freed, Erin Cullen

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
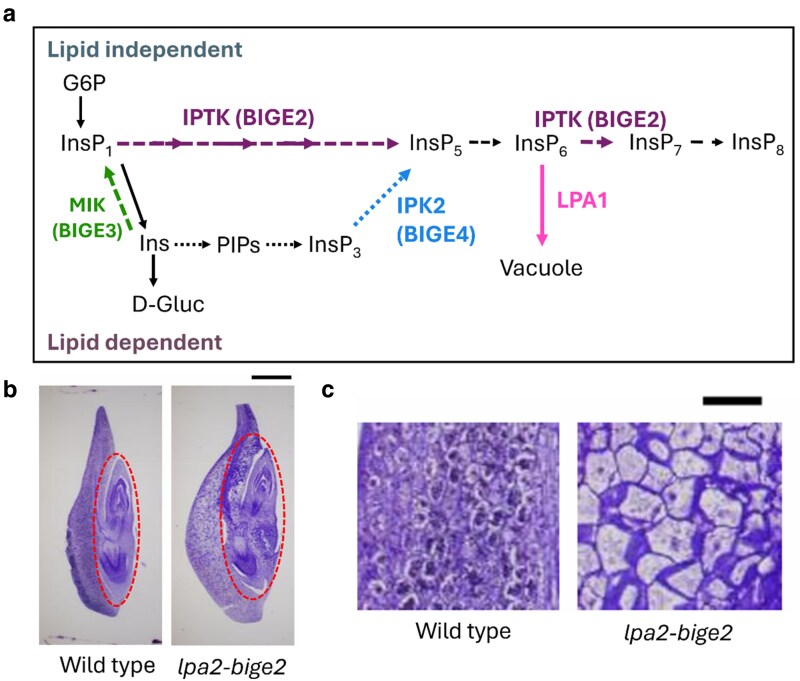 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytase and its Applications · Protein Kinase Regulation and GTPase Signaling · Cellular transport and secretion
Plants utilize complex signaling networks to coordinate nutrient allocation and storage across tissues. In maize and other cereal crops, traits such as embryo and endosperm size are key determinants of oil, protein, starch, and nutrient storage. Understanding the molecular foundations of embryo size and nutrient storage is essential for developing nutrient rich crops.
Inositol phosphates (InsPs) are a diverse group of signaling molecules that, depending on their phosphorylation pattern, can convey unique chemical information (Gillaspy 2011). InsPs are important for plant development, phosphate sensing, hormone signaling, and response to abiotic and biotic stress (Ghosh et al. 2025). Consisting of a 6-carbon inositol ring scaffold, InsPs are phosphorylated by a specific set of enzymes in 2 interconnected pathways that are lipid dependent and lipid independent (Williams et al. 2015; Shears 2018). Although many key enzymes involved in InsP signaling have been identified in plants, there are still many mechanistic unknowns surrounding how specific InsPs contribute to plant development and stress responses. Understanding how InsPs impact specific developmental processes is critical given they are attractive targets for enhancing nutrient use efficiency and stress resilience in crops (Freed et al. 2020),
A recent study in Plant Physiology by Suzuki et al. (2026) investigates the link between InsP signaling and maize embryo development. To determine the genetic controllers of maize embryo size, the authors identified 4 maize mutants with enlarged embryos from a previous screen (McCarty et al. 2005). Interestingly, all 4 identified mutants had mutations in the InsP signaling pathway: big embryo2 (bige2) and big embryo3 (bige3), deficient in lipid independent InsP kinases; big embryo4 (bige4), impaired in a lipid dependent InsP kinase; and low phytic acid 1 (lpa1), lacking a tonoplast InsP_6_ transporter (Fig. 1). Including mutants involved in different aspects of InsP signaling allowed the authors to study how InsP enzymes spanning multiple parts of the signaling pathway contribute to embryo development.
All mutants displayed alterations in scutellum growth based on changes in cell size and/or number at the expense of the endosperm (Fig. 1). Notably, bige mutants showed higher accumulation of anthocyanins in certain areas of the kernels as well as distinct changes in germination frequency compared to wild-type (WT) kernels. Of these, bige2 mutants showed the most distinctive phenotypes in size and coloration compared to the other mutants. Together, this suggests that while these genes have overlapping roles in altering embryo size, they also have distinct roles in maize development.
To dissect changes in increased embryo size, the authors used histological analyses to quantify cell size in bige2 and bige4 embryos at later stages in kernel development. Although both mutants had an enlarged scutellum, the bige2 scutellum was larger due to increased cell size, whereas the bige4 scutellum had an increased number of cells. The authors found that the increase in cell size may be due to an increase in ploidy. Upon further inspection of the embryos at apical, middle, and basal locations, the authors found that bige2 had significantly larger cells in the apical region, while in bige4 cell size was not largely altered at any location. Together, these results establish that alterations in specific InsP signaling genes impact cell size and number in specific regions in the embryo.
The authors also extended their histological analysis to query scutellum size in all 4 mutants at the onset of embryo development. Both bige2 and lpa1 scutellum were significantly larger in the apical region and bige3 scutellum cells were slightly larger compared to WT. In bige4, some areas of the scutellum had larger cells compared to WT, whereas in the apical region, while the cells were not significantly larger in size, they were higher in number. Together, these results establish that alterations in specific InsP signaling genes impact cell size and number in specific regions in the embryo.
Suzuki et al. (2026) also compared the transcriptomes of WT and bige embryos. Of these, bige2 mutant embryos showed the most changes with ∼4,600 differentially expressed genes (DEGs), consistent with their stronger kernel phenotypes, while bige3 mutants had under 100 DEGs and bige4 mutants had ∼2,100 DEGs. These 2 mutants shared lipid accumulation and histone DEGs, suggesting that these genes are uniquely regulated by the lipid independent InsP kinases. A small subset of 13 genes was shared between all 3 bige mutants with roles in RNA biogenesis and ion homeostasis. Interestingly, over one-half of the upregulated DEGs were shared between bige2 and bige4 embryos with roles in protein and ribosomal translation, phosphate and lipid metabolism, DNA replication and repair, and the cell cycle. These data suggests that the InsP kinases from both pathway impact cell cycle regulation in mutant embryos.
The authors also quantified levels of genes previously identified to regulate embryo size in maize, such as CYP78A and BIGE1-LIKE MATE transporter genes. Multiple CYP78A genes were downregulated in bige2 and bige4 mutants, whereas BIGE1-LIKE MATE genes were upregulated in bige2, consistent with past literature (Suzuki et al. 2015). Mutant embryo transcriptome analyses also revealed anthocyanin synthesis genes in the bige mutants that were consistent with the observed kernel phenotypes.
This work uniquely establishes a role for InsPs in regulating embryo size and cell development in maize. Knocking out genes that span multiple parts of the InsP signaling pathway highlights both overlapping and distinct roles for InsP enzymes in regulating embryo development and the cell cycle in maize. This research is valuable given ongoing efforts that target InsPs to enhance crop resilience and nutrient use (Raboy 2007; Freed et al. 2020). Given the importance of InsPs in phosphate sensing and hormone signaling pathways such as auxin and jasmonic acid, future efforts exploring bige and lpa1 maize mutant defense response to abiotic and biotic stress would be informative. Future studies should also quantify and examine how specific InsP levels contribute to bige and lpa1 maize phenotypes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Freed C, Adepoju O, Gillaspy G. 2020. Can inositol pyrophosphates inform strategies for developing low phytate crops? Plants. 9:115. 10.3390/plants 9010115.31963418 PMC 7020182 · doi ↗ · pubmed ↗
- 2Ghosh R, Yadav R, Pullagurla NJ, Rana P, Laha D. 2025. The expanding landscape of inositol phosphate signaling network in land plants. FEBS Lett. 10.1002/1873-3468.70236. Online ahead of print 41288081 · doi ↗ · pubmed ↗
- 3Gillaspy GE . 2011. The cellular language of myo-inositol signaling: Tansley review. New Phytol. 192:823–839. 10.1111/j.1469-8137.2011.03939.x.22050576 · doi ↗ · pubmed ↗
- 4Mc Carty DR et al 2005. Steady-state transposon mutagenesis in inbred maize. Plant J. 44:52–61. 10.1111/j.1365-313X.2005.02509.x.16167895 · doi ↗ · pubmed ↗
- 5Raboy V . 2007. The AB Cs of low-phytate crops. Nat Biotechnol. 25:874–875. 10.1038/nbt 0807-874.17687363 · doi ↗ · pubmed ↗
- 6Shears SB . 2018. Intimate connections: inositol pyrophosphates at the interface of metabolic regulation and cell signaling. J Cell Physiol. 233:1897–1912. 10.1002/jcp.26017.28542902 PMC 5694711 · doi ↗ · pubmed ↗
- 7Suzuki M, Sato Y, Wu S, Kang BH, Mc Carty DR. 2015. Conserved functions of the MATE transporter BIG EMBRYO 1 in regulation of lateral organ size and initiation rate. Plant Cell. 27:2288–2300. 10.1105/tpc.15.00290.26276834 PMC 4568504 · doi ↗ · pubmed ↗
- 8Suzuki M, Wu S, Sato Y, Jones NT, Mc Carty DR. 2026. Control of embryo size by inositol phosphate signaling revealed by big embryo mutants of maize. Plant Phys. 20:1–17. 10.1093/plphys/kiag 005. Online ahead of print 41554099 · doi ↗ · pubmed ↗