More than a defense: salicylic acid's secret role in fruit color
Ved Prakash

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
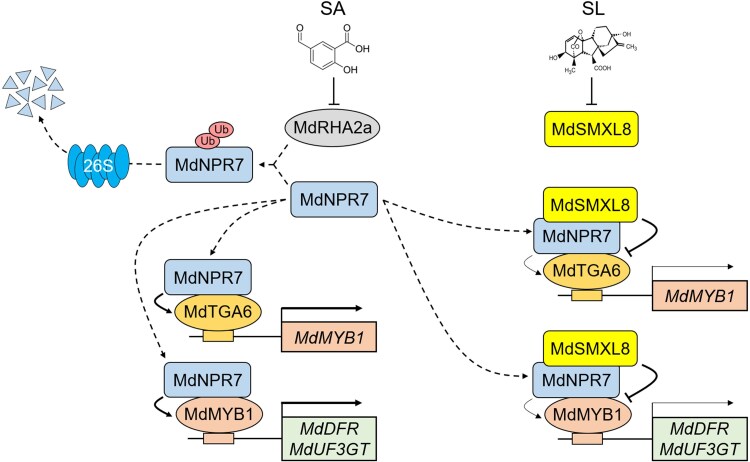 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Postharvest Quality and Shelf Life Management · Horticultural and Viticultural Research
The vibrant red skin of an apple (Malus domestica) is not only a visual delight for consumers but also an important biological trait. The variation in the color of different apple varieties comes from varying proportions of a flavonoid pigment, anthocyanin, in the apple fruit skin. Anthocyanin not only provides coloration but also serves to attract seed dispersers and protect the fruit from the UV radiation and fluctuating temperatures (Steyn et al. 2002; Shang et al. 2011; Zhao et al. 2023). Although the role of ripening hormones such as ethylene and abscisic acid is well documented in promoting pigmentation in fruits, the influence of defense-related hormones in fruit pigmentation has remained less explored.
In a recent study, Lei Zhao and colleagues (Zhao et al. 2026) examined how salicylic acid (SA) induces anthocyanin biosynthesis (see Fig. 1). Besides the well-established role of SA in plant immunity, the authors found that SA treatment can induce anthocyanin accumulation in apple. This induction was mediated by increased accumulation of MdMYB1 transcripts, a master transcription factor that increased in 30 min and peaked 1 hour after SA treatment. Hampering the MdMYB1 expression using tobacco rattle virus–based silencing impaired SA-induced anthocyanin accumulation in the fruit. The authors observed similar results through antisense suppression of MdMYB1 expression in apple callus, suggesting that MdMYB1 is essential for SA-induced anthocyanin biosynthesis.
The authors further identified another SA-responsive transcription factor, MdTGA6, which binds to the promoter of the MdMYB1 gene to activate its transcription. TGA transcription factors belong to the basic region leucine zipper (bZIP) protein family. Using chromatin immunoprecipitation-PCR and electrophoretic mobility shift assays, the authors confirmed the interaction of MdTGA6 with the MdMYB1 promoter; as well, using GUS reporter and luciferase assays, they confirmed that MdTGA6 activates MdMYB1. Importantly, this process requires a copilot, MdNPR7, an apple ortholog of the well-known SA receptor and transcriptional coactivator NON-EXPRESSOR OF PATHOGENESIS-RELATED GENES 1 (NPR1) (Chen et al. 2021). MdNPR7 physically interacts with MdTGA6, as confirmed by 4 independent methods: yeast 2-hybrid assay, pull-down assay, chromatin immunoprecipitation, and bimolecular fluorescence complementation. The interaction between MdNPR7 and MdTGA6 significantly enhanced the transcription of MdMYB1. Additionallly, when the authors silenced MdNPR7, they found a visible decrease in anthocyanin biosynthesis in apple fruit as well as in callus, confirming the role of MdNPR7 in promoting anthocyanin synthesis. MdNPR7 was shown to have another crucial function as it interacted directly with the MdMYB1 protein. This physical interaction of MdNPR7 with MdMYB1 enhanced MdMYB1's affinity for the promoters of downstream anthocyanin biosynthesis genes, effectively promoting the anthocyanin-production machinery in apple.
Furthermore, in the absence of SA, the RING-type E3 ubiquitin ligase MdRHA2a targets MdNPR7 for degradation by the 26S proteasome. This constant turnover of MdNPR7 ensures that anthocyanin production remains low under normal conditions. However, when SA is present, it interferes with the MdNPR7 degradation process, allowing the MdNPR7-TGA6-MYB1 module to accumulate and initiate anthocyanin biosynthesis.
Another important finding of the study is integration of SA with strigolactone (SL) hormones in that anthocyanins accumulated more strongly in fruit during co-treatment with SA and SL compared to individual treatments. When SL signaling is activated, a repressor protein, MdSMXL8, is degraded, which allows MdNPR7 to promote anthocyanin production. Thus, MdNPR7 is a central component of the SA-mediated anthocyanin biosynthesis that integrates defensive signals, growth regulators, and metabolic outputs. The findings from this research broaden our understanding of SA from being a defense hormone to also being a versatile regulator of fruit color.
This research identified several targets for molecular apple breeding. By interfering with the stability of MdNPR7 or its interaction with MdSMXL8, breeders may develop apple varieties that have increased coloration and better resilience. This is particularly crucial under climate change because increased temperature can affect pigmentation. Furthermore, the increase in SA-mediated defense may extend fruit life by improved resistance to pathogens. This study not only identifies an intricate molecular mechanism of SA-mediated anthocyanin biosynthesis in apple but also presents a new case for the multi-hormonal control of plant secondary metabolism.
Recent related articles in The Plant Cell:
An et al. (2024) identified that the SL response factor AGL9 and SL signaling repressor SMXL8 mediate the crosstalk between SL and gibberellin, and this crosstalk regulates anthocyanin biosynthesis in apple. Q. Sun et al. (2025) explored ethylene-mediated citrus peel reddening. The authors found that the transcription factor CsERF25 binds to the promoter of the CsRP1 gene, thus enhancing its transcription. CsRP1 protein then binds to the CsCCD4b promoter, activating its transcription and thus leading to reddening of citrus peel.Y. Sun et al. (2025) showed that the auxin and endoplasmic reticulum stress pathways determine the inhibition of anthocyanin biosynthesis in grape during high temperature.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1An J-P et al 2024. The SMXL 8-AGL 9 module mediates crosstalk between strigolactone and gibberellin to regulate strigolactone-induced anthocyanin biosynthesis in apple. Plant Cell. 36:4404–4425. 10.1093/plcell/koae 191.38917246 PMC 11448916 · doi ↗ · pubmed ↗
- 2Chen J et al 2021. More stories to tell: NONEXPRESSOR OF PATHOGENESIS-RELATED GENES 1, a salicylic acid receptor. Plant Cell Environ. 44:1716–1727. 10.1111/pce.14003.33495996 · doi ↗ · pubmed ↗
- 3Shang Y et al 2011. The molecular basis for venation patterning of pigmentation and its effect on pollinator attraction in flowers of antirrhinum. New Phytol. 189:602–615. 10.1111/j.1469-8137.2010.03498.x.21039563 · doi ↗ · pubmed ↗
- 4Steyn WJ, Wand SJE, Holcroft DM, Jacobs G. 2002. Anthocyanins in vegetative tissues: a proposed unified function in photoprotection. New Phytol. 155:349–361. 10.1046/j.1469-8137.2002.00482.x.33873306 · doi ↗ · pubmed ↗
- 5Sun Q et al 2025. Red peel regulator 1 links ethylene response factor 25 and β-citraurin biosynthetic genes to regulate ethylene-induced peel reddening in citrus. Plant Cell. 37:koaf 010. 10.1093/plcell/koaf 010.PMC 1176093939792899 · doi ↗ · pubmed ↗
- 6Sun Y et al 2025. Vv FHY 3 links auxin and endoplasmic reticulum stress to regulate grape anthocyanin biosynthesis at high temperatures. Plant Cell. 37:koae 303. 10.1093/plcell/koae 303.PMC 1166357239539042 · doi ↗ · pubmed ↗
- 7Zhao L et al 2026. The NPR 7-TGA 6-MYB 1 module promotes salicylic acid-induced anthocyanin biosynthesis and integrates salicylic acid with strigolactone signaling via interaction with SMXL 8 in apple. Plant Cell. 38:koag 030. 10.1093/plcell/koag 030.41712403 · doi ↗ · pubmed ↗
- 8Zhao Y et al 2023. Colorful hues: insight into the mechanisms of anthocyanin pigmentation in fruit. Plant Physiol. 192:1718–1732. 10.1093/plphys/kiad 160.36913247 PMC 10315290 · doi ↗ · pubmed ↗