Genomic epidemiology of extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in Mwanza, Tanzania
Vitus Silago, Benson R. Kidenya, Katarina Oravcova, Louise Matthews, Conjester I. Mtemisika, Stephen E. Mshana, Heike Claus, Jeremiah Seni

TL;DR
This study identifies high-risk antibiotic-resistant bacteria in a hospital in Tanzania, highlighting the spread of dangerous clones and the need for better infection control.
Contribution
First documentation of ESBL-producing Klebsiella pneumoniae ST2390 in neonatology units and genomic analysis of ESBL clones in Mwanza, Tanzania.
Findings
ESBL-producing Escherichia coli ST131 is the predominant high-risk clone in Mwanza hospitals.
ESBL-producing Klebsiella pneumoniae ST2390 is newly identified and associated with neonatal bloodstream infections.
Genomic analysis reveals clonal clusters of ESBL bacteria in medical and neonatology wards.
Abstract
•Predominance of globally recognised high-risk clone extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli ST131.•First documentation and predominance of ESBL-producing Klebsiella pneumoniae ST2390 associated with bloodstream infections in the neonatology units.•The cgMLST-based Neighbour-Joining (NJ) phylogenetic analysis revealed clonal clusters involving high-risk clone ESBL-EC ST131 in medical and neonatology wards, and newly detected ESBL-KP ST2390 in the neonatology ward/unit. Predominance of globally recognised high-risk clone extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli ST131. First documentation and predominance of ESBL-producing Klebsiella pneumoniae ST2390 associated with bloodstream infections in the neonatology units. The cgMLST-based Neighbour-Joining (NJ) phylogenetic analysis revealed clonal clusters involving high-risk clone ESBL-EC…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
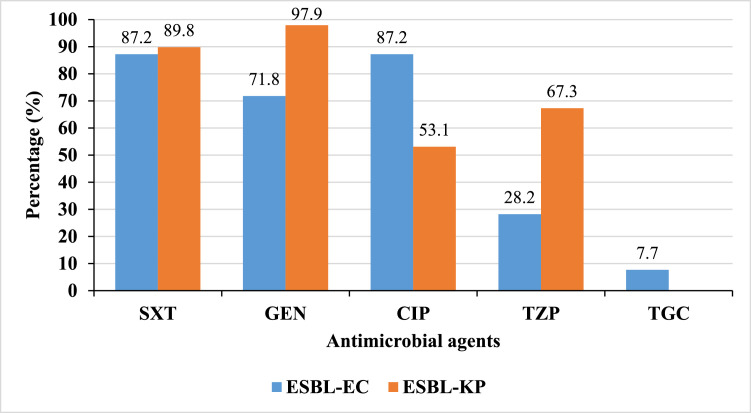 Figure 1
Figure 1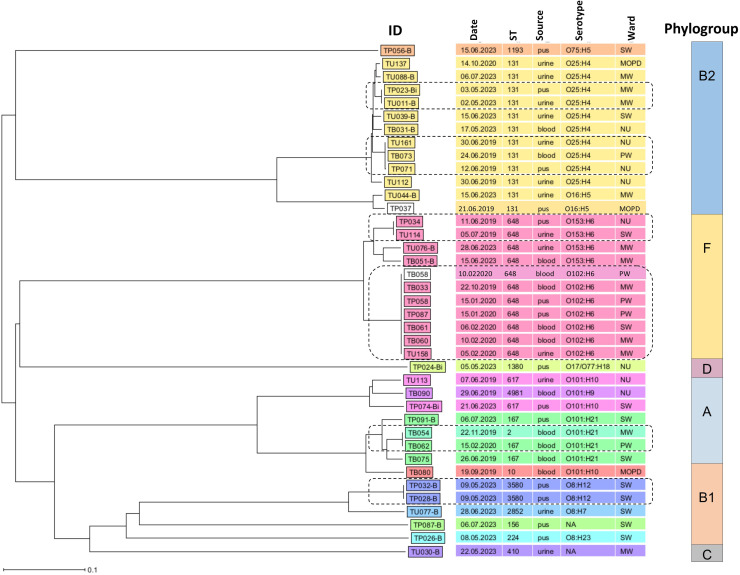 Figure 2
Figure 2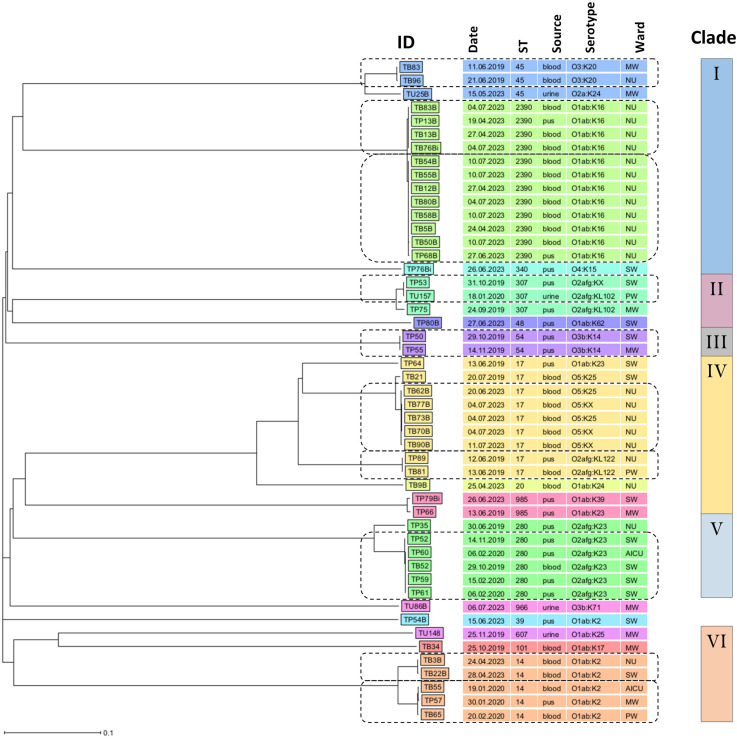 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Escherichia coli research studies · Vibrio bacteria research studies
Introduction
1
Antimicrobial resistance (AMR) represents a critical global health threat, with extended-spectrum beta-lactamase (ESBL)-producing Enterobacterales (PE), particularly Escherichia coli and Klebsiella pneumoniae, at the forefront [1]. These pathogens cause life-threatening bloodstream infections (BSIs), urinary tract infections (UTIs), and skin and soft tissue infections (SSTIs), with disproportionate high burden in low- and middle-income countries (LMICs) where diagnostic capacity and antimicrobial stewardship programs are limited [2,3].
A global burden study from 2019 estimated nearly 929 000 deaths associated with drug-resistant E. coli and K. pneumoniae, with significantly higher mortality in sub-Saharan African countries [2]. Globally, high-risk clones of ESBL-producing E. coli (ESBL-EC), i.e., sequence type 131 (ST131) and serotype O25:H4 and ESBL-producing K. pneumoniae (ESBL-KP), i.e., ST307 and serotype O2afg:KL102 have emerged as key resistance drivers [4,5]. In Tanzania, limited research-based genomic data are showing predominance of ESBL-EC ST131 and ESBL-KP ST14 and ST45 implicated in both gastrointestinal colonisation and infections [[6], [7], [8]]. These STs exhibit multidrug resistance, often harbouring blaCTX−M-15 alongside resistance to fluoroquinolones and aminoglycosides, with enhanced pathogenicity through virulence factors related to adhesion, invasion, immune evasion, and iron acquisition [4,5].
The World Health Organization (WHO) has prioritised third-generation cephalosporin-resistant Enterobacterales comprising ESBL-EC and ESBL-KP in its Global Action Plan on AMR and its Bacterial Priority Pathogens Lists, 2017 and 2024 [[9], [10], [11]]. Despite adopting national action plans on AMR (NAP-AMR) across LMICs, including Tanzania [12], genomic AMR surveillance data shortages persist, hampering infection prevention and control (IPC) efforts. Moreover, understanding local clonal clusters and resistance mechanisms is essential for effective targeted public health interventions. This study bridges this knowledge gap by utilising whole genome sequencing (WGS) to characterise ESBL-EC and ESBL-KP isolates causing BSIs, UTIs, and SSTIs among patients admitted or attending at Bugando Medical Centre (BMC), a zonal referral hospital in Mwanza, Tanzania. This research seeks to generate insights to guide targeted surveillance and IPC strategies in resource-limited healthcare settings through a comprehensive analysis of genomic epidemiology (diversity, resistance determinants, virulence factors, and clonal relationships) of ESBL-PE.
Materials and methods
2
Study design
2.1
This cross-sectional hospital-based study employed WGS to characterise ESBL-EC and ESBL-KP isolates recovered from blood, urine, and pus samples of patients with clinical diagnoses of BSIs, UTIs, and/or SSTIs, respectively. The samples were collected from patients attending or admitted to BMC, a zonal referral hospital in Mwanza, Tanzania, during June 2019–June 2020 and March–July 2023, as part of a previous research project [13,14].
Bacterial isolation and antimicrobial susceptibility testing
2.2
Routine bacterial culture, biochemical tests, and disk diffusion methods by Kirby-Bauer technique were employed for the isolation, species identification, and antimicrobial susceptibility testing (AST) of bacterial pathogens implicated in BSIs, UTIs, and SSTIs following standard operating procedures [15,16]. All isolates were stored at −80 °C until further analysis. Species verification was performed using an automated analyser, VITEK MS (Matrix-assisted laser desorption/ionisation – time of fright [MALDI-TOF]; bioMérieux, Germany). Whereas AST was performed using the VITEK 2 (bioMérieux, Germany) system with the AST-N214 card to determine the minimum inhibitory concentrations (MICs) of bacterial pathogens against antimicrobial agents. The AST-N214 card included ampicillin, cefpodoxime, cefuroxime, cefotaxime, trimethoprim-sulfamethoxazole, gentamicin, ciprofloxacin, piperacillin-tazobactam, tigecycline, and meropenem. The AST also included screening for ESBL production and assessment of multidrug resistance levels. Interpretation of MIC values was performed using the EUCAST 2023 guidelines [17].
Isolate selection and DNA extraction for WGS
2.3
ESBL-positive E. coli and K. pneumoniae strains classified as multidrug-resistant Gram-negative bacteria (MRGN) level ≥3 (resistance to at least one or more antimicrobials from three or more different antimicrobial classes) were selected for WGS. The isolates were subcultured on 5% sheep blood agar and incubated aerobically at 35 ± 2 °C for 24 h prior to DNA extraction using the Wizard® Genomic DNA Purification Kit (Promega, Germany). DNA samples with absorbance ratio A260/A280 ≥1.80 and A260/A230 2.0–2.2 by Nanodrop Spectrophotometer (bioMérieux, Germany) were accepted for WGS and stored at −20 °C.
WGS and bioinformatics analysis
2.4
WGS was performed on an Illumina NextSeq 500/550 instrument (Illumina, San Diego, CA) using Nextera XT library preparation and 2 × 150 bp paired-end sequencing. Raw sequences underwent quality control using FastQC, trimming with Trimmomatic, and de novo assembly using Velvet in Ridom SeqSphere+ v.8.4.0 (Ridom GmbH, Münster, Germany; https://www.ridom.de/seqsphere/). MLST sequence types were identified using the PubMLST database (https://pubmlst.org/mlst) within Ridom SeqSphere+. Additionally, Neighbour Joining (NJ) phylogenetic trees for ESBL-EC and ESBL-KP were later generated using the Bruker MBioSEQ Ridom Typer (formerly Ridom SeqSphere+), with the clonal clustering threshold set at ≤10 cgMLST allelic distance. In the current study, a clonal cluster represents bacterial isolates with minimal genetic divergence (≤10 cgMLST allelic differences) within a clone. A clone denotes a set of bacterial isolates forming a genetically uniform group as a result of recent shared ancestry (e.g., ST131), whereas a lineage represents a higher-order phylogenetic group encompassing multiple related clones with shared ancestry.
Comprehensive genomic characterisation employed multiple bioinformatics tools from the Center for Genomic Epidemiology (CGE) of the Technical University of Denmark (https://www.genomicepidemiology.org/services/): SeroTypeFinder 2.0 [18] for serotype identification, PlasmidFinder 2.1 [19,20] for plasmid replicon detection and typing, and ResFinder 4.6.0 [19,21] for antimicrobial resistance gene (ARG) identification. When ResFinder failed to detect third-generation cephalosporin resistance genes, the Resistance Gene Identifier (RGI) v6.0.5 [22] from the Comprehensive Antibiotic Resistance Database (CARD v4.0.1; https://card.mcmaster.ca/analyze/rgi) was utilised. All analyses performed using bioinformatics tools within CGE and CARD were conducted with default settings.
The CGE VirulenceFinder 2.0 [19] was used for virulence gene detection in ESBL-EC, whereas the virulence factor database (VFDB; https://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi) and Pathogenwatch (https://pathogen.watch/) were used to determine virulence factors and serotypes in ESBL-KP, respectively. Additionally, Kleborate [23] on Galaxy Europe (https://usegalaxy.eu/) was used for species identification and virulence scoring in ESBL-KP. Moreover, the ClermonTyping [24,25] was used for phylotyping of ESBL-EC into major phylogroups.
Statistical analysis
2.5
Data were analysed using STATA 15.0 [26]. Chi-square or Fisher's exact tests were used to compare categorical variables, while Wilcoxon rank-sum tests compared continuous variables. Statistical significance was set at P ≤ 0.05 and 95% confidence interval.
Results
3
Characteristics of sequenced isolates
3.1
A total of 88 ESBL-producing E. coli and K. pneumoniae isolates classified as MRGN level ≥3 were selected for WGS. These isolates were obtained from patients with a median age of 9.5 y [IQR: 0–42.5], the majority of whom were inpatients (96.6%; n = 85). Most isolates originated from blood samples (43.2%; n = 38), primarily from neonates admitted to the neonatology unit (34.1%; n = 30). Furthermore, 30.7% (n = 27) of patients reported a history of fever, 19.3% (n = 17) had used antibiotics within the past 3 months, and 13.6% (n = 12) had comorbid conditions (Table 1).Table 1. The demographic and clinical characteristics of patients whose ESBL-EC and ESBL-KP were sequenced.Table 1 dummy alt textVariablesFrequencyPercentageMedian [IQR]/n%Median [IQR] age in years9.5 [0–42.5]–SexFemale4551.1Male4348.9ResidenceRural4551.1Urban4348.9Patient categoryInpatient8596.6Outpatient33.4WardNeonatology3034.1Surgical2528.4Medical2326.1Others1011.4Sample typeBlood3843.2Pus3337.5Urine1719.3History of feverNo6169.3Yes2730.7History of antibiotic useNo7180.7Yes1719.3History of admissionNo8394.3Yes55.7Comorbidities*No7686.4Yes1213.6⁎Comorbidities: Hypertension only (n = 4), sickle cell disease (n = 3), diabetes mellitus only (n = 2), hypertension and diabetes mellitus (n = 2), and HIV (n = 1).
Sequence types, serotypes, and plasmid replicons
3.2
Of the 88 sequenced ESBL-PE isolates, 39 were ESBL-EC and 49 were ESBL-KP. Among the ESBL-EC, 13 sequence types (STs) were identified, with ST131 (30.7%; n = 12) and ST648 (28.2%; n = 11) being the most prevalent. The majority of ESBL-EC ST131 (7 out of 12) were isolated from urine samples. Nearly all ESBL-EC (94.9%; n = 37) were serotypable, predominantly O25:H4 (27.0%; n = 10) and O102:H6 (18.9%; n = 7) serotypes were detected. In addition, plasmid replicons were detected in 92.3% (n = 36) of the ESBL-EC, with the most common replicon types being IncFII (63.9%; n = 23), IncFIB (50.0%; n = 18), and IncFIA (38.9%; n = 14) (Table 2).Table 2. Sequence types, serotypes, and plasmid replicons of sequenced ESBL-EC and ESBL-KP.Table 2 dummy alt textVariablesFrequencyPercentagesIsolatesCharacteristicsCategoriesN%ESBL-EC (N = 39)STs1311230.76481128.2167410.261725.1Othersa1025.6SerotypableYes3794.9No25.1SerotypesO25:H41027.0O102:H6718.90101:H21410.8O153:H6410.8Othersb1027.0Detected plasmid repliconYes3692.3No37.7Plasmid repliconsIncFII2363.9IncFIB1850.0IncFIA1438.9Col440I616.7Othersc719.4ESBL-KP (N = 49)STs23901224.517918.4280612.214510.2Othersd1734.7SerotypableYes4591.8No48.2SerotypesO1ab:K161226.7O1ab:K2613.3O2afg:K23613.3O5:K2548.9Otherse1328.9Detected plasmid repliconYes49100No00.0Plasmid repliconsIncFII4183.7IncFIB3979.6IncR1836.7repB1428.6Othersf1734.7ESBL-EC.ESBL-KP.aOthers: ST-10 (n = 1), ST-156 (n = 1), ST-224 (n = 1), ST-410 (n = 1), ST-1193 (n = 1), ST-1380 (n = 1), ST-2852 (n = 1), ST-3580 (n = 2), and ST-4981 (n = 1).bOthers: O101:H10 (n = 3), O16:H5 (n = 2), O101:H9 (n = 1), O8:H12 (n = 2), O17/O77:H18 (n = 1), O75:H5 (n = 1), O8:H23 (n = 1), and O8:H7 (n = 1).cOthers: IncQ1 (n = 2), IncX4 (n = 2), IncY (n = 3), IncI1-I (n = 3), and ColB (n = 1).dOthers: ST-20 (n = 1), ST-39 (n = 1), ST-45 (n = 3), ST-48 (n = 1), ST-54 (n = 2), ST-101 (n = 1), ST-307 (n = 3), ST-340 (n = 1), ST-607 (n = 1), ST-966 (n = 1), and ST-985 (n = 2).eOthers: O2afg:KL102 (n = 2), O2afg:KL122 (n = 2), O3:K20 (n = 2), O3b:K14 (n = 2), O1ab:K17 (n = 1), O1ab:K23 (n = 1), O1ab:K25 (n = 1), O1ab:K24 (n = 1), O1ab:K39 (n = 1), O1ab:K62 (n = 1), O2a:K24 (n = 1), O3b:K71 (n = 1), and O4:K15 (n = 1).fOthers: Col440II (n = 9), Col440I (n = 3), IncM1 (n = 2), IncHI1B (n = 1), p0111 (n = 1), and IncFIA (n = 1).
All ESBL-KP were identified by Kleborate as K. pneumoniae subsp. pneumoniae. Fifteen STs were detected, with ST2390 (24.5%; n = 12) and ST17 (18.4%; n = 9) being predominant. The ST2390 was reported in Tanzania for the first time in this study. Notably, 10 of the 12 ESBL-KP ST2390 isolates were recovered from blood samples. Most ESBL-KP (91.8%; n = 45) were serotypable, with frequent serotypes including O1ab:K16 (26.7%; n = 12), O1ab:K2 (13.3%; n = 6), and O2afg:K23 (13.3%; n = 6). Plasmid replicons were detected in all ESBL-KP isolates, with the most common replicons being IncFII (83.7%; n = 41), IncFIB (79.6%; n = 39), and IncR (36.7%; n = 18). Plasmid replicon repB was exclusively detected in ESBL-KP and notably in all ESBL-KP ST2390 (n = 12) (Table 2).
Phenotypic antimicrobial resistance patterns
3.3
ESBL-PE demonstrated high resistance rates (>50%) to most antimicrobial agents, with the exception of meropenem, against which no resistance was detected in either ESBL-EC or ESBL-KP. ESBL-EC exhibited significantly higher resistance to ciprofloxacin compared to ESBL-KP (87.2% vs. 53.1%, P < 0.01), whereas ESBL-KP showed markedly greater resistance to gentamicin (97.9% vs. 71.8%, P < 0.01) and piperacillin-tazobactam (67.3% vs. 28.2%, P < 0.01). Low resistance to tigecycline was observed among ESBL-EC (7.7%; 3/39); however, this agent was not tested for ESBL-KP, and thus, corresponding results are not presented (Fig. 1).Fig. 1. Percentage resistance of ESBL-EC (blue bars) and ESBL-KP (orange bars) against antimicrobial agents. Key: CIP = ciprofloxacin; GEN = gentamicin; TGC = tigecycline (ESBL-EC only); TZP = piperacillin-tazobactam; and SXT = trimethoprim-sulfamethoxazole.Fig 1 dummy alt text
Genomic determinants of AMR in ESBL-EC and ESBL-KP
3.4
The blaCTX−M-15 gene was the predominant determinant of resistance to third-generation cephalosporins (3GCs), and thus ESBL production, in both ESBL-EC (87.2%; n = 34) and ESBL-KP (95.9%; n = 47). Despite this similarity, notable differences were observed in the distribution of other ARGs between the two species, ESBL-EC and ESBL-KP. For aminoglycoside resistance, aac(6′)-Ib-cr was most common in ESBL-EC (61.5%), whereas aac(3)-IIa predominated in ESBL-KP (67.3%). Regarding trimethoprim resistance, dfrA17 (61.5%) was most frequent in ESBL-EC, while OqxB (53.1%) was predominant in ESBL-KP. Similarly, for amphenicol resistance, catB3 (43.6%) was the leading gene in ESBL-EC, compared to OqxB (53.1%) in ESBL-KP. Interestingly, fosA (36.7%; n = 18) and arr-3 (30.6%; n = 15), conferring resistance to fosfomycin and rifamycin, respectively, were detected exclusively in ESBL-KP (Table 3).Table 3. Genomic determinants of AMR in ESBL-EC and ESBL-KP.Table 3 dummy alt textAntimicrobial classResistance geneESBL-EC (N = 39)ESBL-KP (N = 49)n (%)n (%)Beta-lactamsbla_CTX−M-15_34 (87.2)47 (95.9)bla_TEM-1B_26 (66.7)39 (82.6)bla_OXA-1_24 (61.5)13 (26.5)blaSHV-65–12 (24.5)blaSHV-1–10 (20.4)blaSHV-100–7 (14.3)Others5 (13.0)20 (40.8)Aminoglycosidesaac(6′)-Ib-cr24 (61.5)30 (61.2)aadA520 (51.3)1 (2.0)aph(6)-Id17 (43.6)26 (53.1)aac(3)-IIa15 (38.5)33 (67.3)aph(3″)-Ib9 (23.1)11 (22.4)aadA23 (7.7)1 (2.0)aph(3′)-Ia2 (5.1)12 (24.5)aac(3)-IId2 (5.1)6 (12.2)AminocyclitolaadA520 (51.3)1 (2.0)aadA23 (7.7)1 (2.0)aadA16–10 (20.4)aadA1–1 (2.0)Quinolonesaac(6′)-Ib-cr24 (61.5)30 (61.2)qnrS15 (12.8)–qepA41 (2.6)–OqxB–26 (53.1)qnrB1–8 (16.3)Sulfamethoxazolesul123 (59.0)14 (28.6)sul217 (43.6)34 (69.4)sul31 (2.6)–TrimethoprimdfrA1724 (61.5)–dfrA144 (10.3)13 (26.5)dfrA123 (7.7)–dfrA11 (2.6)–OqxB–26 (53.1)dfrA27–3 (6.1)dfrA72–2 (4.1)Tetracyclinestet(A)19 (48.7)20 (40.8)tet(B)15 (38.5)–tet(D)–12 (24.5)AmphenicolscatB317 (43.6)–OqxB–26 (53.1)catA2–2 (4.1)FosfomycinfosA–18 (36.7)Rifamycinarr-3-15 (30.6)Others: ESBL-EC: blaOXA-320 (n = 1), blaOXA-534 (n = 1), blaTEM-34 (n = 1), blaTEM-206 (n = 1), and blaTEM-216 (n = 1) and ESBL-KP: blaSHV-187 (n = 3), blaSHV-28 (n = 2), blaSHV-148 (n = 2), blaSHV-172 (n = 2), blaSHV-27 (n = 1), blaSHV-56 (n = 1), blaSHV-106 (n = 1), blaSHV-161 (n = 1), blaSHV-182 (n = 1), blaTEM-1C (n = 1), blaTEM-30 (n = 1), blaTEM-234 (n = 1), blaOXA-320 (n = 1), blaOXA-534 (n = 1), and blaCTX−M-27 (n = 1).
Five ESBL-EC lacked detectable 3GCs-resistance genes when screened using ResFinder at the CGE. However, further analysis with the RGI revealed the presence of genes encoding AmpC β-lactamases, including chromosomally encoded variants (EC-8, EC-15, and Ecol_ampC_BLA) and the plasmid-mediated gene blaCMY-181. Overall, discrepancies were noted between genotypic and phenotypic resistance profiles, underscoring the complexity of resistance mechanisms in these Yisolates (Supplementary Table 1).
Genomic determinants of virulence factors in ESBL-EC and ESBL-KP
3.5
In ESBL-EC, the detected virulence genes (VGs) were classified into seven functional categories: (i) adhesion and colonisation, (ii) capsule and biofilm formation, (iii) invasion and intracellular survival, (iv) immune evasion and serum resistance, (v) iron acquisition systems, (vi) toxin production, and (vii) antibacterial activity and survival. The most prevalent genes included fimH (89.7%), aslA (66.7%), and csgA (66.7%) encoding adhesion and colonization; nlpI and yehA/B/C/D (100%, each) encoding capsule and biofilm formation; iha (38.5%) associated with invasion and intracellular survival; terC (100%) and traJ/T (64.1%) encoding immune evasion and serum resistance; fyuA (82.1%), irp2 (82.1%), and sitA (76.9%) encoding iron acquisition systems; hlyA/E/F (74.4%) encoding pore forming haemolysins/toxins; and cma and clpK1 (5.1%, each) encoding proteins associated with antibacterial activity and survival (Table 4).Table 4. Proportions and distributions of genes encoding virulence factors in ESBL-EC.Table 4 dummy alt textFunctional categoriesVirulence gene (VG)FrequencyPercentagesn%Adhesion and colonisationfimH3589.7aslA2666.7csgA2666.7yfcV2564.1Hra2256.4lpfA1846.2afaA/B/C/D/E1641.0papA/C1743.6fdeC410.3faeF12.6aap12.6aggR/4C25.1Capsule and biofilm formationnlpI39100yehA/B/C/D39100kpsE/M2564.1capU12.6Invasion and intracellular survivaliha1538.5Tia25.1Cia25.1Immune evasion and serum resistanceterC39100traJ/T2564.1Iss1846.2ompT1435.9gad410.3eatA12.6usp12.6Iron acquisition systemsfyuA3282.1irp23282.1sitA3076.9iutA2769.2chuA2666.7iucC2564.1ireA820.5mchC/F25.1iroN12.6neuC12.6ToxinshlyA/E/F2974.4sat1333.3cnf11128.2senB820.5astA410.3sepA12.6Antibacterial activity and survivalcma25.1clpK125.1colE412.6
On the other hand, VGs identified in ESBL-KP were grouped into three main functional categories: (i) adhesion and colonisation, (ii) capsule formation and immune evasion, and (iii) iron acquisition systems. The genes fimA-I/K and mrkA-D/F/H/I/J associated with adhesion and colonisation; acrA/B and rcsA/B encoding capsule formation and immune evasion; and entA-E/F/S, fepA-D/G, fes, and iroE associated with iron acquisition systems were detected in all ESBL-KP (Table 5). All ESBL-KP ST2390 isolates were found to harbour entA-E/F/S, fepA-D/G, fes, and iroE, while irp1/2 and ybtA/E/P/Q/R/S/T/U/X were detected in 11 of the 12 isolates. According to the Kleborate virulence scoring system, 61.2% (30/49) of ESBL-KP isolates had a virulence score of 1.Table 5. Proportions and distributions of genes encoding virulence factors in ESBL-KP.Table 5 dummy alt textFunctional categoriesVirulence gene (VG)FrequencyPercentagesn%Adhesion and colonisationfimA/B/C/D/E/F/G/H/I/K49100mrkA/B/C/D/F/H/I/J49100Capsule formation and immune evasionacrA/B49100rcsA/B49100wzi3775.5Iron acquisition systemsentA/B/C/D/E/F/S49100fepA/B/C/D/G49100Fes49100iroE49100irp1/23163.3ybtA/E/P/Q/R/S/T/U/X3061.2fyuA3061.2
Phylogenetic analysis of ESBL-PE
3.6
The NJ phylogenetic tree based on cgMLST analysis of ESBL-EC isolates demonstrated considerable genetic diversity, identifying six phylogroups and clonal clusters within four phylogroups: B2, F, A, and B1. Phylogroup B2 had two clonal clusters of ST131, serotype O25:H4: the first clonal cluster with two strains from pus (TP23Bi; 3 May 2023; medical ward) and urine (TU11B; 2 May 2023; medical ward) and the second clonal cluster with three strains from urine (TU161; 30 June 2019; neonatology unit), blood (TB73; 24 June 2019; paediatric ward), and pus (TP71; 12 June 2019; neonatology unit). Phylogroup F had two clonal clusters: one clonal cluster contained pus (TP34; 11 June 2019; neonatology unit) and urine (TU114; 5 July 2019; surgical ward) isolates, both ST-648, serotype O153:H6, and the other clonal cluster comprised seven isolates from blood samples (TB33; 22 October 2019; medical ward), (TB58 and TB60: 10 February 2020; medical ward), (TB61; 6 February 2020; surgical ward), pus samples (TP58 and TP87: 1 January 2020; paediatric ward), and urine sample (TU158; 5 February 2020; medical ward), all ST-648, serotype O102:H6. Phylogroup A contained one clonal cluster of two blood isolates: TB54 (22 November 2019; medical ward) and TB62 (15 February 2020; paediatric ward) belonging to ST167, serotype O101:H21. Lastly, phylogroup B1 had one clonal cluster of two pus isolates: TP28B and TP32B (9 May 2023; surgical ward) of ST3580, serotype O8:H12 (Fig. 2).Fig. 2. An NJ phylogenetic tree of ESBL-EC, based on cgMLST analysis. In the isolate IDs, the letter ‘T’ denotes Tanzania, followed by ‘B’, ‘U’, or ‘P’, which specify the sample type, blood, urine, and pus, respectively. The dashed rounded rectangles indicate clonal clusters corresponding to each phylogroup. Abbreviations: MOPD = medical outpatient; MW = medical ward; NU = neonatology unit; PW = paediatric ward; SW = surgical ward; NA = not serotypable.Fig 2 dummy alt text
The cgMLST-based NJ phylogenetic analysis of ESBL-KP isolates revealed high genetic diversity, with six clades (as visually delineated by the authors) and clonal clusters within clades. Clade I comprised three clonal clusters: the first with two blood isolates (TB83; 11 June 2019; medical ward) and (TB96; 21 June 2019; neonatology unit) of ST45 and serotype O3:K20; the second with four isolates (ST2390, serotype O1ab: K16) from neonatology unit, including TB83B and TB76Bi (blood; 4 July 2023, both), TP13B (pus; 19 April 2023), and TB13B (blood; 27 April 2023); and the third with eight isolates (ST2390, serotype O1ab: K16) also from neonatology unit, including TB54B and TB55B (blood; 10 July 2023, both), TB12B (blood; 27 April 2023), TB80B (blood; 27 June 2023), TB58B (blood; 10 July 2023), TB5B (blood; 24 April 2023), TB50B (blood; 10 July 2023), and TP68B (pus; 27 June 2023). Clade II contained one clonal cluster of two isolates belonging to ST307: TP53 (pus; 31 October 2019; surgical ward) and TU157 (urine; 18 January 2020; paediatric ward). Clade III had once a clonal cluster of ST54, serotype O3b: K14, made of two isolates: TP50 (pus; 29 October 2019; surgical ward) and TP55 (pus; 14 November 2019; medical ward). Clade IV had two clonal clusters belonging to ST17: the first clonal cluster had five blood isolates, TB62B (20 June 2023), TB77B, TB73B, TB70B (4 July 2023, both) and TB90B (11 July 2023), all from neonatology unit, serotype O5:K25, and the second clonal cluster had two isolates, TP89 (pus; 12 June 2019; neonatology unit) and TB81 (blood; 13 June 2019; paediatric ward), serotype O2afg: KL122. Clade V had one clonal cluster of ST280, serotype O2afg: K23, made up of five isolates: TP52 (pus; 14 November 2019; surgical ward), TP60 (pus; 6 February 2020; AICU), TB52 (blood; 29 October 2019; surgical ward), TP59 (pus; 15 February 2020; surgical ward), and TP61 (pus; 6 February 2020; surgical ward). Clade VI contained two clonal clusters of ST14, serotype O1ab:K2: the first with two blood isolates TB3B (24 April 2023; neonatology unit) and TB22B (28 April 2023; surgical ward), and the second with three isolates, two from blood samples TB55 (19 January 2020; AICU) and TB65 (20 February 2020; paediatric ward), and one from pus sample TP57 (3 January 2020; medical ward) (Fig. 3).Fig. 3. An NJ phylogenetic tree of ESBL-KP, based on cgMLST analysis. In the isolate IDs, the letter ‘T’ denotes Tanzania, followed by ‘B’, ‘U’, or ‘P’, which specify the sample type, blood, urine, and pus, respectively. The dashed rounded rectangles indicate clonal clusters corresponding to each clade. Abbreviations: AICU = adult intensive care unit; MW = medical ward; NU = neonatology unit; PW = paediatric ward; SW = surgical ward.Fig 3 dummy alt text
Discussion
4
This comprehensive genomic epidemiological study reveals the presence of the global high-risk clone ESBL-EC ST131 and the emergence of ESBL-KP ST2390 reported in Tanzania for the first time in this study. The ESBL-KP ST2390 isolates harboured key virulence factors, notably iroE/N and ybtA/E/P/Q/R/S/T/U/X, suggesting that this ST may represent a potentially high-risk clone [27,28]. However, ESBL-KP ST2390 strains have been previously reported elsewhere, for instance, in China in 2019 [29]. Furthermore, we observed clonal transmission/expansion events of these global high-risk strains at this setting, mostly within neonatology units, highlighting potential gaps in infection prevention and control strategies.
In line with a previous study from the same setting [6] and elsewhere [30], we observed the predominance of ST131 phylogroup B2 in ESBL-EC. Alongside ST131 phylogroup B2, we identified ST648 phylogroup F as the second most prevalent lineage. Both ST131 phylogroup B2 and ST648 phylogroup F are well-documented extraintestinal pathogenic E. coli (ExPEC) lineages, characterised by their heightened virulence and extensive multidrug resistance [5,31]. Additionally, the dominance of uropathogenic ESBL-EC ST131 may be associated with specific virulence factors of this clone, including P fimbriae and type 1 fimbriae for superior bladder epithelial adhesion; iron-acquisition systems crucial for the iron-limited urinary tract environment; and increased biofilm-forming capacity on urinary bladder epithelium [32]. On the other hand, the predominance of ST2390 in ESBL-KP and particularly its clonal expansion in neonatology units, underscores the BSIs outbreak in the neonatology unit and the potential risk to vulnerable neonates. Generally, the predominance of K. pneumoniae in BSIs is favoured by its multiple and effective iron acquisition systems [33]. In contrast to the sustained dominance of ESBL-EC ST131, the predominant STs of ESBL-KP in our setting have fluctuated over time. For instance, Mshana et al. [7] identified ST14 as the leading clone in 2009–2010 [8], while Marando et al. later reported ST45 as predominant in 2016. The fluctuation of ESBL STs over time in our setting may partly be attributed to differences in the sampled patient populations or ward distributions. Additionally, despite the fluctuations of STs among ESBL-KP over time, the overall trend highlights the continued presence of a globally recognised high-risk clone within healthcare environments [34].
Consistent with previous studies [7,35], IncFIB was the most prevalent plasmid replicon among ESBL-EC and ESBL-KP, supporting a major role for conjugative IncF-type plasmids in the dissemination of resistance and virulence determinants [36]. Nevertheless, this study did not examine the specific localisation of ARGs or VFs within these plasmid replicons. Further, our observation of the dominance of high-risk global clone ESBL-EC ST131 and the newly detected ESBL-KP ST2390 highlights that clonal expansion contributes substantially to the spread of ARGs and rapid dissemination of AMR in our setting.
Notably, all ESBL-PE isolates in this study were susceptible to meropenem, aligning with previous studies in Tanzania [37] and Sri Lanka [38], underscoring its continued reliability as a last-resort agent for MDR infections, including those caused by ESBL-EC and ESBL-KP. This observation further illustrates the positive impact of antimicrobial stewardship efforts in curbing inappropriate use of this critical agent within our setting. However, consistent with previous studies in India [39] and Turkey [40], resistance to ciprofloxacin was significantly higher among ESBL-EC than ESBL-KP. Although ciprofloxacin resistance in ESBL-EC, particularly the ST131 clone, is predominantly mediated by chromosomal mutations [41], our findings show that most ESBL-EC isolates harboured acquired ciprofloxacin resistance genes. Conversely, we observed higher resistance to gentamicin and piperacillin-tazobactam among ESBL-KP than ESBL-EC. Collectively, these findings point to a species-specific dissemination of ARGs, likely driven by clonal expansion, as exemplified by the predominance of ESBL-EC ST131 and ESBL-KP ST2390 in our setting.
We observed the predominance of blaCTX−M-15 in both ESBL-EC and ESBL-KP, underscoring its widespread dissemination as a key determinant of 3GCs resistance among Enterobacterales. Nonetheless, we observed genotype-phenotype discordance, exemplified by the absence of detectable acquired resistance genes, particularly blaCTX−M genes, in five ESBL-EC isolates. The AmpC β-lactamase-mediated resistance, an intrinsic mechanism in most E. coli [42], conferring resistance to β-lactams, including third-generation cephalosporins, may plausibly explain the ESBL-like phenotypes observed in the five ESBL-EC isolates in which no ESBL genes were detected. On the other hand, cryptic resistance genes present in the bacterial genome may remain phenotypically silent due to low expression levels, regulatory mutations, or suboptimal promoter activity, thereby accounting for the susceptible phenotypes observed among genotypically resistant isolates [43].
The virulence landscapes of ESBL-EC and ESBL-KP differ markedly in both diversity and functional emphasis. ESBL-EC harboured a wide array of VFs spanning multiple categories, indicative of high adaptability and opportunistic pathogenicity typical of ExPEC, such as ST131 [44]. By contrast, ESBL-KP exhibited a more conserved virulence profile, largely centred on adhesion, capsule production, and iron acquisition, underscoring the importance of colonisation and nutrient uptake. Additionally, the predominance of low Kleborate scores among ESBL-KP further suggests that multidrug resistance, rather than enhanced virulence, constitutes its primary advantage in healthcare-associated infections [45]. Nonetheless, a comprehensive understanding of VFs would require direct comparisons between 3GC-resistant and susceptible or ESBL-PE and non-ESBL-PE isolates, which were beyond the scope of this study.
The NJ phylogenetic analysis based on cgMLST revealed clonal clusters, suggesting potential intrahospital transmission of ESBL-PE. Specifically, this included the high-risk clone ESBL-EC ST131 and the newly identified ESBL-KP ST2390, occurring across patients and clinical conditions, particularly BSIs, UTIs, and SSTIs. Notably, ESBL-EC ST131 was detected circulating in medical, neonatology, and paediatric wards, while ST648 circulated within medical, surgical, and paediatric wards. In contrast, ESBL-KP ST2390 showed persistent circulation within the neonatology unit, indicative of extensive clonal expansion in this setting. Collectively, these findings point to shortcomings in IPC measures and suggest the potential role of environmental reservoirs or healthcare-associated transmission routes in sustaining these clones [46].
Importantly, our study aligns with the implementation gaps highlighted in the WHO-JEE 2023 assessment of Tanzania, which noted the country’s limited capacity for systematic genomic surveillance of AMR, IPC, and AMS [47]. By identifying an emerging ESBL-KP ST2390 in the neonatology unit and documenting its spread in critical care settings, our findings contribute to bridging this gap. However, significant challenges remain regarding nationwide representativeness, integration of genomic data into routine AMR surveillance, and linkage with clinical outcomes. Overall, the predominance of high-risk clone ST131 and the emergence of ST2390 underscore the urgent need to strengthen genomic surveillance systems and implement targeted infection prevention measures, particularly in vulnerable populations such as neonates, through improved hand hygiene, rigorous environmental monitoring, and strict contact precautions.
Study limitations
4.1
This single-centre study may limit generalizability to other healthcare settings in Tanzania or similar LMICs. The relatively small sample size and lack of environmental screening may have underestimated transmission pathways. Additionally, the absence of patient outcome data prevents assessment of the clinical impacts of these genomic changes.
Conclusion
5
This study provides a comprehensive WGS-based genomic epidemiological characterisation of ESBL-EC and ESBL-KP causing BSIs, UTIs, and SSTIs at Bugando Medical Centre in Mwanza, Tanzania. The presence of globally recognised high-risk clone ESBL-EC ST131 and the emerging ESBL-KP ST2390, together with evidence of clonal clusters, especially within medical wards and neonatology units, highlights substantial gaps in IPC. The dominance of blaCTX−M-15, species-specific resistance patterns, and distinct virulence landscapes underscore the role of clonal expansion in driving AMR. Collectively, these findings emphasise the urgent need to strengthen genomic surveillance, integrate WGS into routine AMR monitoring, and implement targeted IPC strategies to mitigate the spread of high-risk ESBL-producing pathogens in resource-limited healthcare settings.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abay G.K.Shfare M.T.Teklu T.G.Kidane K.M.Gebremeskel T.K.Kahsay A.G.Extended-spectrum β-lactamase production and antimicrobial resistance among Enterobacteriaceae causing clinical infections in Africa: a systematic review and meta-analysis (2012–2020)Eur J Med Res 3020251143977333010.1186/s 40001-024-02267-8PMC 11706086 · doi ↗ · pubmed ↗
- 2Murray C.J.Ikuta K.S.Sharara F.Swetschinski L.Aguilar G.R.Gray A.Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis The Lancet 399202262965510.1016/S 0140-6736(21)02724-0PMC 884163735065702 · doi ↗ · pubmed ↗
- 3Pokharel S.Raut S.Adhikari B.Tackling antimicrobial resistance in low-income and middle-income countries BMJ Specialist J 42019 e 00210410.1136/bmjgh-2019-002104 PMC 686112531799007 · doi ↗ · pubmed ↗
- 4Mazumder R.Hussain A.Abdullah A.Islam M.N.Sadique M.T.Muniruzzaman S.International high-risk clones among extended-spectrum β-lactamase–producing Escherichia coli in Dhaka, Bangladesh Front Microbiol 12202173646410.3389/fmicb.2021.736464 PMC 852114434671331 · doi ↗ · pubmed ↗
- 5Peirano G.Chen L.Kreiswirth B.N.Pitout J.D.Emerging antimicrobial-resistant high-risk Klebsiella pneumoniae clones ST 307 and ST 147Antimicrob Agents Chemother.64202010.1128/aac.01148-01120 PMC 750859332747358 · doi ↗ · pubmed ↗
- 6Seni J.Peirano G.Mshana S.E.Pitout J.D.De Vinney R.The importance of Escherichia coli clonal complex 10 and ST 131 among Tanzanian patients on antimicrobial resistance surveillance programs Eur J Clin Microbiol Infect Dis 4020211103400952910.1007/s 10096-021-04271-w · doi ↗ · pubmed ↗
- 7Mshana S.E.Hain T.Domann E.Lyamuya E.F.Chakraborty T.Imirzalioglu C.Predominance of Klebsiella pneumoniae ST 14 carrying CTX-M-15 causing neonatal sepsis in Tanzania BMC Infect Dis 1320134662409928210.1186/1471-2334-13-466PMC 3851032 · doi ↗ · pubmed ↗
- 8Marando R.Seni J.Mirambo M.M.Falgenhauer L.Moremi N.Mushi M.F.Predictors of the extended-spectrum-beta-lactamases producing Enterobacteriaceae neonatal sepsis at a tertiary hospital, Tanzania Int J Med Microbiol 30820188038112998037210.1016/j.ijmm.2018.06.012PMC 6171784 · doi ↗ · pubmed ↗