Promoter competition and Polycomb response elements govern transvection efficiency between co-regulated engrailed and invected genes in Drosophila
Yuzhong Cheng, Adrienne T Perkins, Fountane W Chan, Natalie D Gehred, Jack R Bateman, Judith A Kassis

TL;DR
This paper explores how gene regulation occurs between paired chromosomes in fruit flies, revealing how promoters and enhancers interact to control gene activity.
Contribution
The study identifies promoter competition and Polycomb response elements as key regulators of transvection efficiency between co-regulated genes.
Findings
The en promoter in cis prevents trans-activation of inv, but its removal enables robust transvection.
Polycomb response elements enhance transvection reliability but are not essential for trans-activation.
Transvection at the en-inv locus is developmentally regulated and can be reconstituted in transgenic constructs.
Abstract
Transvection is a phenomenon where gene regulatory elements interact between different chromosomes, adding an additional layer of regulatory control beyond traditional cis-interactions. Although transvection effects have been characterized for many individual genes in Drosophila, it remains unclear how trans-interactions occur among multiple co-regulated genes where enhancers are shared. Here we demonstrate that transvection is supported at the engrailed-invected (en-inv) locus, where transcription of the two developmental genes is coordinated by common enhancers. Our data show that the presence of the en promoter in cis to the enhancers prevents trans-activation of inv, but removal of this promoter enables robust transvection, demonstrating competition between heterologous promoters in trans. We also find that local Polycomb response elements (PREs) enhance transvection reliability but…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
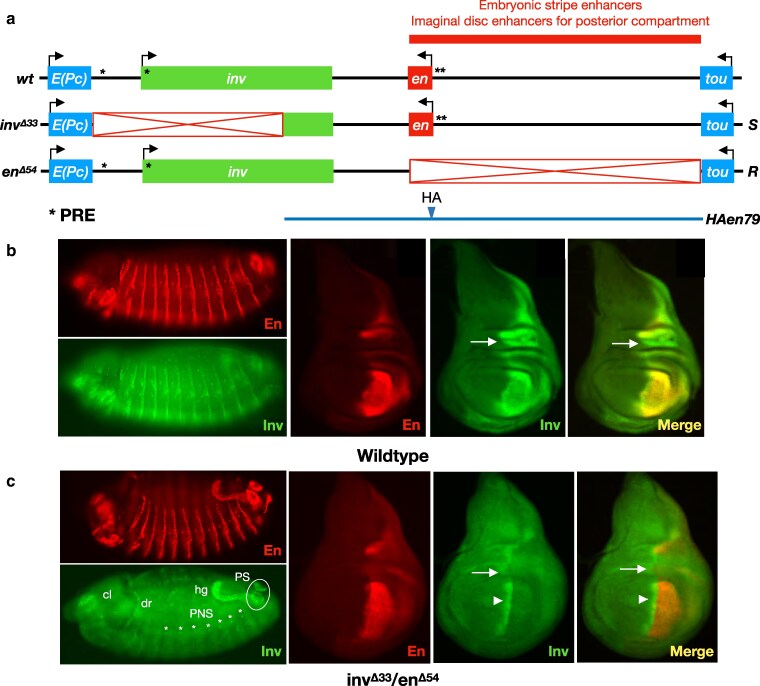 Fig. 1
Fig. 1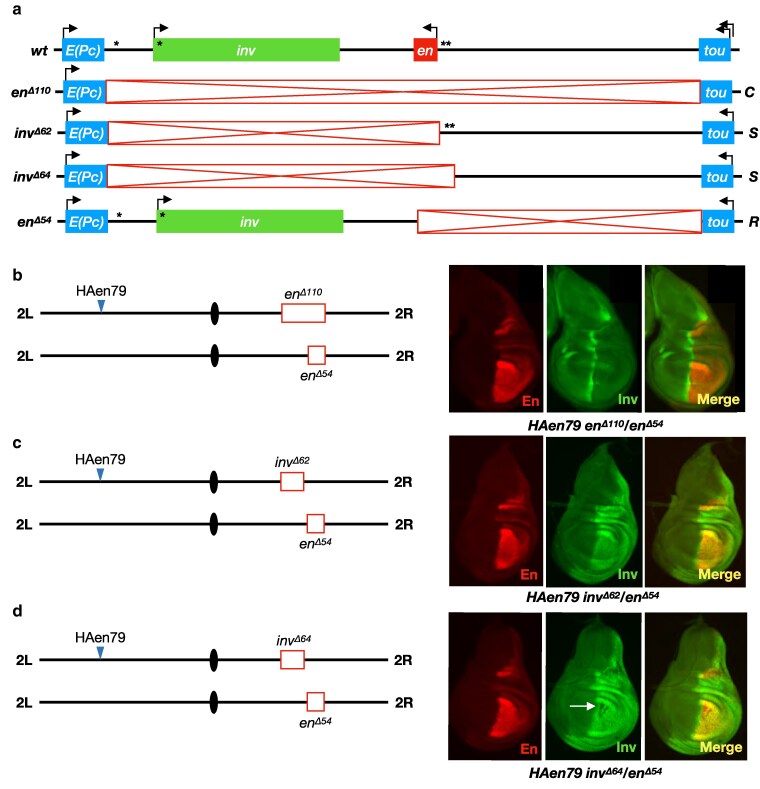 Fig. 2
Fig. 2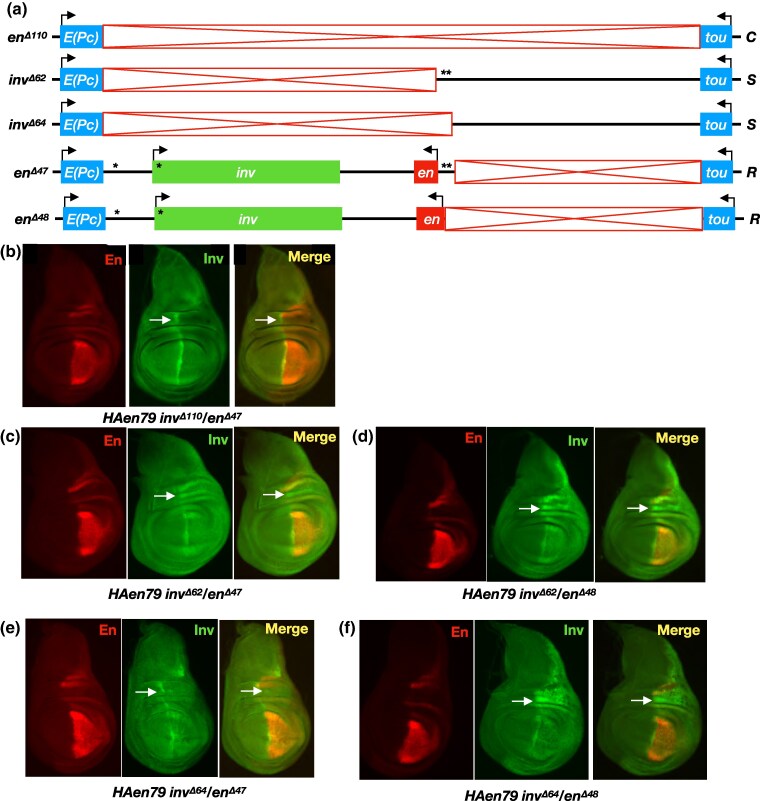 Fig. 3
Fig. 3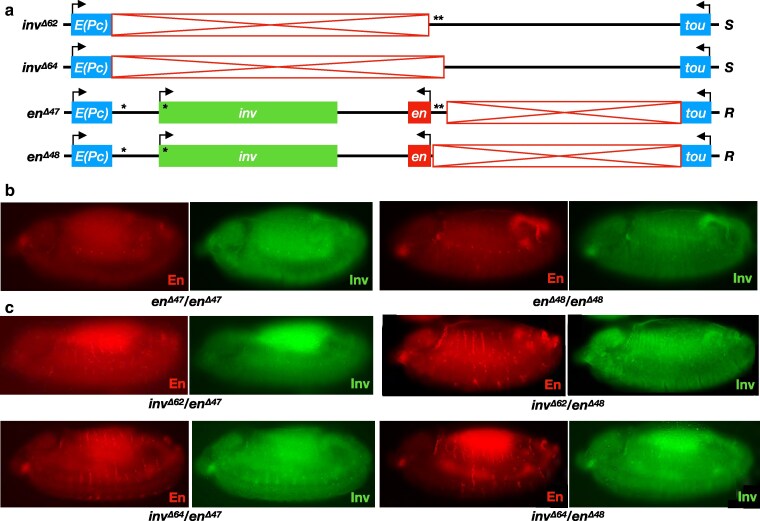 Fig. 4
Fig. 4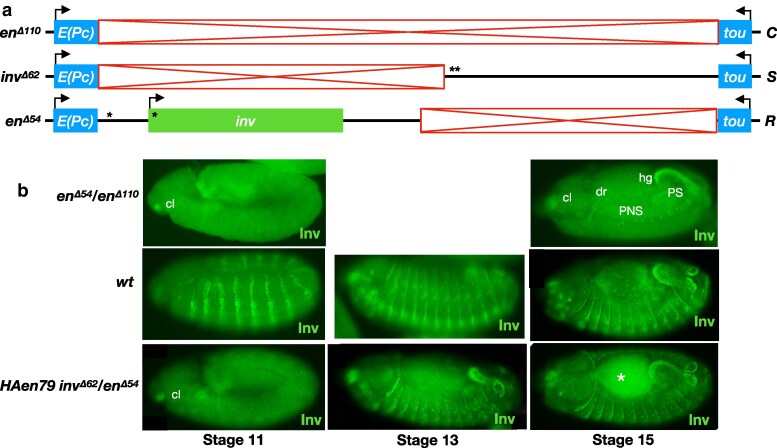 Fig. 5
Fig. 5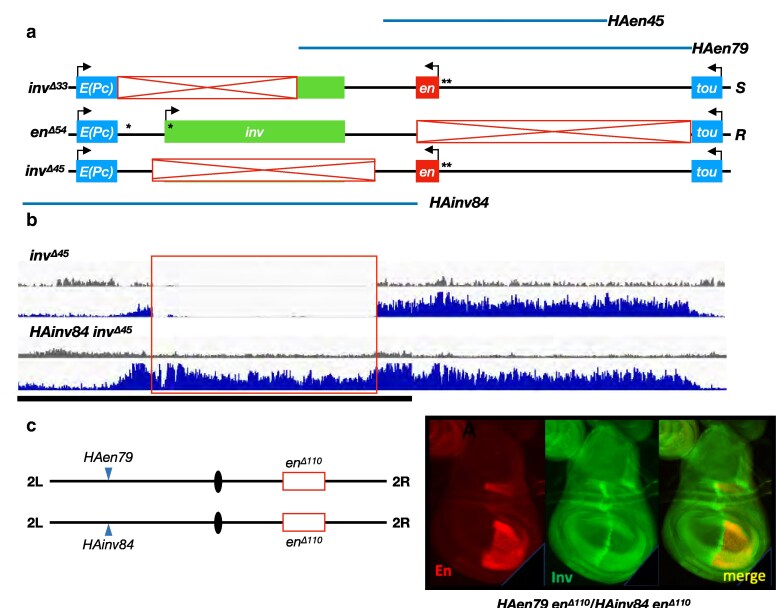 Fig. 6
Fig. 6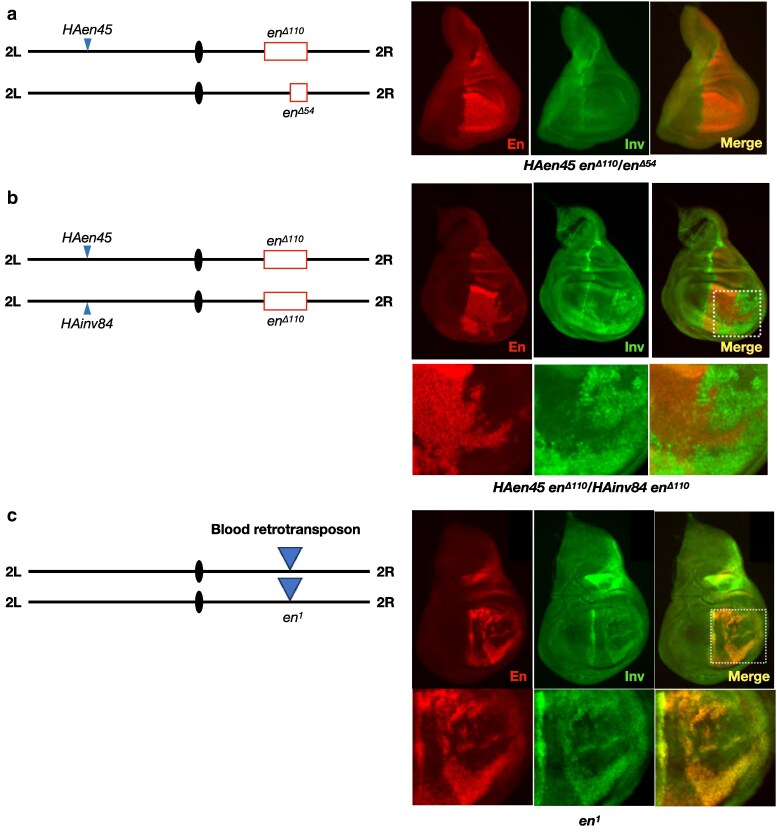 Fig. 7
Fig. 7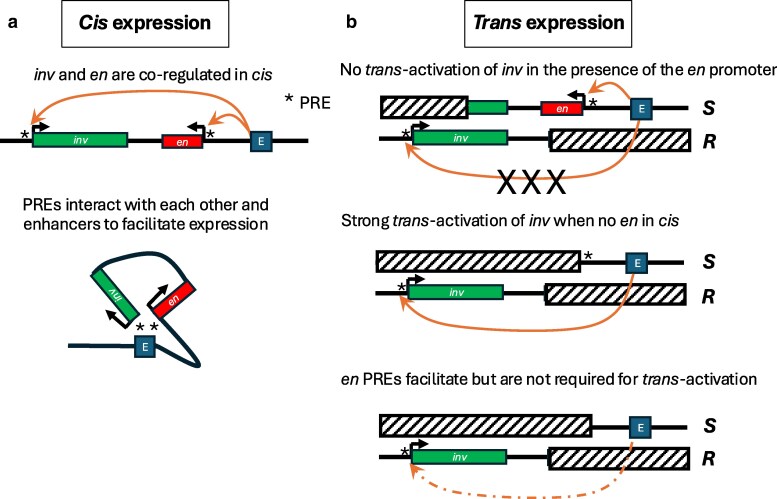 Fig. 8
Fig. 8| Name | Deleted sequences | Reference |
|---|---|---|
|
| 11466259..11499376 | ( |
|
| 11469702..11514649 | ( |
|
| 11466241..11528210 | this study |
|
| 11466241..11529821 | this study |
|
| 11529803..11576474 | this study |
|
| 11528279..11576472 | this study |
|
| 11522241..11576472 | this study |
|
| 11466238..11577472 | ( |
|
| 11496174..11537511 | ( |
- —National Institute of Child Health and Human Development10.13039/100009633
- —Department of Health and Human Services10.13039/100012737
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Genomics and Chromatin Dynamics · Neurobiology and Insect Physiology Research
Introduction
The spatial organization of the genome has emerged as a fundamental mechanism governing gene expression in eukaryotes. Chromosomes adopt hierarchical three-dimensional conformations that bring regulatory elements and target promoters into precise spatial relationships while simultaneously partitioning the genome into active and inactive compartments, enabling transcriptional programs that define cell identity (reviewed by Acemel and Lupiáñez 2023; Hehmeyer et al. 2023). Understanding how three-dimensional genome structure influences gene regulation is important for deciphering normal development, disease pathogenesis, and the potential for novel therapeutic interventions.
Many layers of 3D genome organization appear conserved across Eukarya, but the proteins that establish these conformations can differ from species to species. Common to diverse organisms are topologically associated domais (TADs), chromosomal segments of high self-interaction that tend to be flanked by insulator sequences (reviewed by Bhattacharya et al. 2024). In mammalian systems, a model has emerged where TADs are formed by cohesin-dependent loop extrusion, which can be halted by CTCF proteins bound to insulators, thus forming TAD boundaries. While the Drosophila genome is also characterized by partitioning into TADs, cohesin and CTCF do not play a major role in TAD formation in this species (reviewed by Matthews and White 2019). Rather, other mechanisms appear to drive local compartmentalization of the Drosophila genome, including preferential clustering of similarly modified chromatin states and/or the binding activities of species-specific architectural proteins (Ulianov et al. 2016; Rowley et al. 2017; Bhattacharya et al. 2024).
In addition to the local chromatin folding that creates TADs, eukaryotic genomes also exhibit long-range interactions between chromosomally distal loci that play important roles in coordinating gene expression across multiple genomic regions. In Drosophila, Polycomb response elements (PREs) and their associated Polycomb Group complexes are found at many distally-interacting loci (Li et al. 2015; Ogiyama et al. 2018; Brown et al. 2024; Gurgo et al. 2024), and transgenic insertions of PRE sequences can initiate ectopic long-distance interactions (Bantignies et al. 2003; Grimaud et al. 2006; Vazquez et al. 2006; Kwon et al. 2009). Additionally, several studies have identified Tethering Elements (TEs) that mediate long-range interactions between distant genomic loci (reviewed by Li and Levine 2024). Recent analyses have shown that a significant subset of TEs in Drosophila overlap with known PREs (Batut et al. 2022; Levo et al. 2022), highlighting that some regulatory elements may serve dual functions in both Polycomb-mediated silencing and broader chromosomal organization.
Adding to the complexity of genome organization in Drosophila and other Dipterans is the phenomenon of somatic homolog pairing, where homologous chromosomes are intimately paired from end to end in somatic cells (reviewed by Joyce et al. 2016; Peterson et al. 2021). The close juxtaposition of homologs can permit regulatory DNA to communicate in trans between the two alleles of a gene, a phenomenon known as transvection (reviewed by Duncan 2002; Kennison and Southworth 2002; Galouzis and Prud’homme 2021a). In many classical examples of transvection, a mutant allele lacking essential enhancer sequences can be complemented by pairing with a second mutant allele that retains the enhancers but carries mutations in the promoter or coding region, effectively sharing regulatory elements across the paired chromosomes to restore gene function. Several studies have also uncovered important roles for transvection underlying developmental events in otherwise wild-type genetic backgrounds (Johnston and Desplan 2014; Galouzis and Prud’homme 2021b; Antel et al. 2022), emphasizing the importance of understanding mechanisms governing trans-gene regulation in this system. However, most classical studies of transvection have focused on individual genes in isolation, leaving open the question of how long-range chromatin interactions are coordinated simultaneously across both cis and trans configurations, particularly for co-regulated genes that must respond to common developmental or environmental cues.
The Drosophila engrailed (en) and invected (inv) gene complex is one of many developmentally important gene pairs in Drosophila (Levo et al. 2022). Like other gene pairs, en and inv encode highly related proteins that share expression patterns and function (Gustavson et al. 1996). En is required for many aspects of Drosophila development, including embryonic segmentation and the formation of the posterior compartment of imaginal disks (reviewed by Joyner et al. 2024), while Inv plays a more supporting role and is not required for viability in the laboratory (Gustavson et al. 1996). Both en and inv are contained within a 113-kb domain between the genes Enhancer of Polycomb (E(Pc) and toutatis (tou)), two genes with unrelated expression patterns and functions (Fig. 1a; De et al. 2020; Jenkins et al. 2022). Transcription of inv-en is controlled by a large number of enhancers distributed throughout the inv-en domain, many of which are located upstream of the en transcription unit (Cheng et al. 2014), and by the Polycomb-group of transcriptional repressors (Moazed and O'Farrell 1992). There are four PREs in the inv-en domain, two at inv and two at en (Cunningham et al. 2010; Brown and Kassis 2013). When inv and en are not expressed, the PREs at both genes interact and the nucleosomes within the domain are modified with the repressive Polycomb chromatin mark, H3K27me3 (Brown et al. 2024). When the two genes are transcribed, part of the domain is modified with the active chromatin mark H3K27ac and the promoter-proximal PREs interact, suggesting they serve as TEs to coordinate the transcription of these two genes (Brown et al. 2024).
Trans-activation of inv is not supported in the presence of the en promoter in cis. a) schematic of the en-inv locus. The region of the red bar contains enhancer DNA required for embryonic stripes and the imaginal disk posterior compartments. Structures of wild type, invΔ33, and enΔ54 alleles are shown (red boxes, deleted regions; asterisks, PREs; “S,” “Sender” chromosome carrying intact enhancers for the imaginal disk posterior compartment and embryonic stripes but lacking the inv promoter; “R,” “Receiver” carrying an intact inv promoter but lacking the enhancers carried on the S chromosome. Line below the genomic locus shows the extent of DNA in the transgene HAen79 with an HA tag encoded on the N-terminus of the En protein. b) wild-type pattern of En and Inv staining in embryos (left) and wing imaginal disks (right). Embryo is stage 13, anterior left, dorsal up, lateral view. c) En and Inv staining in invΔ33/enΔ54 embryos (left) and wing disks (right). Inv staining is only observed in regions controlled by enhancers in cis on the R chromosome (peripheral nervous system (PNS, white asterisk), dorsal ridge (dr), posterior spiracles (PS), hindgut (hg), and clypeolabrum (cl) in embryos, and a stripe at the A/P boundary in disks (arrowhead), and not in regions controlled by enhancers in trans from the Sender chromosome. Arrows indicate a region at the wing hinge with higher Inv staining relative to En in wild-type wing disks. Embryo is late stage 13, anterior left, dorsal up, dorsolateral view. At least ten wing disks and stage 13 embryos were examined for each genotype, and a representative disk or embryo is shown.
Knowing that somatic chromosomes are paired in Drosophila, we sought to test whether enhancers that regulate en and inv when they are on the same chromosome (in cis) could also regulate these two genes when they are on homologous chromosomes (in trans). Our data show that enhancers upstream of the en transcription unit can act in trans, but only when the en cis-promoter is removed, suggesting a cis-preference that was not anticipated. Furthermore, we showed that the PRE/TEs facilitate but are not required for trans-activation, consistent with experiments that show these elements coordinate but are not required for cis-activation of paired genes (Levo et al. 2022). Finally, we demonstrated transvection between two transgenes, one carrying en and one carrying inv, which suggests the enhancers often activate only one promoter or the other, something not seen at the endogenous locus.
Materials and methods
Deletions in the endogenous inv-en domain
All new deletions were made using CRISPR/Cas9 following the procedures in https://flycrispr.org as described in Cheng et al. (2023). The coordinates for the deletions are listed in Table 1.
Genetic crosses and immunostaining
For all crosses except inv^Δ33,^ which is homozygous viable and vigorous, mutant chromosomes were balanced over either CyO, actin-GFP, or CyO, Kr-GFP. For embryo collections, the Kr-GFP chromosome was used because GFP is expressed in early embryos. For larval collections, 4–7 female virgins of one genotype were crossed to males of the other and transferred every 2–3 d. All crosses were kept at 25 °C. Ten to 12 GFP negative 3rd instar larvae were collected (with the exception of HAen45/en^Δ54^ where 6 larvae were used), dissected, and stained with antibodies as previously described (Cheng et al. 2014). Primary antibodies used were guinea pig anti-Inv (1:5000, Cheng et al. 2014), rabbit anti-EN (1:500, Santa Cruz Biotechnology, Inc), rabbit anti-GFP (1:2000, Invitrogen A11122) or mouse anti-GFP monoclonal (1:250, sc-9996). Alexa Fluor secondary antibodies (Invitrogen) were used: goat-anti-guinea pig 488 at 1:1000; goat-anti-rabbit 555 Superclonal at 1:1000 for larval stainings, 1:500 for embryo stainings; and Donkey-anti-mouse 488 at 1:500. Embryos and disks were mounted in Vectashield with DAPI (Vector Labs). For embryos, overnight collections (0–16 h) and daytime collections (0–7 h) were done, and hundreds of embryos (stages 1 to 16) were immunostained and examined.
ChIP-seq
ChIP was performed on larval brains and disks as previously described (Langlais et al. 2012). Two biological samples were taken for each genotype. ChIP-seq libraries were made and analyzed as described in De et al. (2019).
Results
To address whether transvection is supported at the en-inv locus, our general strategy was to create flies where inv transcription could only occur if enhancers act in trans. To accomplish this, we used CRISPR/Cas9 genome editing to engineer “Sender” (S) chromosomes carrying various deletions of the inv region, leaving intact the enhancers upstream of en that control en-inv expression in embryonic stripes and the posterior compartment of imaginal disks (Fig. 1a) (Gustavson et al. 1996; Cheng et al. 2014). We further engineered “Receiver” (R) chromosomes that delete the enhancer region upstream of en but leave intact the inv transcription unit. In trans-heterozygous flies carrying R/S chromosomes, transcription of inv in embryonic stripes and imaginal disk posterior compartments is only possible if the enhancers from the S chromosome activate the inv promoter on the R chromosome in trans. In some experiments, deletions on the S and R chromosomes each remove the en gene, which is required for embryonic development. In those cases, we employed a transgene, HAen79, that carries an HA-tagged functional copy of en that is inserted at the attP40 landing site on chromosome arm 2L (Fig. 1a). The 79-kb transgene includes the en transcription unit and regulatory DNA and supports development in the absence of the inv-en domain (Cheng et al. 2014, 2023; De et al. 2019) but should not participate in trans-interactions at the endogenous en-inv locus due to its distal location.
The upstream en enhancers show cis-preference for the en promoter
As a first test, we used inv^Δ33^, an S chromosome carrying a 33 kb deletion including the inv promoter and much of the transcription unit, and en^Δ54^, an R chromosome that deletes the en transcription unit and the upstream enhancers (Fig. 1a, see Table 1 for the coordinates of all deletions and transgenes). Note that inv^Δ33^ is homozygous viable and fertile, while en^Δ54^ is homozygous lethal as it lacks the en transcript and regulatory DNA. We crossed flies carrying these chromosomes to create en^Δ54^/inv^Δ33^ trans-heterozygotes and assessed Inv expression in stage 13 embryos and in third instar larval wing disks. In control wild-type organisms, both En and Inv are observed in epithelial stripes in the embryo and in the posterior compartment of the wing disk, where they are largely co-expressed except in the hinge region of the wing disk (arrow in Fig. 1b). In en^Δ54^/inv^Δ33^ trans-heterozygotes, En staining matches the pattern seen in wild type due to the intact en gene on the S chromosome (Fig. 1b). However, we see no evidence of Inv staining in embryonic stripes or in the posterior compartment of the wing (Fig. 1c). Rather, Inv staining is restricted to regions controlled by enhancers present on the en^Δ54^ chromosome, including the clypeolabrum (cl), dorsal ridge (dr), hindgut (hg), posterior spiracles (PS), and cells in the peripheral nervous system (PNS) in embryos and a stripe at the boundary of the anterior (A) and posterior (P) compartments in wing disks (arrowhead points to it in Fig. 1c). Thus, trans-activation of inv is not supported in en^Δ54^/inv^Δ33^ embryos or wing disks.
Previous analyses of transvection have shown that enhancers tend to show a preference for promoters in cis relative to promoters in trans, and that the presence of a cis-promoter can interfere with an enhancer's capacity to support transvection (Geyer et al. 1990, Martínez-Laborda et al. 1992). Although the inv promoter is deleted from the inv^Δ33^ S chromosome, the en promoter remains intact. To address whether the upstream enhancers may be restricted from acting in trans by the presence of the en promoter, we created a new S chromosome, inv^Δ62^, that removes the en promoter and transcriptional unit in addition to those of inv (Fig. 2a). As a control chromosome (C), we used en^Δ110^, which lacks the en-inv domain and therefore cannot act as either Sender or Receiver in transvection. In control HAen79 en^Δ110^/en^Δ54^ wing disks, En staining shows its expected pattern in the posterior compartment due to the presence of the HAen79 rescue construct, whereas Inv staining is entirely absent from the posterior compartment (Fig. 2b) but is expressed at a stripe at the A/P boundary as expected. In contrast, HAen79 inv^Δ62^/en^Δ54^ trans-heterozygotes show strong Inv expression in the posterior compartment, indicative of robust trans-activation of inv on the en^Δ54^ R chromosome by the enhancers on the inv^Δ62^ S chromosome (Fig. 2c).
Deletion of the en promoter in cis permits trans-activation of inv in the posterior compartment of the wing disk. a) structures of alleles at the en-inv locus (red boxes, deleted regions; asterisks, PREs; “C,” “Control” chromosome; “S,” “Sender” chromosome; “R,” “Receiver” chromosome). Note that the deletions in invΔ62 and invΔ64 each remove the entire en transcriptional unit and differ only by 1,611 bp that includes the PREs upstream of en. b–d) trans-activation of inv. In each case, the transgene HAen79 is present to provide expression of en from an ectopic location since each genotype is otherwise embryonic lethal; all En staining therefore results from expression from the transgene, whereas Inv staining results from expression at the endogenous locus. b) control genotype showing no trans-activation of inv. enΔ110 deletes all enhancers from the chromosome in trans to the intact inv promoter on the enΔ54 chromosome, resulting in Inv staining only at the A/P boundary of the wing disk (driven by an enhancer in cis to inv on the R chromosome). A representative example is shown from observations of at least 10 disks. c) trans-activation of inv by the S chromosome invΔ62. Robust staining of Inv is observed throughout the posterior compartment. d) trans-activation of inv by the S chromosome invΔ64. Inv staining is similar to that observed with invΔ62 (c), but with patches of variegated expression in some disks (arrow). Three out of nine HAen79 invΔ64/enΔ54 disks examined had patches of no Inv expression (see also Supplementary Fig. 1a). No such patches were observed in ten HAen79 invΔ62/invΔ54 wing disks.
Transvection at the en-inv locus does not require en PREs or zeste
Notably, the deletions on the R and S chromosomes inv^Δ62^ and en^Δ54^ are overlapping, indicating that there is no requirement for homology across the 113 kb en-inv locus in order to support transvection. However, the two chromosomes carry four well-characterized PREs; two on the R chromosome, with one 6 kb upstream of and one within the inv promoter, and two on the S chromosome, both upstream of the en promoter, that may facilitate enhancer-promoter trans-interactions through the ability of these PRE sequences to interact with each other (Levo et al. 2022; Brown et al. 2024). We therefore created a new S chromosome, inv^Δ64^, which is analogous to inv^Δ62^ but extends an additional 1.6 kb to remove the en PREs. In trans-heterozygous HAen79 inv^Δ64^/en^Δ54^ wing disks, we continue to see robust trans-activation of inv expression in the posterior compartment, indicating that trans PRE-PRE interactions are not necessary for enhancer action in trans at this locus (Fig. 2d). However, 3 of 9 HAen79 inv^Δ64^/en^Δ54^ wing disks had large groups of cells that lacked Inv (Fig. 2d, white arrow), and these patches of cells differed in each disk (Supplementary Fig. 1a). No such patches were seen in ten HAen79 inv^Δ62^/en^Δ54^ disks. We suggest that the lack of the en PREs decreases the ability of the enhancers to act in trans, creating a variegated pattern of expression.
Our analyses thus far have focused on the third instar larval wing disk, but the wild type pattern of larval en and inv expression includes the posterior compartments of other disk tissues. To assess whether inv-en enhancer action in trans is supported across different tissues, we also examined leg disks for Inv protein in R/S trans-heterozygotes. Control HAen79 en^Δ110^/en^Δ^^54^ leg disks show En staining in the posterior compartment due to the presence of the HAen79 construct, while Inv is seen as a discontinuous stripe at the A/P boundary (Supplementary Fig. 2a). However, as we had observed in the wing disk, HAen79, inv^Δ62^/en^Δ54^ and HAen79, inv^Δ64^/en^Δ54^ trans-heterozygotes (with and without en PREs on the S chromosome, respectively) show staining of Inv throughout the leg disk posterior compartment (Supplementary Fig. 2a), indicating that transvection at the en-inv locus is broadly supported across larval tissues.
Prior analyses of transvection have shown that trans-interactions at some loci, but not others, are modified by mutations in zeste (z) (Duncan 2002; Kennison and Southworth 2002). To assess a potential role for z in transvection at the en-inv locus, we tested two classes of z alleles; z^1^, a neomorphic allele that modifies expression of the white gene in a pairing-dependent fashion, and z^a^, a loss-of-function mutation. In either z^1^ or z^a^ mutants, we see no obvious change in Inv staining in HAen79, inv^Δ62^/en^Δ54^ wing or leg disks relative to staining in a wild-type z background (Supplementary Fig. 2b), indicating that z is unlikely to play a significant role in transvection at this locus.
en and inv genes in trans to the upstream enhancers compete for activity
Thus far, our data indicate that the presence of the en gene in cis to the upstream enhancers on the S chromosome can prevent trans-activation of inv on the R chromosome. To further explore a potential role for competition between en and inv, we created a new R chromosome, en^Δ47^, that contains an intact en transcriptional unit in addition to inv but lacks the upstream enhancers (Fig. 3a). In combination with an S chromosome, en^Δ47^ provides two potential target promoters in trans to the upstream enhancers, en and inv. In control HAen79 en^Δ110^/en^Δ47^ wing disks (enhancers deleted from both chromosomes), Inv staining is absent from the posterior compartment as expected, but present in a stripe at the A/P boundary (Fig. 3b). In HAen79 inv^Δ62^/en^Δ47^ trans-heterozygotes, Inv staining can be seen in the posterior compartment; however, the staining is reduced relative to R/S genotypes that contain a deletion of the en promoter and gene body on the R chromosome (compare to Fig. 2, c and d). In HAen79 inv^Δ64^/en^Δ47^ disks, where the en PREs are deleted from the S chromosome, Inv staining is sparse in the posterior compartment and occurs only in small patches of cells (Fig. 3e). In Fig. 3, an arrow points to the hinge region of the disk, where Inv is the predominant protein, even in wild-type disks (see Fig. 1b). When en^Δ47^ is the R chromosome, Inv is present in this region when the PREs are present on the S chromosome (Fig. 3c) and largely absent when they are not (Fig. 3e). Thus, in this case, the PREs on the S chromosome are crucial for robust transvection.
Trans-activation of inv is compromised by the presence of the en promoter on an R chromosome. a) Structures of alleles. C and S chromosomes are identical to those shown in Fig. 2. The R chromosomes enΔ47 and enΔ48 include both the inv and en transcriptional units but differ in the presence or absence of the en PREs. b) control genotype HAen79 enΔ110/enΔ47 shows no trans-activation of inv from the C chromosome. c) HAen79 invΔ62/enΔ47 trans-heterozygotes produce Inv staining in the posterior compartment that is less robust relative to Fig. 2 where the R chromosome lacks the en transcriptional unit. d) trans-activation of inv in HAen79 invΔ62/enΔ48 wing disks is greater relative to panel (c). e and f), HAen79 invΔ64/enΔ47 and HAen79 invΔ64/enΔ48 disks show patterns of Inv staining that are weaker and often variegated relative to comparable genotypes with the sender chromosome invΔ62. Arrows point to the hinge region with high Inv expression in the wild type. At least ten wing disks were examined for each genotype, and a representative disk is shown. HAen79 invΔ62/enΔ54 (Fig. 2c) and HAen79 invΔ62/enΔ47 (Fig. 3c) immunostainings were performed and imaged at the same time. The same was true for the HAen79 invΔ64/enΔ54 (Fig. 2d) and HAen79 invΔ64/enΔ47 (Fig. 3e) immunostainings; though non-quantitative, this difference in intensity between the stainings was well controlled and confirmed by two observers.
We also tested the role of the en PREs on the R chromosome using the allele en^Δ48^, which is analogous to en^Δ47^ with an additional 1.5 kb deleted that includes the en PREs. Both HAen79 inv^Δ62^/en^Δ48^ and HAen79, inv^Δ64^/en^Δ48^ trans-heterozygotes support trans-activation of inv in the posterior compartment (Fig. 3, d and f), with staining levels apparently stronger than those observed for the analogous R chromosome en^Δ47^ with intact en PREs (Fig. 3, c and e). Thus, removing the en PREs from the R chromosome improved its ability to respond to the enhancers on the S chromosome. We hypothesize that when the en PREs are in cis to the inv PREs, these PREs interact and interfere with the trans-activation of inv.
Trans-activation of en and inv is supported during embryogenesis
Thus far, our experiments have focused on the third instar larval stage and have therefore relied on the presence of the HAen79 transgenic construct to rescue the lethality of en deletions at the endogenous en-inv locus, precluding us from further assessing trans-activation of the en gene itself. To further explore transvection of en, we turned to embryos. While embryos that lack both en and inv exhibit defects in embryonic segmentation early, en^Δ47^ and en^Δ48^ have an intact en transcription unit, encode a functional En protein, and embryos that are homozygous for these alleles develop normally through at least stage 13. We suspect this is due to the presence of early stripe enhancers within the en transcription unit that provide a small but undetectable amount of En protein early in development; most of the stripe enhancers are located upstream of the en PREs (Cheng et al. 2014). No En or Inv stripes were observed in homozygous en^Δ47^ or en^Δ48^ embryos at any stage (stage 13 embryos are shown in Fig. 4b), although each protein is found in other tissues for which enhancers are intact on these chromosomes (e.g. PNS, hindgut). In R/S allelic combinations between R chromosomes en^Δ47^ and en^Δ48^ and the S chromosome inv^Δ62^, En protein is observed in many cells of the epidermal stripes beginning at stage 13 (Fig. 4c), indicating that en transcription can be activated by the stripe enhancers in trans (Fig. 4c). In contrast, no Inv staining is apparent in the epidermis of either en^Δ47^/inv^Δ62^ or en^Δ48^/inv^Δ62^ embryos, consistent with our observations in the wing disk that the presence of en on the R chromosome can reduce the trans-activation of inv (Fig. 3). Similar results were obtained using the S chromosome inv^Δ64^ which lacks the en PREs, further supporting that PRE-PRE interactions are not strictly required for transvection at the en-inv locus (Fig. 4c).
Trans-activation of en is supported in epidermal stripes of stage 13 embryos. a) structures of alleles. b) Embryos homozygous for R chromosome enΔ47 or enΔ48 show no staining of En or Inv in epidermal stripes (note that the HAen79 transgene is not carried by any embryos shown here; all en expression is from the endogenous locus). c) Variegated staining of En is observed in stripes of invΔ62/enΔ47, invΔ64/enΔ47, invΔ62/enΔ48, and invΔ64/enΔ48 embryos, with increased staining in genotypes with enΔ48 relative to enΔ47. Inv staining is generally undetectable in each genotype. All embryos are anterior left, dorsal up, lateral view, stage 13. At least 10 mutant embryos at stage 13 were examined for each genotype, and a representative embryo is shown.
To confirm that the lack of inv transvection in embryonic stripes was due to the presence of en on the R chromosome, we once again employed en^Δ54^, which deletes the upstream enhancers and the en transcriptional unit (Fig. 5). In control embryos of en^Δ54^ in combination with en^Δ110^ (the deletion of the en-inv locus), no staining of Inv is observed in epidermal stripes at any stage (stages 11 and 15 are shown) due to the deletion of the upstream enhancers from both chromosomes. In comparison to wild-type embryos, the en^Δ54^/en^Δ110^ control embryos lacking En on both chromosomes have abnormal development, including no head involution, making late staging hard. We therefore used the HAen79 rescue construct and assessed trans-activation of inv in HAen79 inv^Δ62^/en^Δ54^ embryos that have normal morphology. In HAen79 inv^Δ62^/en^Δ54^ embryos, Inv staining is present in cells of the epidermal stripes at stages 13 and 15, indicating that trans-activation of inv is supported by the stripe enhancers on the S chromosome. Notably, we do not see evidence of Inv staining in the epidermal stripes of stage 11 embryos, indicating that transvection at the en-inv locus is not yet supported at this time of development.
Trans-activation of inv in epidermal stripes. a) structures of alleles. b) The genotype of the embryos in each row is listed on the left, and the stage of the embryos in each column is listed at the bottom. Top row, In control enΔ54/enΔ110 embryos (without the HAen79 rescue construct), Inv protein is present in tissues controlled by enhancers present on the R chromosome but is absent from the epidermal stripes. Middle row, in wild type embryos (wt), Inv protein is present in the epidermal stripes at all stages shown. Bottom row, trans-activation of inv in the epidermal stripes of HAen79 invΔ62/enΔ54 is observed at stages 13 and 15 but absent at stage 11. All embryos are anterior left, dorsal up. Asterisk denotes autofluorescence in the yolk. Stage 11 enΔ54/enΔ110 and HAen79 invΔ62/enΔ54 embryos are lateral, and wt embryos are ventral-lateral. wt stage 13 embryo is a lateral view; HAen79 invΔ62/enΔ54 is a dorsolateral view. Stage 15 embryos are dorsolateral views. At least 10 embryos of each stage shown were examined, and a representative embryo is shown.
Trans-activation of inv is supported between transgenic insertions
As a final test of the capacity for the en-inv locus to support transvection, we assessed whether transgenes carrying elements of the locus at an ectopic location are sufficient to allow trans-activation of inv in the posterior compartment of the wing disk using the same logic of “Sender” and “Receiver” elements. The HAen79 transgene used throughout this study is analogous to the S chromosome inv^Δ33^ in that it contains the complete en transcriptional unit and upstream enhancers but lacks the inv promoter and part of the inv transcription unit (Fig. 6a). In addition, a second Sender transgene, HAen45 (also inserted at the attP40 landing site), encodes a truncation of the HAen79 transgene that includes the complete en transcriptional unit but removes regulatory DNA from upstream and downstream enhancer regions (Fig. 6a, Table 1). Finally, the Receiver transgene HAinv84 is analogous to the R chromosome en^Δ54^, encoding the complete inv promoter and transcriptional unit but lacking the en gene and its upstream enhancers (Fig. 6a). Inv expression from the HAinv84 transgene is similar to that seen from en^Δ54^ including no stripes in embryos and a stripe at the A/P boundary in disks (Cheng et al. 2014). As stated above, inv and en are regulated by the Polycomb group genes. In cells where these two genes are not transcribed, the Polycomb repressive chromatin mark H3K27me3 covers the locus from the 3′ end of the e(Pc) gene to the end of the 3′ end of the tou gene. Likewise, the HAen79 transgene is also covered by H3K27me3 (De et al. 2019). We wondered whether the HAinv84 transgene could also form an H3K27me3 domain. To assess this, we performed chromatin-immunoprecipitation followed by next-generation sequencing (ChIP-seq) on third instar larval brains and disks of the genotype HAinv84@attP40 inv^Δ45^. The inv^Δ45^ allele deletes the entire inv transcription unit and some regulatory DNA (Fig. 6a), and ChIP-seq shows H3K27me3 over the inv-en domain except in the region of the deletion (Fig. 6b). In contrast, H3K27me3 is present throughout the domain in HAinv84 inv^∆45*^*larvae, with the middle signal coming from the HAinv84 transgene. Thus, the HAinv84 transgene can also form an H3K27me3 domain, likely initiated by the inv PREs.
Transvection studies using large transgenes. a) schematic of transgenes used. Bars above show the region encoded on Sender transgenic insertions HAen79 and HAen45, whereas the bar below shows the region encoded by Receiver transgene HAinv84. Schematics of alleles invΔ33 and enΔ54 are shown to highlight that the breakpoints of the deletions correspond to the endpoints of the transgenes. invΔ45 deletes the entire inv transcription unit and some flanking DNA. b) ChIP-seq showing the en-inv region on invΔ45 (upper) and HAinv84 invΔ45 third instar brains and disks, Grey-input DNA, Blue-H3K27me3. Note that the H3K27me3 accumulation in the region of the invΔ45 deletion (red box) in the HAinv84 invΔ45 genotype is coming from HAinv84 transgene. The black line below is the extent of the HAinv84 transgene; regions flanking the invΔ45 deletion have higher levels of H3K27me3 because of signal from both the transgene and the endogenous locus. All tracks are 0–5. c) Inv is present at the A/P boundary in HAen79 enΔ110/HAinv84 enΔ110 wing disks, showing there is no trans-activation of inv in the genotype. A representative example is shown from a total of 10 disks observed.
We first examined En and Inv staining in HAen79 en^Δ110^/HAinv84 en^Δ110^, where 110 kb of the en-inv region is deleted from the endogenous locus on both homologs, and the transgenes are therefore the only potential sources of En and Inv proteins. This experiment is analogous to our initial assessment of transvection in en^Δ54^/inv^Δ33^ trans-heterozygotes, where no transvection was evident; as expected, we once again observed no evidence of inv trans-activation in this transgenic combination (Fig. 6c). We then examined the ability of the much smaller HAen45 transgene to work as a Sender chromosome for transvection. We attempted to assess En and Inv staining in control HAen45 en^Δ110^/en^Δ110^ wing disks, but this genotype fails to develop sufficient disk tissue due to incomplete rescue from the truncated HAen45 transgene (Cheng et al. 2014). However, replacing one of the en^Δ110^ alleles with the smaller en^Δ54^ deletion (HAen45 en^Δ110^/en^Δ54^) produced somewhat deformed but recognizable wing disks where En staining was found throughout the posterior compartment and Inv staining was evident only at the A/P boundary, as expected (Fig. 7a). Finally, we assessed the potential for trans-activation of inv by the Sender transgene HAen45 by placing it in trans to HAinv84, with both inserted at the attP40 site on chromosomal 2L. In this case, HAen45 en^Δ110^/HAinv84 en^Δ110^ (homozygous for the large en^Δ110^ deletion) organisms develop into healthy third instar larvae with robust wing disk tissue. In these disks, both En and Inv proteins are observed in the posterior compartment, indicating that transcription of inv encoded on the HAinv84 transgene is activated in trans by the enhancers encoded on the HAen45 transgene (Fig. 7b). Notably, although some cells show evidence of both proteins, in most cells, only En (produced in cis) or Inv (produced in trans) is seen, resulting in a reciprocal pattern of staining across the wing disk rather than co-expression (Fig. 7b). A similar reciprocal pattern was observed in all 11 disks of this genotype examined, with some variation in the exact staining pattern (see two additional examples in Supplementary Fig. 1b), and therefore appears to be a consistent feature of transvection between the HAen45 and HAinv84 transgenes. The reciprocal pattern of expression was surprising to us since En and Inv are usually co-expressed, exist in a gene complex, and are Polycomb-regulated. We wondered whether a reciprocal expression of En and Inv also occurred in a different genetic background. Specifically, in en^1^ mutants, wing development is disrupted, and staining of wing disks with an antibody that recognized both Inv and En showed variegated expression (Brower 1986), but it was not known whether these two proteins were co-expressed. We therefore stained en^1^ homozygous larval disks with anti-Inv and anti-En antibodies. Unlike En and Inv in HAen45 en^Δ110^/HAinv84 en^Δ110^ wing disks, En and Inv proteins are completely co-expressed in en^1^ wing disks, in variegated patterns that vary in each disk (Fig. 7c, see Supplementary Fig. 3 for more info about en^1^). In sum, transgenes carrying sequences from the en-inv locus are sufficient to support transvection at an ectopic locus, with enhancer action supported either in cis or in trans.
Trans-activation of inv is supported by the transgene HAen45. a) HAen45 modestly rescues enΔ110/enΔ54 embryos that entirely lack en at the endogenous locus. Wing disks of this genotype are slightly misshapen, with strong En staining throughout the posterior compartment but Inv staining primarily at the A/P boundary. Six disks were examined, and all had the same pattern of En and Inv expression. b) trans-activation of inv in HAen45/HAinv84 trans-heterozygotes homozygous for enΔ110. Both En and Inv staining are evident in the posterior compartment, with many cells activating only one gene or the other. The dashed white box highlighted in the merge disk picture surrounds the region of the disk enlarged in the pictures below. Eleven disks were examined, and all had reciprocal patterns of En and Inv expression. c) on the en1 chromosome, a blood retrotransposon (triangle) is inserted approximately 13 kb upstream of the en transcription start site (Cheng et al. 2014). En an Inv are variegated and co-expressed in an en1 wing disk. The dashed white box in the merge disk picture surrounds the region of the disk enlarged in the pictures below. Ten disks all showed En-Inv co-expression in this genotype.
Discussion
Transvection has been well documented at many Drosophila loci, but its regulation at complex co-regulated developmental gene loci remains poorly explored. Here, we investigated whether transvection is supported at the en-inv locus, where the en and inv genes are co-regulated by shared upstream enhancers during embryonic segmentation and imaginal disk development (Fig. 8a). The co-expression of these genes is aided by promoter-proximal PREs, which interact with each other and enhancers to facilitate gene expression. Using a CRISPR-based approach to create Sender (S) chromosomes containing regulatory DNA, and Receiver chromosomes (R) containing the Inv transcription unit, we tested the ability of embryonic stripe enhancers and imaginal disk enhancers to act in trans (Fig. 8b). In the presence of the en transcription unit on the S chromosome, the enhancers could not activate the inv promoter (Fig. 8b, top). When the en promoter was removed (Fig. 8b, middle), strong trans-activation of inv occurred. Note that there is no homology between the S and R chromosomes and that the PREs can facilitate interactions between the two promoters and the enhancer in this configuration. Finally, upon deletion of the en PREs (Fig. 8c, bottom), the trans-activation is less robust, leading to a variegated pattern of expression.
Summary model. a) cis expression. The inv and en transcription units are shown with an arrow showing the start and direction of transcription. The stripe or imaginal disk enhancers are shown by an E in a blue box. In the upper linear model, the enhancers can activate both the en and inv promoters. In the lower model, the promoter-proximal en and inv PREs interact with each other and the enhancers to bring the DNAs together and facilitate transcription (Levo et al. 2022; Brown et al. 2024). b) Trans expression. The striped boxes indicate DNA deleted from the inv-en locus in our transvection assays. The upper panel shows that, in the presence of the en transcription unit, the enhancers cannot activate the promoter on the homologous chromosome (indicated by XXX interrupting the arrow). The middle panel shows that when the en promoter and transcription unit is removed, the enhancer can now activate the inv promoter. In this case, the PREs are present and could act together to facilitate transcription. Bottom panel, when the en PREs are not present on the S chromosome, trans-activation is less robust leading to variegated expression.
We were surprised that the en upstream enhancers were unable to trans-activate inv in the presence of an en promoter in cis. Although preference for a cis-promoter has been well documented in transvection at other loci (Martínez-Laborda et al. 1992; Casares et al. 1997; Gohl et al. 2008; Bateman et al. 2012; Mellert and Truman 2012; Lim et al. 2018; Piwko et al. 2019), for the majority of genes tested, weak activation of a promoter in trans is still permitted in the presence of a promoter in cis. To the best of our knowledge, the yellow gene is the only other case that shows a complete lack of observable transvection in the presence of a cis-promoter (Geyer et al. 1990; Morris et al. 1999), although transgenic studies have shown this to be position-dependent (Kravchuk et al. 2016). At the en-inv locus, the upstream enhancers normally activate both en and inv promoters simultaneously, and micrococcal nuclease and chromatin capture (micro-C) analysis has shown that the promoter-proximal PREs of en and inv are physically looped together (Levo et al. 2022; Brown et al. 2024), which likely facilitates long-range clustering of the two promoters with the upstream enhancers. We hypothesized that this PRE-dependent support of long-range promoter clustering would extend to interactions in both cis and trans, thereby facilitating trans-activation of inv even in the presence of the en promoter, but it must instead be the case that interactions in cis are preferred over interactions in trans. This cis-preference could simply be because the inv promoter is too far away to be activated when it is on the homologous chromosome. Note that in our experiments, there is transvection between the S and R chromosomes that share no homology. It would be interesting to do similar experiments using small deletions or mutations of the en and inv promoters, leaving the two chromosomes otherwise homologous. In support of a role for PRE-PRE interactions in facilitating transvection, loss of the en proximal PREs on the S chromosome in HAen79 inv^Δ64^/en^Δ54^ and HAen79 inv^Δ64^/en^Δ47^ larvae results in less consistent trans-activation of inv relative to the genotypes HAen79 inv^Δ62^/en^Δ54^ and HAen79 inv^Δ62^/en^Δ47^, where the en PREs are still present on the S chromosome. Conversely, when the en promoter and its proximal PREs are present on the R chromosome in en^Δ47^, trans-activation of inv is reduced relative to the R chromosome en^Δ48^ without the en PREs, in this case we hypothesize that the en and inv PREs in cis on the en^Δ47^ R chromosome propagate a very stable “OFF” state, while the loss of the en PREs on en^Δ48^ destabilizes this state allowing trans-activation of inv.
Our analysis of developmental timing of inv transvection is consistent with our understanding of the onset of somatic homolog pairing. Maternal and paternal homologs are largely unpaired through the early syncytial divisions, and pairing increases dramatically beginning in cell cycle 14, with many loci plateauing by stage 13–15 (Fung et al. 1998; Gemkow et al. 1998; Child et al. 2021). It is therefore likely that homolog pairing has not progressed sufficiently by stage 11 to support transvection at the en-inv locus, whereas trans-interactions between homologs have stabilized by stage 13, supporting transvection. Similarly, prior analyses of transvection at other loci have demonstrated transvection in embryos at stage 13 and later (Hendrickson and Sakonju 1995; Sipos et al. 1998; Fujioka et al. 2016), with no evidence of pregastrulation transvection in the absence of the transgenes carrying the gypsy insulator (Lim et al. 2018).
While classic examples of enhancer action in trans in Drosophila typically involve trans-heterozygous allelic combinations of endogenous genes, transgenes that are inserted at common positions on homologous chromosomes can also support this form of transvection. This was first observed using transgenes carrying components of the yellow gene (Chen et al. 2002) and later with synthetic fluorescent reporter genes (Bateman et al. 2012; Mellert and Truman 2012). We were surprised to find that the Sender transgene HAen45 supported trans-activation of inv on the Receiver transgene HAinv84 whereas the Sender HAen79 did not. Both HAen79 and HAen45 carry a complete en transcriptional unit in cis to the upstream enhancers; therefore, we expected both elements to be restricted from acting in trans. HAen79 contains 17 kb more DNA on both the distal and proximal ends relative to the HAen45 transgene, and we propose that those sequences somehow “lock in” the cis-promoter preference of the posterior compartment imaginal disk enhancers.
We were further surprised by the patterns of expression observed for en (activated in cis) and inv (activated in trans) in the posterior compartment of HAen45 en^Δ110^/HAinv84 en^Δ110^ wing disks undergoing transvection, with many cells activating only one or the other gene. Previous analyses of transvection using fluorescent reporters have instead shown that an enhancer will typically co-activate promoters in cis and in trans in the same cells (Bateman et al. 2012; Mellert and Truman 2012; Lim et al. 2018) with one exception (Kassis 2012 ; Mellert and Truman 2012). Furthermore, the relatively large regions of tissue showing expression of one gene or the other suggest that a decision on which gene to activate (or repress) was made in earlier precursor cells and then maintained in clonal patches as the cells of the disk underwent mitosis. Overexpressing En can lead to its silencing in a variegated manner in wing disks (Guillén et al. 1995; Tabata et al. 1995), producing variegated expression patterns similar to those seen in en^1^ homozygous disks. In addition, both HAen79 and HAen45 can be repressed by En produced by the endogenous en-inv domain. This regulation is likely to be direct as En is a repressor, binds to one of the imaginal disk enhancers (IDE), and can repress expression of an IDE-reporter transgene (Cheng et al. 2023). While we have no evidence that En produced from HAen45 can repress the expression from HAinv84 (or vice versa), the reciprocal pattern of expression common in HAen45 en^Δ110^/HAinv84 en^Δ110^ wing disks could, in theory, involve feedback by En and/or Inv proteins, repressing expression of one transgene or the other. To this end, it would be interesting to test whether en and inv genes carrying loss-of-function point mutations would show a similar pattern of expression in paired transgenes, although the complication of embryonic lethality for en loss-of-function mutations would present a challenge.
Three-dimensional analysis of Drosophila genome structure using micro-C has identified a class of elements, called tethering elements (TEs), that facilitate interactions between regulatory DNA, including enhancer-promoter and promoter-promoter interactions (reviewed in Li and Levine 2024). In one study (Levo et al. 2022), more than 20 co-regulated developmental gene pairs, including en-inv, were identified that contain paired TEs located near their promoters. Removal of a TE from one gene in a pair uncoupled expression of the two genes, and led to stochastic expression of the gene without the TE (Levo et al 2022). The promoter-proximal en PRE can function as a TE for the imaginal disk enhancers (Kwon et al 2009), and the PRE at the inv promoter can pair with a presumptive enhancer in micro-C experiments (Brown et al. 2024), suggesting it is also a TE. We suggest that many of the TEs identified in gene pairs may facilitate both cis- and trans-regulatory interactions.
Supplementary Material
iyaf276_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Acemel RD, Lupiáñez DG. 2023. Evolution of 3D chromatin organization at different scales. Curr Opin Genet Dev. 78:102019. 10.1016/j.gde.2022.102019.36603519 · doi ↗ · pubmed ↗
- 2Antel M et al 2022. Interchromosomal interaction of homologous Stat 92E alleles regulates transcriptional switch during stem-cell differentiation. Nat Commun. 13:3981. 10.1038/s 41467-022-31737-y.35810185 PMC 9271046 · doi ↗ · pubmed ↗
- 3Bantignies F, Grimaud C, Lavrov S, Gabut M, Cavalli G. 2003. Inheritance of Polycomb-dependent chromosomal interactions in Drosophila. Genes Dev. 17:2406–2420. 10.1101/gad.269503.14522946 PMC 218078 · doi ↗ · pubmed ↗
- 4Bateman JR, Johnson JE, Locke MN. 2012. Comparing enhancer action in cis and in trans. Genetics. 191:1143–1155. 10.1534/genetics.112.140954.22649083 PMC 3415998 · doi ↗ · pubmed ↗
- 5Batut PJ et al 2022. Genome organization controls transcriptional dynamics during development. Science. 375:566–570. 10.1126/science.abi 7178.35113722 PMC 10368186 · doi ↗ · pubmed ↗
- 6Bhattacharya M, Lyda SF, Lei EP. 2024. Chromatin insulator mechanisms ensure accurate gene expression by controlling overall 3D genome organization. Curr Opin Genet Dev. 87:102208. 10.1016/j.gde.2024.102208.38810546 PMC 11317221 · doi ↗ · pubmed ↗
- 7Brower DL . 1986. Engrailed gene expression in Drosophila imaginal discs. EMBO J. 5:2649–2656. 10.1002/j.1460-2075.1986.tb 04547.x.3536480 PMC 1167165 · doi ↗ · pubmed ↗
- 8Brown JL, Kassis JA. 2013. Architectural and functional diversity of Polycomb group response elements in Drosophila. Genetics. 195:407–419. 10.1534/genetics.113.153247.23934890 PMC 3781969 · doi ↗ · pubmed ↗