Chlamydia trachomatis-specific T Cell Immunity Reflects Widespread Exposure in South African Adolescents and Young Women
Rubina Bunjun, Micaela Lurie, Smritee Dabee, Shaun Barnabas, Venessa Maseko, Shameem Z Jaumdally, Hoyam Gamieldien, David A Lewis, Heather B Jaspan, Katherine Gill, Linda-Gail Bekker, Jo-Ann S Passmore

TL;DR
This study shows that T cell responses to Chlamydia trachomatis in South African adolescent girls and young women are linked to infection status and inflammation, offering insights for vaccine development.
Contribution
The study identifies how T cell immunity correlates with infection outcomes and inflammation in a high-risk population.
Findings
C. trachomatis-specific CD4+ T cell responses were lower in untreated/recurrent infections compared to cleared or primary infections.
Systemic Th1 responses were inversely correlated with genital inflammation, suggesting a protective role.
Polyfunctional Th1 cells were nearly absent in untreated infections, highlighting their importance in immunity.
Abstract
Chlamydia trachomatis remains the most prevalent bacterial sexually transmitted infection worldwide, disproportionately affecting adolescent girls and young women (AGYW). Vaccine development is hindered by a limited understanding of protective immunity, particularly in the context of multiple exposures and immunopathology. We characterized mucosal inflammation and systemic immune responses to C. trachomatis in South African AGYW (n = 145), stratified by exposure history based on nucleic acid amplification testing (NAAT) and serology (Ab). Specifically, cervicovaginal cytokines, cervical T cell activation, and C. trachomatis-specific CD4+ T cell responses were assessed. A NAAT+/Ab + status, signifying untreated/recurrent infection, was associated with increased cervical T cell activation. These women all had detectable C. trachomatis-specific CD4+ T cells in blood; however, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
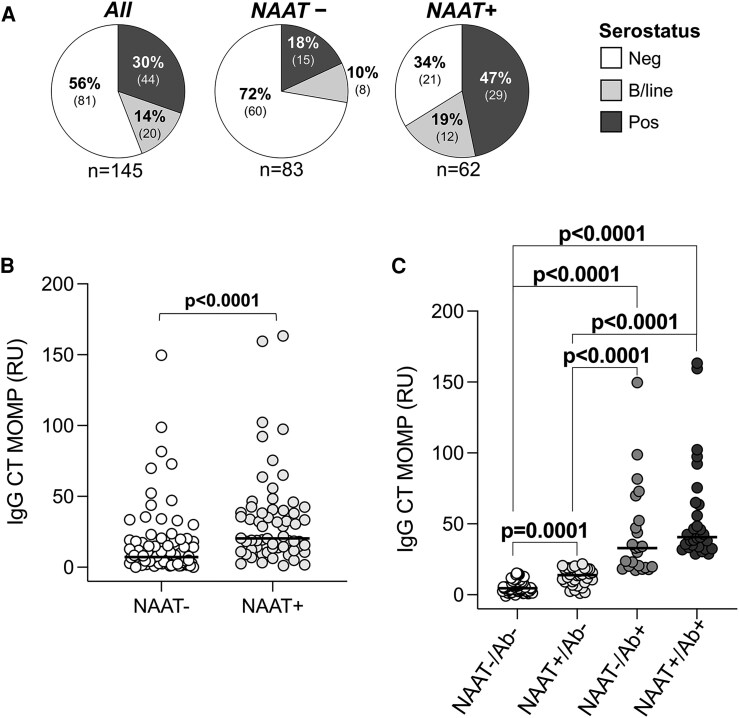 Figure 1
Figure 1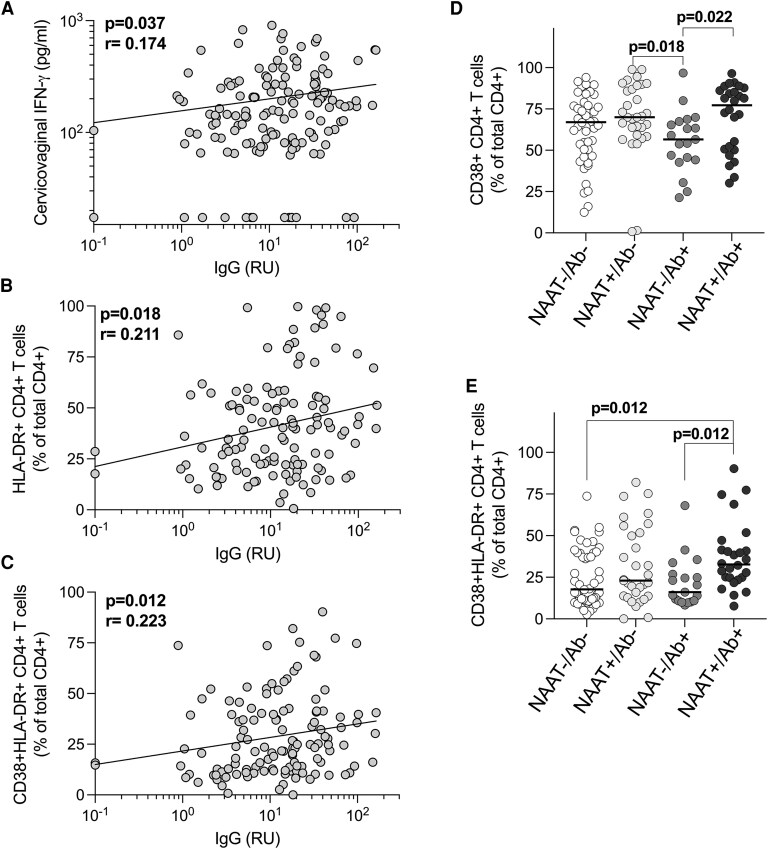 Figure 2
Figure 2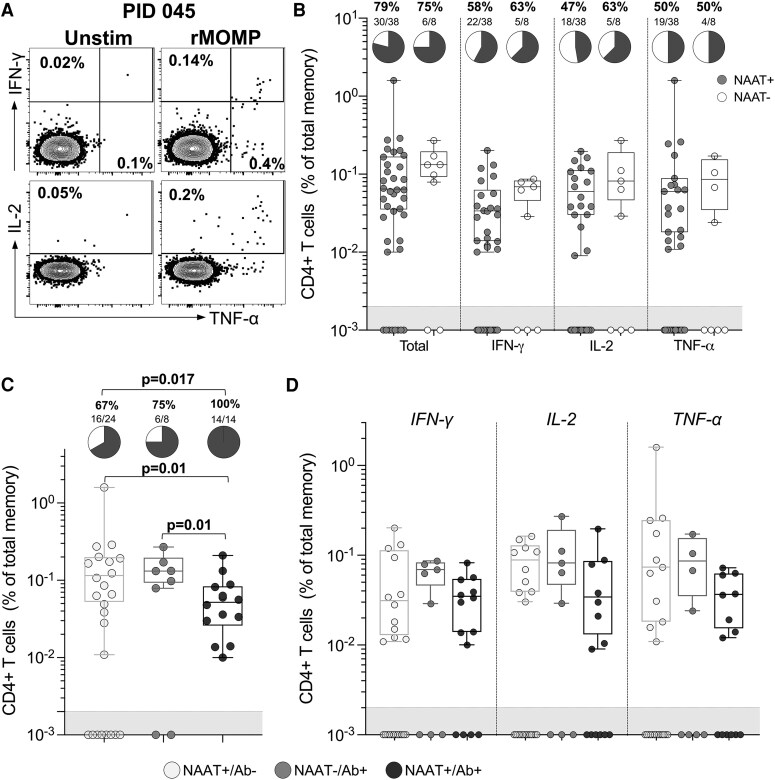 Figure 3
Figure 3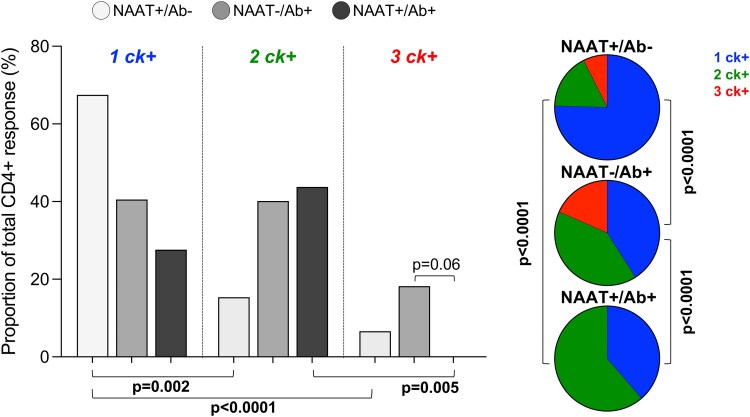 Figure 4
Figure 4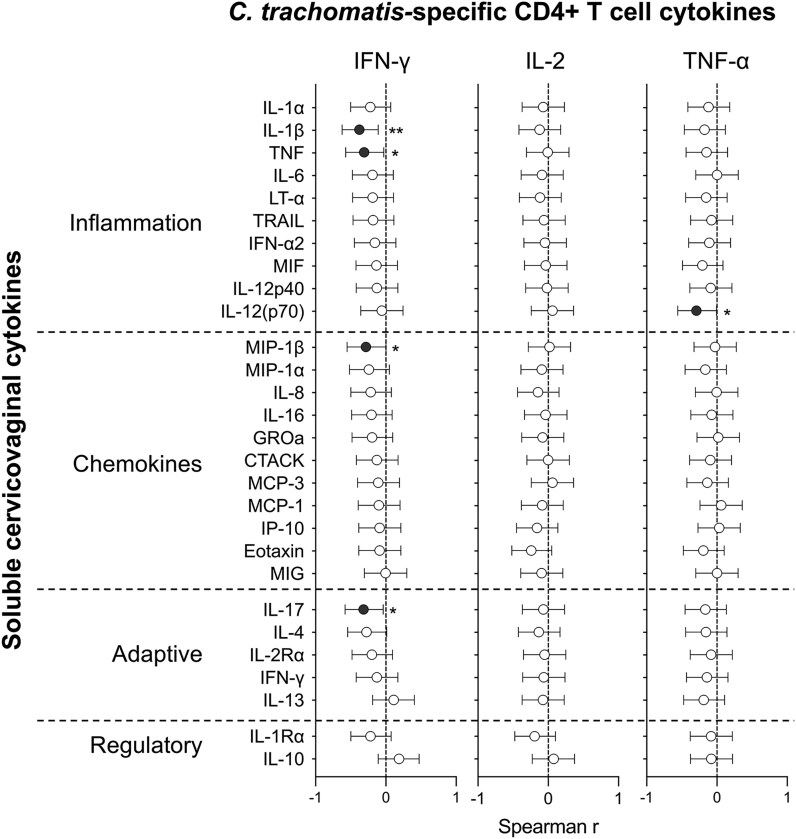 Figure 5
Figure 5| Characteristic | N = 145 |
|---|---|
| Age in y [median (IQR)] | 18 (17–20) |
| Age of sexual debut (y) [median (IQR)] | 16 (15–17) |
| Lifetime sexual partners [median (range)] | 2 (1–13) |
| Multiple sexual partners in the last 12 m [n/N(%)] | 33/111 (30%) |
| Contraceptive use [n/N (%)] | |
| Ever pregnant | 30/122 (25%) |
| Previous contraceptive use | 96/122 (79%) |
| Condom use during the last sex act | 87/121 (72%) |
| Regular condom use in the last 12 m | 42/122 (34%) |
| Hormonal contraceptives | |
| DMPA-IM | 27/145 (19%) |
| Net-En | 99/145 (68%) |
| Combined oral contraceptive | 7/145 (5%) |
| LNG implant | 9/145 (6%) |
| Nuvaring | 1/145 (0.7%) |
| Sexually transmitted infections (STIs) [n/N (%)] | |
|
| 62/145 (43%) |
|
| 17/145 (12%) |
|
| 6/145 (4%) |
|
| 11/145 (8%) |
| Bacterial vaginosis (Nugent score 7–10) | 70/145 (48%) |
| Previous STI (diagnosed or treated) | 17/122 (14%) |
- —European and Developing Countries Clinical Trials10.13039/501100001713
- —Fondation Botnar10.13039/501100011318
- —European Union10.13039/501100000780
- —South African Medical Research Council10.13039/501100001322
- —Desmond Tutu Health Foundation10.13039/501100005860
- —National Research Foundation10.13039/501100001321
- —Poliomyelitis Research Foundation10.13039/501100001323
- —HIV Vaccine Trials Network10.13039/100029382
- —Fogarty Foundation10.13039/100008096
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive tract infections research · Cervical Cancer and HPV Research · Adolescent Sexual and Reproductive Health
Chlamydia trachomatis is the most prevalent bacterial sexually transmitted infection (STI) globally. In 2020, the World Health Organization (WHO) estimated that there were more than 120 million new infections worldwide [1]. Adolescent girls and young women (AGYW), in particular, experience a high chlamydia burden [2]. Although the interruption in sexual healthcare delivery caused by the COVID-19 pandemic contributed to an increase in infections [3], chlamydia prevalence was already increasing in 2019 [4, 5]. Despite the availability of effective antibiotic treatment, many infections remain unnoticed and untreated [6], particularly in low-and-middle income countries (LMICs), which rely on the syndromic management for STIs [7, 8]. The WHO has set a target of a 50% reduction in new cases of chlamydia by 2030 [1]. Achieving this objective requires a prevention-oriented strategy, with an emphasis on vaccines [9].
Chlamydia vaccine development has been hindered by several immune-related obstacles [10, 11]. Live-attenuated vaccines carry the risk of inducing genital tract pathology [12], which is associated with pelvic inflammatory disease and subsequent infertility [13, 14] and may increase the risk of HIV-1 acquisition [15]. Therefore, an effective vaccine needs to induce a protective immune response without the accompanying mucosal immunopathology. Furthermore, determining correlates of protection is complicated by the natural history of C. trachomatis infection. Untreated or recurrent infections are common in young women but decrease with age, suggesting some immune protection after exposure, although incomplete [16–19].
CD4+ T cells play a key role in immunity against chlamydia, primarily due to their production of Th1 cytokines [20–23]. Clinical studies have focused on antigen mapping, with limited data characterizing the T cell response to C. trachomatis, especially in high-risk young women or in relation to exposure history [24–26]. Consequently, there is an urgent need to better understand the development of immunity while also considering the development of immune-mediated pathology due to multiple exposures. We characterized genital tract inflammation and systemic CD4+ T cell responses to C. trachomatis in South African AGYW, stratified by their infection history and relative exposure levels, to better understand immune correlates of prior cleared infections, and the effect of continuous pathogen exposure.
METHODS
Study Participants
AGYW (16–22 years old) from South Africa were enrolled in the Women's Initiative in Sexual Health (WISH) study and provided samples every 2 months for 6 months [27, 28]. A total of n = 149 AGYW completed the baseline study visit, 127/149 completed 2 visits, and 88/149 completed 3. Participants were excluded if they were living with HIV, pregnant, menstruating at the time of sampling, or if they were sexually active, douched, or used spermicides in the last 2 days or took antibiotics in the last 2 weeks. The Human Research Ethics Committee of the University of Cape Town (267/2013) approved the study.
Study Procedures
Samples were collected at each study visit [28, 29]. A vulvovaginal swab was collected for STI testing, and cervicovaginal secretions (CVSs) were collected through insertion of a menstrual cup (Softcup; The Flex Company, USA) into the vagina for 30 minutes. Participants were tested for STIs at every visit. Specimens positive for C. trachomatis were also tested for lymphogranuloma venereum (LGV)-associated L1-L3 genovars. All participants were negative for LGV. Participants who had signs/symptoms of STIs or who tested positive were offered treatment and a partner referral letter.
Sample Processing
Peripheral blood mononuclear cells (PBMC) and plasma were isolated by density gradient centrifugation with Histopaque (Sigma Aldrich, USA) within 4 hours of collection. Cells were cryopreserved in fetal calf serum (HyClone; Cytiva, USA) with 10% DMSO (Sigma-Adrich, USA) and stored in liquid nitrogen until use. Plasma was aliquoted and stored at −80°C. CVSs were removed from the menstrual cup as previously described [28, 30], diluted 1:5 with phosphate-buffered saline (PBS; Sigma Aldrich, USA), aliquoted, and stored at −80°C.
Stimulation of T Cells
We selected participants with a positive C. trachomatis nucleic acid amplification test (NAAT) or who were seropositive for C. trachomatis at any visit during the 6-month study period, where samples were available (n = 46). Cryopreserved PBMC from the baseline visit were thawed and rested in R10 medium (Gibco RPMI 1640; Thermo Fisher Scientific, USA), with the addition of 10% heat-inactivated FCS (HyClone; Cytiva, USA) and 50 U/mL of penicillin-streptomycin (Sigma-Aldrich, USA) for 4 hours prior to stimulation. Cells were incubated in R10 with rMOMP protein (10μg/mL; Uniprot Accession #P17451.1, ProMab, USA) for 18 hours at 37°C in the presence of the costimulatory antibodies anti-CD28 and anti-CD49d (1μg/mL each; BD Biosciences, USA) and Brefeldin A (BFA; 5μg/mL, Sigma-Aldrich).
Multiparameter Flow Cytometry
Stimulated PBMC were stained with a violet viability dye (ViViD; Molecular Probes, USA), followed by staining with CD14-Pacific blue, CD19-Pacific blue, CD4-PE-Cy5.5 (all Invitrogen, USA), CD8-BV711 (Biolegend, USA), CD27-PE-Cy-5 (Beckman Coulter, USA), and CD45RO-BV785 (Biolegend, USA). Cells were permeabilized with Cytofix-Cytoperm (BD Biosciences, USA), stained intracellularly with CD3-APC-H7, IFN-γ-Alexa700, IL-2-APC (all from BD Biosciences, USA), and TNF-α-PE-Cy7 (Biolegend, USA), and fixed with CellFix (BD Biosciences, USA). Cells were acquired on a BD LSRII (BD Biosciences, USA), using FACSDiva software. Data were analyzed using FlowJo v10 (TreeStar, BD Biosciences, USA) and Pestle and Spice [31]. The gating strategy is shown in Supplementary Figure 1. A positive cytokine response was defined as at least double the background frequency of the unstimulated sample and a net response >0.005%. All data are reported after background subtraction.
Measurement of C. trachomatis-specific IgG by ELISA
Baseline visit plasma from 145 women was used to measure IgG using a semiquantitative ELISA kit licensed for diagnostic use with 100% sensitivity and 97% specificity (Cat#EI2191-9601G; EuroImmun; Germany). Plasma was diluted and incubated on the supplied precoated plate. The plate was washed and incubated with peroxidase-labelled anti-IgG, followed by a final incubation with tetramethylbenzidine (TMB) substrate. Stop solution was added, and color intensity was measured at 450 and 650 nm (reference) within 15 minutes. A standard curve was used to calculate the relative units (RU) of IgG. Results of >22 RU/mL were positive, 16–22 RU/mL were borderline, and <16 RU/mL were considered negative. Trachoma (serovars A-C) is not endemic to South Africa, and all participants were negative for LGV, suggesting the IgG detected arose from C. trachomatis serovar D-K infections. Similarly, cross-reactive antibodies generated by C. pneumoniae are unlikely due to their rarity in South Africa [32, 33]. While the derivative serovar(s) of the rMOMP used in the kit are not disclosed, the manufacturer states that the ELISA detects all serovars of human C. trachomatis and is not cross-reactive for human infection with C. pneumoniae or C. psittaci.
Cervicovaginal Cytokine Assays
Diluted CVSs were thawed and filtered through a 0.2μm cellulose acetate filter. The Human Cytokine Group I 27-plex and Human Cytokine 21-plex kits (Bio-Rad, USA) were used to quantify a range of 44 cytokines, chemokines, and growth factors. Values below the detection limit were recorded as half of the lowest measured concentration for each cytokine. IL-2, IL-5, IL-15, and RANTES were excluded for all analyses as the values did not pass QC or >50% of values were below the detection limit.
Statistical Analyses
Statistical analyses were performed using Prism 9 (GraphPad, USA). Nonparametric tests (Mann-Whitney U, Wilcoxon matched pairs, and Spearman Rank) were used for comparisons. Fischer's exact test was used to compare proportions of responders. A P value of <.05 was considered statistically significant.
RESULTS
Cohort Description
We used baseline samples from a well-characterized cohort of HIV-uninfected AGYW (median age of 18 years old) from South Africa [27, 28]. Participants were recruited within 3 years of sexual debut and reported a median of 2 sexual partners (Table 1). Hormonal contraceptive use was high (n = 143/145), but only 34% (42/122) of participants reported regular condom use within the past year. Importantly, a high prevalence of chlamydia (43%) was found, with low rates of previous STI diagnosis or treatment (14%), highlighting the need for better sexual health interventions.
Classification of C. trachomatis Infection History and Exposure Levels Using Nucleic Acid Amplification Testing and Serology
We used anti-C. trachomatis IgG serology to infer infection history. Thirty percent (30%; 44/145) of participants were seropositive, 14% (20/145) were borderline positive, and 56% (81/145) were seronegative (Supplementary Table 1). Of those with a negative nucleic acid amplification test (NAAT), 72% (60/83) were also seronegative (Figure 1A). Similarly, 66% (41/62) of NAAT + AGYW were also seropositive. Notably, 34% (21/62) NAAT + AGYW were seronegative, likely signifying a primary infection. Women who were NAAT + also had nearly 3-fold higher antibody levels than women who were NAAT− (Figure 1B; P < .0001; medians: 20.35RU and 7.14RU, respectively), suggesting higher antigen load due to an active infection [34]. Given the low prevalence of previous STIs, and the asymptomatic nature of infections in this cohort, it is probable that NAAT + AGYW were undiagnosed and untreated, rather than having treatment failure (persistence) or repeated infections (recurring).
Repeated or persistent C. trachomatis infections were common in adolescent girls and young women (n = 145). (A) Plasma serology results from the cohort were measured using semi-quantitative ELISA (Euroimmun; Germany). Relative units (RU) were used to classify participants into negative (<16 RU; neg; white), borderline positive (16–21 RU; b/line; light grey), and positive (≥22 RU; pos; dark grey). (B) Distribution of IgG levels measured by ELISA. (C) IgG levels stratified by exposure. Exposure groups were assigned based on NAAT results and serostatus. Each symbol represents an individual. Data were analyzed using a nonparametric Mann-Whitney U test. P < .05 was considered statistically significant. Ab = serostatus; CT = C. trachomatis; MOMP = major outer membrane protein; NAAT = nucleic acid amplification test.
We next estimated the level of C. trachomatis exposure in this cohort, based on NAAT and serology (Supplementary Table 1). NAAT−/Ab− participants were uninfected, with no evidence of prior or current infection. AGYW with the lowest relative exposure were NAAT+/Ab− and likely had an early, primary infection prior to the development of antibodies [35]. Since antichlamydia antibodies can persist for at least 10 years [34, 36], NAAT−/Ab + participants had previously cleared or treated infections. The group with the highest exposure, NAAT+/Ab + participants, was considered to have an untreated infection of unknown duration or less likely, a recurrent (reinfection) or persistent (treatment failure) infection [36, 37]. Antibody levels increased progressively across exposure groups, with median IgG values of 4.61RU, 13.85RU, 32.98RU, and 40.69RU, respectively (Figure 1C). This grouping framework enabled a nuanced assessment of exposure history in a highly affected cohort and provided a unique window into the development of immune responses during early C. trachomatis infection.
C. trachomatis Infection History and Genital Tract T Cell Activation
Previous work within this cohort showed that C. trachomatis infection was associated with elevated concentrations of genital tract inflammatory cytokines, including IL-6, TNF (also known as TNF-α), IL-1β, and IFN-γ [29]. Given that repeated or long-term exposure to C. trachomatis has been implicated in reproductive tract immunopathology [13], we examined relationships between exposure history, as measured by serology, and the immune environment in the genital tract. Antichlamydia IgG positively correlated with the concentration of cervicovaginal IFN-γ (P = .037, r = 0.174; Figure 2A), suggesting a significant but weak link between exposure and the mucosal IFN-γ response. No associations were observed between antibodies and other cytokines (IL-6, TNF, and IL-1β). Notably, antichlamydia IgG also positively correlated with the frequency of activated cervical CD4+ T cells expressing HLA-DR (P = .018, r = 0.211; Figure 2B) and those coexpressing CD38 and HLA-DR (P = .012, r = 0.223; Figure 2C). A similar association was seen with activated CD8 + HLA-DR + T cells (P = .019, r = 0.209; data not shown), though no correlations were found with CD4 + CD38+ T cells or CD8 + CD38+ T cell subsets alone.
Exposure to C. trachomatis is associated with T cell activation in the female genital tract. Associations between IgG levels and (A) cervicovaginal IFN-γ concentrations (n = 144); and (B, C) frequencies of activated cervical T cells (n = 126). (D, E) Associations between relative exposure and the frequencies of T cells in the female genital tract (n = 126). Nonparametric Spearman correlations and Kruskal-Wallis statistical tests were performed. Statistical analyses were performed with false-discovery rate correction using the step-down procedure where appropriate. P < .05 was considered statistically significant. Ab = serostatus; NAAT = nucleic acid amplification test.
We next compared cytokines and T cell activation across the relative exposure groups. While cytokine concentrations did not differ significantly between groups (data not shown), AGYW with evidence of prior, cleared C. trachomatis exposure (NAAT−/Ab+) had significantly lower frequencies of activated cervical CD4 + CD38+ T cells (median: 56.6%) compared to NAAT+/Ab− (P = .018; median: 70%) or NAAT+/Ab + (P = .022; median: 77.2%) participants (Figure 2D). Furthermore, highly exposed women (NAAT+/Ab+) had significantly greater frequencies of activated cervical CD4 + CD38 + HLA-DR + T cells compared to unexposed AGYW (medians: 32.7% and 17.75%, P = .018) or those with past infection (NAAT−/Ab+; medians: 32.7% and 16.1%, P = .022; Figure 2E), reflective of the active infection in the NAAT+/Ab + women. The associations between cumulative C. trachomatis exposure and elevated cervical T cell activation underscore the potential for immune-mediated pathology due to sustained antigenic persistence.
Circulating C. trachomatis-Specific CD4+ T Cells Reflect Level of Exposure
We measured C. trachomatis-specific CD4+ T cell cytokine responses in PBMCs) from AGYW who were either NAAT + or seropositive at baseline (n = 46; Figure 3A). Most women (n = 36/46) had detectable CD4+ T cell responses to the major outer membrane protein (rMOMP), regardless of NAAT status (Figure 3B). We also found no statistical differences in the proportion of NAAT− or NAAT + participants producing any cytokine (75% and 79%, respectively; P > .99), IFN-γ (63% and 58%, respectively; P > .99), IL-2 (63% and 47%, respectively P = .699) or TNF (50% for both; P > .99) in response to rMOMP (Figure 3B). Similarly, there were no statistical differences in the frequencies of C. trachomatis-specific CD4+ T cells producing any cytokine (median responses: 0.13% and 0.06%, respectively; P = .378), IFN-γ (median responses: 0.069% and 0.034%, respectively; P = .383), IL-2 (median responses: 0.082% and 0.06%, respectively; P = .321), or TNF (median of responses: 0.086% and 0.06%, respectively; P = .771; Figure 3B) between AAT + and NAAT− women. However, when participants were ranked by exposure, clear differences emerged. All the highly exposed AGYW (NAAT+/Ab+) had detectable CD4+ T cell responses (100%), compared to 67% of those with the lowest exposure (NAAT+/Ab-; P = .017). Furthermore, the magnitude of the T cell response was significantly lower in NAAT + Ab + women (median response: 0.05%) compared to NAAT+/Ab− (median response: 0.12%; P = .01) or NAAT−/Ab + women (median response: 0.13%; P = .01; Figure 3C), although this last group was very small (n = 8). There were no significant differences in cytokine-specific responses (P > .05; IFN-γ, IL-2, or TNF) across exposure groups (Figure 3D).
Peripheral CD4+ T cell responses to C. trachomatis are detectable in highly exposed AGYW. (A) Representative flow cytometry plots showing the IFN-γ, IL-2, and TNF response to rMOMP in one individual. (B) Frequency of CD4+ T cells responding to rMOMP in participants with a positive or negative NAAT result at baseline (n = 46). The number of participants with a positive cytokine response is represented by the pie charts. (C) Frequency of CD4+ T cells producing any cytokine in response to rMOMP across the 3 exposure groups (n = 46). Pie charts indicate the number of participants with a positive cytokine response, defined as twice the background value and a net response >0.005%. (D) C. trachomatis-specific CD4+ T cells producing IFN-γ, IL-2, or TNF. The box and whisker plots represent positive cytokine responses only. Each symbol represents an individual. Data were analyzed using a Kruskal-Wallis test. Fisher's exact test was used to determine differences between pie graphs. P < .05 was considered statistically significant. Ab = serostatus; NAAT = nucleic acid amplification test.
We next investigated the quality of the CD4+ T cell response to C. trachomatis by examining cytokine coexpression. Cytokine profiles were significantly different based on relative exposure (P < .0001; Figure 4). NAAT+/Ab− participants had predominantly monofunctional C. trachomatis-specific CD4+ T cells (median: 67.5%, IQR: 51–98.8%), and a low frequency of C. trachomatis-specific CD4+ T cells producing 3 cytokines (median: 6.62%, IQR: 0–19.9%). In contrast, NAAT−/Ab + AGYW had a more diverse response, with roughly equal proportions of cells producing 1 (medians and IQR: 40.5%; 15.9–86.3%) or 2 cytokines (40.2%; 9.1–45.4%), and a median of 18.2% (IQR: 4.6–35.4%) of cells producing all 3 measured cytokines. Interestingly, participants with the greatest exposure, NAAT+/Ab + lacked these highly functional T cells producing 3 cytokines (median: 0%, IQR: 0–5.9%) and showed a predominance of dual cytokine-producing cells (median: 43.8%, IQR: 21–100%). These findings suggest that while persistent or recurrent exposure to C. trachomatis increases the likelihood of detecting systemic T cell responses, it may also be associated with diminished functional quality, particularly a lack of cells coexpressing IFN-γ, IL-2, and TNF.
Exposure to C. trachomatis is reflected in CD4+ T cell function. The graph shows the proportion of C. trachomatis-specific CD4+ T cells producing different combinations of cytokines (n = 36). The pies show the polyfunctionality profiles of C. trachomatis-specific CD4+ T cells stratified by relative exposure (n = 36). Data shown are from participants with a positive total cytokine response. Each bar represents the median. Data were analyzed using a Kruskal-Wallis test. Fisher's exact test was used to determine differences between pie graphs. P < .05 was considered statistically significant. Ab = serostatus; NAAT = nucleic acid amplification test.
C. trachomatis-specific T Cells are Associated With Reduced Genital Inflammation
To understand how C. trachomatis-specific T cell immunity might influence pathogen-associated immunopathology in the genital tract, we examined relationships between peripheral CD4+ T cell responses and cervicovaginal cytokine concentrations in AGYW with matched data. Notably, the frequency of CD4+ T cells producing IFN-γ in response to C. trachomatis stimulation was inversely correlated with levels of several proinflammatory mediators in the genital tract, including IL-1β (P = .007, r = −0.397), TNF (P = .028, r = −0.328), MIP-1β (P = .045, r=−0.301), and IL-17A (P = .024, r = −0.337; Figure 5). Similarly, the frequency of C. trachomatis-specific CD4+ T cells producing TNF was significantly negatively associated with cervicovaginal IL-12p70 (P = .039, r = −0.308). Importantly, none of these mucosal cytokines were associated with exposure, suggesting that these correlations reflect antigen-specific immune regulation rather than cumulative exposure alone. There were no associations found with growth factors (data not shown).
*Associations between peripheral C. trachomatis-specific CD4+ T cell responses and the female genital tract immune milieu. Spearman correlation coefficient (r) and the 95% confidence interval are shown for associations between C. trachomatis-specific CD4+ T cells producing IFN-γ, IL-2, or TNF, and cervicovaginal cytokine concentrations (n = 22). Filled circles indicate statistically significant correlations. P < .05 was considered statistically significant. *P < .05; **P < .01; **P < .001.
DISCUSSION
We examined circulating C. trachomatis-specific CD4+ T cells and their relationship with the genital tract immune milieu in women stratified into 3 groups based on increasing exposure: NAAT+/Ab−, NAAT−/Ab+, and NAAT+/Ab+, which we consider representative of primary infections, cleared/treated prior infections, and untreated/recurrent infections, respectively. Several important observations emerged from our study. The higher exposure to C. trachomatis in NAAT+/Ab + AGYW was associated with activated cervical T cells, consistent with previous studies describing an increased risk of inflammatory sequelae after recurrent C. trachomatis infection in response to persistent antigenic challenge [18, 38, 39]. In terms of C. trachomatis-specific T cell immunity, peripheral blood responses were detected in all NAAT+/Ab + women with untreated/recurrent infections, due to high antigen exposure. Notably, we also found evidence of defects in the CD4+ T cell response to C. trachomatis in this group. These highly exposed women had significantly lower frequencies of C. trachomatis-specific Th1 responses compared to women who were NAAT+/Ab− or NAAT−/Ab + . Furthermore, there was a near-total absence of highly functional C. trachomatis-specific CD4+ T cells simultaneously producing 3 cytokines in NAAT+/Ab + women. Meanwhile, NAAT−/Ab + AGYW had the highest proportion of these cells. These important and novel results emphasize that both the magnitude and quality of the CD4+ T cell response are vital for protective immunity to C. trachomatis.
Our findings may also reflect underlying T cell trafficking dynamics. In NAAT+/Ab + women, antigen-specific CD4+ T cells may have migrated from peripheral blood to the site of current infection, contributing to local immunity. Supporting this, murine studies have shown that C. trachomatis-specific T cells producing IFN-γ circulate between the genital tract and peripheral blood [40]. Clinical studies have similarly reported elevated IFN-γ concentrations in the female genital tract during recurrent C. trachomatis infections [29, 41]. Here, cervicovaginal IFN-γ concentrations correlated with antichlamydia IgG titers, supporting its use as a marker of cumulative exposure. Given the well-established role of IFN-γ in pathogen clearance, we hypothesize that cells producing IFN-γ, including C. trachomatis-specific Th1 cells, contribute to protective immunity at mucosal surfaces. Consistently, we found significant negative associations between the frequency of peripheral C. trachomatis-specific CD4+ T cells and concentrations of cervicovaginal inflammatory mediators like IL-1β and TNF, suggesting a possible reduction in potential immunopathology when higher circulating antigen-specific T cells are detected. These findings suggest that systemic C. trachomatis-specific Th1 responses may not only reflect effective immune control but could also contribute to the regulation of immunopathology. Although technically challenging, C. trachomatis-specific T cells should be studied at the site of infection to fully disentangle the development of protective immunity and immunopathology.
Earlier studies sought to identify correlates of protection from subsequent reinfection following treatment. Bakshi et al. [42] demonstrated that C. trachomatis-specific CD4+ IFN-γ T cell responses were largely absent in women who were reinfected, compared to those who were not. Although the differences in our studies preclude direct comparison, we found that women with untreated/recurrent infections (NAAT+/Ab+) had fewer highly functional CD4+ T cells, despite a high prevalence of detectable responses. This lack of functional diversity may reflect T cell exhaustion or impaired differentiation, processes commonly associated with chronic antigenic stimulation in other infectious settings [43].
We classified participants into exposure groups using baseline NAAT and serology to gain a more nuanced understanding of C. trachomatis infection history. There were some caveats to this approach. Since the treatment data was incomplete, we cannot infer the duration of current or past infections. The short time since sexual debut and the documented persistence of antichlamydia antibodies led us to classify NAAT+/Ab− as primary infections prior to the development of antibodies [35]. However, we cannot entirely rule out failure to seroconvert, rather than early infection. Thus, future studies should also measure IgM, since IgM seroconversion remains poorly characterized [44]. Nevertheless, these groupings still reflect the relative level of C. trachomatis exposure and provide valuable insights into chronic antigen exposure and immunity. A limitation of this study was that NAAT was only performed on vulvovaginal swabs, potentially missing anorectal infections [45], which would affect the classification of NAAT− participants.
We assessed T cell responses using a highly immunogenic recombinant protein antigen, the major outer membrane protein [46]. Although recombinant proteins have the potential to activate nonantigen-specific T cells, a short-term assay was used specifically to minimize this effect. While we cannot completely exclude the possibility of nonspecific stimulation, the immune response to rMOMP, including some degree of nonspecific activation, remains biologically relevant, as in vivo immune responses would similarly involve both antigen-specific and nonspecific T cell activation.
C. trachomatis-specific Th1 responses in blood occur at low frequencies, making them difficult to consistently detect in humans, depending greatly on assay conditions [25, 45]. Our findings need to be validated in larger cohort studies, with additional antigens, eg, heat-shock protein 60, or chlamydial protease-like activity factor (CPAF) to capture a greater breadth of the response [47]. Mucosal-acting cytokines like IL-17A or immunoregulatory cytokines such as IL-10 may also be important in the response to C. trachomatis. Indeed, previous work reported that women who remained uninfected had higher frequencies of bulk Th17 cells, suggesting a role in protection [48]. The ability to identify robustly detectable responses, ideally in blood, will be crucial for future evaluation of vaccine-induced immunity and potential correlates of protection.
In conclusion, we showed that women with untreated or recurrent infections had lower frequencies of C. trachomatis-specific CD4+ T cells, and a smaller proportion of these cells were highly functional. Furthermore, peripheral T cell responses negatively correlated with inflammation in the genital tract. Our data clearly indicate that both the magnitude and function of the CD4+ T cell immune response to C. trachomatis are important components of protective immunity. These findings underscore the importance of including mucosal immune markers and functional systemic responses in the evaluation of candidate chlamydia vaccines, especially in high-risk populations such as AGYW.
Supplementary Material
jiaf595_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO . Chlamydia fact sheet [Internet]. 2024. Available at: https://www.who.int/news-room/fact-sheets/detail/chlamydia
- 2Torrone EA, Morrison CS, Chen P-L, et al Correction: prevalence of sexually transmitted infections and bacterial vaginosis among women in sub-Saharan Africa: an individual participant data meta-analysis of 18 HIV prevention studies. P Lo S Med 2018; 15:e 1002608.29944660 10.1371/journal.pmed.1002608 PMC 6019393 · doi ↗ · pubmed ↗
- 3Pinto CN, Niles JK, Kaufman HW, et al Impact of the COVID-19 pandemic on Chlamydia and Gonorrhea screening in the U.S. Am J Prev Med 2021; 61:386–93.34020848 10.1016/j.amepre.2021.03.009PMC 8131393 · doi ↗ · pubmed ↗
- 4Du M, Yan W, Jing W, et al Increasing incidence rates of sexually transmitted infections from 2010 to 2019: an analysis of temporal trends by geographical regions and age groups from the 2019 Global Burden of Disease Study. BMC Infect Dis 2022; 22:574.35754034 10.1186/s 12879-022-07544-7PMC 9233762 · doi ↗ · pubmed ↗
- 5Zheng Y, Yu Q, Lin Y, et al Global burden and trends of sexually transmitted infections from 1990 to 2019: an observational trend study. Lancet Infect Dis 2022; 22:541–51.34942091 10.1016/S 1473-3099(21)00448-5 · doi ↗ · pubmed ↗
- 6Detels R, Green AM, Klausner JD, et al The incidence and correlates of symptomatic and asymptomatic Chlamydia trachomatis and Neisseria gonorrhoeae infections in selected populations in five countries. Sex Transm Dis 2011; 38:503–9.22256336 PMC 3408314 · pubmed ↗
- 7Kularatne RS, Niit R, Rowley J, et al Adult gonorrhea, chlamydia and syphilis prevalence, incidence, treatment and syndromic case reporting in South Africa: estimates using the Spectrum-STI model, 1990–2017. P Lo S One 2018; 13:e 0205863.30321236 10.1371/journal.pone.0205863 PMC 6188893 · doi ↗ · pubmed ↗
- 8Kaida A, Dietrich JJ, Laher F, et al A high burden of asymptomatic genital tract infections undermines the syndromic management approach among adolescents and young adults in South Africa: implications for HIV prevention efforts. BMC Infect Dis. 2018; 18:499.30285705 10.1186/s 12879-018-3380-6PMC 6171143 · doi ↗ · pubmed ↗