Sex chromosome pairing and multivalent associations during meiosis in diploid and polyploid Silene latifolia
Václav Bačovský, Pavla Novotná, Dylan Phillips, Lucie Horáková, Jana Kružlicová, Jana Čížková, Bohuslav Janoušek, Radim Čegan

TL;DR
This study explores how sex chromosomes in Silene latifolia pair and interact during meiosis, revealing differences in synapsis and recombination patterns in diploid and polyploid plants.
Contribution
The study provides the most detailed cytogenetic analysis of sex chromosome pairing in a plant with young sex chromosomes.
Findings
Both Silene species show class I crossover frequencies similar to mammals.
Sex chromosome configurations vary from rod bivalents in diploids to open-ring tetravalents in autopolyploids.
Y chromosome behavior differs across XXY, XXXY, and XXYY karyotypes.
Abstract
Sex chromosomes undergo various modifications that affect their synapsis during meiosis. While most of the genome achieves full synapsis by the end of pachytene, the nonrecombining regions of XY (or ZW) chromosomes often remain asynaptic and fail to form physical associations at metaphase I. Despite significant progress in animal models, the meiotic behavior dynamics of plant sex chromosomes remain largely unexplored. In this study, we employed super-resolution microscopy to analyze 3D chromosome organization and the localization of key meiotic proteins. Namely, we studied the dynamics of Asynaptic1 (ASY1), Zip1 homolog (ZYP1), and HEI10 across the leptotene to pachytene stages, and compared sex chromosome behavior in dioecious white campion (Silene latifolia) with related gynodioecious species bladder campion (Silene vulgaris). Our findings show that both species exhibit a class I…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1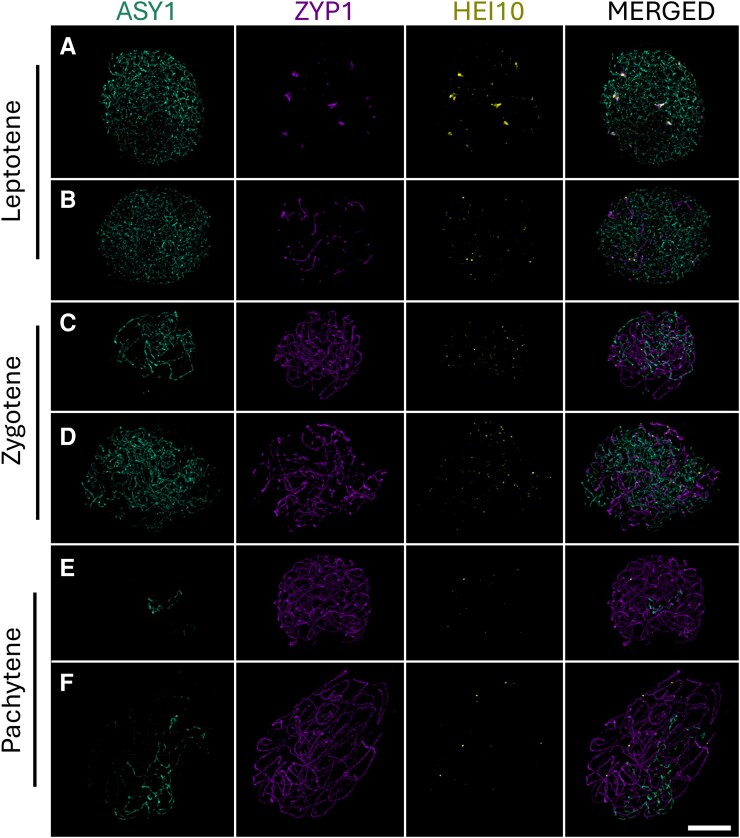 Figure 2
Figure 2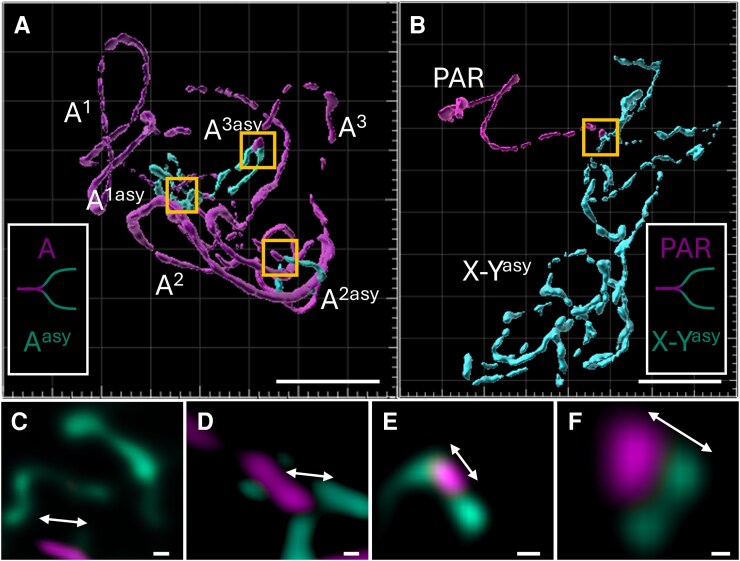 Figure 3
Figure 3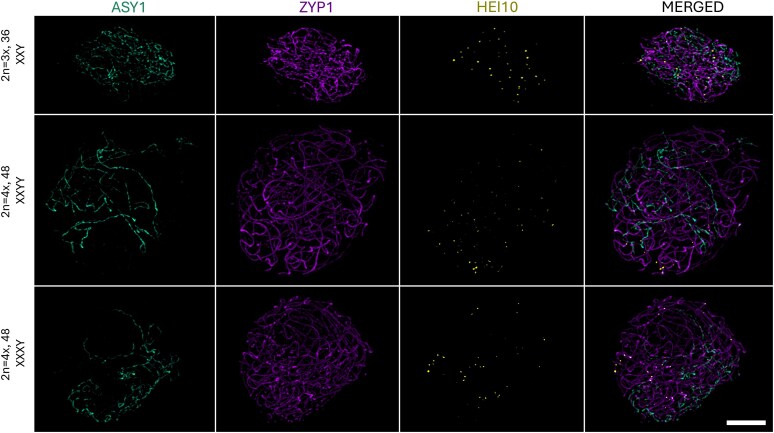 Figure 4
Figure 4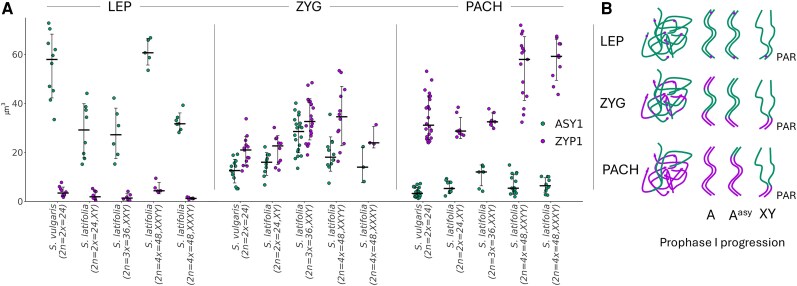 Figure 5
Figure 5- —TowArds Next GENeration Crops
- —ERDF Programme Johannes Amos Comenius
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Chromosomal and Genetic Variations · DNA Repair Mechanisms
Introduction
Sex chromosomes evolved independently across various taxa in animals and plants (Graves 2006; Charlesworth 2016; Vicoso 2019). Though their independent origin, they exhibit a core set of characteristics that include accumulation of deleterious mutations, recombination suppression, Y degeneration, and evolution of dosage compensatory mechanism (Wright et al. 2016; Kratochvíl et al. 2021; Muyle et al. 2021). Following successive steps of recombination suppression, the sequence divergence allows to form “evolutionary strata” (stratum = regions that stopped recombining at different times, and their age is inferred from the level of synonymous substitution between X- and Y-linked alleles; Olito and Abbott 2023). Deleterious mutations disrupt the reading frame of the Y-linked genes (Bergero and Charlesworth 2009) and, together with the accumulation of transposable elements (TEs), cause a decrease in the expression of other mutated sequences, leaving the heterologous chromosome weak (Lenormand et al. 2020; Muyle et al. 2022). In this evolutionary stage, the nonrecombining region often occupies a significantly large part of the Y (W), which in time, becomes heterochromatic and epigenetically inactive (Wright et al. 2016; Sardell and Kirkpatrick 2020).
The genetic and morphological characteristics of sex chromosomes have significant implications for their meiotic behavior (reviewed in Page et al. 2006). Meiosis, a process in which sexually reproducing organisms form gametes, in plants spores (sperm and egg cells), that contain half the chromosome number and display canonical features across different phylogenetic groups (Harrison et al. 2010). The variability among subsequent generations is produced by recombination during prophase I, which involves several key steps that also ensure faithful chromosome segregation. This includes chromosome synapsis via the formation of a protein scaffold along the chromosomes (SC), the formation of double-strand breaks and DNA exchange (crossovers, COs), and the separation of homologous chromosome pairs (bivalents; Mercier et al. 2015). Most notably, the process of synapsis is completed at the end of zygotene of prophase I. The proper meiotic pairing, manifested as full synapsis, is typically associated with DNA exchange. COs are later resolved at the diplotene–diakinesis stage (Qiao et al. 2012; Mercier et al. 2015). To prevent the transmission of damaged or improperly organized genetic material found on sex chromosomes, asynapsed (unsynapsed) chromosomes are subjected to the silencing mechanisms termed as meiotic silencing of unsynapsed chromatin (MSUC) (Turner et al. 2005). This process ensures the chromosomes that fail to synapse are transcriptionally silenced, preventing the expression of potentially harmful genes that might be present on unpaired chromosomes and thereby reducing the risk of gametogenic failure (Cloutier and Turner 2010). In eutherian mammals, asynapsis is a prominent feature of nonhomologous regions of the X and Y chromosomes (Page et al. 2006; Turner 2007). The asynapsed regions are subjected to the process of meiotic sex chromosome inactivation (MSCI), and it may involve changes to the tripartite proteinaceous axis of the synaptonemal complex (SC), particularly the axial and central elements, which may undergo structural and functional modifications (Cahoon and Libuda 2019; Ishiguro 2023). These architectural alterations contribute to sex-specific differences in recombination landscapes and may have profound consequences for meiotic stability, CO formation, or in some cases affect fertility (Sardell and Kirkpatrick 2020; Alavattam et al. 2022). While such phenomena have been extensively studied in animals, the behavior of sex chromosomes in plants remains largely underexplored. In fact, the asynaptic behavior of sex chromosomes, to date, has only been described in the genus Rumex, namely R. suffruticosus, R. acetosella, and R. acetosa, which display distinct XY or XYY meiotic configurations (Cuñado et al. 2007). Interestingly, Vasconcellea parviflora represents an early evolutionary stage of sex chromosomes and appears to exhibit limited asynaptic behavior in the heterochromatic domain on the Y chromosome (Iovene et al. 2015). However, the dynamics during prophase I remain unknown.
Dioecious plants offer a unique opportunity to study meiotic synapsis because the sex chromosomes are still ongoing degenerative processes and are of independent origin (Garcia et al. 2023). The dioecious plant model, Silene latifolia, possesses a large heteromorphic sex chromosome that evolved 11 MYA (Moraga et al. 2025). Both sex chromosomes have accumulated a large proportion of tandem repeats, some of which are X- or Y-chromosome specific, and some that are predominantly enriched in gene regulatory regions (Akagi et al. 2025; Moraga et al. 2025). The sex chromosomes display modest heterochiasmy between linkage groups (LGs) 5, 6, and 11, with higher recombination in males compared to females (Filatov 2023). The recombinationally inactive regions are highly enriched with LTR-Gypsy and LTR-unknown class of TEs, which is similar to the observed phenomenon in related species with no sex chromosomes, S. vulgaris (Akagi et al. 2025). Despite the large developments in the genomic resources available for this model plant, the knowledge of Y chromosome pairing behavior, sex chromosomes association during prophase I, and progression through meiotic division remains limited.
To uncover the sex chromosome dynamics of S. latifolia, we used high-resolution microscopy and a 3D specimen preparation technique to study XY pairing during prophase I. We developed a polyclonal antibody to label the ZYP1 proteins that form the transverse filaments of the SC of Silene, and visualized ongoing synapsis between autosomes and sex chromosomes. Next, we generated artificial triploids and autotetraploids in S. latifolia and analyzed the XY pairing in the various polyploid backgrounds. We specifically aim to answer (i) whether the sex chromosome dose impairs sex chromosome pairing and synapsis, and (ii) what is the behavior of the Y chromosome in the presence of another homologous partner. Using an XY-oligo painting probe, we show preferred associations of various sex chromosome doses during metaphase I and discuss the Y chromosome pairing capability that would have implications for the evolution of nonrecombining region.
Results
Sex chromosome configurations vary with ploidy level and XY chromosome dose
To understand how sex chromosome composition and ploidy level influence meiotic chromosome pairing in Silene, we examined male meiocytes at metaphase I across diploid white campion and bladder campion (S. vulgaris, S. latifolia), and (derived) artificial triploid (XXY), and autotetraploid (XXXY and XXYY) individuals. Plants with a higher ploidy level were derived from flower buds treated with 5 mM colchicine 2 DAP (Supplementary Fig. S1, Supplementary Table S1, Supplementary Note S1). We systematically reconstructed the autosomes configurations and sex chromosome associations during metaphase I by 2D specimen preparation with previously published oligo-painting probe (XY-PAR), and subtelomeric (X43.1), and centromeric (STAR-C) repeats. This combination of probes allows to differentiate 3 orthologous autosomes in related S. vulgaris (2n = 2x = 24), and to position the pseudoautosomal region (PAR) on the q-arm of Y chromosome and on the p-arm of X chromosome in S. latifolia (2n = 2x = 24, XY) (Fig. 1). This XY-PAR positioning was consistent with a previously published model for both species (Bačovský et al. 2020). Cytogenetic observation of the probes in the neotriploid and neotetraploid S. latifolia confirmed their order was preserved (Supplementary Figs. S2 to S5). In Silene vulgaris, the 3 orthologous autosomes identified by XY-oligo painting probe consistently formed 2 ring bivalents and 1 rod bivalent (Fig. 1, Supplementary Fig. S2). In contrast, male plants of S. latifolia formed a rod bivalent between the X and Y chromosomes, with the physical association and CO confined to the PAR (Figs. 1 and 2, Supplementary Fig. S2). Additionally, the subtelomeric repeat (X43.1) was positioned within 3 other autosomal bivalents in S. vulgaris (Fig. 1, Supplementary Fig. S2) and was not colocalized with the XY-PAR oligo painting probe.
Meiotic segregation and sex chromosome pairing during male metaphase I in Silene. In S. vulgaris, 3 autosomes differentiated by XY-PAR positioning, form 2 ring bivalents and rod bivalent configurations at the metaphase I plate. The 3 regions are orthologous to XY chromosomes (dashed line). In contrast, in S. latifolia, the X and Y chromosomes consistently form a rod bivalent at the metaphase I plate. In cases of multiple sex chromosomes (2n = 3x = 36, XXY; 2n = 4x = 48, XXXY/XXYY), the sex chromosomes form from left to right (the upper line): type A—a single rod bivalent and physically separated univalent (X-Y and X), type B—a Y-shaped trivalent (X-X-Y), type C—a chain-like (Y-X-X-X), and type D—an open-ring tetravalent with 3 Xs and a single Y. From left to right (bottom line): type E—an open-ring tetravalent, type F—a single bivalent (X-X) and rod bivalent structure (Y-Y), type G—a chain-like tetravalent (Y-X-X-Y), type H—an open-ring tetravalent in which 2 X chromosomes are connected through the q-arm (Y-Y-X-X), or type I—all 4 sex chromosomes connected via the PAR (YY-X-X). Type J consists of a chain tetravalent structure with a loosely associated Y chromosome (Y-X-X/Y). The formation of all displayed configurations was consistently observed in individual analyzed cells (at least 20 per slide) across 8 replicates. Sex chromosomes in S. latifolia and polyploid individuals were identified using a combination of subtelomeric (X43.1; green) and centromeric tandem repeat (STAR-C; magenta) markers. Arrows indicate PAR. Chromosomes were counterstained with DAPI (gray). Scale bar = 10 µm is applicable to all the images.
Formation of SCs and immunolocalization of meiotic proteins ASY1, ZYP1, and HEI10. A to F) The progression of prophase I in S. vulgaris (A, C, E) and S. latifolia (B, D, F) is visualized through the dynamic localization of ZYP1 along the axial elements. A, B) During leptotene ASY1 (cyan) labels the axial elements, and central elements (ZYP1, magenta) become increasingly distinct. HEI10 forms numerous small foci along the chromosomes. C, D) In zygotene, the ASY1 signal becomes fragmented as synapsis progresses, marked by the extension of ZYP1 along the central element, facilitating SC assembly. HEI10 foci subsequently coalesce into large foci. E, F) By pachytene, ASY1 (axial elements) was restricted to asynaptic regions of the nucleus, while fully assembled SCs were observed between homologous chromosome pairs. These are 3 autosome bivalents in S. vulgaris, and PAR in S. latifolia. HEI10 foci become fewer and larger, indicating a process of coarsening, marking future CO events. The behavior of ZYP1, ASY1, and HEI10 was consistent across all analyzed cells (n = 74; Supplementary Table S3). Scale bar = 4 µm is applicable to all the images.
In triploid S. latifolia males (2n = 3x = 36, XXY), 2 configurations were observed at metaphase I: the predominant type A configuration, where 1 X chromosome remains univalent while the other pairs with the Y, forming a rod bivalent (72.4%). In the second type B configuration, all 3 sex chromosomes link at the PAR, forming a trivalent Y-shaped association (27.6%) (Fig. 1, Supplementary Fig. S3).
Autotetraploid individuals (2n = 4x = 48) with XXXY or XXYY karyotypes showed a wider diversity of metaphase I configurations (Fig. 1, Supplementary Figs. S4 and S5). In XXXY plants, chain-like (type C) and open-ring tetravalents (type D) configurations were frequently observed, indicating at least 2 to 3 CO points among the X chromosomes and their association with the Y (Supplementary Fig. S4). In XXYY individuals, at least 5 distinct configurations were detected (types E to J), ranging from open rings and chains to bivalent combinations, often with asymmetrical Y chromosome involvement (Supplementary Fig. S5).
Among the observed configurations, the open-ring tetravalent was one of the most recurrent and structurally diverse formations. In XXXY individuals, the open-ring tetravalent (type D) was the most frequent configuration, accounting for 63.6% of observed cells (Fig. 1, type D). This configuration typically involved a ring bivalent of 2 Xs connected at both arms and a third X linked to the Y via the PAR, linked together with the Xs bivalents. Thus, this configuration of XXXY resulted in an open-ring tetravalent (Fig. 1, Supplementary Fig. S4).
In XXYY individuals, open-ring tetravalents appeared in multiple structural variants, including types E and F, which together represented over 50% of observed cells (Fig. 1, Supplementary Fig. S5). These included configurations where 2 X chromosomes were connected via the q-arm, and one or both Y chromosomes were associated with Xs via the PAR, located on the X p-arm. Additionally, chain tetravalents, types G and I, formed associations in which Xs pair via q-arm, and both Ys were paired via PAR at X p-arm. Overall, the recurrent presence of open-ring tetravalents across different polyploid karyotypes with 3 Xs or 2 Xs suggested that this structure may represent a preferred mode of synapsis and CO resolution under conditions of increased sex chromosome dose and partial homology. In this regard, type H and type I were the least preferred pairings as they were observed in 3.8% and 1.9% of cells, respectively. All configurations were reproducible across biological replicates (n ≥ 20 cells per slide; 8 replicates), demonstrating stable yet diverse meiotic behavior in response to increasing sex chromosome complexity.
Progression of synapsis from leptotene to pachytene in relation to sex chromosome dose
To further investigate the configurations observed at metaphase I in diploid, triploid, and tetraploid S. latifolia (Fig. 1), we examined the dynamics of meiotic pairing during early prophase I—specifically at the leptotene, zygotene, and pachytene stages (Fig. 2, A to F). To study meiotic synapsis, we produced a polyclonal antibody against ZYP1, isolating and characterizing Silene ZYP1 gene (Supplementary Fig. S6, A to E). In both diploid S. vulgaris and S. latifolia, loading of Asynaptic 1 (ASY1), a marker associated with axial element formation, occurred along all autosomes and sex chromosomes (Fig. 2, A and B), accompanied by the dot-like emergence of central element signals Zip1 homolog (ZYP1). As meiosis progresses in zygotene, ZYP1 extends along axial elements, and numerous HEI10 foci become visible, gradually increasing in size from zygotene into pachytene (Fig. 2, C to F). By pachytene, most chromosomes were fully synapsed, as indicated by continuous ZYP1 signals and gradual disappearance of ASY1 (Fig. 2, E and F).
Three-dimensional (3D) reconstructions of pachytene nuclei revealed persistent asynaptic regions: 6 distinct ASY1-positive axes corresponding to 3 autosomal bivalents in S. vulgaris, and a single unpaired region composed of 2 ASY1-marked axes in S. latifolia corresponding to the X and Y chromosome (Figs. 2 and 3, A and B; Supplementary Video S1 and S2). These unpaired regions are positioned adjacent to a short, fully synapsed domain marked by ZYP1, corresponding to the PAR. Dissection of individual Z-stacks confirmed the presence of discrete ASY1–ZYP1 transition zones in all 3 orthologous chromosome pairs (Fig. 3, C to E), as well as a well-defined PAR boundary (PAB) in S. latifolia (Fig. 3F).
3D analysis of meiotic pairing in pachytene chromosomes of S. vulgaris and S. latifolia. A) In S. vulgaris, 3 orthologous chromosome pairs, depicted by ZYP1 (magenta) axis (A1 to A3), link to 3 asynaptic regions (cyan) designated as A1asy, A2asy, and A3asy. B) In S. latifolia, a single large asynaptic region of 2 axes (X–Yasy) corresponds to the X and Y chromosomes. PAR is visualized by ZYP1 immunolocalization. White boxes in (A, B) represent the boundary of asynaptic region in both species. C, D, E) The insets (yellow boxes) of asynaptic regions in S. vulgaris (A), the boundaries between synapsed and asynaptic regions are sharply defined, as revealed in maximum intensity projections, marked by arrows. F) The inset (yellow box) of asynaptic region in S. latifolia (B), the asynaptic region displays a clear boundary at the PAB, extending into 2 protruding ASY1-marked axes representing the unsynapsed portions of the X and Y chromosomes. The whole 3D reconstruction and optical sectioning of pachytene chromosomes for both species are shown in Supplementary Videos S1 and S2. Scale bars in main panels (A, B) = 3 µm. Scale bar in insets (C to F) = 0.5 µm.
The same synaptic dynamics were observed in polyploid individuals, including XXY, XXXY, and XXYY genotypes, where clearly distinguishable asynaptic regions persisted despite increased sex chromosome dose (Fig. 4, Supplementary Figs. S7 and S8). The use of 3D imaging nucleus segmentation, followed by axis simulations, allowed us to quantify the ASY1/ZYP1 ratio per cell across different ploidy levels (Fig. 5). As expected, the volume of ASY1/ZYP1 signal increased with genome size, accompanied by a proportional increase in HEI10 foci per cell (Supplementary Fig. S9). In line with the immunolocalization results, volumetric quantification of the ASY1/ZYP1 differences across leptotene–zygotene–pachytene stages confirmed progressive SC assembly. However, the relative size of the asynaptic region, measured by the ASY1 signal, remained unchanged regardless of increasing sex chromosome dose, and the presence of another X or Y homologous partner in XXXY/XXYY individuals (Fig. 5, A and B). This observation aligns with the dominant presence of open-ring sex chromosome configurations at metaphase I (Fig. 1), typically involving partially synapsed X chromosomes with Y chromosomes attached solely via the PAR. The same holds true in the case of XXYY plants in which 2 Ys are attached in the PAR to the Xs bivalent, forming open-ring or chain-like configuration (type E, Fig. 1). It should be stressed that the chromosome configuration as shown in Fig. 1 could not be differentiated at pachytene and correlated to the ASY1/ZYP1 measurements taken from the 3D specimens.
Synaptonemal complex formation in male triploid and autotetraploid S. latifolia individuals. During pachytene, in triploid and autotetraploid plants with XXY, XXYY, and XXXY sex chromosome constitutions, ASY1 (cyan) is confined to asynaptic regions of the nucleus, while fully assembled SCs are present between homologous chromosome pairs (ZYP1; magenta). HEI10 (yellow) foci are observed exclusively on fully assembled SCs. Scale bar = 4 µm is applicable to all the images.
Dynamics of ASY1 and ZYP1 loading during meiotic pairing in prophase I. A) Quantification of ASY1 (cyan) and ZYP1 (magenta) signal volumes during leptotene (LEP), zygotene (ZYG), and pachytene (PACH) in S. vulgaris, S. latifolia, and polyploid individuals. A progressive increase in ZYP1 signal corresponds with a reduction in ASY1 as synapsis proceeds. Despite differences in ploidy and sex chromosome constitution, the overall ASY1-to-ZYP1 transition pattern is conserved. Each dot corresponds to an individual cell. Error bars represent standard deviation (SD). B) Schematic representation of ASY1 and ZYP1 loading during leptotene-to-pachytene progression in prophase I. In S. vulgaris, 3 autosomal pairs (Aasy) display asynaptic behavior while the rest of the autosomes are fully synapsed (A). In S. latifolia, as well as in XXY, XXXY, and XXYY individuals, the nonhomologous regions of the X and Y chromosomes (X–Yasy) are asynaptic, marked by strong ASY1 adjacent to short ZYP1-positive PAR segments. The autosomes are synapsed (A).
Quantification of HEI10 foci during meiotic prophase I reveals temporal and karyotype-specific dynamics
The HEI10 foci were classified from leptotene to pachytene into 2 types—type A immature HEI10 foci (<0.07 µm) and type B (>0.08 µm), similarly as described in Randall et al. (2022). At early stages (LEP and ZYG), type A foci predominate and are more abundant, with broad variance observed particularly in zygotene (Supplementary Fig. S9). As meiosis progresses to pachytene, type A foci resolve into fewer, brighter type B foci, marking class I COs designation. Notably, a significant reduction in HEI10 foci is observed at pachytene across all genotypes (P = 0.001, horizontal bars) (Supplementary Fig. S9). The total decrease is expected given to the coarsening hypothesis, which proposes the formation of more stable clusters of HEI10 at the expense of smaller and less stable foci (Morgan et al. 2021; Fozard et al. 2023).
In pachytene, the number of class I COs was significantly increased in XXY individuals compared to both S. vulgaris and XY S. latifolia (P = 0.001; Supplementary Fig. S9). The moderate increase in CO number between XXXY and XXYY individuals was expected, given their frequent type D (open-ring tetravalent) configuration. This likely reflects the doubled number of chromosome pairs, and potentially contribution of additional X chromosomes, paired via PAR regions, allowing to have 2 to 3 times more COs than in XY individuals. In comparison, S. latifolia exhibited on average 1.65 times more HEI10 foci per cell in pachytene than S. vulgaris (P ≤ 0.1). The difference between S. vulgaris (1.09 class I CO per bivalent) or S. latifolia (1.24 class I CO per bivalent) XY and polyploid individuals displays a significant increase in type B foci, reaching from 1.45 to 1.69 average number of class I COs per bivalent (P = 0.001; Supplementary Fig. S9).
Together, these results indicate that the number and distribution of potential COs between sex chromosomes, reflected by HEI10 foci and the structure of physical linkage, vary across pachytene configurations and are closely tied to sex chromosome dose. These patterns range from single COs in simple XY rod bivalents to multiple COs in complex multivalent configurations (Fig. 1), with implications for recombination frequency and meiotic stability.
Discussion
In contrast to animal species, the configuration of sex chromosome during prophase I and in metaphase I has only been investigated in a few plant species. The most notable examples are in Rumex species, namely Rumex acetosa, R. acetosella (Cuñado et al. 2007), R. hastatulus (Kasjaniuk et al. 2019), and minor extent in S. latifolia (Warmke and Blakeslee 1939; Zluvova et al. 2007). To address this gap, we conducted a comprehensive cytogenetic analysis of sex chromosome pairing during meiosis in Silene, using the advantage of recently developed tools and 3D super-resolution microscopy.
Implications of limited sex chromosome association in higher-ploidy level
The observed architecture of meiotic chromosome pairing in Silene was shaped by sex chromosome composition and ploidy level. In diploids S. vulgaris and S. latifolia, a canonical bivalent configuration was observed, while the XXY, XXXY, and XXYY karyotypes displayed increasing variability in trivalent and tetravalent formation (Fig. 1). In S. latifolia, the consistent formation of rod bivalents between X and Y chromosomes supports previous findings of distant pairing in wild-type male S. latifolia (Zluvova et al. 2007). In this study, we clearly identified the distal pairing regions using the XY-PAR oligo-painting probe, corroborating physical association and limited recombination restricted to the PAR (Bergero et al. 2008, 2013; Campos et al. 2017; Akagi et al. 2025).
In XXY individuals, the Y chromosome was typically associated with 1 X via the PAR, while the second X remained unpaired, forming either a univalent or, possibly, an early-separated component of a transient trivalent (Fig. 1, type B). While the latter scenario cannot be entirely excluded, the univalent interpretation is more likely given the physical separation and more frequent occurrence of type A configuration (Fig. 1, Supplementary Fig. S3). This is further supported by the total number of HEI10 foci which is comparable to tetraploid plants (Supplementary Fig. S9). The number of HEI10 foci suggests a large number of bi-trivalent configurations, leading to potential interlocks that need to be resolved prior to completing meiosis. The higher number of interlocks in turn may impair or delay synapsis, SC continuity and chromosome structure, influencing HEI10 diffusion and coarsening (Capilla-Pérez et al. 2021; Zickler and Kleckner 2023; Olaya et al. 2024).
Early cytogenetic studies in S. latifolia reported chain tetravalents as the most prevalent type of metaphase I association (Warmke and Blakeslee 1939), though Westergaard (1938) did not observe any such configurations. In our study, the behavior of the Y chromosome in XXXY and XXYY individuals was in fact much more variable and configuration-dependent. In open-ring tetravalents (e.g. types D and E), the Y was stably integrated into a multivalent structure through the PAR, while in other configurations (e.g. type J), it appeared only loosely associated or partially excluded from the main chromosomal complex. In chain-like tetravalents (Fig. 1, type G), the Y chromosome often occupied a terminal position, suggesting limited CO engagement and weaker integration into the SC. Thus, while our findings are consistent with those of Warmke and Blakeslee (1939), we observed a higher frequency of open-ring tetravalent configurations, possibly due to the greater number of cells scored in our analysis and increased resolution. Similarly to our results, 2 configurations of sex chromosomes in XYY individuals were described in R. acetosa (Cuñado et al. 2007). In the case of the Y-X-Y association, the 2 Ys pair with the X chromosome, leading to 2 asynapsed y axes, while in the Y-Y-X configuration, both Ys undergo at least partial synapsis, leaving a substantial part of the X chromosome asynapsed. The latter configuration in R. acetosa is similar to what we observed in XXYY individuals (Fig. 1, type F), though the ratio of cells with Y-X-Y and Y-Y-X configurations is currently not known. Compared to R. acetosa or the species with multivalent sex chromosome configurations, such as Leptodactylus pentadactylus (Noronha et al. 2020), our observations indicate that the Y chromosome is capable of synapsis through the PAR, but its pairing capacity remains strongly constrained. This became more obvious in the polyploid context with additional and potential homologous partner (Fig. 1). Therefore, we conclude that the pairing capacity, is already repressed, reflecting (i) sequence collinearity, (ii) the evolutionary history, and (iii) epigenetic differences between PAR, the X chromosome and the nonrecombining region of the Y (Bačovský et al. 2019; Akagi et al. 2025; Moraga et al. 2025). This is also supported by the evidence of uniform tetravalent configuration in Humulus lupulus var. cordifolius that exhibits exclusively a chain-type multivalent at metaphase I, indicating a conserved pairing behavior likely driven by fixed structural rearrangements (Ono and Suzuki 1962).
Interestingly, in S. vulgaris, a single rod bivalent alongside 2 ring bivalents may represent autosomes with pre-existing recombination patterns that will predispose it to a sex chromosome-like configuration. Indeed, comparison of S. vulgaris and S. latifolia genomes has identified 4 orthologous chromosomes containing regions with limited recombination (Akagi et al. 2025; Moraga et al. 2025). Therefore, we anticipate that the same chromosomes contribute to the asynaptic behavior observed in our study (Fig. 3, C to F). Phillips et al. (2012) described the termination of partially synapsed bivalents near the nucleolus in Hordeum vulgare, indicating delayed synapsis in regions associated with nucleolar organizing regions. Although a similar mechanism is likely at play in S. vulgaris, this species possesses a substantially higher number of rDNA loci (7 chromosomal pairs for 25S rDNA and 2 pairs for 5S rDNA) (Siroký et al. 2001), which could result in late synapsis or incomplete pairing during pachytene (Phillips et al. 2012). Therefore, the observed asynaptic behavior of specific autosomal chromosome regions in S. vulgaris more closely resembles the pairing dynamics of low-recombination domains and sex chromosomes, suggesting shared structural or epigenetic constraints that impede full synapsis.
In line with the physical positioning and immunolocalization results, volumetric quantification of ASY1- and ZYP1-labeled axes across leptotene, zygotene, and pachytene stages revealed a consistent decrease in ASY1 signal and a corresponding increase in ZYP1 as synapsis progressed (Fig. 5A). This transition was observed in both S. vulgaris and S. latifolia, as well as in polyploid individuals (XXY, XXXY, and XXYY). Despite the differences in sex chromosome number and genome size, the overall trend in ASY1/ZYP1 ratio dynamics was conserved. Consistently, quantification of residual ASY1 signal across ploidy levels confirmed that additional sex chromosomes do not facilitate extended synapsis beyond the PAR, highlighting the persistent structural and epigenetic constraints governing XY interactions (Fig. 5B). The observation of large HEI10 foci during pachytene corresponds to the phenomenon observed in other species, supporting coarsening hypothesis (Morgan et al. 2021; Fozard et al. 2023). In our study, the observed average number of class I COs per bivalent, as measured by HEI10 foci at pachytene, was 1.09 in S. vulgaris and 1.24 in S. latifolia, and 1.45 to 1.69 in higher ploidy individuals. This is comparable to, or even exceeds, CO frequencies reported in reptiles such as Sceloporus lizards, where total COs (including both class I and II) range between 1.14 and 1.45 per bivalent in males and females (Marín-Gual et al. 2022). The authors concluded that the rate of COs in reptiles is low compared to that in mammals. Given that our estimates are based solely on class I COs, the actual average number of COs is likely underestimated, indicating that Silene does not exhibit a particularly low recombination rate per bivalent (Supplementary Fig. S9). In fact, the CO ratio in male Silene meiosis is comparable to that observed in A. thaliana (Li et al. 2021) or maize (Luo et al. 2019), aligning with the typical average of 1 to 3 COs per chromosome pair found across most eukaryotic species (Brazier et al. 2025).
Asynapsis during prophase I is shaped by Y chromosome architecture and epigenetic features
The Y chromosome in S. latifolia has undergone at least 2 inversion events that disrupted collinearity with the X chromosome, accelerating recombination suppression and reducing the extent of homologous pairing (Hobza et al. 2007; Bergero et al. 2008, 2013; Marais et al. 2025). In addition, the large nonrecombining region of the Y chromosome is depleted of active histone marks, notably H3K4me3 and H3K27me3 (Bačovský et al. 2019), and exhibits extensive TE accumulation and distinct patterns of DNA methylation (Akagi et al. 2025; Moraga et al. 2025). In maize, recombination hotspots are positively correlated with histone occupancy and enrichment of H3K4me3 and H3K27me3, particularly in gene-rich euchromatic regions (Kianian et al. 2018; Chowdary et al. 2023). In S. latifolia, both histone modifications are substantially enriched only within the PAR of the Y chromosome and at the distal ends of the X chromosome (Bačovský et al. 2019). Therefore, we anticipate that both sequence homology and epigenetic compatibility are essential prerequisites for successful synapsis of sex chromosomes in S. latifolia, potentially explaining the observed metaphase I configurations and pachytene pairing dynamics (Fig. 5, A and B).
The limited recombination near the X chromosome centromere and across the Y-specific region likely reflects not only structural divergence (Akagi et al. 2025) but also progressive epigenetic erosion, which may further contribute to gene down-regulation and functional decay (Bachtrog et al. 2011; Zhou and Bachtrog 2012; Muyle et al. 2021). The observed patterns of sex chromosome pairing during pachytene and metaphase I may have broader implications for the regulation of sex-linked genes, particularly those located in older strata and nonrecombining regions. Although our model represents a relatively advanced state of sex chromosome differentiation, similarly as R. acetosa and R. acetosella (Cuñado et al. 2007), it is tempting to speculate that similar regulatory mechanisms may be present in species with younger sex chromosomes. This includes species already exhibiting asynaptic sex chromosome behaviors, such as papaya (Zhang et al. 2008; Wai et al. 2012) and related V. parviflora (Iovene et al. 2015), and species yet to be tested for this phenomenon, including among others spinach (Deng et al. 2013) and wild strawberry (Tennessen et al. 2016). We suggest that both meiotic pairing behavior and post-meiotic development may influence X- and Y-gene expression, providing further opportunities to link sex chromosome biology with phenotypic outcomes in dioecious crops. Whether the asynaptic regions identified in this study, similar to those seen in the Y-X-Y configuration of R. acetosa, undergo MSUC, a process analogous to MSCI described in animals (Turner 2007; Alavattam et al. 2022), remains an open question for future investigation.
Materials and methods
Plant material
Seedlings of white campion (S. latifolia) inbred population U16, made by 16 generations of full-sib mating, were used as a parental population, as described previously in Bačovský et al. (2020). The ecotype of bladder campion (S. vulgaris) originating from Brno region was used as an outgroup. The seeds of both species are owned by the Institute of Biophysics of the Czech Academy of Sciences. Plants were kept in a greenhouse under a 16 h light/8 h dark cycle at 24 °C in controlled conditions.
Polyploid generation
Triploid and tetraploid individuals were generated using a previously published protocol (Jones et al. 2008), using 5 µM oryzalin and 5 mm colchicine. Due to the low survival rate of young, treated seedlings (Supplementary Note S1, Supplementary Table S1), we used the following protocol treating female flowers 2 days after pollination (DAP) (Supplementary Table S1). Diploid female flowers of U16 generation were first fertilized with diploid U16 male flowers, and 2 DAP the same flowers were washed for 1 h in deionized water, incubated for 7 h in 5 mm colchicine solution, and washed again in deionized water for 1 h. After 14+ days, mature seeds from the treated flowers were sterilized as described previously (Bačovský et al. 2022a). Seeds were grown in a greenhouse under the same conditions as the parental plants. Mean DNA nuclear content of adult individuals was analyzed with flow cytometry, and fluorescence in situ hybridization (FISH) using a previously described protocol (Bačovský et al. 2020, 2022b). For determination of the number of sex chromosomes within the polyploid individuals, we used 2 satellites—subtelomeric (X43.1) and centromeric (STAR-C) satellites, and X-chromosome PAR oligo painting probe (XY-PAR) as described in Bačovský et al. (2020). Hermaphrodite flowers of tetraploid individuals were self-crossed and were used to generate the F2 population. Subsequently, the same plants were used as pollen donors and crossed with U16 diploid females to produce triploid individuals.
Genome size measurement
The nuclear genome size of tetraploid and triploid individuals was estimated following the protocol described in Doležel et al. (2007). Briefly, fresh leaf tissue of S. latifolia or S. vulgaris and the internal reference standard were chopped together in a 1 mL volume of Galbraith's buffer using a razor blade (Galbraith et al. 1983). The crude suspension was filtered through a 50 µm nylon mesh. The filtrate was made up to 50 µg/mL propidium iodide and 50 µg/mL RNase. Samples were analyzed using a CytoFLEX flow cytometer (Beckman Coulter, United States) equipped with a 488 nm blue laser. Pisum sativum “Ctirad” (2C = 9.09 pg; Doležel et al. 1998, 1992) was used as an internal standard. Each sample was analyzed 3 times, each time on a different day. A total of 5,000 nuclei per sample were analyzed, and 2C DNA contents (in pg) were calculated from the means of the G1 peak positions by applying the formula: 2C nuclear DNA content = (sample G1 peak mean) × (standard 2C DNA content)/(standard G1 peak mean). Mean nuclear DNA content (2C) was then calculated for each accession. DNA contents in pg were converted to genome lengths in bp using the factor suggested by Doležel et al. (2003), i.e. 1 pg DNA = 0.978 Gbp (Supplementary Table S1, Supplementary Fig. S1).
ZYP1 antibody development
H. vulgare and Arabidopsis thaliana sequences of ZYP1 gene (Higgins et al. 2005) were used to search Silene orthologs in the NCBI database (Supplementary Fig. S6A). Based on the quality and sequence length, we designed primers for S. colpophyla ZYP1 sequence, annotated in Geneious Prime (2023.1.1). The whole sequence was amplified from S. latifolia gDNA using SlZYP1 F1/R1 primers (SlZYP1 F1—TGGCTAGGTCTCGAGTCGAA and R1—CTCAGCTTGGCGATTGATGC), synthesized by GeneriBiotech (Hradec Kralove, Czech Republic). The gene was amplified using Q5 High-fidelity DNA polymerase (M0491S; NEB), following the manufacturer’s instructions. Resulting PCR products were purified using QIAquick PCR Purification Kit (28104, QIAGEN), and used for blunt cloning using CloneJET PCR Cloning Kit (K1231; ThermoFisher). Ligation reactions were incubated overnight at 16 °C. After desalting, competent Escherichia coli cells were transformed by electroporation using a MicroPulser Electroporator (Bio-Rad). Transformed cells were allowed to recover for 30 min and then plated on LB agar supplemented with ampicillin (100 mg/L). Plates were incubated overnight at 37 °C. The following day, PCR on resulted colonies was performed using pJET1.2-specific primers to identify positive clones. Individual bacterial colonies were used as templates. Primers and nucleotides were removed using the ExoSAP reaction. Sequencing primers (pJET1.2 F/R) were used in all reactions. Samples were sequenced by Macrogen (Amsterdam, Netherlands) and aligned in Geneious Prime (Supplementary Table S2, Supplementary Fig. S6B). The whole consensus protein sequence was analyzed by the Kyte & Doolittle algorithm with linear weight variation model (Kyte and Doolittle 1982), and compared to barley and Arabidopsis (Supplementary Fig. S6C, Supplementary Table S2). To verify the protein structure and similarity to published ZYP1 sequences, the whole protein sequence was computed by AlphaFold2 (Jumper et al. 2021; Supplementary Fig. S6, D and E). Following the verification steps, the whole protein was used for immunization of 2 Guinea pigs to generate a complex polyclonal antibody. The codon optimization, protein synthesis, immunization, and antibody purification were performed by GenScript (New Jersey, United States).
Chromosome preparation and meiotic analysis
S. vulgaris (2n = 24), S. latifolia U16 (2n = 2x = 24, XY), triploid U17 (2n = 3x = 36, XXY), and autotetraploid U17 (2n = 4x = 48, XXYY; 2n = 4x = 48, XXXY) were used for further meiotic analysis (Supplementary Table S1). Chromosomes in metaphase I were prepared from young flower buds. Anthers were dissected under Olympus Stereomicroscope SZX16 (Evident) supplemented by LED fluorescence and light system, and Olympus DFLPLAPO 0.8PF objective. Anthers were first squashed in 1% acetocarmine, and desired stage was verified under Light Microscope CX43 (Evident), supplemented with PlanC 40× objective (NA 0.65) and phase contrast. The remaining anthers of the same size within the same flower were placed into Clarke's fixative (ethanol:glacial acetic acid, 3:1, v/v) for 72 h at RT, washed with freshly prepared Clarke's fixative again, and stored at −20 °C until use. Chromosomes for FISH experiments were prepared by the squashing technique as described in Bačovský et al. (2022b), with minor modifications. Briefly, the incubation time in the enzymatic mixture was 20 min, and slides with a well-preserved number of chromosomes were stored in 96% EthOH at −20 °C until use. The FISH was performed using the same combination of probes as for karyotype screening: X43.1, STAR-C, and XY-PAR oligo painting probe. Chromosome pictures were captured with Olympus AX70 microscope equipped with CCD1 camera, fluorescence PhotoFluor LM-75 light source, PlantAPO oil objective 100× (NA 1.35), and filter set for 405, 488, 561, and 642 nm fluorescent wavelength. All channels were processed in Adobe Photoshop v. 26.9.0.
Immunostaining and super-resolution microscopy
Anthers in prophase I were screened in the same way as for metaphase I preparation. Specimens for 3D immunolocalization were prepared as previously described in Hurel et al. (2018), with minor modifications. Briefly, anthers from young flower buds were fixed in 2% (w/v) paraformaldehyde, macerated with a brass rod in buffer A, and embedded in acrylamide. Embedded meiocytes were washed in washing buffer (1xPBS supplemented with 1% Triton X-100 and 0.5 mM EDTA) for 2 h, and incubated with following polyclonal antibodies diluted in 1% blocking buffer (1% BSA (w/v) in 1xPBS, 0.1% Tween20): α-ASY1 (rat, 1:200; Hurel et al. 2018), α-HEI10 (1:200; Chelysheva et al. 2012), α-SlZYP1 (guinea pig, 1:300; this study). After incubation with primary antibodies (48 h) at 4 °C, embedded meiocytes were washed 4× 1 h in 1xPBS at RT and incubated with secondary antibodies conjugated with fluorescein (donkey α-rat; 712-097-003), Cy3 (goat α-rabbit; 111-167-003), or Alexa 647 (donkey α-mouse; 715-606-003), diluted 1:400. Immunostained cells were imaged using structured illumination microscopy (3D-SIM^2^) on a Zeiss Elyra 7 microscope equipped with metal halide HXP 120 V fluorescence light source, Plan-apochromat 63× oil objective (1.46 NA), filter set for 405, 488, 561, and 642 nm fluorescent wavelength and 2xPCO edge sCMOS camera with pixel size resolution 6.5 × 6.5 µm^2^ at CELLIM facility (Brno). Images were captured in ZEN Black software with SIM^2^ deconvolution.
Image analysis and HEI10 classification
For image deconvolution in ZEN Black, we applied adjusted parameters with standard-fixed values for ASY1 and ZYP1 across all 3 meiotic stages, using the scale to raw image option in advanced histogram settings. For improved detection of HEI10 signals during leptotene and zygotene, the regularization weight was set to 0.03. In pachytene, HEI10 signal was processed using strong-fixed values and again scaled to raw image histogram settings.
Subsequent image analysis was performed using Imaris (v10.0, Oxford Instruments). To segment the SCs, surface creation was applied using a smooth surface detail value of 0.05 and absolute intensity thresholding. The filter surface tool was manually adjusted (semi-automatic mode) to retain all relevant ASY1 and ZYP1 signal intensities, with no additional subclassification of detected objects. Leptotene, zygotene, and pachytene stages were identified based on ASY1/ZYP1 distribution and the number of HEI10 foci.
HEI10 foci were classified according to their size and signal intensity as described by Randall et al. (2022), with minor modifications and differentiating only 2 types. We used the shortest-distance algorithm for spot detection, with either variable spot sizes or a fixed estimated x, y-diameter of 0.150 µm. Point spread function elongation along the z axis was modeled with a diameter of 0.350 µm. These stricter size estimates, compared to Randall et al. (2022), were calculated by measuring HEI10 foci in x–y–z projections from slice view mode in 10 representative nuclei. Foci were further filtered by mean intensity and then classified into type A or type B based on both maximum intensity and average x/y-diameter: type A foci were <0.07 µm, and type B foci >0.08 µm. Once parameters were optimized for each meiotic stage, all images were processed using the batch analysis tool. Quantitative data were exported to Excel and further analyzed in R Studio. Statistical significance of class I CO differences was assessed using 2-way ANOVA followed by Tukey's multiple comparisons test.
Accession numbers
Sequence data from this article can be found in the GenBank/EMBL data libraries under accession numbers KC747118, AT1G22260, and AT1G22275.
Supplementary Material
kiaf599_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akagi T, Fujita N, Shirasawa K, Tanaka H, Nagaki K, Masuda K, Horiuchi A, Kuwada E, Kawai K, Kunou R, et al Rapid and dynamic evolution of a giant Y chromosome in Silene latifolia. Science. 2025:387(6734):637–643. 10.1126/science.adk 907439913598 · doi ↗ · pubmed ↗
- 2Alavattam KG, Maezawa S, Andreassen PR, Namekawa SH. Meiotic sex chromosome inactivation and the XY body: a phase separation hypothesis. Cell Mol Life Sci. 2022:79(1):18. 10.1007/s 00018-021-04075-3PMC 918843334971404 · doi ↗ · pubmed ↗
- 3Bachtrog D, Kirkpatrick M, Mank JE, Mc Daniel SF, Pires JC, Rice W, Valenzuela N. Are all sex chromosomes created equal? Trends Genet. 2011:27(9):350–357. 10.1016/j.tig.2011.05.00521962970 · doi ↗ · pubmed ↗
- 4Bačovský V, Čegan R, Šimoníková D, Hřibová E, Hobza R. The formation of sex chromosomes in Silene latifolia and S. dioica was accompanied by multiple chromosomal rearrangements. Front Plant Sci. 2020:11:205. 10.3389/fpls.2020.0020532180787 PMC 7059608 · doi ↗ · pubmed ↗
- 5Bačovský V, Čegan R, Tihlaříková E, Neděla V, Hudzieczek V, Smrža L, Janíček T, Beneš V, Hobza R. Chemical genetics in Silene latifolia elucidate regulatory pathways involved in gynoecium development. J Exp Bot. 2022 a:73(8):2354–2368. 10.1093/jxb/erab 53835045170 · doi ↗ · pubmed ↗
- 6Bačovský V, Houben A, Kumke K, Hobza R. The distribution of epigenetic histone marks differs between the X and Y chromosomes in Silene latifolia. Planta. 2019:250(2):487–494. 10.1007/s 00425-019-03182-731069521 · doi ↗ · pubmed ↗
- 7Bačovský V, Janíček T, Hobza R. The sister chromatid division of the heteromorphic sex chromosomes in Silene species and their transmissibility towards the mitosis. Int J Mol Sci. 2022 b:23(5):2422. 10.3390/ijms 2305242235269563 PMC 8910698 · doi ↗ · pubmed ↗
- 8Bergero R, Charlesworth D. The evolution of restricted recombination in sex chromosomes. Trends Ecol Evol. 2009:24(2):94–102. 10.1016/j.tree.2008.09.01019100654 · doi ↗ · pubmed ↗