Heterochromatin-based silencing of a foreign tandem repeat in Drosophila melanogaster shows unusual biochemistry and temperature sensitivity
Tingting Gu, Elena Gracheva, Michael Lee, Wilson Leung, Sophia Bieser, Alixandria Nielsen, Adam T Smiley, Nhi N T Vuong, Matthias Walther, Gunter Reuter, Sarah C R Elgin, Andrew M Arsham

TL;DR
Fruit flies can form heterochromatin around a foreign DNA sequence, but this process behaves differently from natural ones and is affected by temperature.
Contribution
The study reveals a novel mechanism of heterochromatin formation triggered by a foreign DNA sequence in Drosophila, distinct from known systems.
Findings
A foreign lacO tandem repeat can induce heterochromatin formation in Drosophila.
This heterochromatin is HP1a and histone deacetylation-dependent but lacks H3K9 methylation.
Silencing is temperature-sensitive, active at 25°C but suppressed at 18°C.
Abstract
Eukaryotic genomes are packaged into chromatin, a regulatory nucleoprotein assembly. Establishment, maintenance, and interconversion of chromatin states is required for correct patterns of gene expression, genome integrity, and survival. Transcriptionally repressive heterochromatin minimizes mobilization of transposable elements and limits expansion of other repetitive DNA, but mechanisms for recognition of the latter sequences are not well established. We previously demonstrated in Drosophila melanogaster that transcripts derived from 1360 and Invader4 transposon insertions can trigger local conversion of transcriptionally permissive euchromatin to heterochromatin through the piRNA system, but only in a subset of genomic locations near existing blocks of heterochromatin. Here we show that a ∼9 kb tandem array of the 36-nucleotide lac operator (lacO) sequence of Escherichia coli can…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
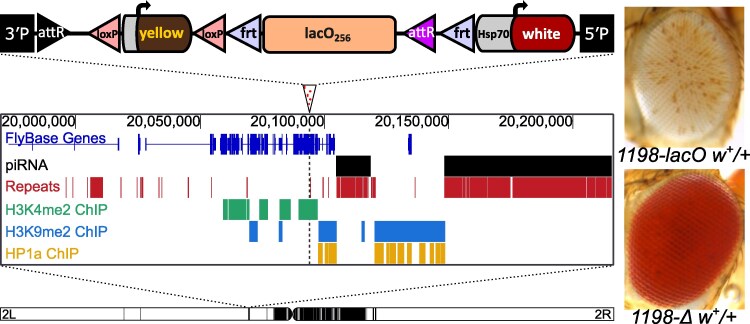 Fig. 1
Fig. 1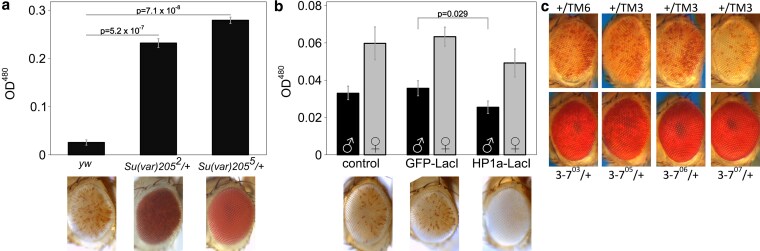 Fig. 2
Fig. 2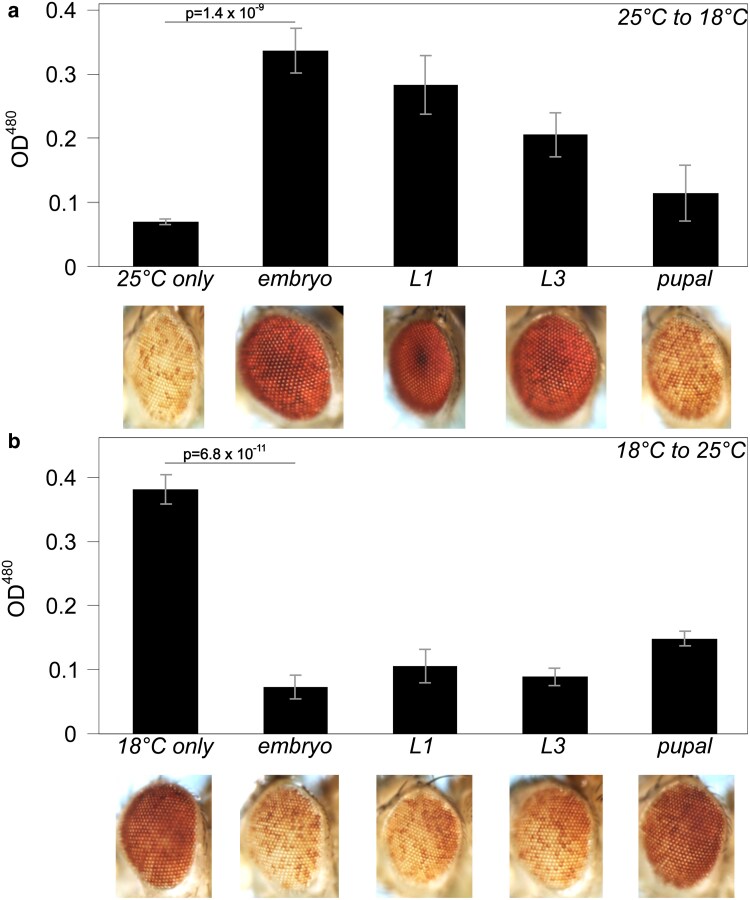 Fig. 3
Fig. 3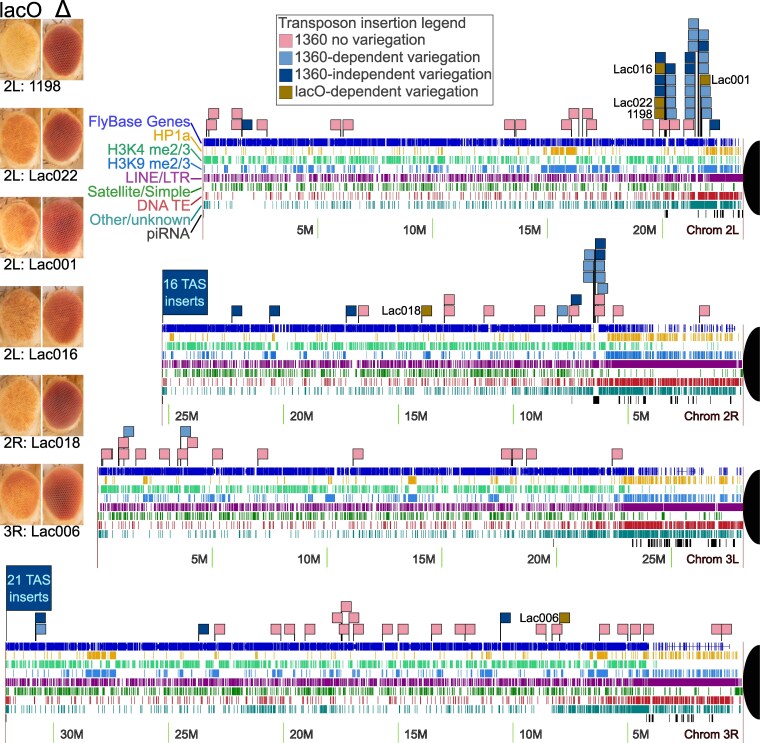 Fig. 4
Fig. 4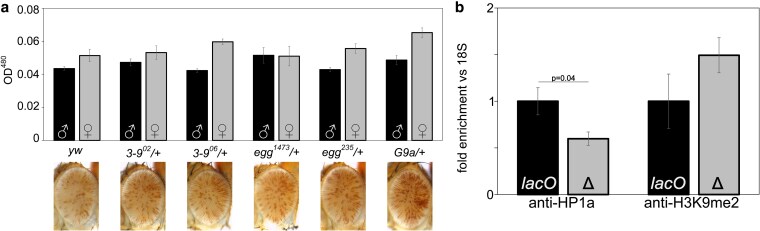 Fig. 5
Fig. 5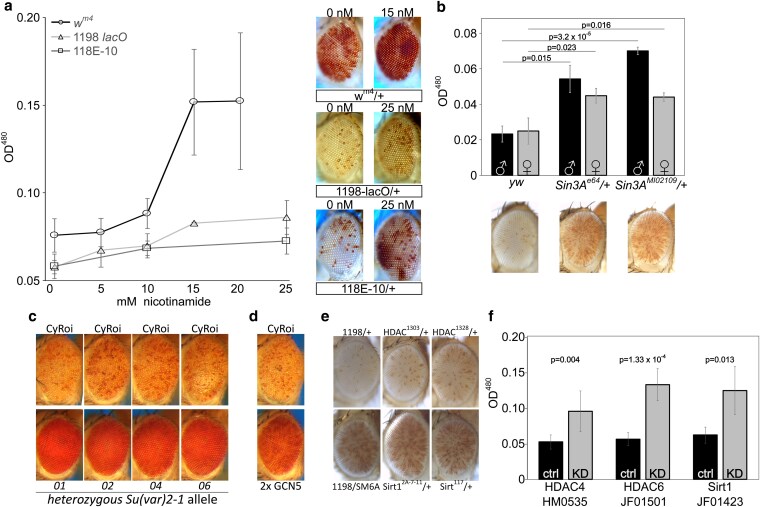 Fig. 6
Fig. 6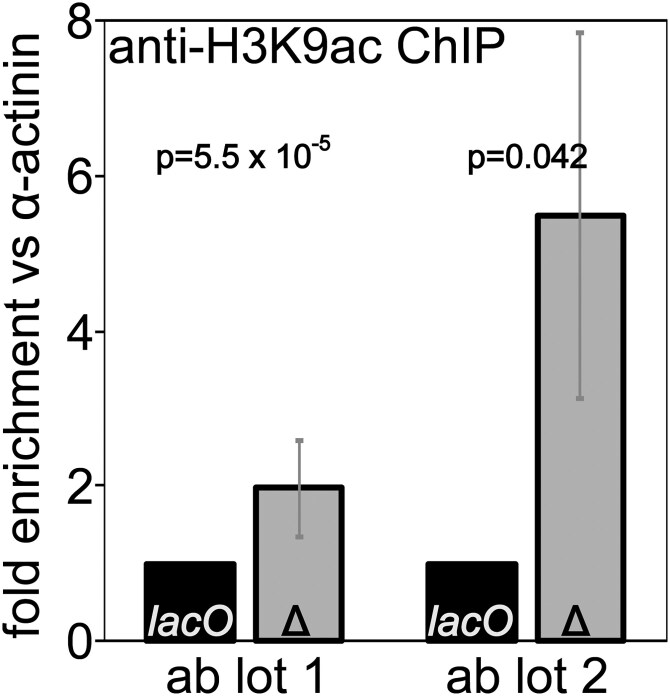 Fig. 7
Fig. 7| Source or Stock # | Mutant genotype | Gene of Interest |
|---|---|---|
| Arsham lab |
| |
| Elgin lab |
| |
| Elgin lab |
| |
| Elgin lab | yw; | |
| Elgin lab |
|
|
| Elgin lab |
|
|
| Elgin lab |
|
|
| Reuter lab |
| |
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| Reuter lab |
|
|
| RRID_BDSC:1033 |
| |
| RRID_BDSC:24470 |
|
|
| RRID_BDSC:24483 |
| |
| RRID_BDSC:24857 |
|
|
| RRID_BDSC:26791 |
|
|
| RRID_BDSC:26792 |
|
|
| RRID_BDSC:28549 |
|
|
| RRID_BDSC:30715 |
|
|
| RRID_BDSC:31053 |
|
|
| RRID_BDSC:31093 |
|
|
| RRID_BDSC:31191 |
|
|
| RRID_BDSC:31399 |
|
|
| RRID_BDSC:31633 |
|
|
| RRID_BDSC:31636 |
|
|
| RRID_BDSC:32480 |
|
|
| RRID_BDSC:33402 |
|
|
| RRID_BDSC:33984 |
|
|
| RRID_BDSC:34279 |
|
|
| RRID_BDSC:34530 |
|
|
| RRID_BDSC:34774 |
|
|
| RRID_BDSC:34778 |
|
|
| RRID_BDSC:3605 |
|
|
| RRID_BDSC:3612 |
|
|
| RRID_BDSC:36512 |
|
|
| RRID_BDSC:36588 |
|
|
| RRID_BDSC:36802 |
|
|
| RRID_BDSC:53697 |
|
|
| RRID_BDSC:5576 |
|
|
| RRID_BDSC:6309 |
| |
| RRID_BDSC:8516 |
|
|
| RRID_BDSC:8838 |
|
|
| RRID_BDSC:8862 |
|
|
| RRID_BDSC:9722 |
|
|
| Wallrath lab |
|
|
| Wallrath lab |
|
|
| Allele | Silencing System | PEV Effect |
|---|---|---|
|
| HP1 | None |
|
| HP1 | None |
|
| Polycomb | None |
|
| siRNA | None |
|
| piRNA | None |
|
| piRNA | None |
|
| piRNA | None |
|
| piRNA | None |
|
| piRNA | None |
|
| piRNA | Su(var) |
| Line | Coordinates | BG3 9-state | S2 9-state | Approximate distance from repeat block(s) |
|---|---|---|---|---|
| 1198- | 2L: 20,094,149 | 1 | 1 | 11 kb from 15 kb block; 52 kb from 77 kb |
| Lac022 | 2L: 20,097,756 | 4 | 5 | 7 kb from 15 kb block; 50 kb from 77 kb |
| Lac016 | 2L: 20,115,973 | 6 | 8 | 0 kb from 15 kb block; 31 kb from 77 kb |
| Lac001 | 2L: 21,630,011 | 1/3 | 1 | 17 kb from 15 kb block; 57 kb from 30 kb |
| Lac018 | 2R: 13,964,566 | 3 | 3/1 | 128 kb from 16 kb block |
| Lac006 | 3R: 7,955,506 | 6 | 9 | 87 kb from 6 kb block; 218 kb from 8 kb; 241 kb from 20 kb |
- —NIH10.13039/501100012264
- —Deutsche Forschungsgemeinschaft10.13039/501100001659
- —Max Planck Society10.13039/501100004189
- —Louis Stokes North Star STEM Alliance10.13039/100017648
- —Bemidji State University’s New Faculty Innovation10.13039/100007260
- —Professional Improvement Grants, and Richard Beitzel Research Fund (AMA)10.13039/501100012596
- —National Institutes of Health10.13039/100000002
- —National Science Foundation10.13039/100019394
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Genomics and Chromatin Dynamics · Bacterial Genetics and Biotechnology
Introduction
Eukaryotic genomic DNA is packaged in chromatin, a dense, dynamic, and heterogeneous nucleoprotein complex. Chromatin packaging and the three-dimensional organization of the nucleus impact gene activity, establishing patterns that enable the selective expression required for cell differentiation and development in multicellular organisms. Chromatin also helps define the boundaries of key genome structures, including telomeres and centromeres, which transmit epigenetic information over cellular, organismal, and evolutionary time scales (reviewed by (Janssen et al. 2018)).
The primary repeating unit of chromatin is the nucleosome, with a core of 147 bp of DNA spooled around a barrel-shaped histone protein octamer. The chromatin state is generally defined by the post-translational modification of the N-terminal tails of histone proteins (Strahl and Allis 2000) and the differentially associated chromosomal proteins (reviewed by (Elgin and Reuter 2013)). Chromatin can broadly be categorized into two types: euchromatin—gene-rich, loosely packed, and transcriptionally accessible—is characterized by high levels of histone acetylation; in contrast, heterochromatin—gene-poor and repeat-rich, more densely packed, and generally transcriptionally inactive—is characterized by low histone acetylation, distinct histone methylation, and the presence of heterochromatin proteins such as Heterochromatin Protein 1a (HP1a). “Heterochromatin” is a useful oversimplification of a large and complex portion of the genome. Heterochromatic domains in fact comprise many subdomains (some with active genes) containing different kinds and lengths of repetitive sequences and transposable elements (TEs) with diverse histone marks. Heterochromatin accumulates these repetitive sequences in part by mitigating their deleterious impact through inhibition of transcription and recombination. In addition to the major blocks of heterochromatin that define centromeres and telomeres, short heterochromatic domains can also be found interspersed throughout otherwise euchromatic regions. Some of these regions may be characterized by TEs or other repeats (reviewed in (Janssen et al. 2018)).
Because heterochromatin silences genes it is imperative that the cell correctly identifies regions of the genome to target for heterochromatin assembly. To explore this issue, we earlier established a transgene system in Drosophila melanogaster (D. melanogaster) whereby the addition of a repetitious sequence from the 1360/Hoppel DNA transposon or from the Invader4 retrotransposon can tip the balance, converting euchromatin to heterochromatin. The presence of the TE sequence stochastically silences an adjacent reporter generating Position Effect Variegation (PEV), a well-known indicator of heterochromatin packaging. Genetic and biochemical analyses have shown that in this case, the change is dependent on the piRNA system (Sentmanat and Elgin 2012).
piRNA-mediated silencing of TEs by heterochromatin has been investigated in detail by numerous labs (reviewed by (Czech et al. 2018)), but much less is known about the silencing of other deleterious repeats, including tandem arrays of short sequences. Could tandem repeats also drive ectopic heterochromatin formation in this reporter system? If so, would small RNA or other sequence-specific signals be involved, or is there something broadly inherent to DNA tandem repeats that can trigger silencing? Cloning and manipulating repetitive DNA fragments is difficult, so to address this question we used an existing 256-copy array of 36 bases of prokaryotic DNA from the lac operon (lacO) of Escherichia coli (E. coli), using a P element vector to insert this fragment into the Drosophila genome. Here we show that this lacO array can induce ectopic heterochromatin formation in a subset of genomic regions. These regions substantially overlap those that we previously found susceptible to TE-induced heterochromatin formation (Haynes et al. 2006; Huisinga et al. 2016). However, unlike common TEs, the lacO repeat sequence used here can be defined as “foreign,” i.e. not previously seen by the organism. It is not present in 78 D. melanogaster genome assemblies searched, including the reference genome (Release 6 plus ISO1 MT assembly), a selection of geographically distant strains (Srivastav et al. 2024), and a recent PacBio HiFi assembly which includes the highly repetitive regions of the D. melanogaster genome (Shukla et al. 2025). Thus the experiments reported here argue against recognition based on pre-existing genomic sequences (such as an RNAi mechanism) and instead point to properties inherent to repetitive DNA.
The heterochromatin formed at lacO repeats has a number of interesting and useful properties, with a distinct histone modification pattern and an unusual temperature sensitivity which we have exploited to probe the silencing process. We anticipate that the system we have developed will be useful in future studies examining how organisms defend themselves against invading non-TE repetitious sequences.
Materials and methods
Fly husbandry
All fly stocks (Table 1) were cultured at 25 °C in 60 to 70% relative humidity unless otherwise indicated. Most experiments used a fly food recipe consisting of 45 g agar, 155 g brewer's yeast, 674 g sucrose, 858 g cornmeal, 8.5 L water, 100 mL propionic acid, and 70 mL of 10% tegosept. Generation and characterization of the lacO transposition mutants was carried out on the standard sucrose cornmeal media recipe used by the Bloomington Drosophila Stock Center with a sprinkle of dry yeast pellets.
Generation of the 1198-lacO-hsp70-white stock
Construction of the original 1360 hsp70 w+ P element reporter P{T1} has been described previously (Haynes et al. 2006); relevant primer sequences are listed in Supplementary Table 1. To swap 1360 for the lacO array the loxP-frt elements from vector pCR2.1-loxP-frt were cloned into the XhoI site of pCR2.1-attB1-loxP-y-attB2 using primers XhoI8-24F/R to make pCR2.1-attB1-loxP-y-loxP-frt-attB2 (Sentmanat and Elgin 2012). The lacO cassette is comprised of 256 copies of a 36 nt E. coli lac operator sequence CCACATGTGGAATTGTGAGCGGATAACAATTTGTGG (Sasmor and Betz 1990) and its cloning has been previously described (Li et al. 2003; Memedula and Belmont 2003). The repeat array was excised from pBS KS(-)-lacO using XhoI and inserted into pCR2.1-attB1-loxP-y-loxP-frt-attB2 to make pCR2.1-attB1-loxP-y-loxP-frt-lacO-attB2. The size of the insertion was confirmed by enzymatic digest and agarose electrophoresis. Male 1198-1360 flies (Sentmanat and Elgin 2012) were crossed to females expressing the phiC31 integrase in germline cells driven by the vasa promoter (BDSC 24483) and embryos were injected with the lacO-containing vector. Putative F1 male recombinants were selected by the presence of the yellow+ phenotype and full cassette exchange was verified by PCR (Supplementary Fig. 1) as previously described (Sentmanat and Elgin 2012).
Homology search for lacO sequences in D. melanogaster genome
NCBI blastn (Altschul et al. 1997) was used to compare the 36 nucleotide lacO sequence (CCACATGTGGAATTGTGAGCGGATAACAATTTGTGG) and a 256-copy array of that sequence to 78 D. melanogaster genome assemblies (see Supplementary Table 3 for full list). This list includes the Release 6 plus ISO1 MT assembly (RefSeq accession GCF_000001215.4) maintained by the FlyBase consortium (Öztürk-Çolak et al. 2024)), a recent PacBio HiFi assembly (GenBank accession GCA_042606445.1) (Shukla et al. 2025), and a collection of geographically distant strains sequenced to study piRNA cluster evolution (Srivastav et al. 2024). Parameters were: Expect threshold = 0.05, Word Size = 11, Match/Mismatch Scores: 2, −3, Gap Costs: Existence: 5 Extension: 2, Low complexity regions filter = off.
In vivo excision of the lacO repeat array
The lacO repeats are flanked by FRT sites (Fig. 1), allowing precise removal of the repeat array from the rest of the reporter construct in live flies expressing the FLP recombinase. Creation of stable repeat-deletion lines has been described previously (Sentmanat and Elgin 2012). Briefly, 1198-lacO males were crossed to females expressing a heat shock-inducible FLP on the X chromosome (BDSC 8862), and progeny were incubated at 37 °C for 1 h on days 3 through 7 of larval development. F1 males with mosaic eye phenotypes suggesting high levels of FLP-mediated recombination were crossed to second chromosome balancer stocks and individual F2 males were selected to create stable balanced stocks—excision of lacO repeats was verified by PCR using primers A412F and 200R (Haynes et al. 2006; Supplementary Table 1).
A tandem array of lacO repeats initiates variegated silencing of a reporter gene when inserted at site 1198. Top: schematic of the lacO-hsp70-white reporter construct; middle: genome browser showing the lacO construct insertion site (marked by an arrowhead with dotted line below); the presence in the surrounding 250 kb of piRNA clusters and repetitive DNA; and H3K4me2, H3K9me2, and HP1a maps derived from modENCODE ChIP data from S2 cell lines (Kharchenko et al. 2011). Bottom: position of the 1198 insertion site on the left arm of D. melanogaster chromosome 2 (Sentmanat and Elgin 2012). Black domains indicate pericentric heterochromatin; white domains indicate euchromatin. Inset: heterozygous reporter eye phenotypes in the presence (1198-lacO w+) and absence (1198-Δ w+) of lacO repeats in the construct.
Testing of dominant modifiers of lacO-directed silencing
Candidate variegation-modifying mutant flies were crossed with 1198-lacO/CyO flies in plastic bottles, approximately 20 to 25 females and 10 to 15 males. The presence of mutants was deduced by the absence of dominant balancer phenotypes. Dominant eye effects were assessed qualitatively by visual comparison of pigment levels between mutants and balancer, and between mutant and a parallel cross to yw^67c23^ which does not carry a known PEV modifier and is treated as wild type for the purposes of these studies. For most experiments we crossed mutant males (for example Spn-E, mael, AGO2, E(z), or HP2) with 1198-lacO/CyO females. To account for the possibility of maternal effects in chromatin mutant lines (Gu and Elgin 2013) we crossed mutant females with 1198-lacO males.
RNAi knockdown cross protocol
RNAi-driven knockdown of a variety of genes of interest was done using stocks from the Transgenic RNAi Project (TRiP; Zirin et al. 2020). 10 female flies with 1198-lacO and a ubiquitously expressed GAL4 driven by the daughterless promoter (yw; 1198*-lacO/CyO; da-GAL4/Tm3 Sb^1^) were crossed to 5 balanced or homozygous males with GAL4-driven RNAi hairpins targeting specific genes of interest. As a control, 2 TRiP lines (HDAC4^HM05035^ and Su(z)12^HMS00280^) were crossed to 1198-lacO* flies without the da-GAL4 driver. To eliminate confounding X chromosome markers common to all TRiP stocks only male progeny with no balancer alleles were selected for phenotypic analysis. In a subsequent version of the experiment, each TRiP line was crossed to lacO flies with and without the da-GAL4 driver to provide more consistent controls.
Fly eye imaging
Two distinct eye imaging protocols were used in this study. For all except Fig. 4, Drosophila adults were randomly collected within 24 h after eclosion and aged for 3 to 4 d at 25 °C for full pigment development. They were anesthetized and mounted on microscope slides. Pictures of eyes were taken with a digital camera installed on a Leica S8APO dissecting microscope and processed using ImageJ software. Initial pictures were taken using a white background. Later pictures, especially for flies utilizing RNAi knockdown, used a blue background for greater contrast with the red eye pigment. Detailed methods for Fig. 4 are described in (Pipkin et al. 2024 ). Briefly, 3 to 4 d old adult progeny were frozen overnight and photographed using a full frame digital camera and macro lens mounted on a horizontal focus stacking rail. Images were acquired in RAW format with exposure of 1/25, f2.8, ISO 500. Rail travel from top to bottom is 2750 uM made up of 56 steps at 50 uM each. The 56-image stack was automatically exported from Helicon Remote to Helicon Focus version 7.7.5 and a composite image combining the most-focused pixels of each individual image was generated using the “C, smoothing 4” setting and saved as TIF files.
Eye pigment assay and statistical analysis
In white mutant flies, eye pigmentation reflects expression of the transgenic white reporter gene. Spectrophotometric measurement of eye pigments (described in (Sun et al. 2004)) were made on pools of 15 to 25 3-day old adult flies from at least 4 crosses, and carried out as described in (Sun et al. 2004) with minor changes. Flies were mechanically homogenized in Pigment Assay Buffer (0.01 N HCl in ethanol), followed by incubation at 50 °C for 10 min, centrifugation, and measurement of 480 nm absorbance of the supernatant homogenate to quantitatively determine the level of red drosopterin pigments. We applied a homoscedastic Student's t-Test in Microsoft Excel to generate 2-tailed P-values and, where appropriate, adjusted for multiple comparisons by multiplying the P-value by the number of tests within a given panel. All bars show the average of at least three biological replicates and error bars show standard deviations.
ChIP protocol
Chromatin immunoprecipitation followed the modENCODE protocols (Kharchenko et al. 2011; Riddle et al. 2011) from 300 to 1000 mg of whole 4-day old adult 1198 lacO/CyO or 1198 Δ/CyO flies. Each ChIP experiment included at least two independent experimental replicates, and at least two technical replicates for each sample. Chromatin was sheared over six rounds of 30 s on/30 s off in Bioruptor (Diagenode) resulting in average fragment sizes of ∼100 to 200 bp and immunoprecipitated with antibodies to HP1a (W191) and H3K9me2 (Abcam 1220). The relative enrichment of each mark at the designated region was determined by quantitative PCR (iQ SYBR Green Supermix, Bio-Rad) for 18S and hsp70-white, using 18S to normalize hsp70-white between the two genotypes (relevant primer sequences are listed in Supplementary Table 1). The Δ strain was generated from the 1198-lacO line and was maintained on a similar lab standard yw^67c23^ genetic background, so the two strains should have similar 18S genomic copy number. Two replicates of each PCR were run and input control values were subtracted before calculation of the cycle threshold from the average of the replicates.
The transient nature of histone acetylation required modification to standard protocols. To preserve chromatin acetylation state, the PBS-EDTA+, nuclear extraction buffer, and wash buffers were supplemented with 0.01 mM TSA and 10 mM nicotinamide. Immunoprecipitations were performed using a mouse monoclonal antibody against H3K9ac (MAB Institute, 309-32379, Lots 13012 and 16009). Mock IPs were performed using mouse IgG (Jackson ImmunoResearch Laboratories, 015-000-003, Lot 150101). DNA extracted from immunoprecipitated chromatin fragments was subjected to three real-time PCR analyses using primers for hsp70-white (above) and for α-actinin. Each PCR experiment included H3K9ac IP, mock IP, and input DNA from 1198 lacO/CyO or 1198 Δ/CyO flies. Acetylation enrichment levels were determined based on input-normalized PCR results obtained for the hsp70-white region, and then normalized again to mock IP. Acetylation values obtained for 18S were used to normalize the 1198-lacO and 1198 Δ values for comparison. Relevant primer sequences are listed in Supplementary Table 1.
The lacO transposition mutagenesis screen
To facilitate a transposition mutagenesis screen for genomic locations on the autosomes that support lacO repeat-mediated silencing we first needed to mobilize the 1198 lacO P element and recover insertions on the X chromosome. Unmated w; sp/CyO; delta2-3 sb/TM6B (BDSC 3612) females which express the P element transposase were crossed with w; 1198 lacO/CyO males. Female F1 lacO/CyO; delta2-3 sb were crossed to w; net; sbd; spa males homozygous for viable recessive markers on all autosomes. Individual male F2 progeny with CyO (indicating the absence of the parental lacO insert) and eye color (indicating the presence of a novel lacO insertion) and without stubble (indicating absence of the transposase) were back-crossed to multiply marked recessive females. Stocks with red eyes and all three recessive phenotypes were presumed to carry a new lacO insertion on the X chromosome. Two X chromosome lacO stocks were recovered, and the presence of the full length lacO insert was confirmed by Southern blot (see Supplementary Data) and mapped by inverse PCR as previously described (Huisinga et al. 2016). Southern blots were performed using 10 µg of total genomic DNA from the stocks of interest, hybridized to probes labeled with DIG-High Prime DNA Labeling and Detection Starter Kit II (Millipore Sigma/Roche 11585614910) according to the manufacturer's instructions. Inverse PCR mapping was performed as described in (Sun et al. 2004) and Southern blots as described herein.
Insertion of the full length lacO construct was confirmed by Southern blot (Supplementary Fig. 2). Total DNA was extracted from about 100 adult flies in 500 µl of lysis buffer (0.1 M EDTA, 0.1 M Tris, 1% SDS, 1% DEPC), incubated 30 min at 75 °C. 70 µl of 8 M potassium acetate was added to homogenate, mixed vigorously, and incubated on ice for 1 h before 10-minute spin at top speed in a desktop centrifuge, phenol/chloroform extraction, isopropanol precipitation, and 70% ethanol wash. Approximately 10 µg of genomic DNA was incubated with HindIII, EcoRI, and XhoI restriction endonucleases and separated by electrophoresis on 0.8% agarose. DNA transfer to PVDF membrane and membrane hybridization was performed as described in manufacturer's instructions for DIG-High Prime DNA Labeling and Detection Starter Kit II (Roche 11585614910). Hybridization probe for the transgenic yellow sequence in our lacO cassette was produced by PCR using primers Pvu yellow F and Bgl yellow R. These newly isolated X chromosome lacO lines were used for subsequent transposition screens to recover lines exhibiting lacO repeat-induced silencing; the insertion sites were mapped by inverse PCR as previously described (Sentmanat and Elgin 2012).
We used a custom mirror of the UCSC genome browser (https://gander.wustl.edu/) to assess the proximity of the insertion sites to transposons in the D. melanogaster “Aug. 2014 (BDGP Release 6 + ISO1 MT/dm6)” genome assembly (NCBI accession GCF_000001215.4 (Hoskins et al. 2015)) and to create map displays of insertion sites relative to other genomic features. Genomic regions surrounding the insertion sites were shown with the “Combined Repeats” evidence track showing on the which shows the regions of the D. melanogaster genome with similarity to sequences in the “centroid” Drosophila repeat library that have been detected by RepeatMasker (Smit et al. 2013) using the following parameters: -nolow -s -e wublast. The “centroid” Drosophila repeat library consists of Drosophila transposons in release 20150807 of the RepBase repeat library (Jurka et al. 2005), Drosophila Helentrons (Thomas et al. 2014), and Drosophila ananassae transposons identified by five de novo repeat finders. The five de novo D. ananassae repeat library include sequences from the ReAS repeat library produced by the Drosophila 12 Genomes Consortium (Clark et al. 2007), LTRHarvest (Ellinghaus et al. 2008), RepeatModeler (Smit and Hubley 2008), Tedna (Zytnicki et al. 2014), and dnaPipeTE (Goubert et al. 2015). The protocol used to construct the “centroid” Drosophila repeat library has previously been described ((Leung et al. 2017) in Supplementary File 7 pages 22–25). To create the maps in Fig. 4 we used the same UCSC browser instance displaying tracks for FlyBase genes, modENCODE ChIP Data for Histone Modifications (H3K4 and H3K9 methylation), modENCODE ChIP Data for Chromosomal Proteins (HP1a/Su(var)205), piRNA clusters (Brennecke et al. 2007), and the repeat detection tracks described above.
As described above, the lacO repeats are flanked by FRT sites to allow FLP recombinase-mediated excision of the repeat cassette. Repeat-dependence of variegation in newly isolated lacO lines was tested by crossing them to flies expressing eye-specific FLP recombinase driven by the eyeless promoter (yw; eyFLP/TM3, yw; eyFLP/Sb, or yw; eyFLP Sb/TM3, all derived from BDSC 5576)—silencing was considered repeat-dependent if eyFLP caused a substantial suppression of variegation relative to siblings without eyFLP. To assess temperature dependence of repeat-induced silencing, newly isolated lacO lines were crossed to yw at 18 °C or 25 °C for 24 h and 2 to 4 d old adults were collected and photographed, and eye pigment levels were measured and compared as described in (Pipkin et al. 2024). Data from these experiments are shown in Supplementary Fig. 4.
Temperature shift experiments
Stocks were propagated at the experimental starting temperature in standard media for several generations prior to starting the temperature shift experiments. Roughly 10 female and 3 adult male yw; 1198-lacO/CyO flies were placed in vials were placed in vials of fresh food for 48 h and transferred for 24 h to an enriched semi-defined media containing 10 g of agar, 80 g of brewer's yeast, 20 g of yeast extract, 20 g of peptone, 30 g of sucrose, 60 g of glucose, 0.5 g of MgSO_4_ × 6H_2_O, 0.5 g of CaCl_2_ × 2H_2_O, 6 mL of propionic acid, 10 mL of 10% p-Hydroxy-benzoic acid methyl ester in 95% ethanol in 1 L distilled water (BDSC Semi-Defined Food). For each timepoint 3 independent vials were moved from the starting temperature to the shifted temperature for the remainder of development. Timepoints were 24 h (embryo group), after hatching of first instar larvae (L1 group), after most larvae had entered the wandering stage (L3 group), and after most larvae had formed pupae (Pupal group). The control group remained at the starting temperature. For experiments starting at 18 °C we increased the number female parents to compensate for decreased rate of egg laying at the lower temperature.
Nicotinamide feeding experiments
Nicotinamide is water soluble and was added as powder to warm (∼50 °C) semi-defined fly food to make a concentrated stock of 100 mM. This 100 mM nicotinamide food was diluted with unmodified food to final concentrations of 10 mM, 15 mM, 25 mM. Five female and 2 to 3 male flies from each PEV stock was transferred to fresh medium for 48 h before transfer to enriched semi-defined medium containing nicotinamide. Parents were discarded after 24 h and F1 adults were collected, aged, sexed, and subjected to pigment assays upon eclosion.
Isolation and testing of novel Su(var)2-1 and Su(var)3-7 alleles
To isolate novel Su(var) mutations we carried out a mutagenesis screen in w^1118^ males (BDSC 3605) on 2.5 mM ethyl methanesulfonate (EMS) crossed to PEV flies with a sensitized genetic background containing a spontaneous enhancer of variegation mutation (E(var)1^01^) on a multiply marked balancer. Mutagenized males were crossed with In(1)w^m4^; T(2;3)ap^Xa^ + In(2L)Cy, ap^Xa^ Cy E(var)3-1^01^. The presence of the enhancer of variegation allele E(var)3-1^01^ silences the w^m4^ allele leading to absence of eye color—even modest suppression of variegation can be detected on this background (Reuter and Wolff 1981; Reuter et al. 1985; Wustmann et al. 1989), and flies with this phenotype were isolated and sequenced as described in (Walther et al. 2020). The impact of these novel alleles on 1198-lacO were tested by crossing 1198-lacO females with w^m4^; Su(var)2-1/CyRoi or w^m4^; Su(var)3-7/TM3 Sb Ser males and comparing eye color between mutant and balancer male siblings.
GCN5 overexpression
To test the impact of overexpression of GCN5 on lacO-mediated silencing we injected the P{FlyFos028109-Gcn5-V5-3xFLAG} vector containing a transgenic copy of Gcn5 under the control of the endogenous Gcn5 promoter into embryos with a 2nd chromosome attP docking site for phiC31 integrase-mediated transformation (BDSC 9722). Transgenic flies were generated according to (Bischof et al. 2007; Ejsmont et al. 2009) and males were crossed to 1198-lacO females.
Results
A tandem array of repetitive foreign DNA directs HP1a-dependent silencing
Euchromatic genes placed in close proximity to heterochromatin by rearrangement or transposition can be stochastically silenced by heterochromatin spreading to produce a variegating phenotype. In particular, a white (w) transgenic reporter inserted into heterochromatin by P element mobilization can assume a PEV eye phenotype; the transgene appears to be packaged in the same fashion as nearby heterochromatic genes (Riddle et al. 2008, 2012; Messina et al. 2023). We have exploited this phenomenon to ask whether an array of foreign tandem repeats can induce ectopic heterochromatin formation, using an adjacent white gene as the reporter.
Prior work from our lab has shown that the DNA transposon remnant 1360 can trigger HP1a-dependent silencing of an adjacent hsp70-white reporter gene, but only when inserted in specific genomic locations (Haynes et al. 2006; Sentmanat and Elgin 2012; Huisinga et al. 2016). In this investigation we used a phiC31 landing-pad system to replace the 1360 sequence at insertion site 1198 (2L:20,094,149) with a ∼9 kb tandem array of 256 copies of a 36 nucleotide E. coli lac operator sequence, lacO (Sasmor and Betz 1990; Li et al. 2003). The 1198 insertion site was identified using 1360 to screen for genomic sites where a variegating phenotype is dependent on the presence of a repetitious element (Sentmanat and Elgin 2012). Site 1198 is located in the 5'UTR of the nesd gene, within a gene-rich euchromatic region on the left arm of chromosome 2. This euchromatic location, while several megabases away from pericentric heterochromatin, is approximately 3 kb from an H3K9me2- and HP1a-enriched heterochromatin domain (Kharchenko et al. 2011) and about 10 kb from a piRNA cluster (Brennecke et al. 2007).
The insertion of 256 copies of the lacO sequence resulted in striking silencing of the white transgenic PEV reporter (Fig. 1, top right), stronger than that observed with 1360 (Sentmanat and Elgin 2012). This silencing does not affect the expression of a nearby yellow transgene, consistent with prior reports showing that the strong yellow promoter is resistant to heterochromatin-induced silencing (Yan et al. 2002). We used flanking FRT (Flippase Recognition Target) sites to excise the lacO array in the germline by in vivo heat-shock-induced FLP recombinase. This eliminated silencing, producing progeny with red eyes (Fig. 1, bottom right), confirming that the silencing is repeat-dependent.
In D. melanogaster, local heterochromatin formation is typically associated with high levels of HP1a (encoded by the gene Su(var)205). Consistent with a role in lacO repeat-induced silencing, variegation is significantly suppressed (Fig. 2a) in flies heterozygous for Su(var)205^5^ (a frameshift truncating the protein after amino acid 10 (Eissenberg et al. 1992)) or Su(var)205^2^ (a point mutation disabling the chromo domain (Platero et al. 1995)).
LacO repeat-induced silencing is HP1a-dependent. a) HP1a mutations suppress variegation. 1198-lacO/CyO flies were crossed with one of two distinct HP1a mutant lines, (Su(var)2052 or Su(var)2055) or a yw control. Variegation was suppressed in flies heterozygous for HP1a compared to controls. All pigment assays were performed on male flies, and all photographs are of male fly eyes except for the yw control. b) Introduction of an HP1a-LacI fusion protein enhances silencing at the lacO array. 1198-lacO/CyO flies were crossed with mutants expressing heat shock-activated HP1a-LacI or GFP-LacI fusion proteins that can bind to a lacO repeat array. Crossing in HP1a-LacI resulted in significant loss of expression of the downstream hsp70-white reporter relative to the yw and GFP-LacI controls. Both males and females were tested in the pigment assay (black bars = males; grey bars = females); males are pictured. Two-tailed P-values comparing at least three replicates of the two groups at either end of the horizontal bar are shown in (a) and (b). P-values were corrected for multiple tests; error bars show standard deviation. c) Four heterozygous mutations in the heterochromatin-associated protein Su(var)3-7 suppress lacO repeat-induced silencing (males are pictured; mutation present, bottom row; controls, top row).
Fusion proteins with the lacO-binding domain of the E. coli LacI protein can be used to tether domains of interest to lacO DNA arrays (Memedula and Belmont 2003). Expression of a GFP-LacI fusion protein had no impact on reporter expression. This suggests that protein binding per se at the 1198 site has no impact on the local heterochromatin state, though it is possible that binding is limited by the ectopic heterochromatin. In contrast, expression of an HP1a-LacI fusion protein, previously shown to bind to lacO repeats and to retain full HP1a silencing function (Danzer and Wallrath 2004), enhanced variegation in the 1198-lacO flies, reducing pigmentation by 29% in males (P = 0.029 after correcting for multiple tests) and 22% in females (P = 0.079 after correction). This provides supporting evidence that HP1a, and hence heterochromatin formation, play a role in lacO repeat-induced silencing (Fig. 2b). Further confirming the involvement of HP1a, four distinct EMS-induced mutations in the gene for the HP1a-interacting partner Su(var)3-7 (Cléard et al. 1997) are dominant suppressors of lacO repeat-induced variegation (Fig. 2c).
Silencing induced by the lacO repeat is insensitive to disruption of HP2 or of the polycomb or RNAi complexes
Using PEV reporters, over 150 suppressors and enhancers of variegation (referred to respectively as Su(var) and E(var)) have been identified in D. melanogaster in genetic screens (reviewed in (Elgin and Reuter 2013)). Most of these encode proteins that are either components or post-translational modifiers of chromatin (Elgin and Reuter 2013). To further investigate the nature of the lacO repeat-induced variegating phenotype, we crossed 1198-lacO males to females carrying mutations in genes coding for heterochromatin-associated proteins (identified as Su(var) genes), visually scoring progeny for dominant suppression or enhancement of the PEV phenotype. Heterozygous mutations in Su(var)2-HP2, the gene encoding HP1a binding partner and suppressor of variegation HP2 (Shaffer et al. 2006), have no impact on lacO repeat-induced silencing (Table 2). The Polycomb system maintains silencing of key developmental regulatory genes by a chromatin-based complex that is distinct from HP1a-associated heterochromatin formation. Not surprisingly, heterozygous mutations in the Polycomb complex gene Enhancer of zeste (E(z)) had no impact on lacO repeat-induced PEV (Table 2).
TEs in the Drosophila genome are targeted by the piRNA system for germ line silencing utilizing transcripts from endogenous piRNA cluster loci to provide target specificity (Czech et al. 2018). To evaluate the possibility that transcripts from the lacO repeats are activating endogenous RNAi-based genomic silencing systems based on this sequence, we used BLAST to compare a single copy of the 36 nucleotide lacO sequence and a 256-copy array of that sequence to 78 D. melanogaster genome assemblies. These included the reference assembly (Release 6 plus ISO1 MT; NCBI RefSeq Assembly GCF_000001215.4); a recent PacBio HiFi assembly that resolved the highly repetitive regions of the D. melanogaster genome (ASM4260644v1; NCBI GenBank Assembly GCA_042606445.1) (Shukla et al. 2025), and a selection of geographically distant strains sequenced to study piRNA cluster evolution (Srivastav et al. 2024). No matches with an E-value < 0.05 were found in any case. While this list does not include the exact strain used here, the broad result argues for the absence of *lacO-*matching sequences from existing piRNA clusters, as well as elsewhere in the genome.
Consistent with this, no loss of silencing was observed in flies with heterozygous mutations in Argonaute 2 (AGO2), spindle E (spn-E), aubergine (aub), and piwi, all known components of the piRNA system (Table 2 and Supplementary Fig. 3). This is in marked contrast to piRNA-dependent silencing of the TEs 1360 and Invader4 inserted at the same genomic location (Sentmanat and Elgin 2012). A mutation in maelstrom (mael), a nucleic acid binding protein involved in piRNA regulation (Onishi et al. 2020), does suppress lacO PEV (Table 2); this effect was highly variable and only statistically significant in males. Given that mutations in other piRNA genes had no consistent impact on PEV, we suggest that mael facilitates lacO repeat-induced silencing via a mechanism unrelated to the piRNA system.
Impact of developmental temperature on silencing by lacO repeats
The mottled pigmentation characteristic of PEV is thought to be established in two stages—a stringent and uniform initial silencing step during early embryogenesis followed by leaky maintenance. Stochastic decay of heterochromatin occurs during differentiation and development, allowing expression in some cells while maintaining silencing in others, giving the characteristic mottled appearance (Lu et al. 1996, 1998; Bughio et al. 2019; Bughio and Maggert 2022). While HP1a is required for both initial silencing and maintenance of heterochromatin, the piRNA machinery, required where the target element is a TE, is needed only for initial silencing (Gu and Elgin 2013). Prior studies of temperature dependence have found that PEV is enhanced by culturing at temperatures lower than that typical for the organism (Gowen and Gay 1933; Hartmann-Goldstein 1967). Thus we were surprised to observe a strong inverse temperature-dependence for lacO repeat-induced silencing, which was enhanced at 25 °C and suppressed at 18 °C.
To determine the developmental stage at which this difference is established, we cultured variegating lacO flies for several generations at the silencing-permissive temperature of 25 °C and then carried out timed matings, shifting vials to 18 °C at different stages in development. Transfers were done 24 h after egg laying (‘embryo’); 48 h after egg laying, when larval movement was visible on the food (“L1”); 5 d after egg laying, when larval wandering was observed (“L3”); and 7 d after egg laying, when most larvae had pupariated (“pupal”). Control vials remained at 25 °C for the duration of the experiment. Silencing was dramatically decreased in a developmental stage-dependent manner: the earlier the transfer, the more complete the suppression, indicating that lacO repeat-induced silencing requires active maintenance throughout embryonic and larval development (Fig. 3a). No significant impact was seen when transfer occurred at the pupal stage, presumably because the pigment pattern had already been established.
Impact of developmental temperature on silencing by lacO repeats. a) Silencing triggered by the lacO repeat is robust when animals are raised at 25 °C but is suppressed when animals are transferred to 18 °C during embryonic or larval development. b) Silencing triggered by the lacO repeat is weak in flies raised at 18 °C but is enhanced when flies are transferred to 25 °C during late embryonic or larval development. P-values compare the averages of at least three vials maintained at the starting temperature to vials transferred during the embryo stage; error bars denote standard deviation. The partial effects seen on transfer at the pupal stage suggest that many ommatidia in these flies had already completed their developmental use of the white product before the transfer.
To test whether lacO repeat-induced heterochromatin could be assembled later in development, we reversed the temperature transition, culturing flies for several generations at 18 °C before transfer of progeny to 25 °C at different developmental stages, observed as described above (Fig. 3b). Transfer to the silencing-permissive temperature any time before pupariation restored variegation. Transfer to the permissive temperature after pupariation resulted in a red eye with intermediate pigmentation as shown by direct pigment measurement. This suggests that the transition from a malleable to a fixed phenotype (complete deposition of the pigment in the eye) happens sometime after the onset of pupariation; our experiments lacked the temporal resolution to precisely identify this developmental timepoint. The results suggest that the lacO repeat site remains capable of completing assembly of heterochromatin throughout development. They further imply the existence of a novel temperature-sensitive component required for maintenance of lacO repeat-induced heterochromatin, but not for initial recognition and licensing.
Additional insertion sites support lacO repeat-dependent silencing
Initial testing of the impact of the lacO repeats was done at a single genomic location, 1198 (2L:20,094,149). To test the capacity of the lacO repeats to trigger silencing elsewhere in the genome we carried out a transposition screen, mobilizing the entire reporter P element with the lacO cassette intact (Fig. 1, top) from the X chromosome and looking for white expression from new sites on the autosomes, as described previously (Sentmanat and Elgin 2012). The cassette includes a second reporter gene, the yellow (y) gene of D. melanogaster (scored by observing body pigmentation). The two reporter genes, yellow and white, provide additional insight into the establishment of ectopic heterochromatin. Several pilot screens were carried out to test feasibility and optimize workflows before scaling up. Out of 14,137 unique F2 progeny in the scaled-up screen, approximately 2% (288) had visible transgene expression, a very low rate of transposition. There is no visible difference between failed transposition and a completely silenced insertion, so the true rate of transposition is unknown. Of the F2 male progeny with visible reporter expression, 82% (236 or 1.7% of all F2) displayed full reporter gene expression in eye and body color, suggesting insertions into euchromatic regions. These results demonstrate that in flies, short tandem repeats do not trigger ectopic heterochromatin formation in most genomic locations. This is consistent with prior findings using other repetitive DNA, specifically TE remnants (Haynes et al. 2006; Sentmanat and Elgin 2012; Huisinga et al. 2016).
About one fifth of the recovered mutants (52 or 0.4% of all F2) show some silencing of at least one of the two reporter genes, with the most common silencing phenotype (31 or 0.2% of all F2) being complete loss of white expression with visible expression of yellow. We also observed the converse: full or partial loss of yellow expression with strong expression of white, indicating that in some cases the expression of the two reporters is decoupled despite their proximity to one another. The reduction in body pigmentation in these y- w+ flies was uniform, not variegated, and followed typical pigmentation patterns (see Supplementary Fig. 5 for sample images).
Approximately 0.06% (8 males out of 14,137) of all F2 in the scaled-up screen had variegated eye color; males from all three screens were used to establish new variegated stocks. Stably variegating lacO lines show stochastic “salt-and-pepper” pigmentation phenotypes. We have not observed patches or sectors of pigmentation suggestive of clonal expression but occasionally observe line-specific spatial gradients as in Lac006 on chromosome 3R (Fig. 4). We used inverse PCR to map the genomic position of 5 of the new variegating insertions. Figure 4 shows their location in the context of our prior studies of repeat-triggered variegation using the 1360 transposon remnant (Sentmanat and Elgin 2012; Huisinga et al. 2016).
Identification of new insertion sites that support lacO repeat-dependent silencing. Scaled representations of chromosomes 2 and 3 (Release 6 plus ISO1 MT, NCBI RefSeq assembly GCF_000001215.4) with tracks illustrating the presence of annotated genes (FlyBase: Öztürk-Çolak et al. 2024); HP1a binding (yellow), H3K4 methylation (green), and H3K9 methylation (blue) (modENCODE: Kharchenko et al. 2011); algorithmically detected repetitive sequences (multiple colors); and piRNA clusters (black, Brennecke et al. 2007). Centromeres are represented by black partial circles on the right side of each chromosome arm diagram. Brown flags mark the insertion sites of new lacO repeat-dependent lines described in this study. Other flags illustrate insertion of 1360-containing reporter elements from prior studies (Sentmanat and Elgin 2012; Huisinga et al. 2016): pink denotes full expression of the eye color reporter; light blue denotes 1360 repeat-dependent variegation; dark blue denotes 1360 repeat-independent variegation. The latter includes clusters of insertions into telomere-associated sequences (TAS) which appear to be unique to 1360 and are not observed among lacO repeat insertions. Pairs of photos for each new lacO repeat location are shown with the left image representative of each line's PEV phenotype and the right image representative of the phenotype after eye-specific expression of FLP recombinase to remove the lacO repeats.
The PEV phenotype in all 5 new lacO lines is dependent on the presence of the lacO repeats, as shown by the impact of excision of the lacO repeats with eye-specific FLP recombinase (Fig. 4). We raised larvae from the 5 mapped lines at 18 °C and at 25 °C; all exhibited the same temperature-dependence as the original 1198 line (Supplementary Fig. 4). The ability of the lacO repeats to establish ectopic heterochromatin in a context-dependent manner is thus not limited to a single location but is a more general (albeit limited) cis-acting feature. Although there is no obvious pattern in the chromatin states of the different insertion sites (Table 3), many variegating insertions are in promoters or transcribed regions of euchromatic genes, presumably the best available target for insertion of such a large P element (Spradling et al. 1995). Four of the mapped lines are within 4 Mb of a pericentric heterochromatin mass, but Lac018 is in the middle of the 2R long arm, generally considered a euchromatic domain. Analysis of flanking sequences (Table 3) reveals that all inserts except Lac018, including those within gene-rich regions, are located near >5 kb long blocks of repeats identified by RepeatMasker and Tandem Repeats Finder. No clear patterns in the distribution or type of such repetitive DNA is apparent, but proximity to these likely heterochromatin blocks may facilitate formation of ectopic heterochromatin. We also observed that the recovered lacO-repeat-dependent silencing locations were often close to locations identified in previous studies looking at insertion sites of P constructs showing a white PEV phenotype dependent on an adjacent 1360 element (Sentmanat and Elgin 2012; Huisinga et al. 2016). Such regions, indicated by large clusters of light blue flags in Fig. 4, might be considered “silencing prone.” In particular, a disproportionate number of variegating insertions map to short segments near the pericentric heterochromatin of chromosome 2.
Mutations in H3K9 histone methyltransferases show little impact on lacO repeat-dependent silencing
The presence of HP1a and H3K9me2/3 and the absence of histone acetylation are prominent features of pericentric heterochromatin. In most animals HP1a and H3K9me2/3 are linked by a positive feedback loop, with methylated H3K9 recruiting HP1a, which acts as a histone reader and in turn recruits the histone writer SU(VAR)3-9, which di- and tri-methylates H3K9 (Bannister et al. 2001; Lachner et al. 2001; Schotta et al. 2002). The positive feedback of the system is believed to be important for heterochromatin spreading and maintenance during mitosis. D. melanogaster has three known H3K9 methyltransferases encoded by the genes Su(var)3-9, eggless (aka egg or dSETDB1) and G9a. These appear to operate in different chromatin domains; SU(VAR)3-9 plays a major role in pericentric heterochromatin, while eggless plays a major role in the heterochromatic fourth chromosome (Muller F element), and G9a plays a role in facultative heterochromatin and in developmental chromatin remodeling (Mis et al. 2006; Stabell et al. 2006; Seum et al. 2007; Tzeng et al. 2007; Yoon et al. 2008; Brower-Toland et al. 2009; Merkling et al. 2015). While heterozygous mutations in the genes for these three proteins act as dominant suppressors of PEV for reporters in their domain, they had little to no impact on lacO repeat-induced silencing (Fig. 5a), suggesting that the ectopic heterochromatin induced by lacO differs from endogenous heterochromatin.
H3K9 di- and tri-methylation does not play a dominant role in lacO repeat-dependent silencing. a) 1198-lacO males were crossed to females carrying mutations in the three known H3K9 HMTs (Su(var)3-9, eggless, and G9a) and progeny heterozygous for both the reporter and the HMT mutation were compared to controls (crossed to yw) with wild type HMTs (male progeny are pictured). Introducing these mutations had no effect on silencing as shown by examining the eyes (pictured) and determining pigmentation levels (black and gray bars for males and females, respectively). b) Antibodies against HP1a (left) and H3K9me2 (right) were used for ChIP, followed by qPCR amplification of the 18S and hsp70 reporter promoters in mixed male and female adult flies with lacO repeats intact (lacO) or deleted (Δ). The HP1a and H3K9me2 levels are normalized to 18S relative to a lacO value of 1. Error bars denote standard deviation. HP1-binding is significantly reduced (P = 0.04) after excision of lacO repeats while H3K9me2 is increased; the latter difference is of similar magnitude but is not statistically significant.
To further explore lacO repeat-induced heterochromatin we used ChIP-PCR to measure the amount of HP1a and H3K9me2 bound to the hsp70 promoter of our reporter in line 1198-lacO (see construct, Fig. 1), comparing flies with the lacO repeats to those with the repeats removed by FLP-FRT recombination (Fig. 5b). Deletion of the repeats reduces HP1a association by about 40% (P = 0.04), consistent with the loss of silencing observed in HP1a mutants. It is also associated with a substantial but not statistically significant (P = 0.07) 50% increase in H3K9me2. Thus while H3K9me2 is present at the lacO repeat silenced reporter, the results (Fig. 5b) suggest a different relationship than the HP1a-H3K9me2/3 positive feedback loop commonly reported. We infer that the silencing driven by the lacO repeats reflects an HP1a-dependent heterochromatin assembly that is distinct from previously described Drosophila heterochromatin, possibly uncoupled from H3K9 methylation.
Histone deacetylation plays a role in lacO repeat-induced silencing
To investigate the role of histone acetylation on lacO repeat-induced silencing, we tested the impact of nicotinamide, a small molecule histone deacetylase (HDAC)/sirtuin inhibitor with broad efficacy, feeding it to flies with one of three distinct PEV genotypes: our 1198-lacO repeat line, the classic white mottled 4 line (in which an inversion on the X chromosome moves the white gene adjacent to pericentric heterochromatin (Muller 1930; Solodovnikov and Lavrov 2022)), and a line in which an hsp70-white reporter is inserted into the pericentric heterochromatin of the fourth chromosome (Wallrath and Elgin 1995). Dietary nicotinamide suppressed variegation of all three PEV reporters (Fig. 6a), indicating that, unlike the case of H3K9 methylation, lacO repeat-directed silencing shares a requirement for HDAC activity with other PEV reporters.
Histone deacetylation plays a major role in lacO repeat-induced silencing. a) Mutant embryos from three different PEV lines were reared to adulthood on food containing the HDAC/Sir2 inhibitor nicotinamide, revealing a dose responsive suppression of variegation (males pictured; left 0 drug, right maximum drug tested). X-axis: millimolar concentration of nicotinamide; Y-axis: eye pigment, OD480. Line 118E-10 harbors a variegating reporter insert in Chromosome 4 heterochromatin, serving as a positive control for nicotinamide function. b) Heterozygous Sin3A mutations suppressed variegation relative to wild type controls. 1198-lacO/CyO flies were crossed with yw (control) or with mutants for Sin3A and pigment analysis was carried out on adult progeny heterozygous for the reporter and the Sin3A mutation (females pictured; black bars = males, grey bars = females). c, d) Heterozygous mutations in HDAC-interacting gene Su(var)2-1 (c) and overexpression of histone acetyltransferase GCN5 (d) both suppress variegation. e) Heterozygous HDAC1 and Sirt1 mutants have no significant effect on lacO repeat PEV. f) RNAi-mediated knockdown of HDAC4, HDAC6, and Sirt1 suppress variegation, further supporting a role for histone deacetylation in lacO repeat-directed silencing. Gray bars labeled “KD” express the driver and the indicated RNAi construct; black bars are no-RNAi controls.
We next investigated two proteins known to organize multi-subunit complexes that facilitate HDAC activity. Heterozygous mutations in the HDAC-interacting gene Sin3A had a dominant suppressing effect (Fig. 6b), further implicating histone deacetylation in lacO repeat-induced silencing. Similar results were seen with a series of mutations in the gene Su(var)2-1 (Reuter et al. 1982) which facilitates histone H3K4 demethylation (Yang et al. 2019) and general histone deacetylation (Walther et al. 2020). Multiple EMS-induced Su(var)2-1 heterozygous mutants markedly suppressed 1198-lacO variegation (Fig. 6c). Further supporting the role of deacetylation, we observed suppression of silencing in flies with a transgenic extra copy of the GCN5 lysine acetyltransferase gene (Fig. 6d).
We then sought to identify specific histone deacetylases that impact lacO silencing. Mutations in two well-studied histone deacetylases (Sirt1 and HDAC1) had no significant impact on variegation using the 1198-lacO repeat line as reporter (Fig. 6e). This result could be due to functional redundancy; there are five known histone deacetylases and five SIRT gene products with NAD-dependent HDAC activity in D. melanogaster (Feller et al. 2015). Finally, we tested the effects of RNAi-mediated knockdown of histone deacetylases by crossing 1198-lacO flies with UAS-driven shRNAs targeting HDAC and SIRT genes in the presence of a ubiquitous GAL4 driver (Fig. 6f and Supplementary Table 2). After an initial screen for suppression of variegation identified specific genes for further testing, follow up experiments with shRNAs targeting HDAC4, HDAC6, and Sirt1 confirmed that knockdown of these gene products could suppress lacO repeat-induced variegation. It's unclear why the heterozygous Sirt1 mutation had no effect on variegation while Sirt1 knockdown suppressed it; potentially the knockdown achieved lower protein levels. These results suggest that histone deacetylation is involved in lacO repeat-induced silencing.
We used ChIP followed by qPCR across the promoter region to measure H3K9 acetylation levels at our 1198-lacO reporter gene and compared those levels to acetylation levels when the lacO repeats had been excised by FLP recombinase. Deletion of the lacO repeats leads to an increase in H3K9 acetylation (Fig. 7) further confirming the importance of histone deacetylation in establishment of the local 1198-lacO repeat heterochromatin domain.
Deletion of lacO repeats increases H3K9 acetylation at the reporter gene promoter. Chromatin immunoprecipitation (ChIP) for acetylated H3K9 was performed on flies with (black bars) and without (gray bars) lacO repeats adjacent to the reporter. qPCR results for reporter promoter sequences were normalized to results for alpha-actinin promoter sequences; the resulting values are expressed as fold difference between flies with the lacO repeats (lacO) and without the lacO repeats (Δ). Two lots of antibody gave qualitatively similar but quantitatively distinct results, in both cases supporting increased histone acetylation in the absence of the lacO repeats.
Discussion
A tandem array of 36 bp of E. coli DNA can drive HP1a-dependent silencing
Here we show that a 256-copy array of a 36 bp E. coli lacO DNA sequence inserted into the D. melanogaster genome can silence an adjacent reporter, apparently due to ectopic heterochromatin formation. Dependence on the tandem repeats is demonstrated by the complete loss of silencing upon their excision (Fig. 1 and Supplementary Fig. 4). In earlier screens using either white or yellow as a reporter within a P element transposon devoid of repeats, variegation occurs only when the reporter is within or adjacent to a known heterochromatic domain such as the pericentric heterochromatin, the telomere-associated sequences (TAS), the F element (fourth chromosome), or the Y chromosome (Wallrath and Elgin 1995; Yan et al. 2002). In contrast, the variegating reporters we describe here are inserted into euchromatin. Thus we attribute their variegating phenotype to formation of an ectopic heterochromatin domain, driven by recognition of the tandem repeat of foreign DNA and possibly facilitated by proximity to blocks of repetitive DNA or heterochromatin. This reveals a defense mechanism against intrusion by foreign DNA, as the D. melanogaster genome appears to have no prior exposure to this sequence, suggesting that the organism can defend itself against any short tandem repetitious sequence (see Supplementary Discussion for additional references that informed our thinking).
The lacO repeat-induced variegation correlates with HP1a concentration, losing silencing in response to loss of HP1a function and gaining silencing when HP1a is overexpressed (Fig. 2a and 2b). HP1a is associated with the major constitutive heterochromatic domains identified in Drosophila, and is strongly correlated with reporter silencing (Eissenberg et al. 1992; Eissenberg and Reuter 2009; Phalke et al. 2009). Earlier studies from the Wallrath lab showed that tethering of HP1a to this lacO repeat array using an HP1a-lacI fusion protein is sufficient to induce silencing at a wide range of euchromatic insertion sites; parallel to our observations, this silencing does not show dosage dependence on the Su(var)3-9 H3K9 HMT (Li et al. 2003; Danzer and Wallrath 2004). In the cases reported here, prior tethering of HP1a to the lacO repeat array is not required to induce ectopic heterochromatin formation (Fig. 1); however, silencing of the adjacent reporter is dependent on the dosage of HP1a (Fig. 2a) and binding of endogenous HP1a is enriched at the lacO repeats (Fig. 5b), indicating that HP1a is a key component of the novel heterochromatin complex observed. Whether the enhancement of PEV seen on introduction of the lacI-HP1a fusion protein (Fig. 2b) is due to direct binding of the lacI fragment to the lacO elements or to interaction of the HP1a fragment with the local heterochromatin complex has not been established.
Association with HP1a may be sufficient to drive heterochromatin formation, either through interactions with other HP1-binding proteins such as SU(VAR)3-7 (Fig. 2c) or through HP1a-driven phase separation (see Supplementary Discussion). A multi-copy array of a white transgene has also been reported to induce PEV by local heterochromatin formation (Dorer and Henikoff 1994); this PEV is both HP1a and H3K9 methylation dosage-dependent (Phalke et al. 2009). The differences in the repeating unit (e.g. size and presence of regulatory sequences) compared to the lacO repeat make this example quite different, suggesting that different drivers of heterochromatin formation are likely to be at work. Our results here and several prior studies (e.g. (Honda et al. 2012)) imply a basic structural role for HP1a in heterochromatin assembly that functions independently of H3K9 methylation dosage (Fig. 5).
An unusual temperature sensitivity for maintenance of the lacO repeat-induced silencing has allowed investigation of persistence of the licensing tag and PEV plasticity
In general, PEV is modestly suppressed at higher temperatures and enhanced at lower temperatures ((Gowen and Gay 1933; Hartmann-Goldstein 1967); reviewed in (Spofford 1967; Girton and Johansen 2008)). In contrast, lacO repeat-induced variegation is robust at 25 °C but fails at 18 °C (Fig. 3 and Supplementary Fig. 4). This temperature-dependent plasticity operates in both directions (i.e. both gain and loss of silencing) throughout larval development up to pupariation (when the eye phenotype is fixed), indicating that at least one component of the lacO repeat-induced silencing system required for maintenance of heterochromatin fails at the lower temperature. This unexpected loss of silencing at 18 °C allows an examination of plasticity in heterochromatin formation.
Generation of a stable heterochromatin domain in Drosophila is generally thought to occur in two major steps: initial uniform silencing in the early embryo followed by an ongoing “maintenance” stage. The initial silencing requires first a “recognition” step, identifying domains targeted for heterochromatin formation. The plasticity we observe, in conjunction with earlier observations, argues for a second “licensing” or marking step in late blastoderm/early gastrulation (nuclear cycle 14) that enables local heterochromatin formation, and maintains this memory through somatic cell division. Embryos must establish heterochromatin while activating the zygotic genome, and multiple lines of evidence identify the mid-blastula transition as the point at which pericentric heterochromatin is re-established via recruitment of HP1a (Yuan and O’Farrell 2016; Seller et al. 2019). Lu et al. proposed that the stochastic nature of PEV arises in the maintenance phase, with heterochromatin boundaries receding toward a point of origin prior to being fixed after terminal differentiation (Lu et al. 1996, 1998). In accord with this model, Gu and Elgin were able to show that Piwi, a component of the piRNA system used to recognize TEs for silencing (reviewed in (Czech et al. 2018)), is required in the early embryo but not in the larval stages for silencing of a PEV reporter (Gu and Elgin 2013), indicating that some components of the initial recognition step are not needed after licensing. In contrast, they found that HP1a is required both in the early embryo, potentially as part of the licensing step, and subsequently to maintain silencing. Similar results pointing to a key role for HP1a in the initial establishment of heterochromatin in the early embryo (licensing) have also been reported by Bughio and Maggert (2022) .
Two complementary lines of evidence narrow the temperature-sensitive component of lacO repeat-induced silencing to the maintenance phase. When larvae are moved from 25 °C to 18 °C, loss of silencing is observed implying a failure of heterochromatin maintenance (Fig. 3a). Conversely, when larvae are moved from 18 °C to 25 °C, restoration of silencing is observed (Fig. 3b), implying that the recognition and licensing steps are both functioning properly to mark lacO repeats for silencing even when the adult phenotype is suppressed. In other words, the license persists even when the silencing does not. In contrast, the Gu and Elgin (2013) study showed that the depletion of HP1a in the early embryo resulted in loss of silencing in the adult eye, even when normal levels of HP1a were restored during zygotic gene expression. Combining these observations leads to the conclusion that HP1a plays a key role in licensing, which allows the assembly of heterochromatin at the marked site when the shift to 25 °C occurs during the larval stage, permitting heterochromatin assembly.
A small subset of reporter insertion sites supports lacO repeat-dependent ectopic heterochromatin formation
The studies reported here primarily used a single insertion site previously shown to exhibit TE-dependent silencing (Sentmanat and Elgin 2012). Mobilizing the P element lacO reporter construct revealed additional repeat-dependent silencing sites, 5 of which we mapped; all of these displayed the same temperature- and repeat-sensitivity as the original 1198-lacO line (Fig. 4, Supplementary Fig. 4, Table 2). Several studies using the transposon remnant 1360 (Sentmanat and Elgin 2012; Huisinga et al. 2016) or the lacO array (Li et al. 2003; Danzer and Wallrath 2004) have found that insertion of the repeats themselves is not sufficient to induce ectopic heterochromatin formation at most euchromatic sites; the vast majority of the inserted repeat-containing P elements observed in transposition screens, including this one, are distributed throughout the genome and exhibit wild type red eye color. Here we uncover sites that are normally euchromatic (permissive for reporter expression) but can be converted to heterochromatin by the insertion of the lacO repeat array. The small proportion of insertion sites resulting in variegating eye color are strikingly clustered, indicating that this phenomenon is restricted to a small number of chromosomal subdomains (Fig. 4). Combining multiple studies of lacO repeat- or *1360-*dependent PEV lines from our labs, 21 out of 33 inserts (64%) where variegation is confirmed to be repeat-dependent fall into two clusters within a single 1.6 Mb region of chromosome arm 2L that constitutes only 1.3% of the 120 Mb euchromatic genome. Another six (18%) can be found in a 300 kb segment of arm 2R, 0.25% of the euchromatic genome. Thus >80% of all confirmed cases of repeat-dependent variegation using either TE remnant 1360 or lacO repeat inserts fall within the same 1.5% of the euchromatic genome. The two genomic regions identified are characterized by their proximity to repetitive DNA and heterochromatin blocks, but not uniquely so. The extent and consequences of epigenetic silencing of euchromatic TEs is heterogeneous and varies by species and even strain, and this heterogeneity may be rooted in similar localized differences in chromatin context (Lee and Karpen 2017). Other factors which we have not been able to identify must be contributing to the formation of stable heterochromatin in response to the presence of multiple distinct types of repetitive DNA. This indicates that the feature(s) that define these domains are linked to licensing and maintenance, not recognition.
How might a tandem array of a novel foreign DNA sequence be recognized as a target for silencing? Heterochromatin tends to be AT-rich, and experiments inserting bacterial DNA into yeast chromosomes have suggested that differences in AT content might play a determining role (Meneu et al. 2025). In the case of the endogenous tandem repeats in D. melanogaster, multiple mechanisms appear to be in play. Examples of repeats in D. melanogaster range from minisatellites (tandem arrays with a repeat of a few nucleotides) to the histone gene cluster (∼110 copies of an array of the five genes (Crain et al. 2024; Shukla et al. 2025)) and the rRNA genes (80 to 600 copies (Guetg et al. 2010; Lu et al. 2018)), the latter two both repeating on a kilobase scale (Watase et al. 2022; Ahmad et al. 2025). Several studies have identified specific proteins that bind to these endogenous repetitious sequences and appear to be critical for driving heterochromatin assembly: for example D1 in the case of AT-rich satellite DNAs (Aulner et al. 2002; Blattes et al. 2006), HERS in the case of the histone genes (Ito et al. 2012), and NoRC in the case of rRNA genes (Guetg et al. 2010). However, these strategies do not appear to be well-suited for dealing with novel invading DNA repeats. The lacO fragment has a similar GC% as the rest of the D. melanogaster genome; no significant similarity to any endogenous sequences or piRNAs; and no regions of mono-, di-, or tri-nucleotide repeats that might engage known silencing mechanisms. There is no reason to anticipate the presence of a eukaryotic nuclear protein that can specifically recognize the lacO repeat as a foreign invader to be silenced.
Evidence from multiple systems implicates pausing by RNA polymerase (e.g. Homo sapiens (Colak et al. 2014; Groh et al. 2014; Loomis et al. 2014); Caenorhabditis elegans, (Zeller et al. 2016)) or DNA polymerase (Gaggioli et al. 2023), leading to formation of R-loops and/or D-loops at repetitive sites as a trigger for heterochromatin formation and epigenetic silencing. While some of these mechanisms appear to rely on H3K9 methylation, loop formation can also cause DNA damage and recruitment of the DNA repair machinery, including heterochromatin proteins, notably HP1a and HDAC1/2 (e.g. (Luijsterburg et al. 2009; Miller et al. 2010; Zeller and Gasser 2017), reviewed in (Lemaître and Soutoglou 2014)). Such a mechanism is an attractive candidate to explain lacO repeat-induced silencing because it does not depend on a specific DNA sequence, relying instead on the inherent propensity of repetitive transcripts to form RNA:DNA hybrids (reviewed by (Zeller and Gasser 2017)).
We conclude that the lacO repeat array will drive silencing by heterochromatin formation, resulting in a variegating phenotype in an adjacent reporter, in a small number of chromosomal domains. One possible mechanism for recognition of the repeat array is the formation of R-loops and/or D-loops. If this is the case, the lacO repeat insertion sites that trigger heterochromatin packaging must allow accessibility for transcription but also encompass unidentified features that predispose to repeat-dependent silencing.
Heterochromatin induced by the lacO repeat has a distinct biochemistry
The heterochromatin structure detected here, tracked primarily by its ability to induce silencing of a reporter gene, is dependent on the core heterochromatin proteins HP1a and SU(VAR)3-7 (Fig. 1) and on histone deacetylation (Figs 6 and 7), but is unchanged by reduction of H3K9 methyltransferase gene dosage and is not associated with increased H3K9 di-methylation (Fig. 5). This is surprising given that H3K9 di- and tri-methylation is a prominent feature of pericentric heterochromatin as well as silenced TEs (Penke et al. 2016), at least some trinucleotide repeats (Nageshwaran and Festenstein 2015), and tandem gene arrays (van Steensel et al. 2001). A study that parallels our experiments with the lacO repeats in flies reported here, starting from the 1198 construct but inserting the GAA repeat from a Friedrich's Ataxia patient, also shows a requirement for H3K9 methylation for silencing (E. Gracheva et al., manuscript in preparation).
While a heterochromatic silencing system based on localization of HP1a complexes and histone deacetylation is not the predominant model, it is not without precedent. Similar examples have been reported in other systems. For example, HP1-induced ectopic heterochromatin that is H3K9-independent and HDAC-dependent has been reported in Neurospora crassa (Honda et al. 2012; Gessaman and Selker 2017). A recent study in C. elegans showed that the HP1a homolog HPL-2 functions with LIN-13 (a nuclear regulatory protein) to promote heterochromatic foci and gene repression independently of H3K9 methylation (Delaney et al. 2025). There is also evidence for the parallel presence of an HMT-independent mechanism for HP1a recruitment at the 359 bp satellite sequence in D. melanogaster (Yuan and O’Farrell 2016); this system is consistent with the observations here, suggesting a heterochromatin based on the presence of HP1a and maintenance of de-acetylated histones (Figs. 2a, 2b, and 6).
Many studies have identified histone deacetylation as a key early step in the transition from a euchromatic to a heterochromatic state: in cases where H3K9 methylation is a requirement, H3K9 deacetylation appears to be a prerequisite (reviewed in (Elgin and Reuter 2013)). Indeed, broad deacetylation has been suggested to be a necessary step upstream of heterochromatin formation in the Drosophila embryo (Walther et al. 2020) and in S. pombe (Alper et al. 2013). Although analyzing histone acetylation can be difficult in flies due to redundancy among the five known HDACs (Pallos et al. 2008) and five known Sirts, we were able to generate several lines of evidence indicating that histone deacetylation is a key step in generating the local heterochromatin found at the lacO repeat site (Fig. 6). The lacO repeat-induced silencing was reversed by nicotinamide, a broad suppressor of HDAC/sirtuin activity (Fig. 6a); by mutations in Sin3A or Su(var)2-1, both organizers of HDAC complexes (Fig. 6b and 6c); by overexpression of GCN5, a histone acetyltransferase (Fig. 6d); and by knockdown of specific HDACs (Fig. 6e). Su(var)2-1, initially identified as a suppressor of variegation (Reuter et al. 1982), was recently shown to recruit both HDAC1 (Walther et al. 2020) and the H3K4me1/2 demethylase SU(VAR)3-3 (Yang et al. 2019) to sites of transcriptional repression. Sin3A is also part of a protein complex with HDAC1, and interacts physically and genetically with H3K4me3 demethylase KDM5 (Spain et al. 2010; Gajan et al. 2016) to promote silencing. Together these results argue for a primary role for histone deacetylation in promoting silencing.
In sum, this study of the impact of foreign tandem repeats on local gene expression has identified a form of heterochromatin with distinct features, including distribution in the genome, temperature sensitivity, and biochemistry. These features can now be exploited to further our understanding of the formation, specificity, and stability of heterochromatin and its role in genome integrity.
Supplementary Material
iyaf275_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmad K et al 2025 Sep 29. Cell-cycle-dependent repression of histone gene transcription by histone H 4 [preprint]. bio Rxiv 2024.12.23.630206 [accessed 2025 May 16]. https://www.biorxiv.org/content/10.1101/2024.12.23.630206 v 2. 10.1101/2024.12.23.630206. · doi ↗
- 2Alper BJ et al 2013. Sir 2 is required for Clr 4 to initiate centromeric heterochromatin assembly in fission yeast. EMBO J. 32:2321–2335. 10.1038/emboj.2013.143.23771057 PMC 3770337 · doi ↗ · pubmed ↗
- 3Altschul SF et al 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389–3402. 10.1093/nar/25.17.3389.9254694 PMC 146917 · doi ↗ · pubmed ↗
- 4Aulner N et al 2002. The AT-hook protein D 1 is essential for Drosophila melanogaster development and is implicated in position-effect variegation. Mol Cell Biol. 22:1218–1232. 10.1128/MCB.22.4.1218-1232.2002.11809812 PMC 134649 · doi ↗ · pubmed ↗
- 5Bannister AJ et al 2001. Selective recognition of methylated lysine 9 on histone H 3 by the HP 1 chromo domain. Nature. 410:120–124. 10.1038/35065138.11242054 · doi ↗ · pubmed ↗
- 6Bischof J et al 2007. An optimized transgenesis system for Drosophila using germ-line-specific phi C 31 integrases. Proc Natl Acad Sci U S A. 104:3312–3317. 10.1073/pnas.0611511104.17360644 PMC 1805588 · doi ↗ · pubmed ↗
- 7Blattes R et al 2006. Displacement of D 1, HP 1 and topoisomerase II from satellite heterochromatin by a specific polyamide. EMBO J. 25:2397–2408. 10.1038/sj.emboj.7601125.16675949 PMC 1478169 · doi ↗ · pubmed ↗
- 8Brennecke J et al 2007. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell. 128:1089–1103. 10.1016/j.cell.2007.01.043.17346786 · doi ↗ · pubmed ↗