Cross-kingdom RNAi: a universal mechanism of inter-organismal communication with many unknowns
Loukia M Kellari, Kalliope K Papadopoulou, Athanasios Dalakouras

TL;DR
This review explores how small RNA molecules can transfer between organisms of different kingdoms, influencing gene activity, but many details about how this works remain unknown.
Contribution
The paper distinguishes between natural and transgene-based cross-kingdom RNAi and identifies key unresolved questions about its mechanisms.
Findings
Cross-kingdom RNAi occurs between plants and their interacting organisms, affecting gene silencing.
The transfer and amplification mechanisms of RNAi signals are not fully understood.
Both natural and transgene-based cases of ck-RNAi have been observed in plants.
Abstract
Cross-kingdom RNAi (ck-RNAi) is a biological process in which small RNA (sRNA) molecules are transferred between organisms belonging to different kingdoms to silence specific genes. Although numerous instances of reciprocal ck-RNAi have been documented in plants, demonstrating a modulation of the interaction between plants and their pathogens, pests, or symbiotic partners, the underlying molecular mechanisms remain largely elusive. In this review, we distinguish between naturally occurring and transgene-based cases of ck-RNAi, examine the diverse mechanisms governing the transfer of primary ck-RNAi signals from donor to recipient organisms, and explore the prerequisites for their amplification and systemic spread. Finally, we highlight key unresolved questions concerning the mechanistic basis of ck-RNAi and offer a perspective on its potential role in co-evolutionary dynamics. This…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
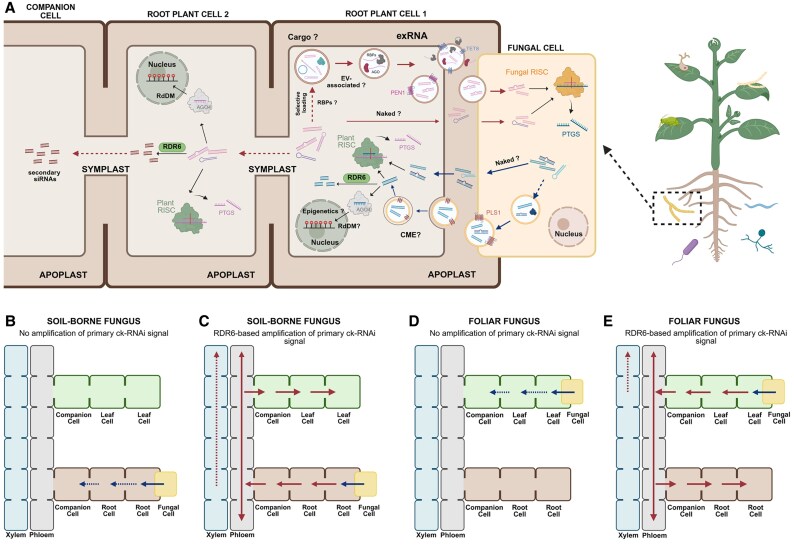 Fig. 1
Fig. 1| From | To | Cargo | EV associated? | Predicted or validated ck-RNAi | Reference |
|---|---|---|---|---|---|
| From plants to microbes and insects | |||||
|
|
| miRNAs, siRNAs | Inside EVs (TET8+) | Validated |
|
|
|
| siRNAs | EV-associated | Predicted |
|
|
|
| mRNA | Inside EVs (TET8+) |
| |
|
|
| Endogenous tyRNAs (10–17 nt) | Outside EVs (PEN1+, PEN3+) associated with RBPs | Predicted |
|
| Inside EVs (TET8+) | |||||
|
|
| sRNAs, long RNAs, AGO1, AGO2 | Outside of high density EVs (PATL1+) | NA |
|
| Medium density EVs (TET8, RIN4, PATL1, ACTIN) | |||||
| Low density EVs (RIN4, PATL1, PEN1, PEN3) | |||||
|
|
| miRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Predicted |
|
|
|
| miRNAs | NA | Validated |
|
|
|
| miRNAs | EV-associated | Validated |
|
|
|
| miRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Validated |
|
|
|
| miRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Validated |
|
| From microbes and insects to plants | |||||
|
|
| sRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Validated |
|
|
|
| sRNAs | NA | Validated |
|
|
|
| sRNAs | Inside EVs (PLS1+) | Validated |
|
|
|
| sRNAs | NA | Predicted |
|
|
|
| tRFs | NA | Validated |
|
|
|
| sRNAs | NA | Validated |
|
|
|
| miRNA | NA | Predicted |
|
|
|
| miRNAs | NA | Predicted |
|
|
|
| miRNA | NA | Predicted |
|
|
|
| miRNA | NA | Validated |
|
|
|
| sRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Validated |
|
|
|
| sRNAs | NA | Predicted |
|
|
|
| sRNAs | NA | Predicted |
|
|
| Soybean | Proteins, lipids | OMV-associated |
| |
|
|
| miRNAs | NA | Predicted |
|
|
|
| miRNAs | NA | Validated |
|
|
|
| sRNAs | Inside OMVs | Validated |
|
- —Horizon Europe project RATION
- —Research Committee of UTH
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Virus Research Studies
Introduction
The ever-moving small RNAs
RNA silencing, also known as RNA interference (RNAi) or post-transcriptional gene silencing (PTGS), is a conserved regulatory mechanism first observed in plants and later in fungi (‘quelling’) and animals (RNAi) (Napoli et al., 1990; Fire et al., 1991; Romano and Macino, 1992). It is initiated by dsRNA, which is processed by DICER-like (DCL) enzymes into small RNAs (sRNAs) that associate with ARGONAUTE (AGO) proteins to guide mRNA cleavage or translational inhibition (Agrawal et al., 2003; Bartel, 2005; Fabian et al., 2010; Jouravleva and Zamore, 2025). In plants, sRNAs can also direct DNA methylation through RNA-directed DNA methylation (RdDM) (Dalakouras and Wassenegger, 2013; Matzke and Mosher, 2014; Erdmann and Picard, 2020; Gallego-Bartolomé, 2020). Moreover, in plants, nematodes, and some fungi, RNA-DEPENDENT RNA POLYMERASEs (RDRs) amplify silencing via transitivity, generating secondary sRNAs from aberrant RNAs in a self-reinforcing cycle (Gazzani et al., 2004; Wassenegger and Krczal, 2006; Baeg et al., 2017; De Felippes and Waterhouse, 2020).
Conserved among all kingdoms are the two main classes of sRNAs, namely the siRNAs and miRNAs. siRNAs derive from perfectly complementary dsRNA precursors, originating either endogenously or exogenously from viruses or transposable elements, with a role in maintaining genome integrity and defense against viruses, transposons, and transgenes (Carthew and Sontheimer, 2009; Borges and Martienssen, 2015; Piombo et al., 2024). Several variants of these siRNAs do exist in plants, each with its particular function and role: epigenetically activated siRNAs (ea-siRNAs), natural antisense siRNAs (nat-siRNAs), trans-acting siRNAs (ta-siRNAs), phased siRNAs (pha-siRNAs), pollen-siRNAs, tapetum-enriched siRNAs (tap-siRNAs), UV-induced siRNAs (uv-siRNAs), and virus-activated siRNAs (va-siRNAs) (Martínez De Alba et al., 2013; Vaucheret and Voinnet, 2024). The second class of sRNAs, miRNAs, derive from endogenous genes, transcribed by RNA polymerase II to imperfect hairpin primary miRNAs, which are eventually processed by DCLs into mature 20–21 nt miRNAs that function as regulators in physiological processes, such as growth and development, immunity, and response to biotic and abiotic stresses (Mallory et al., 2008; Voinnet, 2009). Besides these two canonical sRNA classes, non-canonical sRNAs that may not fulfill the criteria of bona fide siRNAs or miRNAs do exist. Perhaps one of the most important of these is tRNA-derived fragments (tRFs), encountered in both prokaryotes and eukaryotes; tRFs have a size of 17–40 nt, originate from nucleolytic cleavage (not necessarily by DICER) of mature or precursor tRNAs, and, among its other roles, can also associate with AGO and target transcript cleavage (Keam and Hutvagner, 2015; Alves and Nogueira, 2021).
RNAi is not cell autonomous. Thus, in plants, sRNAs (both siRNAs and miRNAs) can move short distances to adjacent cells via the plasmodesmata or long distances via the vasculature to distant parts of the plant to establish systemic RNAi (Baulcombe, 2004; Melnyk et al., 2011; Devers et al., 2020; Voinnet, 2022, 2025). sRNA movement in plants follows the path of the photoassimilates in the ‘source-to-sink’ direction (Tournier et al., 2006; Kobayashi and Zambryski, 2007). During cross-kingdom RNAi (ck-RNAi; Fig. 1Α), sRNAs are suggested to be transported via extracellular vesicles (EVs), and EV-associated sRNAs have been isolated from the apoplast, underpinning the notion that a mechanism of apoplastic sRNA communication cannot be excluded (Baldrich et al., 2019; Voinnet, 2025). In fungi, intercellular transfer of sRNAs can be achieved through the septal pore, a plasmodesmata-like structure associated with endoplasmic reticulum (ER), or the desmotubule, a membranous cell wall-spanning structure that enables sRNA movement, either in naked form or enclosed in vesicles to move throughout the whole mycelial body (Wang and Dean, 2020). In nematodes, systemic RNAi requires the transmembrane proteins SYSTEMIC RNAi DEFICIENT (SID) (Winston et al., 2002, 2007). Insects encode SID-like genes; coleopterans may even have two or three, but Dipterans seem to lack them altogether, perhaps accounting for the fact that the former are much more prone to RNAi than the latter. Yet, Tribolium exhibits systemic RNAi, which is independent of SID-like genes, suggesting that alternative systemic mechanisms do exist (Bucher et al., 2002; Tomoyasu et al., 2008). Systemic RNAi can also be observed in mammals, with sRNA movement being enabled via EVs, RNA-binding proteins, high-density lipoproteins (HDLs), and cell–cell structures, such as gap junctions or synapses (Soutschek et al., 2004; Mittelbrunn and Sánchez-Madrid, 2012).
Summary of the molecular mechanisms underlying ck-RNAi. (A) Bi-directional sRNA translocation between aplant and a soil-borne fungus. Plant sRNAs (miRNAs, siRNAs, and tyRNAs) may be selectively packaged into PEN1/TET8-positive extracellular vesicles (EVs) (Cai et al., 2018; Wang et al., 2024; Koch et al., 2025; Ravet et al., 2025), potentially through the involvement of RNA-binding proteins (RBPs), or exist on the outer surface of EVs either freely or protein bound. They may also be transferred directly in ‘naked’ form to the fungal cells. Inside the fungus, these plant sRNAs are loaded onto the fungal AGO/RISC (RNA-induced silencing complex), triggering post-transcriptional gene silencing (PTGS) of corresponding fungal genes. Reversely, fungal sRNAs can be packaged inside PLS1-positive EVs (He et al., 2023) and incorporated by the plant through clathrin-mediated endocytosis (CME) or even transferred in ‘naked’ form through a mechanism that is not clear. Inside the plant, the fungal sRNAs hijack the RNAi machinery of the host, triggering PTGS. Depending on their target, they may also recruit RNA-dependent RNA polymerase 6 (RDR6) which amplifies the primary sRNAs into secondary sRNAs. Primary or secondary sRNAs having a size of 24 nt may be loaded on host AGO4 and enter the nucleus to trigger RNA-directed DNA methylation (RdDM). Primary and secondary sRNAs can travel through the symplast (via plasmodesmata) to neighboring cells, triggering further PTGS, additional production of RDR6-dependent secondary sRNAs, and reinforcement of RdDM. While primary sRNAs will eventually be diluted away after moving through a certain number of cells, the ever-amplified secondary sRNAs can travel further and reach companion cells in the phloem, to eventually be systemically transported to distant plant parts. Created in BioRender. Kellari, L.M. (2025) https://BioRender.com/spyc57r. (B and C) sRNA translocation from soil-borne fungi to plant cells. Upon contact of the fungus with the plant root, fungal sRNAs are transferred into root cells. If the fungal sRNAs are not RDR6 amplified (blue arrows) (B), they can travel to a few neighboring root cells through the symplast, before their signal diminishes. In contrast, when the primary fungal sRNAs (blue arrows) are RDR6 amplified (C), the resulting secondary sRNAs (red arrows) can reach companion cells and travel through the phloem from source to sink; whether sRNAs can also be transported apoplastically and eventually move through the xylem to the upper parts of the plant is not clear. (D and E) sRNA transmission from foliar fungi to plant cells. Upon contact of the fungus with the plant leaves, fungal sRNAs are translocated into plant cells. If the fungal sRNAs are not RDR6 amplified (blue arrows) (D), they may travel to a few neighboring root cells through the symplast, before their signal diminishes. In contrast, when the primary fungal sRNAs (blue arrows) are RDR6 amplified (E), the resulting secondary sRNAs (red arrows) can reach companion cells and travel through the phloem; whether any apoplastic transportation takes place, is not clear. Created in BioRender. Kellari, L.M. (2025) https://BioRender.com/pfmbowp, https://BioRender.com/umkraom.
Breakthrough studies during recent years have demonstrated that sRNAs can be translocated not only inside an organism but also between organisms belonging to different kingdoms, in a phenomenon termed ck-RNAi (Weiberg et al., 2013; Zeng et al., 2019; Chen et al., 2023; Mahanty et al., 2023; Qiao et al., 2023; Cheng et al., 2025). Focusing on plants, naturally occurring ck-RNAi has emerged as a prominent mechanism of interspecies communication and has been observed during the molecular warfare between plants and their interacting pathogenic organisms (Weiberg et al., 2013; Buck et al., 2014; K. Zhu et al., 2017; Cai et al., 2018; Silvestri et al., 2025). A clear distinction should be made between ck-RNAi and sRNA translocation within the plant kingdom, which has also been demonstrated between the parasite Cuscuta campestris and its host Arabidopsis thaliana (Shahid et al., 2018), or even between two neighboring Arabidopsis plants (Betti et al., 2021). In this review, we will focus only on ck-RNAi and discuss the most important cases of bi-directional ck-RNAi between plants and microbes and insects. Even before the discovery of natural ck-RNAi cases, artificial or transgene-based ck-RNAi methods were developed, mainly to be used as a powerful tool in crop protection platforms (Koch and Kogel, 2014; Koch and Wassenegger, 2021). Such approaches, including host-induced gene silencing (HIGS), spray-induced gene silencing (SIGS), and microbe-induced gene silencing (MIGS), are also briefly presented here. Further, we investigate the mechanisms that regulate the transfer of primary ck-RNAi signals between donor and recipient organisms, as well as the conditions required for their amplification and systemic dissemination (Fig. 1). Lastly, we emphasize the major open questions surrounding the mechanistic foundations of ck-RNAi.
Cross-kingdom RNAi: natural cases
Plants translocate sRNAs to microbes and insects
Plants transmit sRNAs to pathogenic microbes and pests as a natural defense strategy (Table 1). Diverse plant–fungal pathosystems have been reported: upon Verticillium dahliae infection, cotton plants increase the production of miR166 and miR159 and translocate them to the pathogen to silence genes essential for fungal virulence, namely a Ca^2+^-dependent cysteine protease (Clp-1) and an isotrichodermin C-15 hydroxylase (HiC-15) (Zhang et al., 2016); upon Botrytis cinerea infection, A. thaliana transmits sRNAs that target genes related to vesicular trafficking, inhibiting pathogen infection (Cai et al., 2018); upon Fusarium graminearum infection, wheat secretes miR1023 that silences the alpha/beta hydrolase gene of the pathogen, suppressing its invasion (Jiao and Peng, 2018); upon Puccinia striiformis infection, wheat translocates sRNAs to the pathogen to silence various genes (Mueth and Hulbert, 2022). Similarly, comparative analysis of transcriptome and sRNA expression patterns in the Brachypodium distachyon—Magnaporthe oryzae pathosystem predicted (but did not validate) the presence of several plant sRNAs silencing various fungal genes (Zanini et al., 2021). Natural ck-RNAi was validated for the Malus hupehensis—Botryosphaeria dothidea pathosystem, where miR159a from apple silences the sugar transporter gene BdSTP of B. dothidea, affecting fungal growth and proliferation (Yu et al., 2024). Recently, it was shown that upon B. cinerea infection, tomato sends sRNAs and silences serine/threonine kinase genes related to pathogenicity (Cheng et al., 2025). Beyond fungal interactions, during the infection of Arabidopsis by the oomycete Phytophthora capsici, plant secondary siRNAs (siR1310) are translocated to silence P. capsici transcripts (Hou et al., 2019).
So far, only a few cases of sRNA transmission from plants to beneficial microbes have been recorded. One example is the in silico prediction of the putative translocation of miRNAs from two Populus species (Populus deltoides and P. trichocarpa) to either an ectomycorrhizal fungus Laccaria bicolor or an arbuscular mycorrhizal fungus Rhizophagus irregularis during root colonization (Mewalal et al., 2019). The predicted fungal gene targets of the Populus-derived miRNAs include transport proteins, transcription factors, and several genes encoding proteins of unknown function (Mewalal et al., 2019).
For plant-to-microbe sRNA translocation to occur, it is not sufficient for the plant to produce and export sRNAs; the microbe must also be capable of perceiving these exogenous molecules. Indeed, the capacity to take up exogenous RNA is an essential requirement for a ck-RNAi event that not all fungi can manifest. For example, Colletotrichum gloesporioides is resilient to exogenous RNA uptake (Qiao et al., 2021), in contrast to its close relative C. truncatum (Gu et al., 2019). Zymoseptoria tritici is also resilient to exogenous RNA uptake; not surprisingly, a study investigating its interaction with wheat concluded that ck-RNAi was absent (Kettles et al., 2019). However, fungal uptake of exogenous RNA must be distinguished from uptake of plant-derived RNA, as the latter can be delivered through EVs that facilitate sRNA internalization. Notably, even C. gloeosporioides, which is largely resistant to direct uptake of exogenous sRNAs, is susceptible to HIGS, indicating that EV-mediated delivery can overcome this barrier (Mahto et al., 2020). An additional prerequisite for ck-RNAi seems to be the presence of the core RNAi machinery in the recipient organism. Most fungi, indeed, do encode RNAi genes (DCLs and AGOs), but these genes are not universally present across all fungal species. In a notable case, Ustilago maydis lacks the core RNAi machinery (no DCL, AGO, or RDR), in contrast to its close relative U. hordei (Laurie et al., 2008). Yet, even bacteria, which lack canonical RNAi machinery, have been reported to exhibit susceptibility to HIGS (Ravet et al., 2025). Whether this represents an isolated case or indicates a broader phenomenon remains unresolved.
Besides sending sRNAs to fungi and oomycetes, plants may also translocate sRNAs to bacteria and insects. In silico analysis predicts that Arabidopsis translocates several sRNAs, including tRFs, that are taken up by Pseudomonas syringae to silence bacterial genes (Ravet et al., 2025). This was particularly striking, since bacteria were traditionally regarded as not being susceptible to RNAi. Yet, we should perhaps revisit this assumption, since in the same study it was shown that transgenic Arabidopsis plants expressing hairpin-derived sRNAs against P. syringae silenced the corresponding bacterial genes (see also below, ‘Host-induced gene silencing’) (Ravet et al., 2025). Insects encode a well-studied RNAi pathway, and most species are susceptible to exogenous RNAs (Joga et al., 2016; Dalakouras et al., 2024). Plant miRNAs added in larval food regulated caste development of honeybees; when Brassica campestris miR162a was added in honeybee larval food, it targeted amTOR, a stimulatory gene in caste differentiation, preventing larval differentiation (K. Zhu et al., 2017). Interestingly, plant miRNAs in larval food also affected Drosophila melanogaster, causing extended developmental times and reductions in body weight and length, ovary size, and fecundity (K. Zhu et al., 2017). Arabidopsis miRNAs were reported to break the barrier of the insect mid-gut and enter the circulatory system of Plutella xylostella to silence hemocyanin domain-containing genes, eventually regulating insect development (Zhang et al., 2019). Of note, an in silico study of sRNA sequencing data from the gut of the sup-feeding aphid Myzus persicae reveals the presence of 32 miRNAs in aphid gut samples, mapping not only to the genome of the host plant but also to predicted gene targets related to transcription of the aphid (Thompson et al., 2019).
Microbes and insects translocate sRNAs to plants
Ck-RNAi can be bi-directional, namely transmission of sRNAs can also occur from microbes to their plant hosts (Table 1). In a pivotal study, it was shown that B. cinerea sRNAs bind to Arabidopsis AGO1 protein and silence host immunity genes (Weiberg et al., 2013). A few years later, a Botrytis sRNA effector (Bc-siR37) was identified that silences Arabidopsis defense-related genes (M. Wang et al., 2017). In a similar fashion, the fungal pathogen Puccinia striiformis produces a novel miRNA-like (milR1) that suppresses the immune-related PR2 gene during the interaction with wheat (B. Wang et al., 2017). Moreover, Puccinia triticina translocates milR1, milR2, and milR3 to wheat to target host mitogen-activated protein (MAP) kinase, calmodulin, and F-box protein genes, respectively (Dutta et al., 2019). Notably, a Fusarium oxysporum milR1 that is produced upon infection of Solanum lycopersicum was found to associate with plant AGO4a protein to regulate the expression of SlyFGR4, a gene implicated in defense against the pathogen (Ji et al., 2021). The mechanistic details of sRNA uptake are not clear (see below), but a recent study showed that B. cinerea sRNAs ride in EVs to enter Arabidopsis cells through a clathrin-mediated endocytosis pathway (He et al., 2023). Besides these cases, in silico studies have predicted that Blumeria graminis f.sp. hordei and B. graminis f.sp. tritici may send sRNAs to barley and wheat, respectively, to silence various genes (Kusch et al., 2018). Similar predictions on sRNA translocation from pathogen to host have been made in the case of Sclerotinia sclerotiorum and Arabidopsis (Derbyshire et al., 2019). sRNA translocation from fungal pathogens is not restricted to herbaceous species; Valsa mali milR1 and siR1 were shown to silence apple receptor-like kinase and disease resistance-related genes, respectively (Xu et al., 2022; Liang et al., 2024).
Research on sRNA transmission from symbionts to host plants is still limited, but is gradually expanding. The first example was an in silico prediction of sRNAs from R. irregularis with putative mRNA targets in the host M. truncatula, revealing a potential role during symbiosis (Silvestri et al., 2019). Recently, the same research group validated that Rir2216 targets M. truncatula WRKY transcription factor 69 (MtWRKY69) to facilitate successful colonization (Silvestri et al., 2025). Another mycorrhizal fungus, Pisolithus microcarpus, upon colonization of Eucalyptus grandis expresses novel miRNAs, including miR-8, which is transferred to E. grandis roots and silences NB-ARC domain-containing genes to establish symbiosis (Wong-Bajracharya et al., 2022). In silico predictions have revealed probable ck-RNAi phenomena during the interaction of the root-colonizing endophyte Serendipita indica with B. distachyon (Šečić et al., 2021) and miRNA-like molecules from Trichoderma asperellum were predicted to target tomato genes (Y. Wang et al., 2021).
Not only fungi, but also oomycetes, bacteria, and insects translocate sRNAs to plants. Small RNA transmission has also been shown for the oomycete Hyaloperonospora arabidopsidis, which utilizes Arabidopsis AGO1 to silence host genes and thereby enhance its virulence (Dunker et al., 2020). In prokaryotes, the nitrogen-fixing bacterium Bradyrhizobium japonicum produces tRFs that bind to the host AGO1 and silence host target genes during its interaction with Glycine max (Ren et al., 2019). Moreover, Xanthomonas oryzae pv. oryzicola sRNA001 was shown to be loaded on outer membrane vesicles (OMVs) and transported to rice where it silenced the JMT1 gene (Wu et al., 2024), underpinning that our assumptions on the total absence of RNAi-like mechanisms in prokaryotes need urgent revision. Interestingly, insects also translocate sRNAs to their hosts; sRNAs from the whitefly Bemisia tabaci were detected in the phloem of tomatoes, probably transferred during feeding, although their gene targets were not experimentally validated (Van Kleeff et al., 2016). Moreover, a recent study identified salivary miRNAs from the rice plant hopper Nilaparvata lugens, where miR-7-5P was deemed important for feeding and targets rice immunity genes (Zhang et al., 2024).
Cross-kingdom RNAi: artificial cases
Host-induced gene silencing
Even before the documentation of natural sRNA translocation between organisms, RNAi-based technologies were employed in crop protection platforms. Such technologies include HIGS, which utilizes a transgenic plant (host) producing and delivering dsRNAs or sRNAs that target virulence genes to pests or pathogens. Excellent reviews on HIGS are available (Nowara et al., 2010; Koch and Kogel, 2014; Koch and Wassenegger, 2021; Niu et al., 2021; Choudry et al., 2024) and we will not dwell on details here. The most recent studies on HIGS in fungi include maize against Aspergillus flavus (Omolehin et al., 2024), Brassica napus against S. sclerotiorum (Wytinck et al., 2022), soybean against F. oxysporum (Pérez et al., 2021), rice against M. oryzae (L. Zhu et al., 2017), and wheat against F. graminearum (Wang et al., 2020). In the case of oomycetes, HIGS has likewise been explored against Bremia letucae, which causes downy mildew in lettuce (Govindarajulu et al., 2015), as well as Phytophthora infestans, a pathogen of potato (Jahan et al., 2015). More recent studies have demonstrated successful HIGS against insect pests, including maize against Chilo partellus (Adeyinka et al., 2023), cotton against Aphis gossypii (Zhang et al., 2023), tomato against Frankliniella occidentalis (Venkatesh et al., 2023), rice against Chilo suppressalis (Tang et al., 2024), and potato against M. persicae (Murtaza et al., 2022). Applications of HIGS extend further to nematodes—soybean lines expressing chitin synthase gene and tobacco plants expressing different chitin biosynthesis genes were rendered resistant against Heterodera glycines (Kong et al., 2022) and Meloidogyne incognita (Mani et al., 2020), respectively.
Apart from its importance as a plant protection method, HIGS has contributed to the elucidation of the molecular mechanisms that underlie ck-RNAi (reviewed in Koch and Wassenegger, 2021; Zand Karimi and Innes, 2022). For example, fungi preferentially take up long dsRNA molecules, with only some cases showing sRNA uptake (Koch and Kogel, 2014; Baulcombe, 2015; Zand Karimi and Innes, 2022). Herbivorous insects preferably take up long dsRNAs, which are then processed by their RNAi machinery (Koch and Wassenegger, 2021; Zand Karimi and Innes, 2022). In the case of sup-feeding insects such as aphids, HIGS mainly relies on sRNA rather than dsRNA uptake (Zand Karimi and Innes, 2022).
Spray-induced gene silencing
An alternative to HIGS that relies on the exogenous application of dsRNAs or sRNAs to crops instead of using transgenic plants has been developed and termed SIGS (Dalakouras et al., 2020; Chen et al., 2023; Chen et al., 2025; Dubrovina et al., 2025). In contrast to HIGS, SIGS seems to be a less efficient RNAi strategy since the sRNA/dsRNA application is transient, and repetition of application may be required. Yet SIGS can be more effective than HIGS in some cases, as has been shown in barley against F. graminearum (Koch et al., 2019). Moreover, SIGS seems to gain ground due to concerns on GMO (genetically modified organism) use and the environment (Dalakouras et al., 2024; Vatanparast et al., 2024; Tardin-Coelho et al., 2025). During SIGS, chemically synthesized sRNAs or in vitro and/or in vivo transcribed dsRNAs that target essential genes for survival and virulence in pests and pathogens are applied to plants. Methods of application include foliar spraying, trunk injection, or root drenching. Indicative SIGS reports include those against tobacco mosaic virus (Niehl et al., 2018), potato spindle tuber viroid (Carbonell et al., 2008), F. graminearum (Koch et al., 2016), P. infestans (Kalyandurg et al., 2021), Leptinotarsa decemlineata (Rodrigues et al., 2021), Tetranychus urticae (Q. Wang et al., 2025), and weeds such as Amaranthus cruentus (Hendrix et al., 2021). A critical parameter for the efficacy of SIGS is the formulation of exogenous RNA, in order to increase its uptake from the target organism and protect it from environmental degradation; lipid double hydroxide (LDH) nanoclays and carbon dots are some of the most promising formulation agents for use in these cases (Mitter et al., 2017; Schwartz et al., 2020).
Microbe-induced gene silencing
Upon the discovery of ck-RNAi between host and microbial pathogens, the interspecies RNAi (is-RNAi) between rhizospheric microorganisms was also investigated, resulting in the discovery of microbe-induced gene silencing (MIGS) (Wen et al., 2023)—not to be confused with miRNA-induced gene silencing (also called MIGS) (Gauthier et al., 2025). Compared with HIGS, MIGS does not require genetic manipulation of the host plant; compared with SIGS, MIGS does not rely on the use of nanomaterials to carry chemically synthesized RNAs. Still, MIGS is based on the use of a genetically modified microorganism, making it rather unsuitable for field application, at least under the current regulatory landscape. Wen et al. (2023), who first introduced the term MIGS, exploited the beneficial fungus Trichoderma harzianum to express dsRNA targeting the O-mannosyltransferase (PMT) gene, a well-known fungicide target, in V. dahliae and F. oxysporum. Growth inhibition of both pathogens was demonstrated in both in vitro and in planta experiments, with cotton plants showing protection against infection, when treated with the transformed T. harzianum strain (Wen et al., 2023). Recently, MIGS was used to protect maize plants against F. graminearum, proving the applicability of the technology to various crops (Chen et al., 2025). Another use of the MIGS-like case was shown by Porquier et al. (2021), where a B. cinerea strain lacking retrotransposon-derived sRNAs was complemented with the relative genes to reinstate their expression. This has led to enhanced infection and suppression of host defense-related genes, suggesting that retrotransposons are pathogenicity factors that manipulate host plant gene expression by trans-species sRNAs. In our own previous work, a variation of MIGS was implemented in a non-pathogenic, root-residing Fusarium solani strain K (FsK) fungal endophyte. Using a proof-of-concept system that consisted of an RNAi trigger [transgenic FsK expressing host green fluorescent protein (GFP)] and an RNAi-sensor (transgenic Nicotiana benthamiana expressing GFP), systemic silencing and DNA methylation of the host GFP was recorded upon N. benthamiana colonization with FsK (Dalakouras et al., 2023; Kellari et al., 2025).
ck-RNAi in plant and non-plant systems
A common feature between ck-RNAi events in plant and animal systems is the delivery of microbial sRNAs aimed at modulating host immunity to promote infection (Weiberg et al., 2013; B. Wang et al., 2017). In mammals, Heligmosomoides polygyrus and Heligmosomoides contortus, two gastrointestinal nematodes, secrete miRNAs that suppress immunity in mice or murine and ovine organoids, respectively (Buck et al., 2014; Perez et al., 2025), but also during the interaction of a pathogenic bacterium suppressing immunity in human airway epithelial cells (Koeppen et al., 2016). The same was also observed during sRNA transmission from the entomopathogenic fungus Beauveria bassiana to Anopheles stephensi (Cui et al., 2022), reinforcing the conserved nature of this mechanism. In addition, hosts fight back and produce sRNAs to reduce the virulence of the parasites or pathogens, exemplified in this case by the transfer of sRNAs from A. stephensi to B. bassiana (Wang et al., 2021). In an approach similar to MIGS in plants, a pathogen-mediated RNAi (pmRNAi) of B. bassiana transformed to express Aedes aegypti miRNAs (aaemiR-8 and aae-miR-375) acted as a negative regulator of the mosquito Toll immune pathway, thereby increasing fungal virulence (Cui et al., 2022).
One probable common mechanism of sRNA trafficking between different organisms in both plant and animal interactions is through EVs (Fig. 1A). EVs are membrane-bound, phospholipid vesicular structures produced by both prokaryotic and eukaryotic cells, carrying nucleic acids, proteins, and lipids (Maas et al., 2017; Sall and Flaviu, 2023). In recent years, their potential role in inter-organismal interactions has attracted research interest. During plant–pathogenic microbe interactions, plant EVs containing sRNAs (Cai et al., 2018; He et al., 2023; Ravet et al., 2025) and even mRNAs (Wang et al., 2024) were reported to be secreted. Bacteria can also produce OMVs, the bacterial analog of EVs, in pathogenic or mutualistic contexts. The phytopathogen X. oryzae pv. oryzicola produces the sRNA Xosr001d inside OMVs, which down-regulates rice immunity genes (Wu et al., 2024), while Sinorhizobium fredii (now renamed Ensifer fredii) OMVs applied to soybean roots up-regulate nodulation and early symbiosis genes and down-regulate defense-related genes (Li et al., 2022). In the animal kingdom, studies also suggest that sRNAs can move bi-directionally encapsulated in EVs from either the host or the pathogen. For example, in mammals, EVs were reported in the case of Pseudomonas aeruginosa and human airway epithelial cells (Koeppen et al., 2016, 2021). EVs mediate the transfer of sRNAs between human monocyte cells and Candida albicans (Halder et al., 2022). Nematodes are also capable of transferring EVs to their hosts, as observed in D. melanogaster, where they lead to down-regulation of antimicrobial peptide genes (Toubarro et al., 2025), and in mouse cells, where they modulate host gene expression related to immunity and inflammation (Buck et al., 2014).
In contrast to what happens with plant miRNAs, which directly cleave the target genes in pathogens (Zhang et al., 2016; Cai et al., 2018; Jiao and Peng, 2018), a study showed that human erythrocyte miRNAs, when taken up by P. falciparum, which lacks orthologs for AGO and DCL, inhibited mRNA translation by impairing their loading onto ribosomes (LaMonte et al., 2012). A similar mechanism of non-canonical RNAi was also shown for T. harzianum sRNAs targeting V. dahliae through translational inhibition instead of mRNA cleavage (Wen et al., 2023), although the exact mechanism was not uncovered. These examples illustrate that, although ck-RNAi is conserved across kingdoms, its mechanistic implementation varies widely, indicating the need for further investigation.
Exploring the unknowns of ck-RNAi
Many key aspects of ck-RNAi in plants remain elusive, in particular as regards the movement of sRNAs between interacting organisms, their transport in distal parts of the host plant, as well as the factors determining their efficacy and specificity. Some of the current knowledge gaps that need to be addressed and clarified are summarized below.
Are translocated sRNAs inside or outside EVs?
EVs not only protect RNA in the extracellular space where RNA and protein degradation enzymes are abundant, but also facilitate its transfer into destination cells (Hu et al., 2025). Notwithstanding the recent developments in research on EVs and their role in sRNA translocation, the question of how the sRNAs are loaded inside EVs in the origin organism remains largely unresolved. In mammalian systems, several factors have been proposed, including sequence composition and structural features of the RNAs; involvement of RNA-binding proteins (RBPs); and different modifications either in the RNAs themselves or in the proteins they interact with, all of which could contribute to sorting (Dellar et al., 2022). The selective loading of sRNAs inside EVs indicates that this process is also regulated in plants (Cai et al., 2018; Baldrich et al., 2019). It has been demonstrated that fungal sRNAs ride in EVs to enter plant cells through clathrin-mediated endocytosis (He et al., 2023), although it is not clear whether clathrin-independent mechanisms also contribute in this process. However, the question is are the sRNAs inside or outside EVs. Whereas RBPs, like AGO1, RNA helicases, and annexins, can aid the transfer and/or stabilization of sRNAs inside EVs (Hagiwara et al., 2015; He et al., 2021), complexes of sRNAs with RBPs have also been found outside of EVs or in close proximity to the surface of EVs. Zand Karimi et al. (2022), showed that Arabidopsis apoplastic sRNAs are mainly located on the outside of EVs but associated with proteins that protect them against degradation. Similarly, Ravet et al. (2025), by applying various enzymatic treatments that degrade unprotected RNAs or proteins, reported that a significant titer of active sRNAs from apoplastic fluids are located on the outside of EVs (Fig. 1A). In a recent study in Arabidopsis, high-resolution density gradient ultracentrifugation classified the EVs into three categories based on their densities, noting that only the medium- and low-density fractions contain pure EVs associated with known markers, although sRNAs were found in the high-density fraction (Koch et al., 2025). Since in most of the aforementioned studies EVs were isolated using different methods, resulting in the enrichment of different EV subpopulations, improving the accuracy of EV isolation methods will clearly minimize the risk of data misinterpretation. A sound assessment of the quality and purity of the EVs requires combined analysis of TEM, nanoparticle tracking analysis (NTA), and western blotting (Eldahshoury et al., 2024). Intriguingly, it was recently found that various RNAs (tRNAs, rRNAs, mRNAs, miRNAs, and sRNAs), which are neither apoplastic nor in EVs or in protein complexes, are secreted directly onto the leaf surface rather than exuded through stomata or hydathodes (Borniego et al., 2025), suggesting that diverse pathways of sRNA transport exist.
Are sRNAs translocated as free molecules or AGO bound?
In plants, it is well demonstrated that 21, 22, and 24 nt sRNAs move systemically as AGO-free sRNA duplexes (Devers et al., 2020) and, similarly during intraspecies RNAi between plants, miRNAs are translocated in AGO-free form (Betti et al., 2021). During ck-RNAi between Botrytis and Arabidopsis, fungal sRNAs were reported to be translocated in AGO-free form (Weiberg et al., 2013), while, upon uptake of the sRNA molecules, the use of the AGO proteins of the recipient organism seems to be a widespread phenomenon. This is true for tomato sRNAs binding to fungal AGOs (Cheng et al., 2025), fungal (Weiberg et al., 2013; Ji et al., 2021) and oomycete sRNAs binding to plant AGOs (Dunker et al., 2020), and even symbiotic bacterial sRNAs engaging with plant AGOs (Ren et al., 2019). In animal systems, miRNAs from B. bassiana interact with mosquito AGO1, hijacking the RNAi machinery (Cui et al., 2022). Thus, the overwhelming available experimental evidence suggests that sRNAs are translocated to plants as free molecules and are loaded on the host AGOs. In support of this argument comes the fact that during SIGS, the sprayed molecules are naked and not AGO-loaded sRNAs, but still exhibit biological activity once present in the host, indicating that they were eventually loaded on host AGOs.
Onto which host AGO are the translocated sRNAs loaded?
Even if we assume that sRNAs are translocated as AGO-free molecules, it is not clear to which host AGO they will eventually be loaded. In plants, phylogenetic analyses have revealed three major clades of AGO proteins, which are named after Arabidopsis AGOs: AGO1/5/10, AGO2/3/7, and AGO4/6/8/9 (Fang and Qi, 2016). Thus, an AGO-free microbial sRNA, newly introduced into a plant cell, may be offered a variety of AGOs to be loaded onto. The sorting of sRNAs onto plant AGOs is primarily dictated by their size and the nucleotide at their 5′ terminus (Mi et al., 2008). Hence, 21 nt sRNAs with 5′ U are loaded on AGO1 (the main mediator of PTGS), those with 5′ A on AGO2, and those with 5′ C on AGO5, while 24 nt sRNAs with 5′ A are loaded on AGO4/6/9 (the main mediators of RdDM). There is no reason to assume that exogenous/microbial sRNAs do not follow this biochemical rule once present in the plant environment; yet this hypothesis remains to be demonstrated and a comprehensive study analyzing their AGO association (e.g. with AGO-IP seq) and deciphering their eventual biological activity in the host is still lacking.
Are all translocated sRNAs loaded on host AGOs?
In Arabidopsis, miRNAs differ in their AGO loading efficiency, and AGO abundance can be a limiting factor for miRNA incorporation (Dalmadi et al., 2019). Similar constraints are likely to affect ck-RNAi: in cells where AGO levels are limited, not all translocated sRNAs may be loaded onto AGO, reducing ck-RNAi efficacy locally. However, sRNAs that fail to load may remain mobile and may still be incorporated into AGO complexes in adjacent cells, where they can initiate ck-RNAi. Consistent with this hypothesis, recent work in Arabidopsis indicates a negative relationship between AGO loading and mobility, with miRNAs remaining mobile precisely because they are not loaded (Fan et al., 2022; Gonzalo et al., 2025). Together, these observations suggest that non-cell-autonomous ck-RNAi may rely on the stepwise association of mobile sRNAs with AGO in recipient cells, ensuring a continuous pool of unbound sRNAs capable of further intercellular movement.
What is the role of sRNA modification?
Which sRNAs are eventually translocated? All or some of them? It seems counterintuitive that sRNAs are indiscriminately loaded on EVs. Indeed, it has been suggested that in the Arabidopsis–Botrytis pathosystem, although the more abundant sRNAs are more likely to be transported through EVs, there is a clear selection in transferred sRNAs (Cai et al., 2018). A molecular tag could be envisaged to somehow recruit those sRNAs that will preferentially be loaded on EVs over others. Interestingly, it has been recently shown that small and long RNAs associated with EVs from the leaf apoplast are enriched in N^6^-methyladenosine (m6A) modification (Zand Karimi and Innes, 2022), suggesting that such a type of modification might affect sRNA sorting on EVs. Whether additional RNA modifications also contribute to such a selection is not clear. In plants, HUA ENHANCER OF SILENCING 1 (HEN1) is an enzyme that modifies sRNAs by 2′-O-methylation, protecting sRNAs from degradation, particularly from 3′-uridylation, thereby increasing their stability and functional life span. Besides plants, HEN1 homologs that methylate sRNAs for protection have also been identified in mouse, zebrafish, and Drosophila (Ji and Chen, 2012). It is not clear whether fungal sRNAs contain this modification and/or whether this modification has any effect on sRNA translocation.
Which types of RNA molecules are translocated?
A growing body of evidence suggests that sRNAs (miRNAs or siRNAs) of sizes typical of the organism of origin (20–24 nt) are the primary transferable molecules (Zhang et al., 2016; Jiao and Peng, 2018; Hou et al., 2019; Silvestri et al., 2025). Notably, Baldrich et al. (2019) show that plant EVs are also enriched in small RNAs of 10–17 nt in length. These tinyRNAs (tyRNAs), as they were termed, represent degradation products originating from multiple sources, and their potential functional roles remain to be determined. Other reports showed the presence of mRNAs enclosed inside EVs of axenic cultures of U. maydis or Arabidopsis (Kwon et al., 2021; Wang et al., 2024), respectively, with the latter being incorporated by fungal cells and associated with polysomes for translation. Similar phenomena have been observed in mammalian systems, where mRNAs packaged within EVs are transported between different tissues and cell types (Valadi et al., 2007; Yokoi et al., 2017). Conversely, Zand Karimi et al. (2022), reported that Arabidopsis extracellular RNAs (exRNAs) did not contain mRNAs, but rather long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs). Similarly in rice, circRNAs are involved in immune responses against the fungal pathogen M. oryzae (Fan et al., 2020). Intriguingly, the question as to whether long dsRNAs (i.e. the precursors of sRNAs) are also translocated has not been addressed so far.
What is the sequence complementarity prerequisite between the translocated sRNA and its target transcript in ck-RNAi?
It is reasonable to assume that, from a plethora of diverse sRNAs being translocated, only a few will exert meaningful biological activity on the recipient organism, namely only those which exhibit sufficient sequence complementarity with a host transcript. Perfect sequence complementarity will probably ensure maximum RNAi efficiency, but it is unclear how the number and occurrence of mismatches will affect ck-RNAi and to what extent. It is generally accepted that a perfect sequence complementarity of a 21 nt sRNA with its target transcript will be likely to result in mRNA degradation, while a single mismatch at position 10–11 may inhibit such cleavage (Iwakawa and Tomari, 2022). A few mismatches in other positions may not influence mRNA cleavage in some cases, but in others may lead to translational inhibition (Brodersen et al., 2008). Translocated sRNAs having a size of 22 nt may also affect host gene expression by translational arrest (Franco-Zorrilla et al., 2007; Wu et al., 2020). Importantly, translocated sRNAs having a size of 24 nt may induce epigenetic modifications such as DNA methylation (Fig. 1A), even if the 24 nt sRNA contains up to four regularly interspaced mismatches compared with the target DNA (Fei et al., 2021). All in all, how the necessary sequence complementarity in each case is maintained is still elusive; additional studies will be required to answer the question of whether it arises stochastically or through co-evolution of the host with the target organism (Knip et al., 2014; Zhao and Guo, 2019; Nien et al., 2024).
Are the primary ck-RNAi signals sufficient for systemic function?
RDR6 seems to be involved not only in the generation but also in the systemic response to the ck-RNAi signal. Indeed, during ck-RNAi from the plant to a fungal pathogen, host RDR6 generates secondary sRNAs that constitute the primary host ck-RNAi signal to be translocated to the fungus. Thus, Arabidopsis rdr6 mutants are more susceptible to Phytophthora infection, and Phytophthora even encodes a suppressor of silencing effector to inhibit secondary sRNA biogenesis in Arabidopsis and to promote infection (Hou et al., 2019). Similarly, Arabidopsis rdr6 mutants were more susceptible to B. cinerea infection, since Arabidopsis secondary sRNAs mediate the host defense against the pathogen (Cai et al., 2018). On the other hand, we have also shown, using N. benthamiana rdr6 CRISPR/Cas mutant plants, that RDR6 is indispensable for the amplification into secondary sRNAs of the primary ck-RNAi signals that were translocated from a root-residing beneficial fungal strain to the host plant, eventually allowing the onset of systemic silencing (Kellari et al., 2025). It appears that the primary ck-RNAi signal does not suffice (e.g. due to degradation or simply dilution) to mediate systemic RNAi in the host (Fig. 1B, D). However, upon RDR6-based amplification, the secondary sRNAs undergoing a self-reinforcing loop will eventually suffice to move not only cell-to-cell through plasmodesmata but also systemically through the phloem (Fig. 1C, E). Whether RDR6-based amplification will occur depends on the nature of both the sRNA and the target mRNA. Indeed, RDR6 is ideally (but not exclusively) recruited to interactions where sRNAs having an asymmetric bulge and/or a size of 22 nt occupy the 5′ end of a transcript originating from an intronless gene (Chen et al., 2010; Christie et al., 2011; Manavella et al., 2012; Dadami et al., 2014; Uslu et al., 2022). Thus, ck-RNAi does not always become systemic, but it may well do if the above criteria are met. The role of RDRs for RNAi amplification is not limited to plant hosts; fungi also encode RDRs and they may also have a role in the amplification of silencing signals, eventually increasing the efficiency of ck-RNAi (SIGS and HIGS, included). Yet, it seems that despite their presence, fungal RDRs do not display the same RNAi amplification role as plant RDRs (Song et al., 2018).
Do translocated sRNAs trigger epigenetic modifications?
At least in plants, RNAi manifests itself not only as mRNA degradation but as DNA methylation as well, with the two pathways being tightly linked (Jones et al., 1999; Taochy et al., 2019; Trasser et al., 2024). Whether ck-RNAi could have a potential role in inducing epigenetic modifications in recipient organisms, especially in plants where the RdDM pathway is well established, remains another as yet uncharted aspect of the mechanism. The 23 nt miRNA-like Fol-milR1 released by F. oxysporum f.sp. lycopersici preferentially binds S. lycopersicum AGO4a to down-regulate expression of a host calcium-binding protein kinase involved in defense signaling; although cleavage of the target transcript was observed, it was unclear whether Fol-milR1 could also trigger RdDM through its association with SlAGO4a (Ji et al., 2021). Loading endosymbiont sRNAs into host AGO4 proteins opens up the prospect of these driving RdDM and even transcriptional gene silencing (TGS), hence allowing long-term regulation of gene expression in the host. Such regulation would have considerable benefits in a mutualism, potentially adapting one or both partners to increase the stability of the interaction (Qiao et al., 2023). Indeed, DNA methylation is essential in Arabidopsis to establish a beneficial relationship with the root-colonizing Trichoderma atroviride (Rebolledo-Prudencio et al., 2022). The importance of DNA methylation in symbiosis is further highlighted in ectomycorrhizal symbiosis, with hypomethylation in the Populus sp. host associated with decreased association with the Laccaria bicolor mycorrhizal fungus (Vigneaud et al., 2023). Solid evidence for epigenetic modifications induced upon ck-RNAi emerged from our previous work, where it was shown that sRNA translocation from a beneficial fungus triggered DNA methylation (in the CG, CHG, and CHH context) of a host reporter gene in an RDR6-dependent manner (Dalakouras et al., 2023; Kellari et al., 2025). Based on the above, it is reasonable to assume that the onset of epigenetic modifications such as DNA methylation and histone modifications during ck-RNAi may be more common than once thought. Should DNA methylation establishment and maintenance in ck-RNAi be confirmed, it could imply that ck-RNAi may have a pivotal role in crop epigenetic plasticity in response to environmental cues; intriguingly, such plasticity could potentially even be used to reveal cryptic diversity for breeding purposes (Dalakouras and Vlachostergios, 2021).
Conclusions: perspectives on co-evolution
From an evolutionary standpoint, RNAi serves as a defense strategy against viruses and transposons, and its cross-kingdom extension similarly functions as defense against invading pathogens or, conversely, as a means for the pathogens to hijack the immune system of the host. It is reasonable to assume that ck-RNAi reflects a co-evolutionary arms race between plants and microbes. sRNA recipients (plants or pathogens) adapt to newly evolved sRNAs from donors (pathogens or plants, respectively) via target gene or sRNA/miRNA locus mutation, and provided that such adaptation is to the general benefit of the organism. Possible mechanisms of sRNA or miRNA evolution may range from single nucleotide polymorphisms of pre-existing loci to de novo emergence of new ones through inverted gene duplication events. Moreover, horizontal gene transfer (HGT), once thought to occur exclusively in prokaryotes, is now accepted also to occur in eukaryotes such as fungi and plants (Fitzpatrick, 2012; Gonçalves et al., 2024; Y. Wang et al., 2025). HGT has been proposed as a driving force of sRNA evolution in bacteria (Dutcher and Raghavan, 2018); whether HGT contributes to sRNA evolution in eukaryotes remains a possibility (Yang et al., 2019). Interestingly during ck-RNAi, besides novel sRNA/miRNA non-coding loci, coding genes may emerge as well; indeed, microbes may develop RNAi suppressors deactivating or affecting the accumulation of plant sRNAs (Hou et al., 2019; Zhu et al., 2022), reminiscent of viral suppressors of RNAi which were themselves developed as a response to the RNAi antiviral strategy of the plant (Burgyán and Havelda, 2011).
sRNA exchange extends beyond antagonistic interactions to further roles during interspecies communication, since symbiotic and beneficial microorganisms also use ck-RNAi when colonizing their host plants. In a broader view, ck-RNAi seems to represent a piece of a greater co-evolution puzzle between plants and microbes (and, possibly, other organisms as well), where not only RNAs but other molecules are exchanged. As plants evolve, they recruit microbes to assist in the adaptation to available growing environments. Microbes promote plant growth and resilience, and plants, in turn, provide microbes with nutrition (e.g. root exudates) and a desirable habitat (e.g. the rhizosphere or within plant tissues), eventually resulting in the diverse and metabolically rich microbial community that exists in the rhizosphere of terrestrial plants (Lyu et al., 2021). Besides sRNAs, other molecules such as peptides or metabolites participate in the molecular exchange that underlies this evolutionary crosstalk (Frantzeskakis et al., 2020). Yet, as sRNAs are less complex molecular moieties, they may evolve faster in situations demanding an immediate response; for example, even the change of a nucleotide could dramatically change their biological function when needed. Whether sRNAs are indeed the molecular ‘lingua franca’, which affects organismal crosstalk, adaptation, and co-evolution remain to be demonstrated by future studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adeyinka OS, Nasir IA, Tabassum B. 2023. Host-induced silencing of the Cp CHI gene resulted in developmental abnormalities and mortality in maize stem borer (Chilo partellus). P Lo S One 18, e 0280963.36745624 10.1371/journal.pone.0280963 PMC 9901779 · doi ↗ · pubmed ↗
- 2Agrawal N, Dasaradhi PVN, Mohmmed A, Malhotra P, Bhatnagar RK, Mukherjee SK. 2003. RNA interference: biology, mechanism, and applications. Microbiology and Molecular Biology Reviews 67, 657–685.14665679 10.1128/MMBR.67.4.657-685.2003 PMC 309050 · doi ↗ · pubmed ↗
- 3Alves CS, Nogueira FTS. 2021. Plant small RNA world growing bigger: t RNA-derived fragments, longstanding players in regulatory processes. Frontiers in Molecular Biosciences 8, 638911.34164429 10.3389/fmolb.2021.638911 PMC 8215267 · doi ↗ · pubmed ↗
- 4Baeg K, Iwakawa H, Tomari Y. 2017. The poly(A) tail blocks RDR 6 from converting self m RN As into substrates for gene silencing. Nature Plants 3, 17036.28319057 10.1038/nplants.2017.36 · doi ↗ · pubmed ↗
- 5Baldrich P, Rutter BD, Karimi HZ, Podicheti R, Meyers BC, Innes RW. 2019. Plant extracellular vesicles contain diverse small RNA species and are enriched in 10- to 17-nucleotide ‘tiny’ RN As. The Plant Cell 31, 315–324.30705133 10.1105/tpc.18.00872 PMC 6447009 · doi ↗ · pubmed ↗
- 6Bartel B . 2005. Micro RN As directing si RNA biogenesis. Nature Structural & Molecular Biology 12, 569–571.10.1038/nsmb 0705-56915999111 · doi ↗ · pubmed ↗
- 7Baulcombe D . 2004. RNA silencing in plants. Nature 431, 356–363.15372043 10.1038/nature 02874 · doi ↗ · pubmed ↗
- 8Baulcombe DC . 2015. VIGS, HIGS and FIGS: small RNA silencing in the interactions of viruses or filamentous organisms with their plant hosts. Current Opinion in Plant Biology 26, 141–146.26247121 10.1016/j.pbi.2015.06.007 · doi ↗ · pubmed ↗