Evaluation of Phenotypic and Genotypic Susceptibility Testing Methods for Newer β-lactam/β-lactamase Inhibitor Combinations in Multidrug Resistant Pseudomonas aeruginosa
Clayton W Hall, Nicholas Waglechner, Erin Choi, Patryk Aftanas, Kevin Katz, Christie Vermeiren, Finlay Maguire, Robert Kozak, Xena X Li

TL;DR
This study compares methods for testing antibiotic susceptibility in drug-resistant Pseudomonas aeruginosa and finds that genotypic predictions are not yet reliable.
Contribution
The study evaluates the accuracy of commercial susceptibility testing systems and genotypic prediction for newer β-lactam/β-lactamase inhibitor combinations against MDR Pseudomonas aeruginosa.
Findings
The Sensititre panel showed high agreement with broth microdilution for newer β-lactam/β-lactamase inhibitors.
Genotypic testing had high error rates because many resistant isolates lacked identifiable genetic resistance markers.
Acquired β-lactamases were rare causes of resistance in MDR Pseudomonas aeruginosa isolates.
Abstract
Ceftazidime-avibactam (CZA), ceftolozane-tazobactam (CT), and imipenem-relebactam (IMR) are newer β-lactam/β-lactamase inhibitor (BL/BLI) combinations used for treatment of multidrug resistant (MDR) Pseudomonas aeruginosa, although resistance has already emerged. Optimal use of these agents relies on timely, accurate susceptibility testing results and understanding of local resistance mechanisms. 183 MDR P. aeruginosa clinical isolates were used to evaluate the performance of commercial Sensititre and Phoenix panels for CZA, CT, and IMR susceptibility testing compared to broth microdilution (BMD). Genomic resistance determinants were also predicted for each isolate with AMRFinderPlus. Categorical agreement (CA) of the Sensititre panel compared to BMD was 95.8%, 90.1%, and 95.8% for CZA, CT, and IMR, respectively. CA of the Phoenix panel was 83.0% for CZA and 85.7% for CT. The Phoenix…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
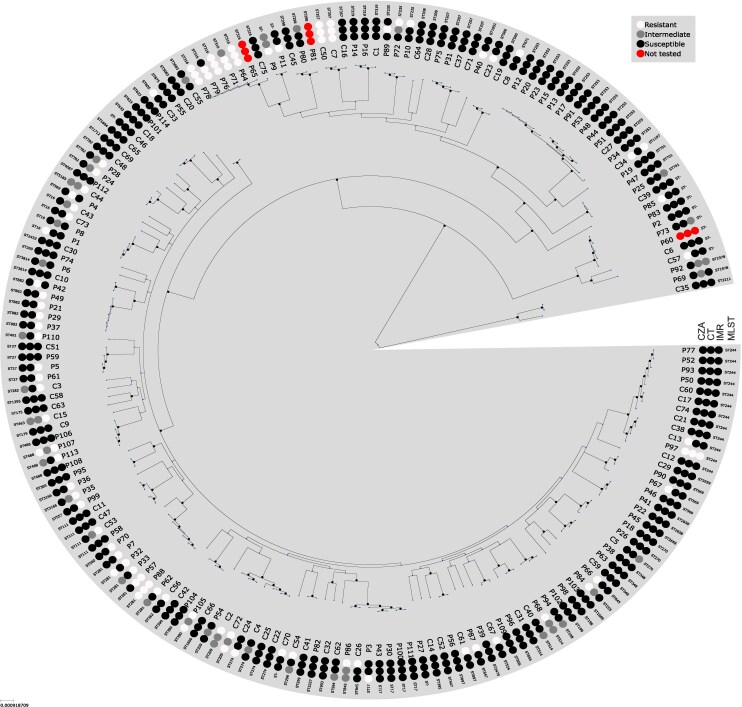 Figure 1
Figure 1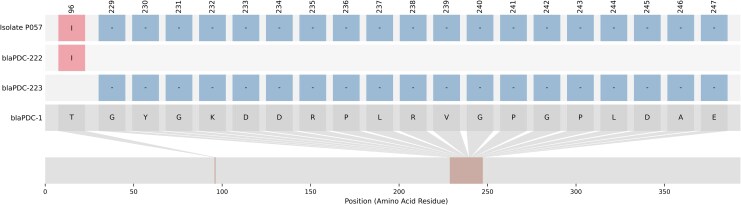 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Antibiotic Use and Resistance
Pseudomonas aeruginosa has an exceptional ability to rapidly develop antimicrobial resistance (AMR) through horizontally acquired resistance genes as well as through mutational adaptation and altered expression of chromosomally encoded resistance determinants [1–3]. Infections due to P. aeruginosa with multidrug resistance (MDR) phenotypes are associated with increased mortality [4, 5], and carbapenem-resistant P. aeruginosa is recognized as a high-priority pathogen by the World Health Organization [6].
Newer antipseudomonal β-lactam/β-lactamase inhibitor (BL/BLI) combinations are recommended for treatment of MDR P. aeruginosa infections with limited first-line treatment options by both the Infectious Diseases Society of America and the European Society of Clinical Microbiology and Infectious Diseases [7, 8]. These BL/BLIs include ceftazidime-avibactam (CZA), ceftolozane-tazobactam (CT), and imipenem-relebactam (IMR).
While the newer BL/BLIs have become invaluable tools to treat MDR P. aeruginosa, resistance to these agents has been increasingly reported worldwide [1, 2]. Oliver et al recently published comprehensive reviews on the known mechanisms of resistance to CZA, CT, and IMR [1, 2]. Acquired, inhibitor-resistant β-lactamases are important mechanisms of BL/BLI resistance in many parts of the world; however, these acquired β-lactamases remain rare in North American P. aeruginosa isolates [9, 10]. In the absence of acquired β-lactamases, BL/BLI resistance appears to be largely driven by adaptive chromosomal mutations [1, 2]. Resistance to CT and CZA has been particularly associated with Ω-loop mutations in the intrinsic Pseudomonas-derived cephalosporinase (PDC) [1, 2]. PDC overexpression can result in low-level CT resistance [11]. PBP3 mutations interfere with β-lactam binding and can result in resistance to CZA, CT, and IMR [12]. CZA and IMR resistance have been linked to overexpression of the MexAB-OprM efflux pump [1, 2].
As the clinical demand for novel antipseudomonal BL/BLIs grows, clinical microbiology laboratories will encounter increasing pressure to provide accurate antimicrobial susceptibility testing (AST) results for these agents in a timely manner [13]. Since CZA, CT, and IMR are tier 3 antipseudomonal antibiotics per current CLSI guidance [14], many laboratories perform AST only on MDR isolates and phenotypic testing may not be available in-house. Results of referred-out AST can take several days, by which time an empiric course may already be completed or the patient may have decompensated on an ineffective regimen. We therefore sought to evaluate the performance of in-house CZA, CT, and IMR phenotypic AST on two commercial platforms compared to referred-out broth microdilution (BMD) results with a large panel of consecutive MDR P. aeruginosa clinical isolates collected between 2023 and 2024 from Toronto, Canada.
In Canada, a national surveillance program for MDR P. aeruginosa is not mandated, and screening for carbapenemase-producing P. aeruginosa is not a routine aspect of infection control practices. Consequently, little is known about the epidemiology and mechanisms of resistance to CZA, CT, and IMR in Canada. We therefore also used whole genome sequencing (WGS) and a publicly available AMR database to identify carbapenemases and other known genotypic mechanisms of BL/BLI resistance. We also evaluated the correlation of genotype with phenotypic susceptibilities.
METHODS
Bacterial Strains
We cultured 6081 P. aeruginosa clinical isolates from blood culture, urine, respiratory, and wound specimens obtained from 3522 patients in 2023 and 2024. Isolates were identified by MALDI-TOF mass spectrometry with the Bruker MALDI Biotyper (MBT Compass Library 2023 revision P). Routine susceptibility testing was performed with the NMIC-451 Gram negative panel on the BD Phoenix M50 or by standard disc diffusion per the laboratory's standard operating procedures. Isolates were considered MDR and included in the study if they were nonsusceptible to ceftazidime, ciprofloxacin, and piperacillin-tazobactam. Overall, a total of 186 MDR P. aeruginosa clinical isolates were isolated from 131 patients during the study period, of which 183 isolates from 130 patients had BMD results.
Reference Broth Microdilution
MDR isolates were referred to the provincial reference laboratory for CZA, CT, and IMR susceptibility testing by reference BMD. BMD was performed once per isolate. MIC calling ranges were ≤4/4 to ≥32/4 µg/mL for CZA and ≤1/4 to ≥16/4 µg/mL for both CT and IMR on the BMD panels.
Sensititre CAN1MSTF Panel Evaluation
Susceptibility testing for CZA, CT, and IMR was performed in-house with a custom Sensititre CAN1MSTF panel (ThermoFisher Scientific) as per the manufacturer's recommendations. MIC calling ranges were ≤4/4 to >16/4 µg/mL for CZA and ≤1/4 to >8/4 µg/mL for both CT and IMR. Panels were read using the Sensititre Vizion Digital MIC Viewing System (ThermoFisher Scientific), and any uncertain MIC reads were resolved with the mirrored Sensititre manual viewbox (ThermoFisher Scientific). Routine QC was performed per the manufacturer's recommendations.
Phoenix NMIC-451 Panel Evaluation
Susceptibility testing for CZA and CT was performed on the BD Phoenix M50 automated susceptibility testing instrument with the Phoenix NMIC-451 panel as per the manufacturer's recommendations. MIC calling ranges were ≤0.25/4 to >8/4 µg/mL for CZA and ≤1/4 to >8/4 µg/mL for CT. IMR was not on the Phoenix NMIC-451 panel and could not be assessed. Routine QC was performed per the manufacturer's recommendations.
Data Analysis and Discrepancy Resolution for Phenotypic Testing
Categorical interpretation of MICs was performed using CLSI M100-Ed35 clinical breakpoints (Supplementary Table 1) [14]. Essential agreement (EA), categorical agreement (CA), and rates of very major errors (VME), major errors (ME), and minor errors (MIN) errors were evaluated for both the Sensititre and the Phoenix panels using BMD as the comparator using standard acceptance criteria per CLSI M52 [15]. Errors were also analyzed using the error-rate-bound method as previously recommended given that >20% of isolates were within ± 1 log_2_ dilution of the BL/BLI breakpoints [16]. For discrepancy resolution, Sensititre or Phoenix testing was repeated in triplicate when one of these systems yielded a VME or a ME. Repeat testing was performed once for isolates with a MIN. Essential errors were not repeated if they did not lead to a categorical error.
Whole Genome Sequencing
Total nucleic acid extraction was performed on EasyMAG (bioMérieux) followed by WGS on the Illumina MiniSeq (2 × 150 bp paired end) with mean coverage depth of 70.1 (interquartile range 51.5–93.7). Sequence quality control and assembly were performed with Bactopia v3.1 using default parameters [17]. Reads were trimmed for quality and adapter removal, filtered using fastp v0.23.4 [18], and then assembled using shovill v.1.0 (https://github.com/tseemann/shovill). To check for contamination and to confirm the taxonomy of each sequenced isolate, assemblies were analyzed using sourmash v4.8.2 [19] against GTDB release 207 and mash v2.3 [20] against NCBI RefSeq release 88. Assemblies were annotated using prokka v1.14.6 [21]. In silico multilocus sequence typing (MLST) was performed using mlst v2.23.0 (www.github.com/tseemann/mlst) with the P. aeruginosa scheme from www.pubmlst.org (downloaded 2025-03-18). A core genome based on loci present in ≥99% of all isolates was estimated using the Bactopia v3.1 pangenome workflow with Panaroo v1.5.0 [22]. ClonalframeML v1.12 [23] was used to mask potentially recombinant sites in the concatenated alignment for core loci, and a maximum likelihood phylogeny was inferred from single nucleotide variants (SNV) in this alignment using IQ-tree v2.2.2.7 with default workflow parameters [24].
Identification of BL/BLI Resistance Determinants
AMR genes and mutations were predicted using AMRFinderPlus v3.12.8 [25]. Isolates were considered genotypically resistant to a given BL/BLI if the isolate carried a resistance gene or mutation previously reported in the literature to confer resistance to that antibiotic. CA between BMD phenotype and genotype was defined as an isolate testing phenotypically susceptible and having no known resistance determinants or an isolate testing phenotypically resistant and having a known resistance determinant. A VME occurred when the genotype predicted false susceptibility (the BMD result was resistant) while a ME occurred when the genotype predicted false resistance (the BMD result was susceptible). Phenotypically intermediate isolates were excluded from this analysis.
Supplementary Material
jiaf585_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Oliver A, Rojo-Molinero E, Arca-Suarez J, et al Pseudomonas aeruginosa antimicrobial susceptibility profiles, resistance mechanisms and international clonal lineages: update from ESGARS-ESCMID/ISARPAE group. Clin Microbiol Infect 2024; 30:469–80.38160753 10.1016/j.cmi.2023.12.026 · doi ↗ · pubmed ↗
- 2Oliver A, Arca-Suárez J, Gomis-Font MA, González-Pinto L, López-Causapé C. Emerging resistance mechanisms to newer β-lactams in Pseudomonas aeruginosa. Clin Microbiol Infect 2025; 31:1790–6.40120758 10.1016/j.cmi.2025.03.013 · doi ↗ · pubmed ↗
- 3Horcajada JP, Montero M, Oliver A, et al Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clin Microbiol Rev 2019; 32:e 00031-19.31462403 10.1128/CMR.00031-19PMC 6730496 · doi ↗ · pubmed ↗
- 4Recio R, Mancheño M, Viedma E, et al Predictors of mortality in bloodstream infections caused by Pseudomonas aeruginosa and impact of antimicrobial resistance and bacterial virulence. Antimicrob Agents Chemother 2020; 64:e 01759-19.31767719 10.1128/AAC.01759-19PMC 6985728 · doi ↗ · pubmed ↗
- 5Morata L, Cobos-Trigueros N, Mart́inez JA, et al Influence of multidrug resistance and appropriate empirical therapy on the 30-day mortality rate of Pseudomonas aeruginosa bacteremia. Antimicrob Agents Chemother 2012; 56:4833–7.22751533 10.1128/AAC.00750-12PMC 3421866 · doi ↗ · pubmed ↗
- 6Jesudason T . WHO publishes updated list of bacterial priority pathogens. Lancet Microbe 2024; 5:100940.39079540 10.1016/j.lanmic.2024.07.003 · doi ↗ · pubmed ↗
- 7Tamma PD, Heil EL, Justo JA, Mathers AJ, Satlin MJ, Bonomo RA. Infectious Diseases Society of America 2024 guidance on the treatment of antimicrobial-resistant gram-negative infections. Clin Infect Dis 2024:ciae 403.39108079 10.1093/cid/ciae 403 · doi ↗ · pubmed ↗
- 8Paul M, Carrara E, Retamar P, et al European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant gram-negative bacilli (endorsed by European society of intensive care medicine). Clin Microbiol Infect 2022; 28:521–47.34923128 10.1016/j.cmi.2021.11.025 · doi ↗ · pubmed ↗