A scaffold attachment factor PHM-2 regulates synaptic transmission through SLO-2 potassium channel in C. elegans
Longgang Niu, Karthika Murugasen, Shannon Hanggodo, Sakia Ferdousy, Lishuang Zhu, Bojun Chen

TL;DR
This study shows that the PHM-2 protein in C. elegans helps regulate nerve-muscle communication by controlling a potassium channel called SLO-2.
Contribution
The study reveals a new role for PHM-2, a SAFB protein, in regulating synaptic transmission through the SLO-2 potassium channel in C. elegans.
Findings
PHM-2 mutants suppress the sluggish movement caused by a hyperactive SLO-2 channel.
PHM-2 reduces SLO-2-mediated neuronal currents and enhances neuromuscular synaptic transmission.
PHM-2 genetically interacts with HRPU-2 to control SLO-2 expression posttranscriptionally.
Abstract
Scaffold attachment factor B (SAFB) proteins are evolutionarily conserved DNA/RNA binding proteins that are involved in multiple processes of gene expression. These proteins are broadly expressed with particular high expression observed in the nervous system. However, their physiological roles in neurons are largely unclear. Here we show that PHM-2, the sole SAFB ortholog in C. elegans, regulates synaptic transmission at the neuromuscular junctions through an effect on SLO-2 potassium channel. We found that phm-2 mutants suppress a sluggish phenotype of worms expressing a hyperactive SLO-2 channel, greatly reduces SLO-2-mediated neuronal whole-cell currents, and enhances neuromuscular synaptic transmission. In addition, we found that PHM-2 genetically interacts with another DNA/RNA binding protein, HRPU-2/hnRNP U, to control SLO-2 expression through a posttranscriptional mechanism.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
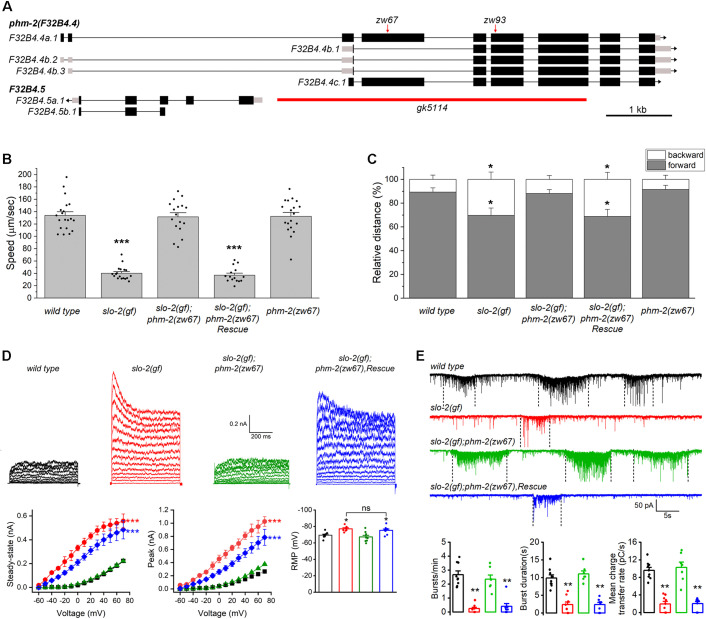 Fig 1
Fig 1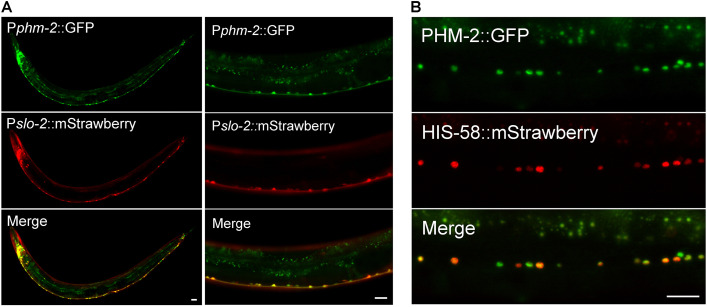 Fig 2
Fig 2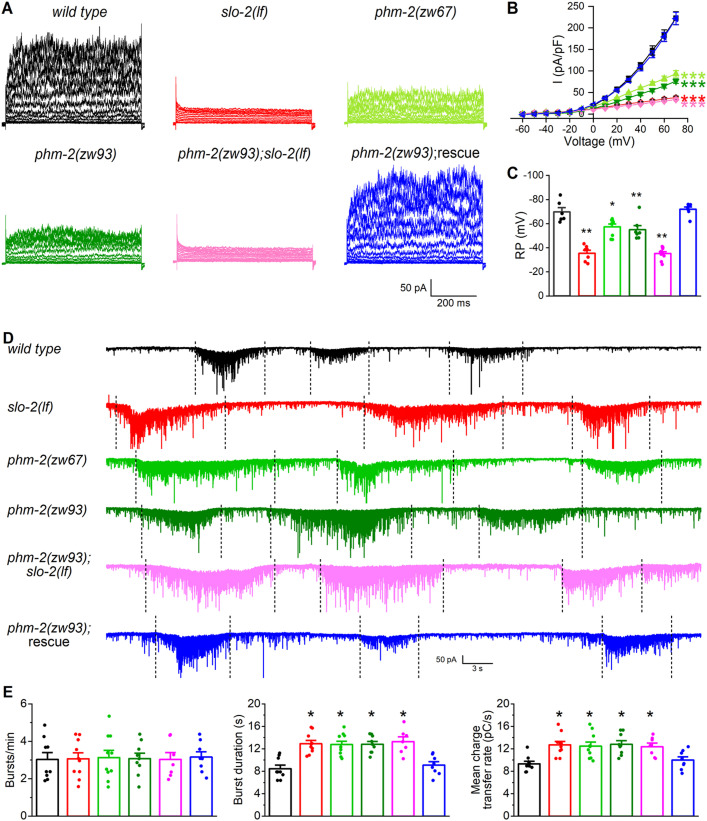 Fig 3
Fig 3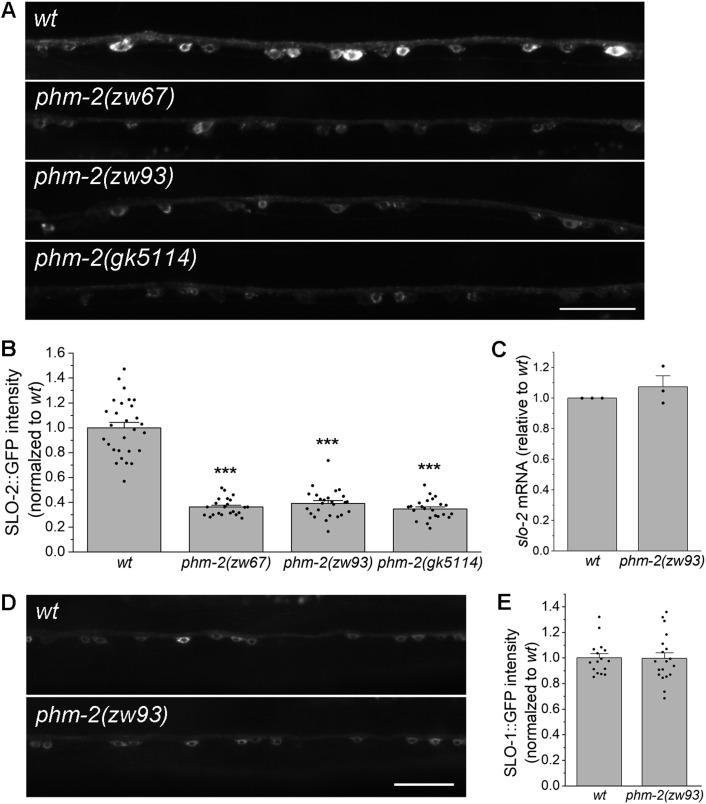 Fig 4
Fig 4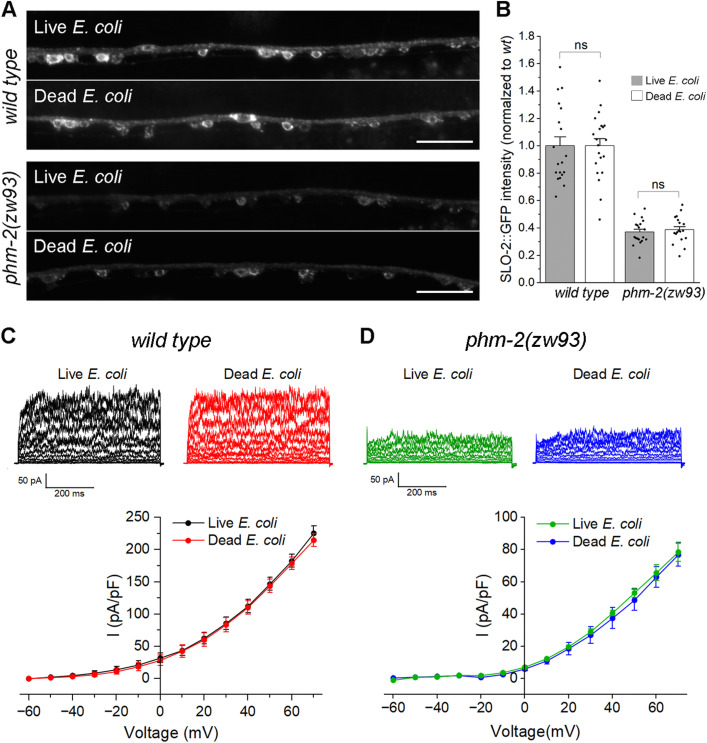 Fig 5
Fig 5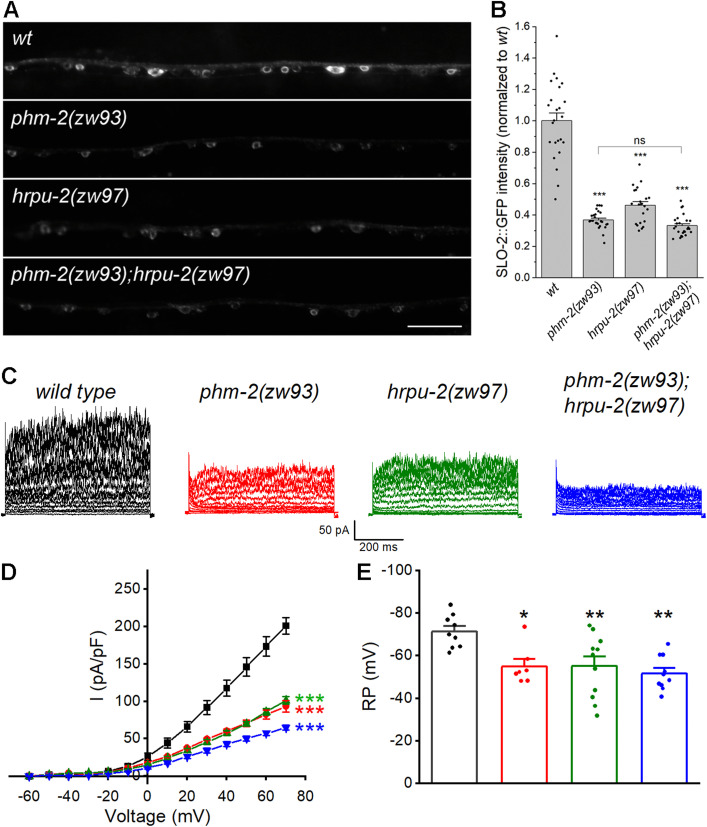 Fig 6
Fig 6- —http://dx.doi.org/10.13039/100000057National Institute of General Medical Sciences
- —http://dx.doi.org/10.13039/100000057National Institute of General Medical Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · RNA Research and Splicing · Genetic Neurodegenerative Diseases
Introduction
RNA-binding proteins (RBPs) constitute one of the largest protein families in eukaryotic cells. These proteins assemble with RNAs to form ribonucleoprotein complexes and play critical roles in RNA biogenesis, stability, function, transport, and cellular localization [1]. In human cells, thousands of RBPs associate with their target RNAs and other proteins to form extensive regulatory networks that regulate cell homeostasis. Analysis of disease association data has identified over a thousand RBPs that are mutated in various human genetic diseases, and Gene Ontology analysis shows that RBPs are predominantly associated with metabolism and nervous system development [2]. In the nervous system, where gene expression is highly dynamic during development and across different regions [3,4], RBPs are essential to neurogenesis, differentiation, and synaptic plasticity, and deficiencies in their expression and/or distribution may cause neurologic disorders such as intellectual disabilities, motor impairments, and neurodegeneration [5,6]. Undoubtedly, mechanistic dissection of the specific roles of RBPs in the brain is crucial for understanding the molecular basis of neurological diseases.
The scaffold attachment factor B (SAFB) proteins are a group of evolutionarily conserved RBPs comprising three members including SAFB1, SAFB2, and SLTM (SAFB-like transcriptional modulator) [7]. Studies have shown that SAFB proteins play important roles in many aspects of cellular processes, such as DNA repair [8,9], cellular stress response [10–12], transcription [13–16], and processing of mRNA and miRNA [17–20]. Evidence also suggests that SAFB proteins may be involved in the progression of various cancers, including prostate cancer [21], pancreatic adenocarcinoma [22], breast cancer [23], and bladder cancer [24]. Like many other RBPs, SAFB proteins are abundantly expressed in the brain [7,25]. In cortical and hippocampal primary neuronal cultures, expression of the SAFB proteins is primarily found in the nuclei, with SAFB2 and SLTM also co-localized in the same dendritic puncta [7]. Adenovirus-mediated expression of SAFB1 in primary hippocampal neurons results in increased dendritic spine size [20]. SAFB1 expression and localization are abnormal in the post-mortem brain tissue of spinocerebellar ataxias and Huntington’s chorea patients [26]. A recent study shows that SAFB interacts with the ribonuclease Drosha to regulate hippocampal stem cell fate [27]. These observations suggest that SAFB proteins are important for neuronal function, but their physiological roles in the nervous system remain largely undefined.
Slo2 channels are large-conductance potassium channels present in mammals and invertebrates [28,29] and are major contributors to delayed outward currents in neurons [30,31]. Humans and mice each have two Slo2 channel, Slick/Slo2.1 and Slack/Slo2.2, which are broadly expressed in the nervous system [32–34]. These channels shape neuronal excitability and act as key suppressors of sensory hypersensitivity, seizures, and excitotoxicity [28,35–37]. Studies have shown that Slack regulates working memory through HCN coupling [38], supports hippocampal plasticity [39], and decreases amygdala excitability to limit anxiety [40]. Slack also links neuronal activity to local translation via two key mRNA translation regulators, FMRP and CYFIP1 [41], and interacts with NaV1.6 to influence excitability of excitatory and inhibitory neurons [42]. In this study, we found that neuronal expression of SLO-2, an ortholog of human Slo2.2/Slack in C. elegans, is regulated by the SAFB ortholog PHM-2. Loss of phm-2 reduces SLO-2-dependent whole-cell currents in motor neurons and enhances synaptic transmission at the neuromuscular junction. We further show that PHM-2 cooperates with another RBP, HRPU-2/hnRNP U, to control SLO-2 expression in neurons. Our findings reveal a molecular mechanism by which a SAFB protein regulates neuronal function and may shed light on the physiological roles of SAFB proteins in the nervous system of other species.
Results
phm-2 mutant suppresses sluggish phenotype of slo-2(gf) worms
In an effort to identify novel regulators of the SLO-2 potassium channel in C. elegans, we performed a forward genetic screen for suppressors of a sluggish phenotype of worms expressing a hyperactive SLO-2 channel. The hyperactive or gain-of-function (gf) SLO-2 channel was engineered by mutating three consecutive amino acid residues, GQT, to AEL in the 6^th^ transmembrane domain of SLO-2 subunit [43]. Worms expressing the SLO-2(gf) channel showed greatly reduced overall locomotion speed and a reduced forward/backward ratio (Fig 1B, C). This phenotype was almost completely suppressed in one of the isolated mutants, zw67. Through an SNP-based mapping approach [44], we mapped the mutation to a small region on the right arm of chromosome I. Analysis of the whole-genome sequencing data identified a nonsense mutation in the gene phm-2 (F32B4.4, www.wormbase.org) within that region (Fig 1A). Expressing the wild type phm-2 cDNA (F32B4.4a.1) in slo-2(gf);phm-2(zw67) double mutant fully reinstated the sluggish locomotion phenotype (Fig 1B, C), which confirms that the suppression of slo-2(gf) phenotype was indeed attributed to the mutation of *phm-*2. PHM-2 is an ortholog of human scaffold attachment factor B (SAFB) proteins [45]. Like other SAFB proteins, PHM-2 has putative DNA-binding and RNA-binding domains, and arginine/glycine motifs. The mutation in zw67 results in a premature stop at arginine 240 of PHM-2. We also analyzed the locomotion behaviors of the phm-2(zw67) mutant and found that phm-2(zw67) mutant has similar speed and forward/backward ratio as those of wild type worms (Fig 1B, C), suggesting that the suppression of slo-2(gf) phenotype by phm-2(zw67) mutant did not result from an additive effect. Thus, phm-2 likely acts in the same genetic pathway to counteract slo-2(gf).
phm-2 mutants suppress the lethargy of slo-2(gf).(A) Genomic organization of phm-2 and the locations of mutations in different phm-2 alleles used in this study. Black boxes and lines represent exons and introns, respectively, and gray boxes represent untranslated regions. zw67 and zw93 carry premature stop codons, and gk5114 contains a large deletion indicated by the red bar. The gene F32B4.5, which is nested within the largest intron of phm-2, is also indicated. (B) Comparison of average locomotion speed among various worm strains. (C) Comparison of the percentage of forward and backward locomotion among various worm strains. For both B and C, the sample sizes were 20 wild type, 18 slo-2(gf), 17 phm-2(zw67);slo-2(gf), 15 rescue, and 19 phm-2(zw67). The asterisk () indicates a statistically significant difference compared with wild type (* p < 0.05, *** p < 0.001, one-way ANOVA with Tukey’s post hoc tests). (D) phm-2(zw67) reverses the effects of slo-2(gf) on whole-cell currents and the resting membrane potential in VA5 motor neuron, including larger amplitudes of peak and steady-state currents, and an apparent voltage-dependent inactivation, and these effects are rescued by expression of the wild-type phm-2 in neurons. The sample sizes were 11 wild type (wt), 9 slo-2(gf), 9 phm-2(zw67);slo-2(gf), and 9 rescue. The asterisk () indicates a significant difference compared with wild type (** p < 0.001, two-way ANOVA with Tukey’s post hoc tests). (E) phm-2(zw67) suppresses inhibitory effects of slo-2(gf) on the frequency, duration, and mean charge transfer of postsynaptic current (PSC) bursts recorded from body-wall muscle cells, and these effects are rescued by expression of the wild-type phm-2 in neurons. Shown are representative traces of spontaneous PSCs and the comparison of PSC burst properties. The vertically dotted lines mark PSC bursts, which are defined as an apparent increase in PSC frequency accompanied by a baseline shift lasting > 3 sec. The sample sizes were 9 wild type, 9 slo-2(gf), 7 phm-2(zw65);slo-2(gf), and 8 rescue. The asterisk () indicates a statistically significant difference compared with wild type (** p < 0.01, one-way ANOVA with Tukey’s post hoc tests).
To determine how phm-2 mutant might alleviate the locomotion defect of slo-2(gf) worms, we recorded voltage-activated whole-cell currents from a representative ventral cord motor neuron (VA5) of different strains including wild type, slo-2(gf), slo-2(gf);phm-2(zw67), and slo-2(gf);phm-2(zw67) with phm-2 rescued in neurons. Similar to what we reported previously [43,46], VA5 whole-cell currents of slo-2(gf) worms were much bigger than those of wild type worms, with increases in both peak and sustained currents, and an apparent voltage-dependent inactivation that was not observed in wild type (Fig 1D). These characteristics of VA5 whole-cell currents of slo-2(gf) worms were completely abolished in slo-2(gf);phm-2(zw67) worms but could be restored in the double mutant by expressing wild-type phm-2 in neurons (Fig 1D), suggesting that phm-2 mutant has a direct effect to mitigate SLO-2(gf) function.
At C. elegans neuromuscular junctions, ventral cord motor neurons control body-wall muscle cells by producing postsynaptic current (PSC) bursts [47], and SLO-2 plays an important role in this function [31]. To obtain further evidence that the suppression of slo-2(gf) phenotype by phm-2 mutant was due to a dysfunction of SLO-2(gf) channel, we compared PSC bursts recorded from body-wall muscle cells of different strains. We found that PSC bursts in slo-2(gf) strain showed significant decreases in the frequency and strength compared with those in wild type worms (Fig 1E). These synaptic phenotypes of slo-2(gf) were not observed in the slo-2(gf);phm-2(zw67) strain but restored when wild-type phm-2 was expressed in neurons of the double mutant (Fig 1E), suggesting that the activity of SLO-2*(gf)* channel was inhibited by the phm-2 mutation. Together, our analyses of locomotion behaviors, neuronal whole-cell currents, and postsynaptic currents indicate that the action of SLO-2(gf) channel depends on PHM-2.
PHM-2 is highly expressed in neurons and localized in the nucleus
Next, we examined the expression pattern of phm-2 by expressing a promoter::GFP transcriptional reporter though an in vivo homologous recombination approach. Because reporters generated by this strategy incorporate larger genomic regions that are likely to contain all relevant cis-regulatory elements, they often better recapitulate endogenous gene expression [48]. We cloned a 1.1-kb genomic fragment that includes part of the first common exon of all phm-2 splice variants along with the upstream intron, and fused it to GFP. The resultant plasmid was then linearized and co-injected with a fosmid clone that contains part of phm-2 coding region and 30-kb upstream sequence. In vivo homologous recombination between the plasmid and the fosmid is expected to result in a promoter::GFP transcriptional reporter that includes the genomic sequence upstream of phm-2 in the fosmid. In transgenic worms, high GFP expression was observed in many neurons in the head and ventral nerve cord (Fig 2A). This expression pattern appears to be more restricted than that reported in a previous study, which observed ubiquitous phm-2 expression using a 475-bp promoter fragment [45]. The differences in cell-type-specific expression pattern observed with our in vivo recombination approach might be due to the existence of enhancer/repressor elements located in the distal upstream region of the phm-2 locus.
Expression and subcellular localization patterns of PHM-2.(A) phm-2 is co-expressed with slo-2 in many neurons in the nerve ring and along the ventral nerve cord. GFP and mStrawberry are expressed under the control of phm-2 promoter (Pphm-2) and slo-2 promoter (Pslo-2), respectively. (B) PHM-2 is localized in the nucleus of neurons. Shown are fluorescence images of a segment of the ventral nerve cord of a transgenic worm expressing a GFP-tagged PHM-2 and an mStrawberry-tagged HIS-58 (nucleus marker). Both fusion proteins are expressed under the control of the pan-neuronal rab-3 promoter. Note that due to mosaic expression from extrachromosomal arrays, some GFP puncta do not overlap with the red nuclear marker. Scale bar = 20 µm in all panels.
To determine whether the expression pattern of phm-2 correlates with that of slo-2, we crossed the Pphm-2::gfp reporter into a strain carrying Pslo-2::mStrawberry, and examined the resulting GFP and mStrawberry signals. Co-expression of the two fluorescent markers was observed in many neurons in the head as well as in ventral nerve cord motor neurons (Fig 2A). Although some differences were noted between the GFP and mStrawberry patterns, both fluorescent proteins were strongly expressed in the same neuronal populations.
To determine the subcellular localization pattern of PHM-2, we cloned the longest phm-2 splice variant and fused GFP at its C-terminus. The PHM-2::GFP fusion protein was co-expressed with a nuclear marker HIS-58::mStrawberry in neurons. We found that the GFP signal overlapped with the mStrawberry signal (Fig 2B), consistent with the nuclear localization of SAFB proteins.
PHM-2 regulates synaptic transmission through SLO-2
In wild-type worms, SLO-2 channels serve as the major carrier of the delayed outward currents in ventral cord motor neurons [31]. To determine whether PHM-2 is required for native SLO-2 function in neurons, we recorded and compared VA5 whole-cell currents from wild type, slo-2(lf), and phm-2 mutant worms. Compared with wild type, VA5 whole-cell currents were greatly reduced in both slo-2(lf) and phm-2(zw67) mutants (Fig 3A, B). This mutant phenotype was confirmed in a new phm-2 mutant allele, zw93, in which a stop codon was introduced after the amino acid residue alanine 495 of PHM-2 using the CRISPR/Cas9 approach. In slo-2(lf);phm-2(zw93) double mutant, VA5 whole-cell currents were not further reduced compared with those recorded from either slo-2(lf) or phm-2 single mutants (Fig 3A, B), suggesting that PHM-2 contributes to motor neuron whole-cell currents through SLO-2. We also compared the resting membrane potential of VA5 between different strains, and found that it was less hyperpolarized in both slo-2(lf) and phm-2 mutants than in wild type, and similar between slo-2(lf);phm-2(zw93) double mutant and the single mutants (Fig 3C), suggesting that PHM-2 plays a role in setting the resting membrane potential through SLO-2.
*PHM-2 contributes to motor neuron whole-cell currents and regulates postsynaptic current (PSC) bursts through SLO-2.(A) Representative VA5 whole-cell current traces. (B) Current (I) - voltage relationships of the whole-cell currents. Sample sizes were 11 wild type, 9 slo-2(lf), 9 phm-2(zw67), 12 phm-2(zw93), 12 phm-2(zw93);slo-2(lf), and 12 phm-2(zw93) rescue. (C) Resting membrane potentials of VA5. Sample sizes were 6 wild type, 7 slo-2(lf), 9 phm-2(zw67), 7 phm-2(zw93), 10 phm-2(zw93);slo-2(lf), and 7 phm-2(zw93) rescue. (D) Representative traces of spontaneous PSCs with PSC bursts marked by vertically dotted lines. (E) Comparisons of PSC burst properties. Sample sizes were 9 wild type, 10 slo-2(lf), 10 phm-2(zw67), 9 phm-2(zw93), 7 phm-2(zw93);slo-2(lf), and 8 phm-2(zw93) rescue. All values are shown as mean ± SE. The asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, **p < 0.001) compared with wild type based on either two-way (B) or one-way (C and E) ANOVA with Tukey’s post hoc tests.
SLO-2 serves as a negative regulator of neurotransmitter release at the neuromuscular junctions [31], and slo-2(lf) mutants exhibit significantly increased duration and charge transfer rate of PSC bursts. To determine whether PHM-2 is also required for this function of SLO-2, we recorded PSC bursts from body-wall muscle cells of wild type and various mutant strains. Similar to slo-2(lf) mutant, the phm-2 mutants showed increased duration and charge transfer rate of PSC bursts without a change in burst frequency. These synaptic phenotypes of phm-2 mutants were not additive with those of slo-2(lf) and were rescued by expressing wild-type phm-2 cDNA specifically in neurons (Fig 3D, E), indicating that PHM-2 regulates neurotransmitter release through presynaptic SLO-2. Our observation that defects in neuronal properties and synaptic transmission in phm-2 mutants can be fully rescued by expression of wild type phm-2 cDNA also excludes a significant contribution from two genes, F32B4.5 and dys-1, that are closely related to the phm-2 locus. F32B4.5 is nested within the largest intron of phm-2 (Fig 1A), and dys-1 is immediately adjacent to phm-2 (www.genebase.org). Nevertheless, we cannot exclude the possibility that phm-2 mutations subtly affect F32B4.5 or dys-1 expression, potentially influencing SLO-2 expression and/or function in ways that are not detectable with our current approaches.
PHM-2 is required for SLO-2 expression
As an RNA/DNA binding protein, PHM-2 might regulate neuronal function through an effect on SLO-2 expression. To test this possibility, an ideal approach would be to compare native SLO-2 protein levels between wild type and phm-2 mutant worms using an antibody against SLO-2. However, no commercial SLO-2 antibodies were available, and our effort to generate a custom SLO-2 antibody was unsuccessful. Therefore, we examined SLO-2 expression using a SLO-2::GFP fusion protein, in which GFP was fused at the C-terminus of the splicing variant F08B12.3a.1. To maintain identical transgene dosage, we crossed a highly stable transgene (> 99% penetrance) expressing SLO-2::GFP in neurons from an existing wild-type strain into the two phm-2 mutants described above, as well as an additional allele (gk5114), in which the majority of phm-2 is deleted (Fig 1A), and quantified the GFP epifluorescence in a segment of the ventral nerve cord. We found that GFP epifluorescence in neurons was greatly reduced in all three phm-2 mutants compared with wild-type worms (Fig 4A, B), suggesting that PHM-2 is required for proper SLO-2 expression. PHM-2 might regulate SLO-2 expression through transcriptional or post-transcriptional mechanisms. To distinguish these possibilities, we quantified slo-2 mRNA levels in wild type and the phm-2(zw93) mutant using our RNA-Seq data. We found that slo-2 mRNA levels were the same between wild type and phm-2(zw93) mutant (Fig 4C), suggesting that PHM-2 regulates SLO-2 expression post-transcriptionally. To determine whether the decrease in SLO-2::GFP in phm-2 mutants reflects a global reduction of protein expression, we compared SLO-1::GFP expression between wild type and phm-2(zw93) worms. SLO-1 is a voltage- and calcium-activated potassium channel and a paralog of SLO-2 in worms. We found that SLO-1::GFP expression in phm-2(zw93) mutants remained unchanged relative to wild type (Fig 4D, E). Thus, loss of phm-2 does not cause a general reduction in protein expression.
PHM-2 regulates SLO-2 expression post-transcriptionally.(A) Representative images of SLO-2::GFP expression in the ventral nerve cord from wild type (wt) and phm-2 mutants. Scale bar = 20 µm. (B) Comparison of SLO-2::GFP signal between wt and phm-2 mutants. The sample sizes were 28 wt, 22 phm-2(zw67), 25 phm-2(zw93), and 23 phm-2(gk5114). The asterisk () indicates a significant difference compared with wt (*** p < 0.001, one-way ANOVA with Tukey’s post hoc tests). (C) Comparison of slo-2 mRNA level between wt and phm-2(zw93) mutants. Shown are results from RNA-seq experiments of three biological replicates per genotype (GEO accession number GSE315272). slo-2 mRNA level in the mutant was normalized to wt = 1. A one-sample t-test indicated no significant difference from wt (p = 0.4045). (D) Representative images of SLO-1::GFP expression in ventral nerve cord from wt and phm-2(zw93) mutants. Scale bar = 20 µm. E, Comparison of SLO-1::GFP signal between wt and phm-2(zw93) mutants. SLO-1::GFP signal was normalized to wt. A two-sample (unpaired) Student’s t-test indicated no significant difference between the two groups (p = 0.9387). The sample sizes were 16 wt and 19 phm-2(zw93).*
Decreased SLO-2 expression in phm-2 mutants is not caused by feeding worms live E. coli
A previous study showed that phm-2 mutants have an abnormal pharyngeal grinder that allows live bacteria to accumulate in the intestine, which is thought to trigger innate immune responses that result in a scrawny body morphology and delayed reproductive aging. These mutant phenotypes can be abrogated by feeding the mutants UV-killed E. coli [45]. To examine whether the effect of phm-2 mutants on SLO-2 expression might be caused by live bacteria, we analyzed SLO-2::GFP expression in wild-type and phm-2(zw93) worms fed either live or UV-killed E. coli. We found that SLO-2::GFP expression was unchanged between the two diets in both wild type and phm-2(zw93) mutants (Fig 5A). We also recorded VA5 whole-cell currents from these worms and found no significant diet-dependent differences within each genotype (Fig 5B). These observations suggest that the effect of phm-2 mutation on SLO-2 expression was not due to exposure to live E. coli.
Reduction of SLO-2 expression in neurons of phm-2 mutants is not caused by feeding worms live E. coli.(A) Representative images of SLO-2::GFP expression in the ventral nerve cord of wild type and phm-2(zw93) mutants grown on either live or dead E. coli. Scale bar = 20 µm. (B) Comparison of SLO-2::GFP signals in worms grown on live versus dead E. coli. The sample sizes were 19 wild type (live E. coli), 21 wild type (dead E. coli), 19 phm-2(zw93) (live E. coli), and 18 phm-2(zw93) (dead E. coli). Statistical significance between live and dead E. coli groups within each genotype was evaluated using a two-sample (unpaired) Student’s t-test. Bonferroni correction was applied for two comparisons (adjusted α = 0.025). No statistically significant differences were detected (p > 0.05). (C) Comparison of VA5 whole-cell currents in wild-type worms grown on live versus dead E. coli. The sample sizes were 10 grown on live E. coli, 10 grown on dead E. coli. (D) Comparison of VA5 whole-cell currents in phm-2(zw93) mutants grown on live versus dead E. coli. The sample sizes were 7 grown on live E. coli, 10 grown on dead E. coli. In both C and D, no statistically significant differences were detected between the groups (p > 0.05, two-way ANOVA with Tukey’s post hoc tests).
PHM-2 acts with HRPU-2 to control SLO-2 expression
Our previous study showed that SLO-2 expression is regulated by HRPU-2, a homologue of mammalian heterogeneous nuclear ribonucleoprotein U (hnRNP U) [43]. Because HRPU-2 regulates SLO-2 expression through a post-transcriptional mechanism, we asked whether PHM-2 and HRPU-2 might function together to regulate SLO-2. To test this, we generated a phm-2(zw93);hrpu-2(zw97) double mutant and compared SLO-2::GFP expression between the double mutant and each single mutant. The hrpu-2(zw97) mutant allele was created by introducing a stop codon after the amino acid residue glutamate 194 of HRPU-2 using the CRISPR/Cas9 approach. We found that the SLO-2::GFP intensity in neurons was similar among the double and the single mutants (Fig 6A, B). These results suggest that PHM-2 and HRPU-2 likely act in the same pathway to control SLO-2 expression.
PHM-2 acts with HRPU-2 to control SLO-2 expression.(A) Representative images of SLO-2::GFP expression in the ventral nerve cord from wild type (wt) and the single and double mutants. Scale bar = 20 µm. (B) Comparison of SLO-2::GFP signal between wt and the single and double mutants. The sample sizes were 24 wt, 25 phm-2(zw93), 22 hrpu-2(zw97), and 25 phm-2(zw93);hrpu-2(zw97). The asterisk () indicates a significant difference compared with wt (*** p < 0.001, one-way ANOVA with Tukey’s post hoc tests). (C) Representative VA5 whole-cell current traces. (D) Current (I) - voltage relationships of the whole-cell currents. Values are shown as mean ± SE. The asterisks indicate statistically significant differences compared with wt (***p < 0.001, two-way ANOVA with Tukey’s post hoc tests). Sample sizes were 9 wt, 7 phm-2(zw93), 11 hrpu-2(zw97), and 10 phm-2(zw93);hrpu-2(zw97). (E) Resting membrane potentials of VA5. Data are shown as mean ± SE. The asterisks indicate statistically significant differences compared with wt (*p < 0.05, *p < 0.01, one-way ANOVA with Tukey’s post hoc tests). Sample sizes were 9 wt, 7 phm-2(zw93), 11 hrpu-2(zw97), and 10 phm-2(zw93);hrpu-2(zw97).
Both phm-2 and hrpu-2 single mutants partially reduce the SLO-2-mediated neuronal whole-cell currents but fully suppressed slo-2(gf) behavioral phenotypes, which raises the possibility that phm-2 and hrpu-2 are required for the function of other ion channels, particularly the second large-conductance potassium channel SLO-1. To address this, we crossed the slo-1(gf) transgene into the phm-2 and hrpu-2 mutants, respectively, and examined their locomotor behaviors. We found that loss of phm-2 or hrpu-2 did not suppress the overall sluggish locomotion of slo-1(gf) worms (S1 Fig), suggesting that PHM-2 and HRPU-2 do not directly modulate SLO-1 channel function. Interestingly, loss of phm-2 reversed the forward/backward movement ratio in slo-1(gf) worms (S1 Fig), suggesting that PHM-2 may act through other channels or neuronal pathways that influence specific locomotor circuits.
To further examine the relationship between PHM-2 and HRPU-2 with respect to SLO-2 function, we compared VA5 whole-cell currents in the phm-2(zw93);hrpu-2(zw97) double mutant and those in the single mutants. If PHM-2 and HRPU-2 regulate SLO-2 independently, we would expect to see very little currents in the double mutant. Instead, VA5 whole-cell currents in the double mutant were only slightly smaller than those in either single mutant (Fig 6C, D), indicating that the current reductions in the single mutants largely overlap. In addition, VA5 resting membrane potentials were comparable between the single and double mutants (Fig 6E). Taken together, these results suggest that PHM-2 and HRPU-2 contribute to neuronal function by acting together on SLO-2 expression.
Discussion
In this study, we demonstrate that PHM-2, the sole SAFB protein in C. elegans, plays an important role in synaptic transmission through the SLO-2 potassium channel. Our conclusion is supported by multiple lines of evidence at the behavioral, cellular, and molecular levels: first, phm-2 mutants suppress the sluggish phenotype of slo-2(gf) worms; second, PHM-2 contributes to neuronal whole-cell currents and post-synaptic currents through SLO-2; third, PHM-2 controls SLO-2 expression through a post-transcriptional mechanism. In addition, we found that PHM-2 functions with another RBP, HRPU-2/hnRNP U, to regulate SLO-2. These findings define a novel neuronal function of a SAFB protein, and may help our understanding of SAFB proteins and their targets in physiology and disease.
We previously showed that HRPU-2/hnRNP U regulates SLO-2 expression in C. elegans neurons [43]. Our new results suggest that PHM-2 has a similar role and may act together with HRPU-2 to regulate SLO-2. In mammals, SAFB1 and hnRNP U were initially characterized based on their capability of binding to matrix/scaffold attachment region (S/MAR) of DNA elements [49,50]. Subsequent studies suggest that these proteins may interact to regulate specific cellular processes. For example, in a comprehensive analysis of the long non-coding RNA (lncRNA)-bound proteome using ChIRP-MS, both SAFB1 and hnRNP U were identified among proteins that bind Xist [51], an lncRNA required for X-chromosome inactivation in female cells to achieve dosage compensation males [52]. In addition, hnRNP U was efficiently pulled down along with several other hnRNPs as SAFB1-interacting proteins from Hela cell nuclear extracts [12]. In humans, mutations of the Slo2.2/Slack channel are strongly linked with epilepsies and intellectual disability [53–58], and deficiencies of hnRNP U are similarly associated with epileptic encephalopathies and intellectual disability [59–64]. Notably, a clinical study reported that SAFB1 and SAFB2 might be responsible for epilepsy and mental retardation in a patient [65]. The common clinical presentations observed in patients with mutations in these genes, together with our findings in worms, suggest that these molecules may interact to perform critical neuronal functions.
A previous study showed that phm-2 mutants have a pharynx grinder defect, which leads to accumulation of live bacteria in the intestine and activation of innate immune responses, including a bacterial avoidance behavior. As a result, phm-2 mutants exhibit a scrawny body morphology and delayed somatic and reproductive aging [45]. These phenotypes of phm-2 mutants do not appear to be related to either SLO-2 or HRPU-2 because both slo-2(lf) and hrpu-2 mutants are morphologically similar to wild-type worms [43]. In addition, while the morphological phenotypes of phm-2 mutants are abrogated when they are raised on a UV-killed E. coli diet [45], the decreased SLO-2 expression and whole-cell currents in neurons of phm-2 mutants remain unchanged compared with those fed a normal E. coli diet. These observations suggest that PHM-2 has multiple physiological functions, which is consistent with its broad expression pattern. In mammals, SAFB proteins are widely expressed, with high expression observed in the developing and mature brain [25]. Immunoblot data also show high expression of SAFB1 and SAFB2 in the immune system and in hormonally regulated organs such as uterus and ovary [66]. It would be interesting to know whether SAFB proteins may have conserved roles in the immune and reproductive systems.
In the slo-2(gf) strain, we observed large outward K⁺ currents with pronounced voltage-dependent inactivation that is absent in wild type. In contrast, both the increase in current amplitude and the voltage-dependent inactivation were abolished in slo-2(gf);phm-2(zw67) double mutants and were fully restored by neuronal expression of wild-type phm-2. Our results show that SLO-2 expression is reduced by approximately 60% in phm-2 mutants; however, while a decrease in channel abundance would be expected to scale current amplitude, it would not, by itself, be sufficient to abolish the characteristic voltage-dependent inactivation of the channel. Thus, reduced expression of SLO-2(gf) alone is unlikely to account for the altered current kinetics observed in the phm-2 mutant. Although lower channel density could contribute indirectly by disrupting channel clustering, interactions with auxiliary or scaffolding proteins, or localization to specific membrane microdomains, these effects point to broader changes in channel regulation. In this scenario, PHM-2 may regulate the expression or function of additional neuronal factors that modulate SLO-2, such as auxiliary subunits or trafficking regulators, such that the altered kinetics in the phm-2 mutant reflects a combination of reduced SLO-2 abundance and a changed regulatory environment, rather than expression changes alone.
Our results show that, although loss of phm-2 fully suppresses behavioral phenotypes of slo-2(gf) worms, phm-2 single mutants display outward K⁺ currents that are lower than wild type but remain higher than those in slo-2 null mutants. This apparent discrepancy likely reflects the robustness and nonlinearity of locomotor circuits, which can tolerate moderate reductions in K⁺ conductance without producing overt behavioral defects. In the absence of SLO-2, residual outward currents and compensatory activity from other channels may be sufficient to maintain normal circuit output. By contrast, slo-2(gf) may shift neuronal excitability beyond a functional threshold, and loss of phm-2 reduces the currents to a permissive range, thereby suppressing the behavioral phenotypes. These observations suggest that PHM-2 may act as a modulatory factor whose contribution becomes evident primarily when SLO-2 activity is elevated.
Slo2 channels are a family of evolutionarily conserved potassium channels. They are widely expressed in the nervous system and have important roles in shaping neuronal activity. Like many other ion channels, the activity of Slo2 channels is regulated by a variety of signaling molecules, such as G-protein-coupled receptors, protein kinases, cyclic AMP, phosphatidylinositol 4,5-biphosphate (PIP2), nicotinamide adenine dinucleotide NAD + , and others [67]. Interestingly, the physiological functions of Slo2 channels may also be regulated by specific RBPs, either directly or indirectly. In mice, the Fragile X Mental Retardation Protein (FMRP) interacts directly with the cytoplasmic C-terminus of the Slo2.2/Slack subunit and potently activates the channel [68]. In C. elegans neurons, HRPU-2 binds to slo-2 mRNA and regulates its expression post-transcriptionally [43], and PHM-2 plays a similar role to HRPU-2 and may interact with it to regulate SLO-2 expression. In addition, SLO-2 function is indirectly modulated by ADR-1 [46], an RBP that regulates adenosine-to-inosine RNA editing [69,70]. ADR-1 facilitates expression of SCYL-1, an evolutionarily conserved regulator of Slo2 channels, by promoting editing of a single adenosine at the 3’-UTR of scyl-1 transcripts [46]. These findings suggest that neuronal Slo2 channels might be under the control of a complex network of RBPs; however, the physiological significance of such regulation remains to be determined.
Our results show that, similar to HRPU-2, PHM-2 regulates slo-2 expression post-transcriptionally. Several mechanisms could be responsible for this regulation, including alternative splicing, nuclear editing, mRNA trafficking, mRNA stability, and translation. In our experiments comparing SLO-2 expression between phm-2 mutants and wild-type worms, a slo-2 cDNA was used to express the SLO-2::GFP fusion in neurons, and the SLO-2(gf) channel that causes the sluggish phenotype suppressed by phm-2 mutants is also encoded by a slo-2 cDNA with a few modifications. Thus, alternative splicing does not appear to be involved in the regulation of SLO-2 by PHM-2. In addition, no RNA editing events are detected in slo-2 mRNA [46], and slo-2 mRNA levels are similar between wild type and phm-2 mutants. Therefore, it appears that, like HRPU-2 [43], PHM-2 may regulate SLO-2 expression through either slo-2 mRNA transport or translation. Nevertheless, apart from their common role in regulating SLO-2, HRPU-2 and PHM-2 likely have distinct functions since the phenotypes of phm-2 and hrpu-2 mutants are very different [43,45].
In the brain, RBPs are essential for maintaining and controlling neuronal excitability [71]. The interaction between RBPs and their targets may link neuronal activity to gene expression that is critical for synaptic plasticity. Indeed, many studies have implicated RBPs in neuronal activity-dependent regulation of gene expression through various post-transcriptional mechanisms, including alternative splicing [72–74], RNA trafficking [75–77], and translation [78–80]. Given that Slo2 channels play major roles in shaping neuronal firing patterns, their regulation by RBPs may be an important mechanism by which the nervous system maintains its homeostasis in response to changes in neuronal activity.
Materials and methods
C. elegans culture and strains
C. elegans were cultured on nematode growth medium (NGM) plates seeded with a layer of Escherichia coli OP50 at 22^o^C inside an environmental incubator. The strains used in this study are listed in S1 Table.
Mutant screening and mapping
Mutant screen was performed with a C. elegans strain with an integrated transgenic array expressing Pslo-1::slo-2(gf) and Pmyo-2::yfp in the N2 Bristol background. L4-stage hermaphrodite worms were immersed in M9 buffer containing 50 mM ethyl methanesulfonate (EMS) for 4 hours at room temperature, and transferred to NGM plates after washing with M9 buffer 5 time. F1 progeny of EMS-treated worms were placed on NGM plates (~10 worms/plate), and the F2 worms were screened for mutants that moved better than the original slo-2(gf) worms. The isolated mutants were subjected to whole-genome sequencing, and SNP-based genetic mapping was performed to determine the rough chromosome location of the mutations. One of the mutants, zw67, was mapped to F32B4.4 (phm-2) and confirmed by subsequent rescue experiments.
Analysis of locomotion behavior
Locomotion behavior was analyzed using an automated worm tracking and analysis system [81]. Briefly, a single young adult hermaphrodite was placed on an NGM plate without food. After ~30 sec recovery time from the transfer, snapshots of the worm were taken at 15 frames per sec for 30 s using an IMAGINGSOURCE camera (DMK37BUX273) mounted on a stereomicroscope (SMZ800, Nikon, Tokyo, Japan). The worm was constantly kept in the center of the view field with a motorized microscope stage (OptiScanTM ES111, Prior Scientific, Inc., Rockland, MA, USA). Both the camera and the motorized stage were controlled by a custom program running in MATLAB (The MathWorks, Inc., Natick, MA).
Generation of phm-2 and hrpu-2 mutant strains
The CRISPR/Cas9 approach was used to create phm-2 and hrpu-2 mutant strain. To generate phm-2 mutant, a guide RNA sequence 5’- GAGAAGCATGTTGCTGAGG was inserted into pDD162 (Peft-3::Cas9 + Empty sgRNA; Addgene #47549). The resultant plasmid was injected into wild type worms along with a repair primer (5’- AAACTGGCTCGGGAGAAGCATGTTGCTTAATGAAGGCGGCGAGCACAATGAGCACTTCCC) and Pmyo-2::mStrawberry (wp1613) as the transgenic marker. The phm-2 mutant worms were identified by PCR using primers 5’- GGGAGAAGCATGTTGCTTAATGA (forward) and 5’- GGAGATTGGAGGATTAGCGGA (reverse). The mutant worms were confirmed by Sanger sequencing. The hrpu-2 mutant was generated using the same approach. The guide RNA sequence for hrpu-2 was 5’-TGGATCATGATGATGAAGG, and the repair primer sequence was:
5’-TGATGAATTAATGGATCATGATGATGAATAACTGGAGGGCATGATGAACATGAAGAAGAT
Mutant worms were identified by PCR using primers 5’- TTAATGGATCATGATGATGAATAACT (forward) and 5’-TGTGACCGATGTCCAGTAATCC (reverse).
Analysis of expression pattern and subcellular localization
The expression pattern of phm-2 was assessed by an in vivo recombination approach. Specifically, a 1.1 kb fragment immediately upstream of phm-2 initiation site was cloned and fused to GFP using the primers 5’- TTTGGTACCAAGCCAGAAGAAATTCCACACAA (forward) and 5’- ATAACCGGTCCGTATTGGCTCGCGAGT (reverse). The resultant plasmid (Pphm-2::gfp, wp1888) was linearized and co-injected with a linearized (fosmid WRM064cE02), which contains 30 kb of phm-2 upstream sequence and part of its coding region, into the lin-15(n765) strain along with a lin-15 rescue plasmid to serve as a transformation marker. Subcellular localization of PHM-2 was determined by fusing GFP to its carboxyl terminus and expressing the fusion protein under the control of Prab-3 (Prab-3::phm-2::gfp, wp1965). Primers for cloning phm-2 cDNA are 5’- TTTGGTACCATGCCGTTGGAAAGCGGAAAA (forward) and 5’- AATACCGGTCAATAATTTCCGCGATAATTTCCATA (reverse). A plasmid harboring Prab-3::his-58::mStrawberry (wp1749) was used to serve as a nucleus marker. The plasmids were injected into the lin-15(n765) strain along with a lin-15 rescue plasmid to serve as a transformation marker. To determine whether phm-2 is co-expressed with slo-2, the Pphm-2::gfp transgene was crossed into an existing strain expressing Pslo-2::mStrawberry. Images of transgenic worms were taken with a digital CMOS camera (Hamamatsu, C11440-22CU) mounted on a Nikon TE2000-U inverted microscope equipped with EGFP/FITC and mCherry/Texas Red filter sets (49002 and 49008, Chroma Technology Corporation, Rockingham, VT, USA).
RNA-seq and data analysis
Total RNA was extracted from young adult-stage worms using TRIzol Reagent (Invitrogen) and treated with TURBO DNase (Ambion). RNA-seq was performed by Novogene Corp. Sacramento, CA.
Raw reads ware filtered using Trim Galore software (http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/) to remove reads containing adapters or reads of low quality. The filtered reads were mapped to C. elegans genome (ce11) using TopHat2 (Kim et al., 2013). The gene expression level is estimated by counting the reads that map to exons.
Quantification of SLO-2::GFP and SLO-1::GFP fluorescence intensity
cDNA was used for expression of SLO-1 and SLO-2 fusion proteins. For the SLO-2::GFP fusion, GFP was fused to the C-terminus of isoform F08B12.3a.1; for the SLO-1::GFP fusion, GFP was inserted into the linker region of isoform Y51A2D.19a.1 between the amino acids alanine 707 and histidine 708. Young adult worms expressing Prab-3::slo-2::gfp or Pslo-1::slo-1::gfp were immobilized in M9 solution containing 1mM azide. Images of the ventral nerve cords posterior to the vulva were obtained using the Hamamatsu digital CMOS camera with an identical exposure time for each group. The ImageJ software was used to extract straightened ventral cord images and to quantify fluorescence intensity. For each image, GFP intensity was calculated by subtracting the minimum intensity (background fluorescence) from the average intensity.
Electrophysiology
Adult hermaphrodites were used in all electrophysiological experiments. Worms were immobilized and dissected as described previously [47]. Borosilicate glass pipettes were used as electrodes for recording whole-cell currents. Pipette tip resistance for recording muscle cell currents was 3–5 MΩ whereas that for recording motor neuron currents was ~ 20 MΩ. The dissected worm preparation was treated briefly with collagenase and perfused with the extracellular solution for 5–10-fold of bath volume. Classical whole-cell configuration was obtained by applying a negative pressure to the recording pipette. Current- and voltage-clamp experiments were performed with a Multiclamp 700B amplifier (Molecular Devices, Sunnyvale, CA, USA) and the Clampex software (version 11, Molecular Devices). Data were sampled at a rate of 10 kHz after filtering at 2 kHz. Spontaneous membrane potential changes were recorded using the current-clamp technique without current injection. VA5 motor neuron whole-cell currents were recorded by applying a series of voltage steps (−60 to +70 mV at 10-mV intervals, 600 ms pulse duration) from a holding potential of −60 mV. The bath solution contained (in mM) 140 NaCl, 5 KCl, 5 CaCl_2_, 5 MgCl_2_, 11 dextrose and 5 HEPES (pH 7.2). The pipette solution contained (in mM) 120 KCl, 20 KOH, 5 Tris, 0.25 CaCl_2_, 4 MgCl_2_, 36 sucrose, 5 EGTA, and 4 Na_2_ATP (pH 7.2). Spontaneous PSCs were recorded from body-wall muscle cells at a holding potential of -60 mV. A PSC burst is defined as a period of apparently increased PSC frequency accompanied by a sustained baseline shift lasting longer than 3 seconds. The recording solutions were the same as those for neuronal whole-cell currents recording, except that the 113.2 KCl in the pipette solution was substituted by K^+^ gluconate. For all Figs, wild-type example current traces were obtained from independent recordings from different animals corresponding to each experimental group.
Data analyses for electrophysiology
Amplitudes of whole-cell currents in response to voltage steps were determined from the mean current during the last 100 ms of the 600-ms voltage pulses using the Clampfit software. The duration and charge transfer of PSC bursts were quantified with Clampfit software (version 11, Molecular Devices) as previously described [47]. The frequency of PSC bursts was manually counted. Statistical comparisons were performed with Origin Pro 2021 (OriginLab Corporation, Northampton, MA) using either ANOVA or unpaired t-test as specified in Fig legends. p < 0.05 is considered to be statistically significant. The sample size (n) equals the number of cells or membrane patches analyzed. All values are shown as mean ± SE and data graphing was done with Origin Pro 2021.
Supporting information
S1 FigLoss of phm-2 or hrpu-2 do not suppress the sluggish locomotion of slo-1(gf).(A) Comparison of average locomotion speed among various worm strains. (B) Comparison of the percentage of forward and backward locomotion among various worm strains. For both A and B, the sample sizes were 19 wild type, 20 slo-1(gf), 22 slo-1(gf);phm-2(zw93); and 20 slo-1(gf);hrpu-2(zw97). The asterisk (*) indicates a statistically significant difference compared with wild type (** p < 0.01, *** p < 0.001, one-way ANOVA with Tukey’s post hoc tests).(TIF)
S1 TableList of strains.(DOCX)
S1 DataMaster Data Table.Excel file with graph values for Figs 1, 3 – 6, and S1.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Glisovic T, Bachorik JL, Yong J, Dreyfuss G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008;582(14):1977–86. doi: 10.1016/j.febslet.2008.03.004 18342629 PMC 2858862 · doi ↗ · pubmed ↗
- 2Gebauer F, Schwarzl T, Valcárcel J, Hentze MW. RNA-binding proteins in human genetic disease. Nat Rev Genet. 2021;22(3):185–98. doi: 10.1038/s 41576-020-00302-y 33235359 · doi ↗ · pubmed ↗
- 3Kang HJ, Kawasawa YI, Cheng F, Zhu Y, Xu X, Li M, et al. Spatio-temporal transcriptome of the human brain. Nature. 2011;478(7370):483–9. doi: 10.1038/nature 10523 22031440 PMC 3566780 · doi ↗ · pubmed ↗
- 4Colantuoni C, Lipska BK, Ye T, Hyde TM, Tao R, Leek JT, et al. Temporal dynamics and genetic control of transcription in the human prefrontal cortex. Nature. 2011;478(7370):519–23. doi: 10.1038/nature 10524 22031444 PMC 3510670 · doi ↗ · pubmed ↗
- 5Romano M, Buratti E. Targeting RNA binding proteins involved in neurodegeneration. J Biomol Screen. 2013;18(9):967–83. doi: 10.1177/1087057113497256 23954928 · doi ↗ · pubmed ↗
- 6Doxakis E. RNA binding proteins: a common denominator of neuronal function and dysfunction. Neurosci Bull. 2014;30(4):610–26. doi: 10.1007/s 12264-014-1443-7 24962082 PMC 5562623 · doi ↗ · pubmed ↗
- 7Norman M, Rivers C, Lee Y-B, Idris J, Uney J. The increasing diversity of functions attributed to the SAFB family of RNA-/DNA-binding proteins. Biochem J. 2016;473(23):4271–88. doi: 10.1042/BCJ 20160649 27888239 · doi ↗ · pubmed ↗
- 8Xiong F, Wang R, Lee J-H, Li S, Chen S-F, Liao Z, et al. RNA m 6A modification orchestrates a LINE-1-host interaction that facilitates retrotransposition and contributes to long gene vulnerability. Cell Res. 2021;31(8):861–85. doi: 10.1038/s 41422-021-00515-8 34108665 PMC 8324889 · doi ↗ · pubmed ↗