A novel QTL GSE3.1 regulates grain size and weight in rice
Kaiying Wang, Jiahao Dong, Chenjie Wang, Chen Zhou, Zhao Li, Wenjie Jiao, Limin Zhang, Minying Dou, Nannan Han, Baolan Zhang, Ran Xu, Yunhai Li, Zupei Liu, Luojiang Huang

TL;DR
This study identifies a new gene in rice that controls grain size and weight, which could help breed higher-yield rice varieties.
Contribution
A novel QTL, GSE3.1, is identified as a regulator of grain size and weight in rice.
Findings
GSE3.1 encodes a TRM protein that promotes cell division to increase grain size.
The GSE3.1 allele from Daligeng improves grain size, weight, and yield in field trials.
The gene provides a molecular mechanism for grain size and weight control in rice.
Abstract
Grain size is a crucial agronomic trait that directly determines grain yield. To elucidate molecular mechanisms of grain size control will help breeders develop high-yield varieties. In this study, we report a novel quantitative trait locus (QTL) GRAIN SIZE AND WEIGHT ON CHROMOSOME 3 (GSE3.1) for grain size and weight in rice, which encodes a TONNEAU1-Recruiting Motif family (TRM) protein and positively regulates grain size by promoting cell division. In addition, the GSE3.1 allele from Daligeng (DLG) significantly increases grain size and weight, and further improves grain yield in field plot trials. Thus, our findings demonstrate the molecular mechanism of GSE3.1 in grain size and weight control, suggesting this gene could be a promising target for breeding high-yield varieties in rice.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
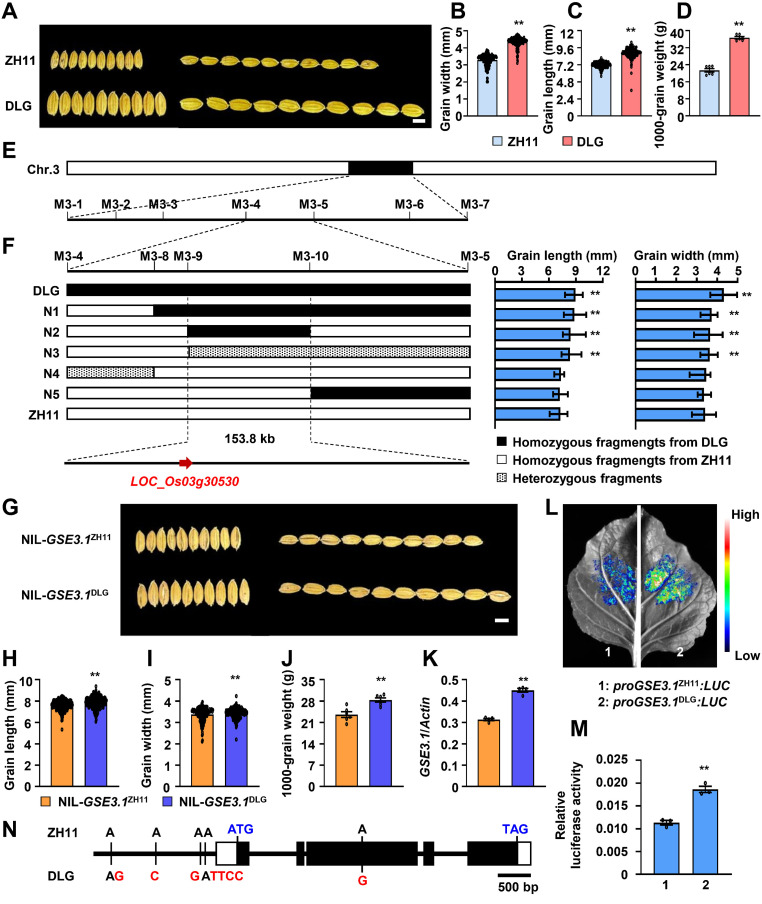 Figure 1
Figure 1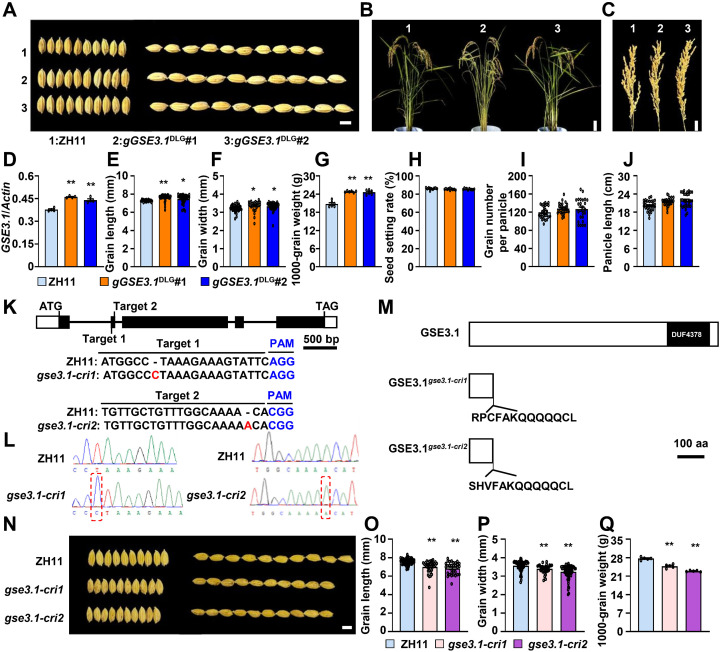 Figure 2
Figure 2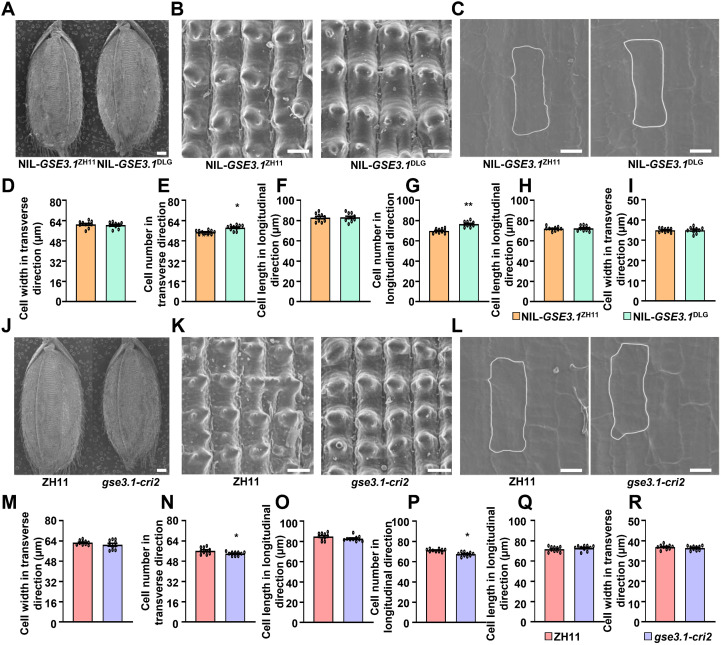 Figure 3
Figure 3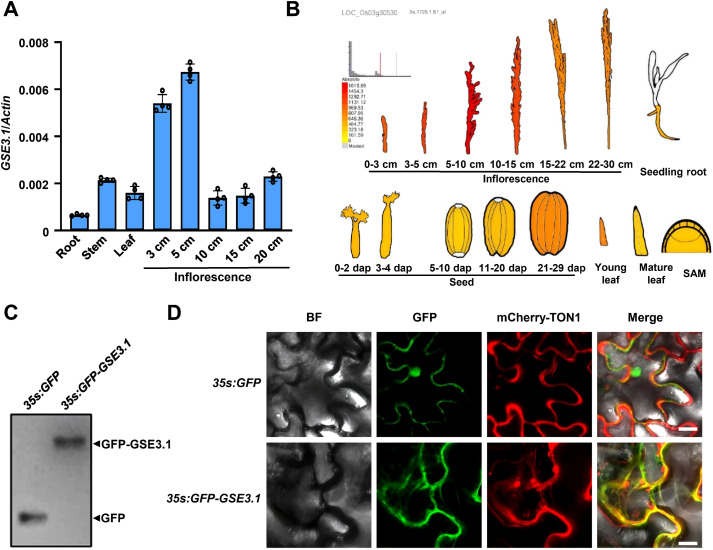 Figure 4
Figure 4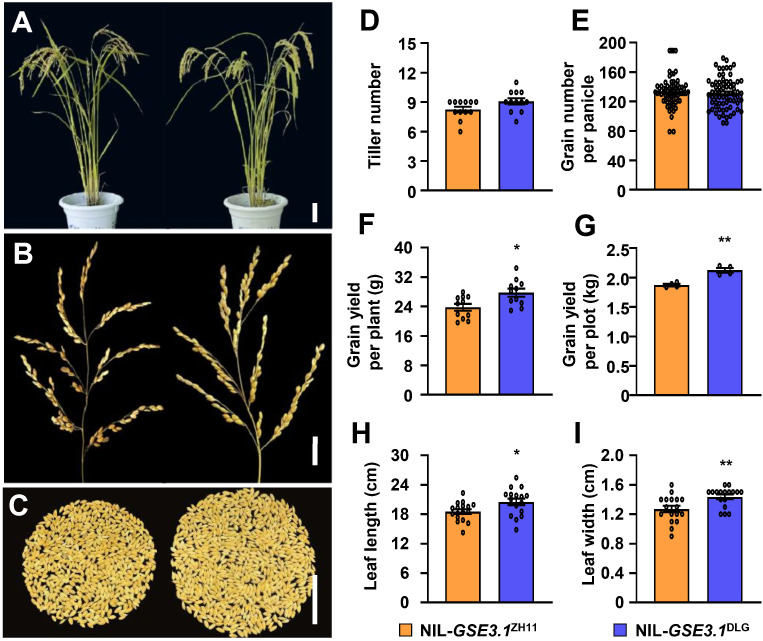 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic Mapping and Diversity in Plants and Animals · Rice Cultivation and Yield Improvement · Plant Molecular Biology Research
Introduction
1
Rice is one of the most important cereal crops worldwide, and feeds more than half of the world’s population (Khush, 2001). Improving rice yield is of vital importance for ensuring global food security. As one of three key determinants of rice yield, grain weight is positively related to grain size (Zuo and Li, 2014). During the past decades, constant efforts have been made by researchers to clone and characterize grain size-related genes, most of which are involved in transcriptional regulation, G protein signaling, plant hormone signaling, the MAPK signaling pathway, and ubiquitin proteasome pathway (Li et al., 2019). Nevertheless, our understanding of the genetic and molecular mechanisms in grain size control is still limited but crucial for improving grain yield.
As plant cytoskeletal proteins, microtubules are indispensable regulators of cell division since diverse microtubule arrangements are associated with distinct stages of the cell cycle (Ehrhardt and Shaw, 2006). During the process of cell division, formation of preprophase band (PPB) of microtubules is the premitotic cytological landmark of the final division plane, and PPB disassembles in late prophase, progressively replaced by a mitotic spindle during metaphase and anaphase (Ambrose and Cyr, 2008). Therefore, precise assembly and disassembly of PPB can ensure the accurate segregation of genetic material and the formation of daughter cells. The PPB assembly is controlled by the TON1/TRM/PP2A (TTP) complex (Schaefer et al., 2017). In Arabidopsis, TRM proteins interact with and target TONNEAU1 (TON1) and type 2A protein phosphatase subunits (PP2A) to cortical microtubules to form the TTP complex, revealing their crucial roles in plant cell division (Drevensek et al., 2012; Schaefer et al., 2017; Spinner et al., 2013). Mutants of TRM genes impair the formation of PPB, which causes inaccuracy in cell division orientation (Schaefer et al., 2017). Up to date, only a few TRM proteins have been reported to participate in plant organ developmental processes (Lee et al., 2006; Wang et al., 2023, 2015, 2015; Wu et al., 2018; Zhou et al., 2015). Arabidopsis TRM1/LNG2 and TRM2/LNG1 regulate organ elongation by positively promoting longitudinal polar cell elongation (Lee et al., 2006). Rice GRAIN LENGTH ON CHROMOSOME 7 (GL7)/GRAIN WIDTH 7 (GW7)/SLENDER GRAIN ON CHROMOSOME 7 (SLG7) encodes a TRM protein, which interacts with OsTON1b and OsTON2, the other two components in TTP complex, and gain-of-function GL7/GW7/SLG7 results in slender grains (Wang et al., 2015, 2015; Wu et al., 2023; Zhou et al., 2015). Therefore, more studies are needed to elucidate molecular mechanisms of TRM proteins in organ size control, which are beneficial for increasing seed/grain weight and yield.
In this study, we performed fine mapping of a novel QTL controlling grain size and weight in rice, and identified the causal gene GSE3.1, which has been previously reported to regulate grain size (Niu et al., 2024; Wu et al., 2023). GSE3.1 encodes a TRM protein, which simultaneously promotes grain length and grain width by positively regulating cell proliferation in both longitudinal and transverse directions in the spikelet hulls. We also demonstrate that the GSE3.1^DLG^ allele, with higher expression of GSE3.1, leads to increased grain size and weight, and significantly increased grain yield in the field plot trials, which suggests its application potential for improving rice grain yield.
Results
2
Map-based cloning of GSE3.1
2.1
To explore QTLs for rice grain size, we selected the japonica variety DLG as a donor and crossed it with the japonica variety Zhonghua11 (ZH11). Compared to ZH11, DLG had significantly increased grain length and grain width, thus resulting in large and heavy grains (Figures 1A–D). We sequenced the whole genomes of ZH11 and DLG, and designed specific markers for mapping (Supplementary Table 3). The F_2_ population was used to map a major QTL GSE3.1 responsible for grain size and weight on chromosome 3. The GSE3.1 locus was further fine-mapped to the 153.8 kb genomic region by using the 2168 BC_3_F_3_ individuals (Figures 1E, F).
*Map-based cloning GSE3.1. (A) mature rice grains of ZH11 and DLG. (B) grain width of ZH11 and DLG (n ≥ 170). (C) grain length of ZH11 and DLG (n ≥ 170). (D) 1,000-grain weight of ZH11 and DLG (n = 6). (E, F) fine mapping of GSE3.1. Left: high-resolution mapping. Right: grain length and grain width analyses of recombinants. (G) mature rice grains of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG. (H) grain length of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG. (N ≥ 210). (I) grain width of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n ≥ 210). (J) 1000-grain weight of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n = 6). (K) relative expression levels of GSE3.1 in young panicles of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n = 4). (L, M) activity analyses of GSE3.1 promoters from ZH11 and DLG by using the dual-luciferase reporter system. The 2.3 kb GSE3.1 promoter sequences from ZH11 and DLG were cloned upstream of the firefly luciferase (LUC) gene to generate the proGSE3.1ZH11:LUC and proGSE3.1DLG:LUC constructs, which were transiently transformed into N. Benthamiana leaves. After 48 h, the image was captured, indicating proGSE3.1DLG:LUC has a higher transcription activity (L). In addition, A renilla luciferase (REN) gene driven by the 35S promoter served as an internal control, and the ratio of LUC to REN luminescence indicates the relative luciferase activity. The diagram shows proGSE3.1DLG: LUC has higher relative luciferase activity than proGSE3.1ZH11:LUC (M). (N) DLG has four variations in the GSE3.1 promoter, and one variation in the coding sequence compared to ZH11. The variations are shown in red and start and stop codons are shown in blue. Values (B–D, H–K, M) are given as mean ± SE. *P < 0.01 compared with ZH11, NIL-GSE3.1ZH11, or proGSE3.1DLG: LUC by Student’s t-test. Bars, 5 mm (A, G).
To identify the candidate gene responsible for the grain size variation between ZH11 and DLG, we carefully compared the genomic sequences of ZH11 and DLG within the 153.8 kb interval and found that three of the total 26 genes contain sequence variations in the promoter, CDS, exon-intron boundary, 5’ UTR and 3’ UTR regions (Supplementary Tables 1, 2). Compared with ZH11, DLG contains four variations in promoter and one variation in CDS in the LOC_Os03g30530 region and has one variation in 3’ UTRs of LOC_Os03g30550 and LOC_Os03g30580 (Figure 1N and Supplementary Table 2). Next, near-isogenic lines (NILs) were obtained in the BC_4_F_3_ population and subjected to further expression analyses. Compared to NIL-GSE3.1^ZH11^, NIL-GSE3.1^DLG^ showed increased grain length, width and weight (Figures 1G–J). Strikingly, expression of LOC_Os03g30530 in NIL-GSE3.1^DLG^ was significantly higher than that in NIL-GSE3.1^ZH11^, while no obvious expression differences of LOC_Os03g30550 and LOC_Os03g30580 were detected (Figure 1K; Supplementary Figure 1). Since there were four variations in the promoter region of LOC_Os03g30530 between ZH11 and DLG, we investigated the activities of the LOC_Os03g30530 promoters from ZH11 and DLG (proGSE3.1^ZH11^ and proGSE3.1^DLG^) in N. benthamiana leaves by using the luciferase reporter system. As shown in Figures 1L, M, the proGSE3.1^DLG:^LUC had stronger luciferase activity than proGSE3.1^ZH11^:LUC, suggesting the LOC_Os03g30530 promoter from DLG resulted in higher expression of GSE3.1. Therefore, LOC_Os03g30530 is the candidate gene for GSE3.1.
GSE3.1 positively regulates grain size and weight
2.2
To further validate whether LOC_Os03g30530 is the GSE3.1 QTL, we carried out a genomic transformation test. A genomic fragment (gGSE3.1^DLG^) from the DLG variety was transformed into ZH11. The gGSE3.1^DLG^ transgenic plants exhibited significantly increased grain length, grain width and 1000-grain weight (Figures 2A, E–G). Next, LOC_Os03g30530 was knocked out by using the CRISPR/Cas9 system and two knockout lines, gse3.1-cri1 and gse3.1-cri2, were obtained. gse3.1-cri1 and gse3.1-cri2 had one base pair insertion in the first and second exon of the GSE3.1 gene (Figures 2K, L), respectively, which resulted in a premature stop codon and truncated GSE3.1 proteins (Figure 2M). Compared to ZH11, both gse3.1-cri1 and gse3.1-cri2 showed decreased grain length, grain width and 1000-grain weight (Figures 2N–Q). Taken together, LOC_Os03g30530 is the causal gene of GSE3.1, and GSE3.1 positively regulates grain size and weight.
*GSE3.1 positively regulates grain size and grain weight. (A) mature rice grains of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (B) plant morphology of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (C) panicle morphology of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (D) relative expression levels of GSE3.1 in young panicles of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (N = 4). (E, F) grain length (E) and grain width (F) of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (N ≥ 40). (G) 1000-grain weight of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (N = 6). (H) seed setting rate of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (N ≥ 10). (I) grain number per panicle of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (N ≥ 25). (J) panicle length of ZH11, gGSE3.1DLG#1 and gGSE3.1DLG#2. (K) schematic of the GSE3.1 gene structure. The start codon (ATG) and stop codon (TAG) are indicated. Black boxes represent exons, white boxes indicate the 5´- and 3´- untranslated regions, and the lines between boxes denote introns. The two target sites (Target 1 and Target 2) are shown, and the PAM sequence are shown in blue. The mutations in gse3.1-cri1 and gse3.1-cri2 are shown in red. (L) sequencing diagrams showing the mutations in gse3.1-cri1 and gse3.1-cri2. The red dotted boxes indicate the insertions. (M) protein structure of GSE3.1. Mutated GSE3.1 proteins of gse3.1-cri1 and gse3.1-cri2 are shown. (N) mature rice grains of ZH11, gse3.1-cri1 and gse3.1-cri2. (O, P) grain length (O) and grain width (P) of ZH11, gse3.1-cri1 and gse3.1-cri2 (n ≥ 30). (Q), 1000-grain weight of ZH11, gse3.1-cri1, and gse3.1-cri2 (n = 6). Values (D–J, O–Q) are given as mean ± SE. **P < 0.01 and P < 0.05 compared with ZH11 by Student’s t-test. Bars, 5 mm (A, N), 15 cm (B), 3 cm (C), 500 bp (K), 100 aa (L).
GSE3.1 promotes cell division to regulate grain size
2.3
Rice grain size determination is crucially influenced by the spikelet hull, in which cell division and cell expansion coordinately regulate grain growth (Li et al., 2019). To understand how GSE3.1 controls grain size, we conducted cell analyses of the NIL-GSE3.1^ZH11^ and NIL-GSE3.1^DLG^ spikelet hulls. As shown in Figures 3A, B, D-G, outer epidermal cell numbers in both longitudinal and transverse directions were significantly increased in the NIL-GSE3.1^DLG^ spikelet hulls, while outer epidermal cell length and cell width in the spikelet hulls of NIL-GSE3.1^DLG^ and NIL-GSE3.1^ZH11^ showed no obvious differences. Additionally, inner epidermal cell length and cell width were almost similar in NIL-GSE3.1^DLG^ and NIL-GSE3.1^ZH11^ spikelet hulls (Figures 3C, H, I). Similarly, cell size of outer and inner epidermal cells in ZH11 and gse3.1-cri2 spikelet hulls exhibited no obvious changes, while cell numbers of outer epidermal cells in both longitudinal and transverse directions of gse3.1-cri2 spikelet hulls were significantly decreased compared to those of ZH11 spikelet hulls (Figures 3J–R). Therefore, these results revealed that GSE3.1 regulates grain size predominantly by promoting cell division.
*GSE3.1 regulates grain size by promoting cell division in spikelet hulls. (A–C) global outer surface (A), outer surface (B) and inner surface (C) of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG spikelet hulls. (D, E) cell width (D) and cell number (E) of outer epidermal cells of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG spikelet hulls in transverse direction (n ≥ 10). (F, G) cell length (F) and cell number (G) of outer epidermal cells of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG spikelet hulls in longitudinal direction (n ≥ 10). (H, I), cell length (H) and cell width (I) of inner epidermal cells of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG spikelet hulls. (J–L) global outer surface (J), outer surface (K) and inner surface (L) of ZH11 and gse3.1-cri2 spikelet hulls. M-N, cell width (M) and cell number (N) of outer epidermal cells of ZH11 and gse3.1-cri2 spikelet hulls in transverse direction (n ≥ 10). (O, P) cell length (O) and cell number (P) of outer epidermal cells of ZH11 and gse3.1-cri2 spikelet hulls in longitudinal direction (n ≥ 10). (Q, R) cell length (Q) and cell width (R) of inner epidermal cells of ZH11 and gse3.1-cri2 spikelet hulls. Values (D-I, M-R) are given as mean ± SE. **P < 0.01 and P < 0.05 indicate significant differences between genotypes by Student’s t-test. Bars, 500 μm (A, J), 50 μm (B, K), 30μm (C, L).
Expression pattern and subcellular localization of GSE3.1
2.4
To elucidate the functions of GSE3.1 in rice developmental processes, quantitative real-time PCR (qRT-PCR) analysis was performed to determine expression pattern of GSE3.1. As shown in Figure 4A, GSE3.1 transcripts were detected in different tissues, including root, stem, leaf and developing inflorescences. Notably, inflorescences at early developing stages (3 cm and 5 cm) had higher expression level of GSE3.1 than those at later developing stages (10 cm, 15 cm and 20 cm), consistent with the GSE3.1/LOC_Os03g30530 expression profiles from public data resources (Figure 4B). Expression pattern of GSE3.1 further supports the role of GSE3.1 in grain size control by promoting cell division.
Expression pattern of GSE3.1 and subcellular localization of GSE3.1. (A) expression levels of GSE3.1 in different rice tissues (n = 4). (B) expression patterns of GSE3.1 in inflorescences, seedling root, developing seed, young leaf, mature leaf and SAM (shoot apical meristem). Data were obtained from botany array resource (http://bar.Utoronto.Ca/efprice/cgi-bin/efpweb.Cgi). (C) detection of the GFP-GSE3.1 fusion protein by Western blot analysis. (D) subcellular localization of GFP-GSE3.1 fusion protein in N. Benthamiana leaf epidermal cells. Images show bright field, GFP fluorescence, mCherry fluorescence and the merged view. The GFP-GSE3.1 fusion protein co-localizes with TON1, a microtubule-binding protein. Bar, 20 μm (D).
GSE3.1 encodes a TRM protein with high sequence identity to Arabidopsis TRM1/LNG2 and TRM2/LNG1 proteins (Supplementary Figure 2). In Arabidopsis, TRM1 interacts with and targets TON1 to cortical microtubules (Drevensek et al., 2012). To investigate the subcellular localization of GSE3.1 protein, we constructed the 35S:GFP-GSE3.1 and 35S:mCherry-TON1 (Arabidopsis TON1 fused with mCherry) constructs, which were transiently coexpressed in N. Benthamiana leaves. The GFP–GSE3.1 fusion protein was detected using a Western blot assay (Figure 4C). Green fluorescence was observed and merged with the microtubule-localized protein TON1, indicating GSE3.1 is localized to microtubules (Figure 4D).
Application potential of GSE3.1 for improving grain yield
2.5
Given that the NIL-GSE3.1^DLG^ plants exhibited larger and heavier grains than NIL-GSE3.1^ZH11^ plants (Figures 1G–J), it is plausible that NIL-GSE3.1^DLG^ plants might increase grain yield. No significant differences between NIL-GSE3.1^ZH11^ and NIL-GSE3.1^DLG^ in terms of overall plant and panicle morphologies, tiller number and grain number per panicle were observed (Figures 5A, B, D, E). Grain yield per plant in NIL-GSE3.1^DLG^ was increased compared with that in NIL-GSE3.1^ZH11^ (Figures 5C, F). To investigate the actual grain yield in NIL-GSE3.1^DLG^ plants, we performed field plot trials. Overall, NIL-GSE3.1^DLG^ plants increased the grain yield by 13.3% in the test plot (Figure 5G). In addition, the leaf length and leaf width of NIL-GSE3.1^DLG^ were increased compared to those of NIL-GSE3.1^ZH11^ (Figures 5H, I). It is possible that the large leaves of NIL-GSE3.1^DLG^ plants might provide more photosynthetic products for grain growth. Therefore, the GSE3.1 allele from DLG increases grain size and weight and has the application potential for improving grain yield.
*NIL-GSE3.1DLG increases grain yield in rice. (A) plant morphology of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG. (B) panicle morphology of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG. (C) grains per plant of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG. (D) tiller number of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n ≥ 12). (E) grain number per panicle of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n ≥ 90). (F) yield per plant of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n ≥ 11). (G) plot yield of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n = 4). (H, I) flag leaf length (H) and width (I) of NIL-GSE3.1ZH11 and NIL-GSE3.1DLG (n ≥ 17). Values (D–I) are given as mean ± SE. **P < 0.01 and P < 0.05 compared with NIL-GSE3.1ZH11 by Student’s t-test. Bars, 15 cm (A), 3 cm (B) and 5 cm (C).
Discussion
3
Grain size is one of important agronomic traits that directly determine rice grain yield. In this study, we report that GSE3.1, a novel QTL allelic to GWL1/IGL1, positively regulates grain size by promoting cell division in spikelet hulls. GSE3.1 encodes a microtubule-localized TRM protein and has a higher expression at early stages of grain development. We also demonstrate that the GSE3.1^DLG^ allele, with higher expression of GSE3.1, leads to increased grain size and weight, and significantly increases grain yield in the field plot trials compared to the GSE3.1^ZH11^ allele. Our findings reveal the molecular mechanism of GSE3.1 underlying grain size regulation and provide a new allele that has application potential for improving grain yield.
GWL1/IGL1 was previously reported to control grain size in rice. GWL1 was firstly identified as a homologous gene of GW7. A knockout mutant line wgl1 in the genetic background of Wuyunjing7 (WYJ7)-DEP1 exhibits short but wide grains, and a WGL1 overexpression line in WYJ7-dep1 shows long and narrow grains, similar to related results of GW7 (Wu et al., 2023). Another study reported that IGL1 functions as a positive regulator for grain length. Knockout mutants and overexpression lines of IGL1 in the Nippobare (NIP) background result in short and long grains, respectively (Niu et al., 2024). Given the two published studies, GWL1/IGL1 is a positive regulator of grain length, consistent with our results of GSE3.1. The latter study didn’t present grain width phenotypes with regard to the IGL1 knockout mutants and overexpression lines, while the former regarded GWL1 as a negative regulator for grain width, which contradicts with our results. This seemingly contradictory phenomenon is not rare in rice because some factors, especially genetic background, may lead to contradictory phenomena in grain size control. Overexpression of GSE9 in ZH11 and NIP results in longer and wider grains, while introgression of GSE9 into Kasalath leads to shorter and wider grains (Chen et al., 2023). The GL3.1/OsPPKL1 knockout mutant in Dongjin background produces longer grains (Zhang et al., 2012), whereas a recent study revealed that its knockout mutant in the ultra-large grain accession (ULG) background results in shorter grains (Li et al., 2024). Therefore, the diverse function of GWL1/GSE3.1 in grain width control might result from different genetic backgrounds. Future molecular and genetic analyses combined with CRISPR/Cas9 knockout system are necessary to solve this problem.
Variations of cis-regulatory elements in promoter regions can influence binding of core transcription factors, which significantly influences gene expression (Wittkopp and Kalay, 2011). By comparing sequence variations of GSE3.1 promoters, GSE3.1 expression and GSE3.1 promoter activity in the NILs, we reveal that the GSE3.1 promoter from DLG causes increased GSE3.1 expression, which is responsible for the large and heavy grains in the NIL-GSE3.1^DLG^. Whether the variations of DLG promoter affects the binding of possible transcription factors needs further investigation. In addition, DLG has an A/G transition in the third exon (A1176G), leading to an Asn/Ser amino acid substitution (Asn338Ser). GSE3.1/WGL1 has been reported to interact with OsTON1b and OsTON2, and future researches are needed to test if this amino acid substitution could impact interactions of GSE3.1 with OsTON1b and OsTON2. Therefore, it would be worth studying the variations in GSE3.1 promoter and possible bound transcription factors, as well as the amino acid substitution (Asn338Ser) and its possible functional differences in the future, which will enrich the molecular mechanisms of GSE3.1 in grain size control.
GSE3.1 protein homologs are found in different plant species (Supplementary Figure 2). Several studies have shown that TRM proteins participate in seed/grain size regulation. Arabidopsis TRM1/LNG2 and TRM2/LNG1, rice GL7/GW7/SLG7 and maize ZmLNG1 positively regulate seed/grain length but negatively regulate seed/grain width. However, our study demonstrates that GSE3.1 acts as a positive regulator in both grain length and grain width. It seems that the former plant TRM proteins, especially rice GW7/GL7/SLG7, contradict GSE3.1 with respect to seed/grain width. Generally, protein homologs, such as GW5 and GW5L, two calmodulin homologs, have similar functions in controlling rice grain size (Duan et al., 2017; Liu et al., 2017; Tian et al., 2019). However, several studies have shown that protein homologs also have diverse roles in regulating grain size. The Kelch-like repeat domain-containing phosphatase OsPPKL2 is a positive regulator of grain length, while OsPPKL1 and OsPPKL3, the closest homologs of OsPPKL2, act as negative regulators of grain length (Zhang et al., 2012). Similarly, the SQUAMOSA Promoter-Binding-Like (SPL) transcription factors, OsSPL13 and OsSPL16, have opposite effects on grain length determination (Si et al., 2016; Wang et al., 2012). In our study, NIL-GSE3.1^DLG^, which has higher GSE3.1 expression, shows increased grain length and grain width compared to NIL-GSE3.1^ZH11^. In addition, grain size analyses in genomic complementation test and GSE3.1 knockout mutants further prove that GSE3.1 is a positive regulator in both grain length and grain width. These results suggest that GSE3.1 has unique advantages over GL7/GW7/SLG7 in improving grain weight and yield in rice.
On cellular level, rice grain size is crucially influenced by cell division and cell expansion in spikelet hulls (Li et al., 2019). TRM proteins in several species have been shown to regulate organ size, especially seed/grain size. In rice, three independent studies with regard to the identical gene GW7/GL7/SLG7 were almost simultaneously reported. However, there is a controversy about cellular mechanisms of GW7/GL7/SLG7 in controlling grain size. The slender grains of gain-of-function GL7/SLG7 result from increased cell length and decreased cell width (Wang et al., 2015; Zhou et al., 2015), demonstrating that GL7/SLG7 regulates grain size by affecting cell expansion rather than cell division. Another study revealed that upregulation of GW7 correlates with slender grains due to increased cell division in the longitudinal direction and decreased cell division in the transverse direction (Wang et al., 2015), suggesting that GW7 regulates grain size by affecting cell division rather than cell expansion. Further cellular analyses of GL7/GW7/SLG7 are needed to resolve this controversy. In dicots, Arabidopsis LNG1/TRM2 positively regulates cell expansion rather than cell division along the proximal-distal axis to control leaf shape (Lee et al., 2006), while tomato SlTRM5 regulates cell division in the proximal-distal and medial-lateral directions to control fruit shape (Wu et al., 2018). Given the above studies, it can be concluded that TRM proteins regulate organ size depending on both cell division and cell expansion. It is well-known that TRM proteins associate with microtubules (Drevensek et al., 2012; Lazzaro et al., 2018), which constitute the structural framework of plant cells and play pivotal regulatory roles in numerous cellular processes, including cell division and cell expansion (Ehrhardt and Shaw, 2006). Thus, it is reasonable that TRM proteins regulate organ size by affecting cell division and cell expansion. In our study, cellular analyses of both NILs and knockout mutant revealed that GSE3.1 positively regulates grain size by promoting cell division in spikelet hulls. In addition, higher expression of GSE3.1 was detected at early stages of grain development, further supporting the role of GSE3.1 in cell division. Therefore, our study is a good example of TRM proteins controlling organ size by cell division, and it will be worth investigating cellular mechanisms of more TRM or cytoskeleton-related proteins in control of organ size.
Grain size usually negatively correlates with grain number, which is also referred to as the trade-off effect (Sadras, 2007). For example, overexpression of the histone acetyltransferase gene GSE3 exhibits increased grain size but decreased grain number, thus resulting in unchanged grain yield (Huang et al., 2024). Strikingly, the NIL-GSE3.1^DLG^ plant produces large and heavy grains without decreasing tiller number and grain number per panicle, and finally increased grain yield by 13.3% in the plot trials, which suggests the application potential of GSE3.1 in breeding high-yield rice varieties.
In summary, our findings report the identification and characterization of a novel QTL GSE3.1 in grain size control. GSE3.1 encodes a TRM protein with subcellular localization to microtubules and positively regulates grain size by promoting cell division. Furthermore, the GSE3.1 allele from DLG significantly increases grain yield in field plot trials. Therefore, our findings reveal the molecular mechanism of grain size control in rice and provide a new allele that could be utilized for breeding high-yield varieties in rice.
Materials and methods
4
Plant materials and growth conditions
4.1
The japonica varieties ZH11 and DLG were used as recipient parent and donor, respectively. ZH11 and DLG were crossed to produce the F_1_ population, which was subsequently backcrossed with ZH11 to generate the BC_3_F_3_ and BC_4_F_3_ populations. The BC_3_F_3_ populations were used for fine mapping. NIL-GSE3.1^ZH11^ and NIL-GSE3.1^DLG^ were obtained in BC_4_F_3_ populations. The gse3.1-cri1 and gse3.1-cri2 mutants were generated using CRISPR/Cas9 technology. Rice plants were cultivated in open fields at Lingshui and Beijing during the natural growing seasons. N. benthamiana plants were grown in a greenhouse under a 16-h light/8-h dark photoperiod.
Morphological analysis
4.2
After plants and grains reached maturity, they were photographed and measured. Grains from the main panicles were scanned using a scanner (MICROTEK ScanMarker i560). Grain length and width were then measured using the Rice Test System (WSEEN). To determine grain weight, one hundred dry grains were weighed, and the average value was calculated from three biological replicates.
Cellular analysis
4.3
SEM and Image J software were used for cellular analyses of mature grains according to a previous study (Li et al., 2025). For outer epidermal cell analysis, intact grains were observed with SEM. We measured grain width and counted total cell number in transverse direction and obtained cell width by dividing the former by the latter. We also measured grain length and counted total cell number in longitudinal direction and obtained cell length by dividing the former by the latter. For inner epidermal cell analysis, the lemmas were separated from grains, then divided and subjected to observation with SEM. We measured lemma width and counted total cell number in transverse direction and obtained cell width by dividing the former by the latter. We also measured lemma length and counted total cell number in longitudinal direction and obtained cell length by dividing the former by the latter.
Vector construction and transformation
4.4
Plasmids were constructed using the Uniclone One−Step Seamless Cloning Kit (Genesand, SC612). The genomic sequence of GSE3.1 was amplified with primers g*GSE3.1*-F and gGSE3.1-R and inserted into the pMDC99 vector to generate the g*GSE3.1* plasmid. The coding sequence (CDS) of GSE3.1 was amplified with primers GFP-GSE3.1-F and GFP-GSE3.1-R and inserted into the pMDC43 vector to generate the 35S:GFP-GSE3.1 plasmid. The GSE3.1 knockout plasmid was constructed using primers gse3.1-cri-F/F0 and gse3.1-cri-R/R0. All constructs were transformed into the japonica variety ZH11 via Agrobacterium tumefaciens strain GV3101 to obtain transgenic plants. Primers used for PCR amplification are listed in Supplementary Table 3.
Analysis of GSE3.1 promoter activity
4.5
The promoter sequences of GSE3.1 from DLG and ZH11 were cloned into the pGreenII0800:LUC vector to generate proGSE3.1^ZH11^:LUC and proGSE3.1^DLG:^LUC, respectively, which were transformed into Agrobacterium GV3101 competent cells. Positive colonies were cultured at 28 °C with shaking at 250 rpm for 24 ;h. Bacterial cells were collected, resuspended in 10 mM MgCl_2_ containing 2 μL mL&^-^¹ of 0.1 M acetosyringone (AS), and incubated for 3 h. Then, 200 ;μL of the suspension was infiltrated into 4-week-old N. benthamiana leaves. After 2 ;d, luciferase activity was quantified to evaluate promoter-driven expression levels.
RNA extraction and quantitative real-time PCR
4.6
Total RNA was extracted from different rice tissues using the RNAprep Pure Kit (TIANGEN, DP439). First strand cDNA was synthesized with the cDNA Synthesis Kit (Vazyme, R211). Quantitative real-time PCR was performed on a LightCycler 480 system (Roche, Switzerland) using SYBR qPCR Mix (Genstar, A301-10). Rice Actin1 was used as the internal control. Primer sequences are provided in Supplementary Table 3.
Subcellular localization of GSE3.1
4.7
The full-length CDS of Arabidopsis TON1 was cloned into the pCAMBIA1300-mCherry vector to generate 35S:mCherry-TON1. The full-length CDS of GSE3.1 was cloned into the pMDC43 vector to generate 35S:GFP-GSE3.1. The construct pairs were transformed into Agrobacterium GV3101 competent cells. Positive single colonies were cultured overnight, and bacterial suspensions were adjusted to OD600 = 0.6. Leaves of 4-week-old N. benthamiana plants were infiltrated, and subcellular localization was observed after 2 d using a laser-scanning confocal microscope (Zeiss LSM710).
Statistical analysis
4.8
Unless otherwise indicated, data were presented as mean ± SE. Statistical analyses were performed using GraphPad Prism 8 (GraphPad Software, Inc., San Diego, CA, USA). Details of statistical tests were provided in the figure legends. P-values were calculated using two−tailed unpaired Student’s t-tests. The sample size was not predetermined by any statistical method.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ambrose J. C. Cyr R. (2008). Mitotic spindle organization by the preprophase band. Mol. Plant 1, 950–960. doi: 10.1093/mp/ssn 054, PMID: 19825595 · doi ↗ · pubmed ↗
- 2Chen R. Xiao N. Lu Y. Tao T. Huang Q. Wang S. . (2023). A de novo evolved gene contributes to rice grain shape difference between indica and japonica. Nat. Commun. 14, 5906. doi: 10.1038/s 41467-023-41669-w, PMID: 37737275 PMC 10516980 · doi ↗ · pubmed ↗
- 3Drevensek S. Goussot M. Duroc Y. Christodoulidou A. Steyaert S. Schaefer E. . (2012). The Arabidopsis TRM 1-TON 1 interaction reveals a recruitment network common to plant cortical microtubule arrays and eukaryotic centrosomes. Plant Cell 24, 178–191. doi: 10.1105/tpc.111.089748, PMID: 22286137 PMC 3289559 · doi ↗ · pubmed ↗
- 4Duan P. Xu J. Zeng D. Zhang B. Geng M. Zhang G. . (2017). Natural variation in the promoter of GSE 5 contributes to grain size diversity in rice. Mol. Plant 10, 685–694. doi: 10.1016/j.molp.2017.03.009, PMID: 28366824 · doi ↗ · pubmed ↗
- 5Ehrhardt D. W. Shaw S. L. (2006). Microtubule dynamics and organization in the plant cortical array. Annu. Rev. Plant Biol. 57, 859–875. doi: 10.1146/annurev.arplant.57.032905.105329, PMID: 16669785 · doi ↗ · pubmed ↗
- 6Huang K. Wang Y. Li Y. Zhang B. Zhang L. Duan P. . (2024). Modulation of histone acetylation enables fully mechanized hybrid rice breeding. Nat. Plants 10, 954–970. doi: 10.1038/s 41477-024-01720-0, PMID: 38831046 · doi ↗ · pubmed ↗
- 7Khush G. S. (2001). Green revolution: the way forward. Nat. Rev. Genet. 2, 815–822. doi: 10.1038/35093585, PMID: 11584298 · doi ↗ · pubmed ↗
- 8Lazzaro M. D. Wu S. Snouffer A. Wang Y. van der Knaap E. (2018). Plant organ shapes are regulated by protein interactions and associations with microtubules. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01766, PMID: 30619384 PMC 6300067 · doi ↗ · pubmed ↗