A review on the genus Kadsura: ethnobotany, pharmacology, and molecular pharmacognosy
Niaojiao Xu, Xianjing Li, Yue Zhang, Lu Qu, Haitao Li, Yunqiang Wang, Meifang Song, Baozhong Duan, Zhonglian Zhang

TL;DR
This review summarizes the traditional and scientific uses of the Kadsura genus, highlighting its medicinal properties and gaps in current research.
Contribution
The paper provides a comprehensive review of Kadsura's ethnobotany, pharmacology, and molecular pharmacognosy, identifying research gaps.
Findings
Kadsura species are traditionally used for circulation and pain relief, with modern validation of anti-rheumatic and liver-protective effects.
Pharmacological mechanisms of Kadsura remain partially understood, with limited safety evaluations and molecular research.
Species-specific therapeutic effects and systematic reviews of Kadsura are currently insufficient.
Abstract
Kadsura Kaempf. ex Juss., belonging to the Schisandraceae family, is a key ethnomedicinal resource in traditional Asian medicine, valued for both dietary and therapeutic roles. Traditionally used to boost blood circulation, relieve pain, and dispel wind-cold-damp pathogens, its metabolites have been validated by modern pharmacology to exhibit potent anti-rheumatoid arthritis, hepatoprotective, antioxidant, and anti-inflammatory properties, supporting clinical applications for rheumatic and hepatic conditions. Literature was retrieved from major databases (Google Scholar, Web of Science, PubMed, ScienceDirect, Baidu Scholar, CNKI, etc.), monographs, and dissertations using “Kadsura” as the core keyword. Species identities were verified via Plants of the World Online (http://www.plantsoftheworldonline.org), and data on ethnobotany, pharmacology, and molecular pharmacognosy were rigorously…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
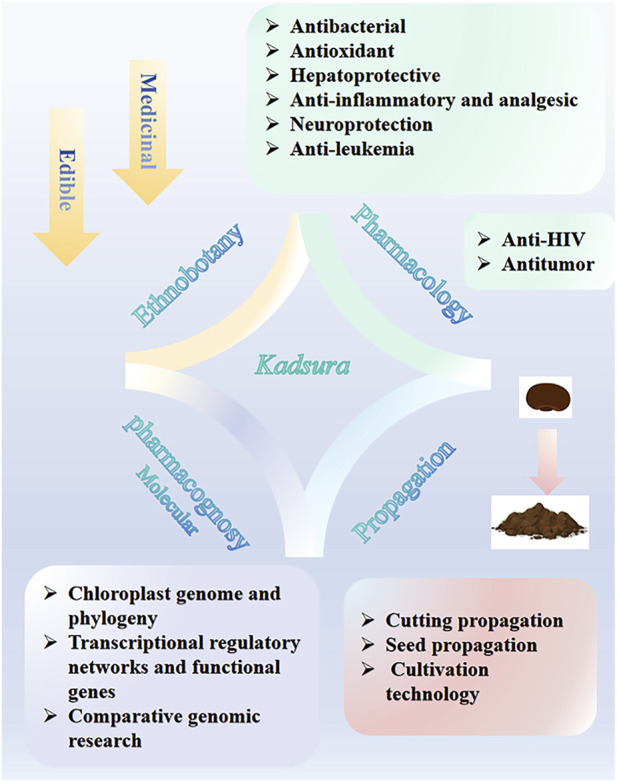 FIGURE 1
FIGURE 1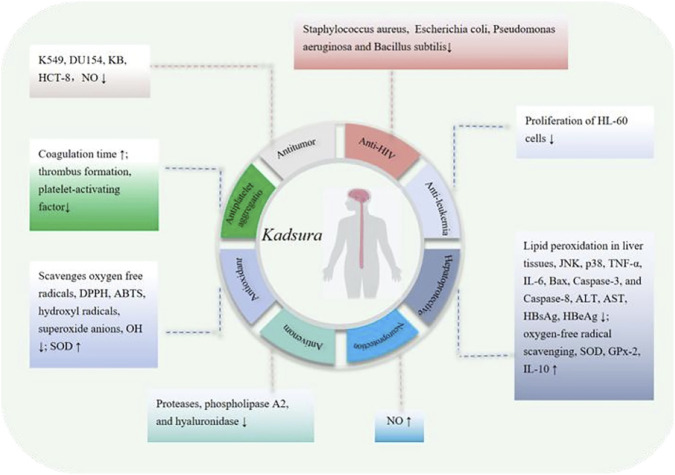 FIGURE 2
FIGURE 2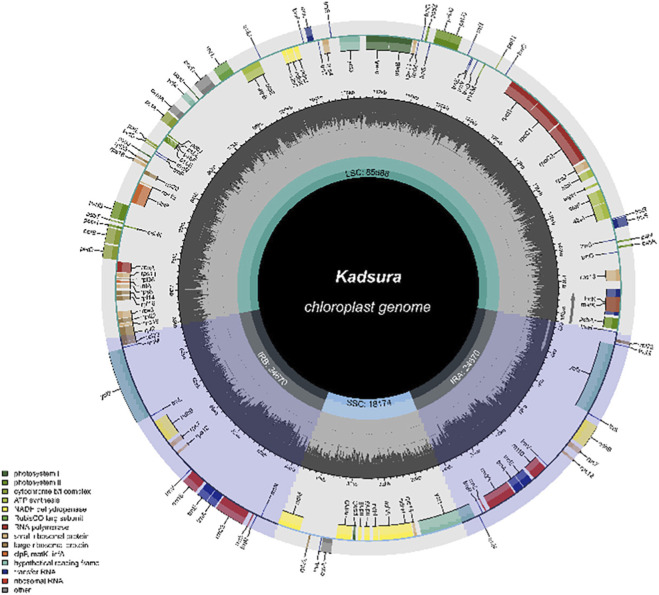 FIGURE 3
FIGURE 3| Species names | Ethnic | Common names | Traditional efficacy and uses | Parts |
|---|---|---|---|---|
|
| Miao, Zhuang, Yao, Tujia, Dong, Dai, Hani, Li, and Da | Len Fantuan, Chou Fantuan, Ru Dishexiang, Shi Bazheng, Hong Zuan | Promotes qi circulation, relieves pain, disperses stasis, reduces swelling, relaxes tendons, and activates collaterals. Treats rheumatoid arthritis, back and leg pain, chronic gastritis, gastric and duodenal ulcers, dysmenorrhea, postpartum abdominal pain, hernia pain, injuries from falls, etc. | Root, bark, stem, leaf |
|
| Dai, Hani, Jinuo, Mulao, Miao, Wa, Zhuang, Tujia | Chui Fengsan, Zhu Naiguo, Len Fantuan, Tong Xuexiang, Di Xuexiang, Feng Qing Jixueteng, Dian Jixueteng | Regulates menstruation, promotes blood circulation, relieves pain, expels wind, and alleviates pain. Treats enteritis, rheumatoid arthritis, gastric and duodenal ulcers, injuries, fractures, external bleeding, lumbar strain, rheumatic bone pain, sciatica, dysmenorrhea, postpartum abdominal pain, colds, bronchitis, neurasthenia, red and white dysentery, etc | Root, stem, fruit |
|
| Dai, Zang, Yi, Miao, Zhuang, Yao, Buyi, Dong, Gelao, Da, Tujia | Hong Muxiang, Zi Jingteng, Zi Jingpi, Xiao Xueteng, Man Shanxiang, Xiang Su | Activates blood, regulates qi, expels wind, promotes joint health, reduces swelling, and relieves pain. Treats stomach pain, abdominal pain, rheumatic pain, dysmenorrhea, irregular menstruation, postpartum abdominal pain, throat swelling, hemorrhoids, undiagnosed swelling, injuries, cuts, cough, poor appetite, night sweats, neurasthenia, etc. | Root, leaf, fruit |
|
| Dai, Miao | Feng Qiangjixueteng, Dian Jixueteng | Clears heat and detoxifies; regulates menstruation; relieves pain; nourishes blood; stops bleeding; and alleviates dysentery. Primarily treats abdominal pain, diarrhea, red and white dysentery, postpartum weakness, etc. | Stem |
|
| Yi | | Promotes blood circulation, removes stasis, regulates qi, and relieves pain. Treats irregular menstruation, rheumatism, fractures, external bleeding, etc. | Whole plant |
|
| Da | Zuan Gufeng, Wen Niuteng, Feng Teng | Treats lung deficiency cough, chronic diarrhea, dysentery, abdominal distention, reversed qi flow, boils, and abscesses. | Root, leaf, seed |
|
| Miao, Yao | Chui Fengsan, Xiao Hongzuan | Treats kidney deficiency and impotence, rheumatic pain, dysmenorrhea, injuries, fractures, enteritis, stomach pain, colds, etc. | Root, stem |
|
| Yao | Tie Zuan | Expels wind and dampness, reduces swelling, and relieves pain. Treats injuries, swelling, rheumatic bone pain, etc. | Stem |
|
| | Xiao Xueteng, Da Hongpao | Dispelling wind and dampness, promoting tendon-muscle relaxation, consolidating astringency, replenishing qi and body fluids, tonifying kidneys, and tranquilizing the mind. Treats chronic cough, asthma, nocturnal emissions, palpitations, insomnia, injuries, rheumatic pain, fractures, irregular menstruation, external bleeding, etc. | Root, stem, leaf, fruit |
| Bioactivity | Metabolites/Extracts | Types | Testing subjects | Doses | Effects/Mechanisms | Control group | Refs |
|---|---|---|---|---|---|---|---|
| Hepatoprotective effects | Ethanol extract |
| HepG2 cells | 100 μM | TC levels ↓ | FFA-induced group/DMEM Medium Supplemented with 10% FBS |
|
| Aqueous extract |
| Clean male and female mice | 2.5, 5.0 g/kg | ALT, AST↓ | Colchicine/Normal Saline |
| |
| Ethanol extract |
| SPF ICR mice | 100, 200, 400 mg/kg | Serum alanine aminotransferase, aspartate aminotransferase levels, apoptosis, oxidative stress, p-Nrf2, Keap1, Cleaved caspase 3 ↓ glutathione, histopathological damage, Bcl-2/BAX ratio, HO-1, and Nrf2 expression↑ | Bicyclol/Normal Saline |
| |
| Aqueous extract |
| KM male and female mice | 20.75, 41.50, 83.00 g/kg | ALT, AST↓ | Administer 200 mg/kg of bifendate/Normal Saline |
| |
| Ethanol extract |
| SPF SD male mice | 0.42, 0.84, 1.68 g/kg | TGF-β1, TNF-α↓ | Administer Fufang Biejia Ruangan pills/Normal Saline |
| |
| Ethanol extract |
| SPF SD male mice | 1.5, 3.0, 6.0 g/kg | FFA, MDA↓ | Administer simvastatin of 4 mg/kg/Normal Saline |
| |
| Ethanol extract |
| Clean male and female mice | 3.0, 5.0 g/kg | ALT, AST, MDA↓ serum, SOD↑ | Administer 25 mg/kg of bifendate/Normal Saline |
| |
| Ethanol extract |
| HSC-T6 cells | 1.5, 3.0, 6.0 g/kg | HSC-T6 cell proliferation↓ miR-193↑ | No HSC-T6 cells were added |
| |
| Ethanol extract |
| ICR male mice | 100, 200, 400 mg/kg | SOD, GPx-2, Bax, Caspase-3, Caspase-8↓ TNF-α, IL-6, MDA, MPO, Bcl-2↑ | Administer a mixture of 25% CCl4 and olive oil/Do not process |
| |
| Ethanol extract |
| SPF SD male mice | 92, 184, 368 mg/kg | ALT, AST↓ | Administer 2.1 mg/kg of bifendate/Normal Saline. |
| |
| Ethanol extract |
| ICR male mice | 200, 400, 800 mg/kg | GPx-2, IL-10, SOD↓ | Administer 200 mg/kg of bifendate/Distilled water |
| |
| Ethanol extract |
| HepG-2 cells | 10 μM | cell viability↑ | AdministerBicyclol//DMEM Medium Supplemented with 10% fetal calf serum |
| |
| Ethanol extract |
| ICR male mice | 100, 200, 400 mg/kg | Inhibited hepatocyte apoptosis; Bcl-2, Bax↑ | Administer 30 mg/kg bifendate/Distilled Water |
| |
| Aqueous extract |
| Healthy mice | 25.65, 51.3, 102.6 g/kg | ALT, AST↓ | Administer 200 mg/kg bifendate/Distilled Water |
| |
| Antioxidant effects | Ethanol extract |
| DPPH,·OH and O2- | 0.025, 0.05, 0.075, 0.1, 0.2, 0.4, 0.6, 0.8 mg/m L | DPPH,·OH and O2-↓ | Vitamin C/Blank solution |
|
| Kadsuralignan F |
| Melan-A cells | 5.94, 11.87, 23.74 μM | The melanin synthesis↓ | DPBS/kojic acid (1%) |
| |
| Ethanol extract |
| phagocytic cells | 2.5, 5, 10, 20, 40 µM | IC50 value of 25.56 µM | Vitamin E/ethanol |
| |
| Aqueous extract |
| DPPH, ABTS | 1.0 mg/mL | DPPH, ABTS↓ | Vitamin C/Distilled Water |
| |
| Ethanol extract |
| DPPH | 125, 250, 500, 1,000 mg/mL | DPPH↓ | Vitamin C/ethanol |
| |
| Aqueous extract |
| DPPH | 0.2, 0.4, 0.6, 0.8, 1.0, 1.2 mg/mL | DPPH↓ | L-ascorbic acid/Distilled Water |
| |
| Anti-inflammatory and analgesic effects | Ethanol extract |
| RAW264.7 cells | Not given | IC50 values of 8.15; IL-6↓ | Methotrexate |
|
| Ethanol extract |
| SPF ICR mice | 200, 400, 800 mg/kg | Inhibiting RA-FLS cell apoptosis and the NF-κB pathway | Not given |
| |
| Aqueous extract |
| Clean SD mice | 0.15, 0.3, 0.6 g/mL | TNF-α, IL-1β↓ | Dexamethasone (0.5 g/lL)/Normal Saline |
| |
| Ethanol extract |
| SPF SD mice | 200, 400, 800 mg/kg | IL-1, IL-2, IL-6, IL-7, TNF-α↓α | Methotrexate (1 mg/kg)/Sodium carboxymethyl cellulose solution (0.3%) |
| |
| Ethanol extract |
| RAW264.7 cells | 1, 3, 10, 30, 50, 100 µg/mL | IC50 value of 34.07 µM | Lipopolysaccharide (100 ng/mL)/DMSO(1%) |
| |
| Ethanol extract |
| RAW264.7 cells | 20, 50, 100 µg/mL | IL-6, IL-12, TNF-α, IL-1β↓ | Not given |
| |
| Ethanol extract |
| SPF ICR mice | 50, 100, 200 mg/kg | TNF-α,IL-1β,IL-6 ↓ | Administer 200 mg/kg of bifendate/Distilled Water |
| |
| Ethanol extract |
| SPF male mice | 1.0, 2.0, 4.0 mg/kg | Promoting apoptosis of mature osteoclasts, inhibiting osteoclast differentiation, and reducing bone resorption; RANKL, RANK, NFATc1, MMP-9, CTSK, TRAP↓ | Sodium carboxymethyl cellulose solution (0.3%) |
| |
| Ethyl acetate extract |
| SPF mice | 4.2, 8.4, 16.8 mg/mL | IL-1β, IL-6, TNF-α↓ | Dexamethasone (0.15 mL/kg)/Normal Saline |
| |
| Antiplatelet aggregation effects | Aqueous extract |
| SPF mice | 7.5, 15 g/kg | Prolonged coagulation time | Warfarin sodium (0.52 mg/kg)/Distilled Water |
|
| Ethanol extract |
| New Zealand White Rabbits | 0.05, 0.5, 1, 5 mg/L | ADP and PAF-induced platelet aggregation↓ | Aspirin (10 mg/L)/Absolute ethyl alcohol |
| |
| Anti-leukemia effects | Ethanol extract |
| HL-60 cells | Not specified | GI50 values of 6.8–42.1 µM | 5-Fluorouracil (50%)/DMSO(0.1%) |
|
| Neuroprotection effects | Methanolic extract |
| BV2 murine microglial cells | 1–50 μg/mL | Regulation of iNOS expression and NF-κB nuclear translocation | NF-κB inhibitor |
|
| Hypolipidaemic effects | Heilaohu seed oil |
| SPF male mice | 150, 300 μL/kg | TC, TG, LDL-C↓ | Feed a high-fat diet/Distilled Water |
|
| Ethanol extract |
| Male C57BL/6N mice | 300 mg/kg | TC, TG, LDL-C↓ | Fed a high-fat diet |
|
| Species names | Number | Size (bp) | LSC length (bp) | IR length (bp) | SSC length (bp) | GC content (%) | Total genes | Protein-coding genes | rRNA genes | tRNA genes | References |
|---|---|---|---|---|---|---|---|---|---|---|---|
|
| HLH-1A | 145617 | 94481 | 16552 | 18032 | 39.7 | 125 | 82 | 8 | 35 |
|
| HLH-2A | 145617 | 94481 | 16552 | 18032 | 39.7 | 125 | 82 | 8 | 35 |
| |
| 145608 | 94457 | 16552 | 18047 | 39.7 | 125 | 82 | 8 | 35 |
| ||
| 145413 | 94511 | 16431 | 18040 | 39.7 | 125 | 82 | 8 | 35 |
| ||
| BN002 | 145875 | 94725 | 16552 | 18046 | 39.7 | 113 | 79 | 4 | 30 |
| |
| 2015091601 | 145412 | 94301 | 16536 | 18039 | 39.7 | 113 | 79 | 4 | 30 |
| |
| 145413 | 94301 | 16536 | 18040 | 39.7 | 113 | 79 | 4 | 30 |
| ||
| 145413 | 94511 | 16431 | 18040 | 39.7 | 126 | 82 | 8 | 35 |
| ||
| 116600 | 145399 | 94287 | 16535 | 18039 | 39.7 | 124 | 82 | 8 | 34 |
| |
| 145608 | 94457 | 16552 | 18047 | 38.6 | 126 | 83 | 7 | 35 |
| ||
| 145399 | 94287 | 16535 | 18039 | 39.7 | 124 | 82 | 8 | 34 |
| ||
|
| 2015090802 | 153029 | 85515 | 24670 | 18174 | 39.6 | 113 | 79 | 4 | 30 |
|
| 153106 | 85593 | 24670 | 18173 | 39.6 | 129 | 84 | 8 | 37 |
| ||
|
| MN 823697/NC_0 57265 | 145903 | 94757 | 16552 | 18042 | 39.7 | 125 | 82 | 8 | 35 |
|
|
| 153201 | 85774 | 24656 | 18115 | 39.6 | 129 | 84 | 8 | 37 |
| |
| 153106 | 85593 | 24670 | 18173 | 39.6 | 129 | 84 | 8 | 37 |
| ||
| 2015092104 | 153127 | 85534 | 24670 | 18253 | 39.6 | 113 | 79 | 4 | 30 |
| |
| 2015082902 | 153096 | 85577 | 24670 | 18179 | 39.6 | 113 | 79 | 4 | 30 |
| |
| 2015090204B | 153362 | 85916 | 24642 | 18162 | 39.6 | 113 | 79 | 4 | 30 |
| |
| BN005 | 153108 | 85829 | 24667 | 17945 | 39.6 | 113 | 79 | 4 | 30 |
| |
| 153289 | 85774 | 24657 | 18201 | 39.6 | 129 | 84 | 8 | 37 |
| ||
|
| 2015121202 | 152810 | 85285 | 24663 | 18199 | 39.6 | 113 | 79 | 4 | 30 |
|
| 153201 | 85774 | 24673 | 18077 | 39.6 | 129 | 85 | 8 | 37 |
| ||
|
| HN2015110801 | 152753 | 85243 | 24670 | 18170 | 39.6 | 113 | 79 | 4 | 30 |
|
|
| TBY2015120104 | 153027 | 85514 | 24670 | 18173 | 39.7 | 113 | 79 | 4 | 30 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-derived Lignans Synthesis and Bioactivity · Bioactive natural compounds · Medicinal Plants and Bioactive Compounds
Introduction
1
The genus Kadsura, belonging to the Schisandraceae family, comprises 29 species predominantly distributed across eastern and southeastern Asia (Guo and Tang, 2021), including China, South Korea, Japan, the Philippines, and Thailand (Sritalahareuthai et al., 2020a; Tai et al., 2022). As a key member of the Schisandraceae family, this plant genus is widely distributed and possesses a long history of medicinal application. Throughout the history of traditional medicine, plants of the genus Kadsura have attracted considerable attention for their remarkable therapeutic properties. Modern pharmacological studies have demonstrated that these plants exhibit strong anti-HIV, antitumor, antioxidant, anti-platelet aggregation, and neuroprotective effects (Liu et al., 2014). Meanwhile, worldwide, the fruits of many Kadsura species also have excellent edible value.
Plants of the Kadsura genus are widely used as traditional folk medicinal materials with various traditional medicinal activities and have great development potential. However, many species of this genus have not yet been studied. Summarizing their medicinal experience is of great significance for the discovery of new chemical substances and the development of new drugs derived from plants of the Kadsura genus. Therefore, it is necessary to summarize current research progress on time and, in combination with traditional medication experience, promptly identify weak research directions and strengthen targeted research in this field, providing a basis for the in-depth development of the clinical application value of Kadsura plants. Furthermore, with increasing global demand for Kadsura plants and rising awareness of ecological protection, research into their introduction, cultivation, and propagation has become crucial. In addition, Molecular pharmacognosy is an interdisciplinary subject derived from molecular biology, specializing in the identification, quality evaluation, and characterization of the molecular constituents of Chinese medicinal materials. In-depth research on the molecular pharmacognosy of the Kadsura plants will provide more scientific and effective technical support for the further development of their medicinal value, and effectively promote the high-quality and sustainable development of the Kadsura industry.
Therefore, this article reviews the progress in its edible characteristics, ethnobotany, and pharmacological research (Figure 1), and proposes further research strategies to provide theoretical support for its modern clinical application in traditional medicine and to lay a foundation for its development in medicine and food. In addition, this article reviews the research progress on Kadsura plants in reproduction and molecular pharmacognosy, providing a basis for analyzing the biosynthetic pathways of their active metabolites and for studying sustainable utilization.
Current research progress of genus Kadsura.
Ethnobotany
2
Edible properties
2.1
The fruits of multiple species of the genus Kadsura can be eaten. In Thailand, the fruits of K. coccinea (Lem.) A.C.Sm. and Kadsura heteroclita (Roxb.) Residents widely consume Craib. Researchers conducted a systematic assessment of the nutritional value of different parts of these fruits (including exocarp, mesocarp, seeds, and pits) (Sritalahareuthai et al., 2020a), verifying their safety for consumption and their nutritional value. In China, the fruits of plants such as K. coccinea, Kadsura longipedunculata Finet and Gagnep., and Kadsura oblongifolia Merr. are also widely consumed as fruits (Idrees et al., 2022; Long et al., 2022; Dong et al., 2023). Among them, the research on K. coccinea is the most extensive. Its fruit has a variety of skin colors (including green, pink, bright red, purplish red, red-black, etc.), is large, non-toxic, and has a sweet-and-sour, juicy taste. Mature K. coccinea seeds are rich in fatty acids, amino acids, vitamins, and minerals, and have healthcare functions such as antioxidation, anti-fatigue, lowering blood lipids and blood pressure, etc., which could meet the material basis conditions for modern human health preservation and balanced nutrition (Li et al., 2025; Liu, 2024; Wang, 2022; Zhang et al., 2022; Zhu et al., 2022). Excitingly, product development for K. coccinea has expanded into the cosmetics industry. Its fruit extract has been included in the “Directory of Names of Used Cosmetic Raw Materials” by the national food and drug regulatory authorities. Currently, five cosmetic products have been developed from the K. coccinea metabolite. Meanwhile, relevant research indicates that its leaf extract also has potential for development as a cosmetic raw material (Ying et al., 2024). Due to the continuous increase in market demand and the growing scarcity of wild resources, southern provinces such as Hunan, Guizhou, and Yunnan in China have already achieved considerable areas of artificial cultivation.
In conclusion, the fruits of many Kadsura species are consumed by people and have definite medicinal value, such as those of K. coccinea, K. heteroclita, and K. oblongifolia. Among them, K. coccinea is currently the most widely used, extensively studied, and largest cultivated species.
Medicinal properties
2.2
The majority of plants in the Kadsura genus have shown considerable therapeutic efficacy, and their traditional medicinal use dates back many years (Table 1). In traditional medicine, the roots, stems, and leaves of these plants are frequently used as botanical drugs, mainly to promote blood circulation, reduce pain, and dispel wind-dampness (Song, 1988; Xu et al., 2008; Zhang et al., 2021; Xiao et al., 2010). Additionally, the fruits of the Kadsura species are widely used for health preservation and possess noteworthy therapeutic properties. Additionally, the fruits of the Kadsura species exhibit notable medicinal properties (Guo, 2016; Huang et al., 2021) and are widely utilized in health preservation. For example, K. coccinea is valued not only for its medicinal properties but also widely consumed as a fruit in Thailand (Sritalahareuthai et al. (2020a)). Their unique pharmacological properties and nutritional profiles have garnered increasing scientific attention and recognition.
Combining traditional folk medicinal knowledge, people have already conducted corresponding pharmacological activity studies on some species. K. heteroclita, found in regions such as China, Sikkim, Bangladesh, Vietnam, Laos, Myanmar, Thailand, India, and Sri Lanka (Editorial Committee Of Flora Reipublicae Popularis Sinicae, 2004), demonstrates significant pharmacological benefits, including anti-inflammatory, anticancer, and hepatoprotective effects (Wang et al., 2021; Yu et al., 2019a; Yu et al., 2019b; Deng and Zhang, 2016). Sritalahareuthai et al. (2020b) reported that the phenolic profiles of this species exhibit potent antioxidant and inhibitory activities. The vine stems of K. longipedunculata, traditionally known as “Hongmuxiang” in China, are widely used to treat inflammatory and infectious conditions. In Germany, they are appreciated for their diverse pharmacological activities, including antimicrobial, trypanocidal, anti-inflammatory, and cytotoxic effects (Mulyaningsih et al., 2010). Furthermore, revealed that its lignans play a significant role in modulating the GABA A receptor. Similarly, the lignans in K. interior have demonstrated inhibition of ADP-induced platelet aggregation (Liu et al., 2018b).
The species involved in this article exhibit both similarities and distinct differences in their traditional applications. For example, most of the Kadsura species listed in Table 1 have the effects of promoting qi circulation and relieving pain, activating blood circulation and removing blood stasis, dispelling wind and dampness. They are mainly used to treat symptoms such as injuries from falls and blows, rheumatic pain, and menstrual disorders. In addition, some species have unique medicinal properties. For instance, K. interior possesses the unique effects of clearing heat and detoxifying, as well as replenishing and stopping bleeding among the species of the Kadsura genus. K. japonica has the effect of treating lung deficiency cough, K. oblongifolia has the effect of treating kidney deficiency impotence, and K. ananosma has the effects of astringency, tonifying qi, and nourishing the kidney. The above fully demonstrates the chemical diversity of the Kadsura species and the unique chemical properties of each species.
These species are integrated into various regional folk medicine practices, although their specific applications and therapeutic effects vary. Preliminary analyses suggest that these discrepancies may arise from differences in chemical compositions, growing environments, harvesting seasons, and the theoretical and practical frameworks of traditional medicine, which contribute to their unique medicinal profiles. Ongoing research continues to reveal the diverse ethnic uses and potential applications of the genus Kadsura, providing new insights and resources for modern drug development.
Cultivation
2.3
Propagation
2.3.1
Scholarly investigations into propagation techniques for the genus Kadsura, including cutting propagation, seed germination, and tissue culture, have largely been confined to a narrow subset of species, leading to significant disparities in the depth and breadth of research across taxa. With respect to cutting propagation, K. ananosma exhibits optimal growth and survival rates of up to 86% when cultured in a substrate consisting of 35% forest topsoil blended with 65% perlite (Li and Liu, 2014). For K. longipedunculata, stem cuttings measuring 9∼10 cm in length yield survival rates exceeding 60%, with thicker stem segments positively correlating to improved survival outcomes (Lin et al., 2015). In contrast, K. coccinea has been the subject of the most intensive research on cutting propagation. Suitable substrates include humus-rich soil and slightly acidic loam (Wu, 2018; Zhang, 2018), while pre-treatment of stem segments with 100 mg/L naphthalene acetic acid (NAA) significantly enhances rooting efficiency (Huang et al., 2018). Furthermore, when cuttings are grown in a plastic greenhouse with 70% light transmittance, survival rates can exceed 95%, underscoring the critical role of environmental conditions in cutting propagation success (Lin and Zhou, 2021). Moreover, field cultivation with a planting density of 1.5 m × 2 m and a single-layer trellis system results in a survival rate of 68.3% (Yin et al., 2017).
Turning to seed propagation, most research on Kadsura species has focused exclusively on K. coccinea. Studies demonstrate that seed treatment with either a mixed solution of 0.5% CaCl_2_ and 50 μg/mL gibberellic acid (GA) (Zhao and Zhang, 2011) or a 30 mg/L GA solution alone (Li and Liang, 2018) can boost germination rates to over 90%. Soaking seeds in optimal concentrations of plant growth regulators not only breaks seed dormancy and stimulates germination but also enhances seedling vigor; notably, the combined application of 200 mg/L 6-benzyladenine (6-BA) and 100 mg/L salicylic acid (SA) further improves germination efficacy (Jiang, 2023). Additional effective dormancy-breaking treatments include: soaking seeds in concentrated sulfuric acid for 30 s (83.77% germination rate), a 60 °C hot water bath (68.53% germination rate), cryogenic treatment in liquid nitrogen for 20 min (82.47% germination rate), and immersion in 100 mg/L GA solution (89.96% germination rate) (He, 2022). Furthermore, 60 days of cold stratification followed by 30 days of incubation results in a germination rate of 78.18% (He, 2022).
In comparison, research on tissue culture techniques for Kadsura species remains relatively scarce, with current studies limited to K. coccinea. Lu et al. (2022) identified optimal media for K. coccinea shoot tip culture: 1/2 MS medium supplemented with 2.0 mg/L 6-BA and 0.2 mg/L indole-3-acetic acid (IAA) for primary culture, and 1/2 MS medium with 3.0 mg/L 6-BA and 0.1 mg/L 3-indolebutyric acid (3-IBA) for subculture. Deng et al. (2024) explored seed-based tissue culture for K. coccinea, developing specialized media for proliferation and rooting that achieved a 100% rooting rate and robust seedling growth. They also established optimal acclimatization durations, transplant substrates, shading protocols, and cultivation cycles for tissue-cultured plantlets.
Cultivation technology
2.3.2
Research on cultivation techniques for medicinal plants of the Kadsura genus has focused on two species: K. coccinea and K. longipedunculata. Among these, K. coccinea has been the most extensively studied. Two primary planting seasons have been identified for K. coccinea: spring (late February to early March) and autumn (late October to early November) (Chen, 2015; Tao et al., 2002). This species demonstrates shade tolerance, strong stress resistance, and adaptability to a wide temperature range (both low and high) (Qi et al., 2014; Yin et al., 2017; Liang et al., 2017). Optimal growth conditions include slightly acidic sandy soils or loose, fertile loam, paired with a light intensity of 30%–50% (Wu, 2012).
Furthermore, soil phosphorus content, cation exchange capacity, and pH are recognized as critical yield determinants (Liang and Li, 2019). Thus, soil selection, targeted fertilization, and pH adjustment are essential for successful cultivation and optimal yield. For K. longipedunculata, seeds should be collected between September and October upon full fruit maturation and sown via row-sowing from March to April. And we recommend transplanting in the autumn of the same year or in the spring of the following year (Wang, 2013).
The impact of environmental factors
2.3.3
Research shows that environmental factors, such as light, water, and altitude, significantly influence the growth of Kadsura plants. For example, Zhong et al. (2009) suggested that moderate shading is beneficial for K. japonica growth, as it enhances the activity of various enzymes under these conditions. In the case of K. coccinea, light plays a particularly critical role. The optimal light intensity for its growth ranges between 30% and 50% (Qi et al., 2014; Zhao et al., 2021), and it is advisable to cultivate this species in areas with ample water sources to ensure robust development (Yang et al., 2020). Additionally, K. ananosma can grow normally in tropical regions at an altitude of 1,300 m (Yang et al., 2016). In Xishuangbanna, it is best to select plots at an altitude of about 1,000 m for optimal growth (Yuan et al., 2019). Wu et al. (2022) noted that K. longipedunculata is more suited to growing on shady or semi-shady slopes with high humidity. It thrives in loose, well-aerated sandy loam, gravelly soil, or yellow clay soil at altitudes below 800 m.
In summary, researchers have conducted studies on cuttings propagation techniques for various species of the genus Kadsura, including K. ananosma, K. longipedunculata, and K. coccinea. The survival rate of cuttings depends not only on the quality of the cutting material itself but also on factors such as the selection of rooting substrates, the application of rooting hormones, and field management practices. Significant progress has been made in the cultivation research of multiple Kadsura species: standardized propagation protocols for cuttings and seeds have been established for species such as K. coccinea, achieving cuttings survival rates exceeding 95% and seed germination rates over 90%. Additionally, cultivation techniques have identified key conditions, including planting seasons, soil types, and light requirements, while environmental adaptability studies provide a scientific basis for precise site selection, thereby offering solid support for large-scale cultivation.
While these advancements lay a solid foundation for practical application, notable limitations persist in the current research landscape: uneven species coverage remains a critical issue, with in-depth studies concentrated on K. coccinea and research gaps across most other species; tissue culture technology is restricted to K. coccinea and remains underdeveloped overall; environmental factor analysis primarily focuses on single variables, lacking investigations into multi-factor interactions; and comprehensive comparative studies on alternative propagation methods and cross-regional cultivation techniques are insufficient. These gaps collectively hinder the full development, utilization, and technological dissemination of the genus as a whole.
Pharmacology
3
Hepatoprotective effects
3.1
Metabolites derived from Kadsura species exert hepatoprotective effects by targeting three core signaling networks, with distinct species-specific variations in pathway prioritization and regulatory mechanisms. First, the regulation of oxidative stress through activation of the Nrf2/ARE signaling axis represents a conserved hepatoprotective mechanism across multiple Kadsura species. For instance, the ethanol extract of K. coccinea has been shown to robustly upregulate the expression of superoxide dismutase (SOD) and glutathione peroxidase (GPX-2), while concurrently suppressing the levels of malondialdehyde (MDA) and myeloperoxidase (MPO) in the livers of mice with carbon tetrachloride (CCl_4_)-induced injury (Yu et al., 2021). Mechanistically, this protective effect is likely mediated by binankadsurin A that directly interacts with Kelch-like ECH-associated protein 1 (Keap1) to facilitate the nuclear translocation of Nrf2. However, direct binding assays are required to validate this proposed mechanism experimentally.
Second, the inhibition of inflammatory responses through the NF-κB and MAPK signaling pathways exhibits notable species-specific divergence. Specifically, K. coccinea suppresses the secretion of pro-inflammatory cytokines TNF-α and IL-6 by reducing the phosphorylation of c-Jun N-terminal kinase (JNK) and p38 mitogen-activated protein kinase (p38) (Wu et al., 2024; Li et al., 2010; Liu et al., 2015; Qu et al., 2003; Zhu, 2007; Liu et al., 2018c; Liu et al., 2018d); in contrast, K. heteroclita harbors hepatoprotective metabolites, including kadsurarin, meso-dihydroguaiaretic acid, kadsuphilol C, and coumarinlignan (Wang et al., 2022; Su et al., 2015), these may also function through the same mechanism. Third, the attenuation of hepatic fibrosis through HSC inactivation is a key hepatoprotective feature observed in several Kadsura species. For example, extracts of K. coccinea inhibit the proliferation of HSC-T6 cells and induce apoptosis through lignan metabolites, such as acetylpigomisin R (Ban et al., 2009). Similarly, certain metabolites in K. longipedunculata, including kadsuraols C, schiarisanrin, micrandilactone I, B22,23-di-epi-micrandilactone J, and longipedlignans F, also play significant roles (Shao et al., 2020; Wang et al., 2017; Wang et al., 2018; Liu et al., 2016; Liu et al., 2018a). Collectively, these findings support the overarching hypothesis that the structural diversity of Kadsura metabolites determines their pathway selectivity: lignans preferentially target oxidative stress and inflammatory responses, whereas terpenoid-lignan hybrids exhibit superior efficacy in inhibiting fibrosis-related HSC activation.
The species-specific profiles of bioactive metabolites further highlight the significant therapeutic potential of the Kadsura genus. For instance, K. coccinea primarily relies on acetylated lignans (acetylpigomisin R, binankadsurin A) to protect hepatocytes against tert-butyl hydroperoxide-induced damage, with acetylation modifications significantly enhancing their bioactivity (Ban et al., 2009). In contrast, K. heteroclita exerts its hepatoprotective effects through the combined action of metabolites (meso-dihydroguaiaretic acid, kadsurarin), which synergistically modulate oxidative stress and inflammatory signaling pathways (Su et al., 2015). On the other hand, the terpenoid-lignan hybrid metabolites of K. longipedunculata (micrandilactone I, longipedlignans F) demonstrate superior anti-fibrotic activity when compared to single-class metabolites (Liu et al., 2018a). These species-specific differences underscore the critical need for targeted research strategies to optimize the selection of lead metabolites for the treatment of distinct liver disease subtypes.
Despite the promising preclinical findings, the translation of Kadsura-derived agents into clinical practice is hindered by several critical bottlenecks. First, most lignans exhibit low oral bioavailability, primarily due to their poor water solubility and susceptibility to first-pass metabolism. To address this issue, structural modifications or formulation strategies, such as nanoliposome encapsulation, may be employed to improve bioavailability and delivery efficiency. Second, significant gaps in standardization persist across Kadsura extract preparations, as variable extraction protocols yield inconsistent metabolite profiles. Therefore, the development of marker-based quality control standards. For example, using kadsurarin as a marker metabolite for K. heteroclita extract is crucial to ensure the reproducibility of preclinical and clinical studies.
Most importantly, to date, no large-scale clinical trials have been conducted to validate the safety and efficacy of Kadsura-derived metabolites for the treatment of liver diseases. Accordingly, future research should prioritize the initiation of phase I/II clinical trials in patients with non-alcoholic steatohepatitis (NASH) or drug-induced liver injury, with primary endpoints including changes in liver enzyme levels and improvements in fibrosis staging. By systematically addressing these translational challenges and rigorously testing the proposed mechanistic hypotheses, species of the Kadsura genus hold significant potential to expand the current therapeutic arsenal for the management of liver disorders.
Antitumor effects
3.2
Multiple studies have confirmed that extracts from Kadsura species exert robust anticancer effects. K. heteroclita stands as one of the most thoroughly investigated species within the genus, with its lignan and triterpenoid metabolites exhibiting robust cytotoxicity against a broad panel of cancer cell lines. For instance, the lignan (+)-1-hydroxy-2,6-bis-epi-pinoresinol and triterpenoid xuetonglactone F from K. heteroclita significantly inhibit the viability of BGC-823 gastric cancer cells and HeLa cervical cancer cells, with notable selectivity compared to normal somatic cells (Shehla et al., 2019; 2022). Furthermore, crude triterpenoid extracts from K. heteroclita have been shown to induce caspase-dependent apoptosis in multiple human cancer cell lines, including OVCAR, HT-29, Hep-G2, and A549, with IC_50_ values ranging from 16.2 to 36.4 µM (Xu, 2006; Minh et al., 2014). Notably, among its purified triterpenoid metabolites, longipedlactones A and F exhibit exceptionally potent cytotoxicity against Hep-G2 and Bel-7402 hepatocellular carcinoma cell lines, highlighting their potential as lead metabolites for subsequent drug optimization and development (Xu et al., 2010).
Beyond K. heteroclita, other Kadsura species contribute a structurally diverse repertoire of antitumor metabolites, with activities spanning both solid tumors and hematological malignancies. For example, volatile oil metabolites isolated from K. longipedunculata, such as camphene and borneol, exhibit moderate cytotoxicity against Hep-G2 hepatocellular carcinoma, MIA-Paca-2 pancreatic cancer, and SW-480 colorectal cancer cells, with IC_50_ values of 133.53, 136.96, and 136.62 mg/mL, respectively (Mulyaningsih et al., 2010). Moreover, the triterpenoid-rich fraction of K. longipedunculata, including metabolites like kadlongilactones A–F and longipedlactones A–C, F, H, demonstrates potent cytotoxicity against K562 chronic myeloid leukemia cells (IC_50_: 0.84–11.38 µM) and solid tumor cell lines such as A549 lung cancer and HT-29 colorectal cancer (IC_50_: 0.49–3.61 µM) (Pu et al., 2005; 2007; Song et al., 2021). Furthermore, penochrochlactone C, a unique triterpenoid isolated from K. longipedunculata, inhibits the proliferation of HeLa cervical cancer cells with an IC_50_ of 9.70 µM (Song et al., 2021). Similarly, taiwankadsurin B, a neolignan from Kadsura philippinensis Elmer, exhibits dose-dependent cytotoxicity against a range of human tumor cell lines, further expanding the structural diversity of antitumor agents within the genus (Shen et al., 2005).
In addition to their direct cytotoxic effects against established cancer cells, Kadsura species also hold significant chemopreventive potential by targeting key tumor-promoting pathways. For example, fourteen neolignans isolated from various Kadsura species have been identified as potent inhibitors of 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced Epstein-Barr virus early antigen (EBV-EA) activation in Raji cells, with neokadsuranin and schisandrin C displaying the highest inhibitory potency (Chen D. et al. 2002). Critically, in vivo studies have validated the therapeutic efficacy of Kadsura extracts in preclinical animal models. For instance, the chloroform extract of K. coccinea significantly suppresses tumor growth and induces apoptosis in a dose-dependent manner in a murine 4T1 breast cancer xenograft model. Mechanistically, this extract exerts its antitumor effects by modulating the MAPK signaling cascade—specifically by upregulating the phosphorylation of ERK, JNK, and p38 MAPK, while concurrently regulating cytokine expression profiles (upregulating pro-inflammatory mediators IL-6, IL-1β, iNOS, and TNF-α; downregulating COX-2 to reshape the tumor microenvironment into an antitumor state (Jin, 2022).
Collectively, these preclinical findings robustly underscore the immense potential of Kadsura species as a prolific source of novel anticancer and chemopreventive agents. The structural diversity of their bioactive metabolites, combined with their multi-targeted mechanisms of action, positions these metabolites as promising candidates for further structural optimization, rigorous in vivo validation using immunocompetent models and patient-derived xenografts (PDXs), and eventual translation into clinical trials to address critical unmet needs in cancer therapy and chemoprevention.
Antimicrobial effects
3.3
To date, three Kadsura species have been scientifically validated for their antibacterial properties: K. coccinea (Feng et al., 2011; Yuan and Zhong, 2009), K. longipedunculata (Li, 2015; Lin et al., 2017; Lin et al., 2019; Liu et al., 2013; Wang et al., 2013; Wu et al., 2012; Lin et al., 2018), and K. angustifolia (Song et al., 2021), each exhibiting distinct species-specific antibacterial profiles that are tightly correlated with their unique secondary metabolite compositions.
Specifically, the ethanolic extract of K. coccinea fruit peel exhibits selective inhibitory activity against Gram-negative Salmonella typhi (Feng et al., 2011) and Gram-positive Streptococcus pneumoniae (Yuan and Zhong, 2009), though its inhibitory efficacy against clinically relevant pathogens such as Bacillus cereus and methicillin-resistant Staphylococcus aureus (MRSA) remains relatively limited. In contrast, K. longipedunculata represents the most extensively investigated species within the genus, with its aqueous and ethanolic extracts demonstrating robust broad-spectrum antibacterial activity against both Gram-positive and Gram-negative bacterial pathogens; notably, it exerts potent inhibitory effects against multidrug-resistant Staphylococcus aureus (MRSA), a leading cause of nosocomial and community-acquired infections (Li, 2015; Liu et al., 2013; Wang et al., 2013). Meanwhile, K. angustifolia, a relatively understudied yet promising species, exhibits moderate antibacterial activity against a panel of clinically significant pathogens, including methicillin-sensitive Staphylococcus aureus (MSSA), Bacillus subtilis, Escherichia coli, and Pseudomonas aeruginosa (Song et al., 2021).
While these findings demonstrate that Kadsura species possess species-specific antibacterial properties, highlighting their differentiated therapeutic potential, current studies primarily rely on in vitro experiments. Such approaches neither replicate host environments nor assess drug resistance evolution, potentially leading to biased antimicrobial efficacy. Furthermore, evaluations of metabolic stability and in vivo distribution remain insufficient. Therefore, integrating in vivo experiments with clinical studies is essential to validate their therapeutic potential and safety comprehensively.
Antioxidant effects
3.4
Extensive preclinical investigations have systematically validated the robust antioxidative activity of several key Kadsura species. For instance, the lignans from K. coccinea exhibit multifaceted antioxidative effects: they directly scavenge free radicals including DPPH, ·OH, and superoxide anions; upregulate the activity of endogenous antioxidant enzymes such as SOD; and modulate oxidative stress signaling pathways to suppress melanin synthesis, highlighting potential applications in both disease treatment and cosmetic formulations (Goh et al., 2013; Liu et al., 2018e; Ai and Li, 2005; Li et al., 2020; Tang, 2020; Wang, 2022; Yan et al., 2013; Zuo, 2021). Concurrently, K. heteroclita produces a distinct array of antioxidative metabolites, including cytochalasin H, isolariciresinol, and gomisin J, which demonstrate potent reactive oxygen species (ROS)-scavenging capacity and protect against oxidative damage in cellular models of neurodegeneration and liver injury (Shehla et al., 2022; Cao et al., 2019).
Current research on the sources of antioxidants in Kadsura remains predominantly descriptive, with most studies relying on simple in vitro experiments (free radical scavenging assays) that fail to replicate the complex in vivo physiological environment of oxidative stress. Additionally, its underlying molecular mechanisms remain unelucidated.
Anti-inflammatory and analgesic effects
3.5
Current research on the anti-inflammatory and analgesic activities of Kadsura species primarily focuses on two species: K. coccinea and K. heteroclita (Li et al., 2006; Yang et al., 2021; Yang et al., 2022; Hu and Chen, 2018; Ji, 2011; Li et al., 2014; Liu and Wang, 1989; Lu, 2021; Shi and Li, 2016; Sun, 2020; Sun et al., 2022; Li, 2023; Yu et al., 2019a; Yu et al., 2019b; Yu, 2017; Zheng et al., 2024). K. coccinea exerts broad-spectrum anti-inflammatory and analgesic activity via multi-pathway regulation: in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages and collagen-induced arthritis (CIA) mouse models, its lignan-rich extracts inhibit the NF-κB and COX-2 signaling cascade to reduce the production of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6), activate the nuclear factor erythroid 2-related factor 2 (Nrf2) antioxidant pathway to mitigate oxidative stress, and directly suppress RA synoviocyte proliferation and osteoclast differentiation, thereby alleviating joint destruction and pain (Lu et al., 2021; Li et al., 2023). In contrast, K. heteroclita demonstrates targeted activity particularly relevant to bone-related inflammatory disorders: its unique triterpenoid metabolites (xuetonglactones A/B, xuetongsu) specifically inhibit inducible nitric oxide synthase (iNOS), all COX subtypes, and bone resorption-related proteins (MMP-9, CTSK, TRAP), with IC_50_ values ranging from 3.2 to 12.8 µM against key inflammatory enzymes. This makes it a promising candidate for the treatment of RA and osteoporosis, where excessive bone resorption drives disease progression (Shehla et al., 2022; Cao et al., 2019).
However, current research on K. longipedunculata and K. interior remains relatively limited; the mechanism of anti-inflammatory action has not been elucidated, most studies rely on in vitro assays or rodent models with limited pharmacokinetic data, and no clinical trials have validated human safety or efficacy (Tan et al., 2017; Zhang et al., 1991; Tang et al., 2024; Li et al., 1999b). To facilitate more effective development and utilization, future work should use multi-omics to identify precise molecular targets, validate in vivo efficacy in disease-relevant models, optimize formulations to enhance bioavailability, and advance clinical trials to unlock Kadsura’s potential as novel anti-inflammatory and analgesic therapies.
Anti-HIV effects
3.6
Species of the genus Kadsura have emerged as a promising source of natural anti-HIV agents, with triterpenoids and lignans identified as the primary bioactive metabolites exhibiting diverse mechanisms of action against HIV-1 replication (Sun et al., 2011; Gao et al., 2008). Triterpenoid extracts from K. heteroclita demonstrate selective cytotoxicity against HL-60 human promyelocytic leukemia cells. This activity is hypothesized to indirectly suppress HIV replication by modulating immune cell populations and reducing viral reservoir formation in infected hosts (Xu, 2006; Xu et al., 2010). Meanwhile, K. longipedunculata produces a suite of potent anti-HIV-1 metabolites with distinct modes of action: schisanlactone A, longipedunin A, and lancilactone C inhibit early-stage HIV-1 entry and post-entry viral processing, while the dibenzocyclooctadiene lignan kadlongirin B directly targets the HIV-1 replication machinery, likely interfering with reverse transcriptase activity or viral integration into host chromatin (Sun et al., 2006; Pu et al., 2008). Notably, these metabolites exhibit low cytotoxicity against normal human peripheral blood mononuclear cells (PBMCs), suggesting a favorable therapeutic index for further drug development.
However, the clinical translation of anti-HIV agents derived from the genus Kadsura still faces two critical bottlenecks. On the one hand, most existing studies rely on simple in vitro assay systems that cannot accurately replicate the complex interactions between HIV and the human immune system, leaving the core molecular mechanisms underlying their anti-HIV activity incompletely elucidated. On the other hand, in vivo efficacy, pharmacokinetic profiles, and long-term safety data of these agents remain severely lacking. Therefore, multi-dimensional translational research strategies are required to drive breakthroughs in future studies. First, structural biology techniques should be employed to identify the precise molecular targets of lead metabolites. Second, in vivo validation should be conducted in humanized mouse models of chronic HIV infection to systematically evaluate the agents’ inhibitory effects on viral load, regulatory roles in viral reservoirs, and tissue tropism. Meanwhile, structure-activity relationship (SAR) studies should be conducted to optimize metabolite structures, enhancing antiviral activity, metabolic stability, and oral bioavailability while reducing cytotoxicity. Through systematic exploration of the medicinal potential of Kadsura plants, it is expected to supplement existing antiretroviral therapy (ART) with novel mechanism-oriented anti-HIV lead metabolites, advancing the clinical translation of natural-source anti-HIV agents.
Antiplatelet aggregation effects
3.7
Studies have demonstrated the antiplatelet aggregation potential of Kadsura species. Specifically, seco-coccinic acid I, isovaleroylbinankadsurin A, and acetyl-binankadsurin A from K. coccinea, as well as heteroclitin D, (+)-anwulignan, meso-dihydroguaiaretic acid, and acid-kadsulignan L from K. heteroclita and K. angustifolia, all demonstrate significant antiplatelet aggregation activity (Association and Prevention, 2022; Su et al., 2017; Li et al., 1999a; Chen et al., 1998). Notably, among these metabolites, heteroclitin D effectively inhibited vascular constriction responses induced by high-potassium depolarization, calcium chloride, and norepinephrine (NA), and exerted a more pronounced inhibitory effect on platelet aggregation (Li et al., 1999a). Additionally, meso-dihydroguaiaretic acid and acid-kadsulignan L demonstrated antagonistic activity against platelet-activating factor (PAF) (Chen et al., 1998).
Platelet aggregation plays a pivotal role in activating blood circulation and eliminating blood stasis through the action of Kadsura plants (Jiang et al., 2005). This discovery not only explains the effectiveness of Kadsura genus plants in traditional Chinese medicine for treating various cardiovascular diseases but also opens up new avenues for modern medicine. However, the research on its mechanisms is not sufficiently in-depth; no studies have been conducted on toxicity or long-term effects, and individual differences have not been adequately considered. Future research should focus on elucidating mechanisms, optimizing doses, and conducting long-term toxicity and individual-difference studies to enhance the reliability of research findings and their clinical application.
Anti-leukemia effects
3.8
Current research has identified four Kadsura species with anti-leukemic activity: K. coccinea (Wang et al., 2008), K. heteroclita (Wang et al., 2006), K. longipedunculata (Liu and Huang, 1991; Liu and Pan, 1991), and K. ananosma (Chen et al., 2006; Yang et al., 2010). However, significant interspecific differences have been observed in their bioactive metabolite profiles and anti-leukemic efficacy. For instance, seco-coccinic acids A-C and seco-coccinic acid E isolated from K. coccinea significantly inhibit the proliferation of HL-60 human promyelocytic leukemia cells, with IC_50_ ranging from 6.8 to 42.1 µmol/L (Wang et al., 2008). In contrast, K. longipedunculata produces changanic acid, schisanlactone E, and schisanlactone F, which exhibit inhibitory effects on murine P-388 leukemia cells with IC_50_ values of 10 µg/mL, 1 µg/mL, and 5 µg/mL, respectively (Liu and Huang, 1991; Liu and Pan, 1991). Notably, while K. heteroclita and K. ananosma have been reported to possess anti-leukemic activity, their specific bioactive metabolites and underlying mechanisms remain largely uncharacterized, leaving critical gaps in our understanding of their therapeutic potential.
Neuroprotection effects
3.9
Plants of the genus Kadsura also exhibit notable neuroprotective potential. For instance, extracts from K. heteroclita significantly promote the growth and development of hippocampal neurons (Xiao et al., 2002). Ananolignans F and L isolated from K. ananosma exhibit potent neuroprotective effects (Yang et al., 2011). Additionally, specific metabolites from K. japonica and Kadsura polysperma Y.C.Yang also exert protective effects against neuronal injury (Dong et al., 2012; Liu et al., 2023).
Existing research lacks systematic identification of the active metabolites responsible for their neuroprotective effects, as well as studies on the underlying mechanisms. To advance the development and use of botanical drugs, future efforts should integrate modern research methodologies, such as multi-omics and single-cell technologies, to further explore their pharmacological value. This will facilitate the translation from basic research to clinical applications, enabling a shift from single-metabolite mechanism elucidation to multi-target system intervention. Such progress will provide novel botanical drug options for the treatment of neurodegenerative diseases.
Others
3.10
Beyond their shared pharmacological properties, individual Kadsura species exhibit distinct therapeutic potentials. For instance, the fruits of K. coccinea have the potential to prevent hyperlipidemia. The bitter taste-removing oils and polysaccharides in its fruits exert lipid-lowering effects. The underlying mechanism may involve enhancing the activity of superoxide dismutase, glutathione peroxidase, and catalase while reducing the levels of malondialdehyde, total cholesterol (TC), triglycerides (TG), and low-density lipoprotein cholesterol (LDL-C) (Zhang et al., 2022; Long et al., 2022). Additionally, its fruit extracts have preventive effects against bacterial diarrhea and effectively mitigate senna-induced diarrhea (Ling, 2021). Furthermore, the alcohol extract inhibits multiple venom enzymes from K. coccinea (Huang et al., 2020). For K. heteroclita, the leaf essential oil (EO) and its major chemical metabolites (δ-Cadinene, Calarene, and δ-4-Carene) exhibit varying degrees of cytotoxicity toward larvae of three mosquito vector species (Ling, 2021). These findings provide a basis for developing new, safer natural larvicides to combat diseases such as malaria and dengue fever. Meanwhile, the metabolites anwuweizonic acid, coccinic acid, and manwuweizic acid, isolated from the stems of K. angustifolia, exert significant antifertility effects (Chen et al., 2001; Chen Y. et al. 2002).
In summary, plants of the genus Kadsura not only exhibit common pharmacological effects such as hepatoprotective (Qu et al., 2003), antibacterial (Feng et al., 2011), antioxidant (Ai and Li, 2005), antitumor (Jin, 2022), and anti-HIV(Xu, 2006) activities but also demonstrate diverse pharmacological properties, including lipid-regulating (Zhang et al., 2022), antidiarrheal (Ling, 2021), antifertility (Chen et al., 2001), anticomplementary (Huang, 2010), and insecticidal (Govindarajan et al., 2016) effects. These findings highlight the potential therapeutic value of various metabolites in Kadsura plants, indicating broad prospects for their development and utilization. Furthermore, these research outcomes provide a robust theoretical foundation for future drug development based on the Kadsura species. A recapitulative summary is presented in Table 2 and Figure 2.
Main pharmacological effects of the Kadsura gene and their corresponding effect mechanisms.
Molecular pharmacognosy
4
Chloroplast genome and phylogeny
4.1
Currently, there has been relatively extensive research on the chloroplast genome of Kadsura species (Figure 3). In 2017, Guo (2017) conducted the first study on the chloroplast genomes of multiple Kadsura species, revealing that K. heteroclita, K. longipedunculata, K. japonica, K. interior, and K. oblongifolia share close phylogenetic relationships. Subsequently, numerous studies on the chloroplast genome of Kadsura have been published, with detailed information summarized in Table 3. Table 2 shows that the chloroplast genome lengths of Kadsura species range from 145,399 bp to 153,362 bp, with the longest being K. heteroclita and the shortest being K. coccinea. The lengths of the LSC, IR, and SSC regions range from 85,243 bp to 94,757 bp, 16,431 bp to 24,673 bp, and 17,945 bp to 18,253 bp, respectively. The total number of genes ranges from 113 to 129, with protein-coding genes, rRNA genes, and tRNA genes ranging from 79 to 85, 4 to 8, and 30 to 37, respectively.
The chloroplast genome of Kadsura species.
In addition, researchers conducted the phylogenetic analysis of the Kadsura plant and its closely related species using chloroplast genomes. For example, Wang et al. (2003) were the first to perform a sequence analysis of the chloroplast rbcL gene in Schisandraceae plants. Their study suggested a close relationship between the genera Schisandra Michx. and Kadsura, with interconnections and overlaps, leading to the hypothesis that the two genera may share a common ancestor. Li (2019) categorized the boundaries of the IR region in Schisandraceae into the S type and the L type, and species of the genus Kadsura include both types. Specifically, K. coccinea has an S-type IR region, whereas the other Kadsura species have an L-type IR region. Moreover, phylogenetic analysis shows that the genera Kadsura and Schisandra are non-monophyletic. It is also recommended that Schisandra plena A.C.Sm. and Schisandra propinqua (Wall.) Baill. be transferred to the genus Kadsura. From these results, it can be inferred that the genera Schisandra and Kadsura are non-monophyletic and probably share a common origin.
As can be seen from the above, researchers have completed the sequencing assembly and structural analysis of multiple chloroplast genomes of Kadsura species, and have used these data to conduct partial phylogenetic evolution analysis of Kadsura and its close relatives, providing a good data foundation for the later development of DNA barcode labeling and molecular marker-assisted breeding of this genus.
Transcriptional regulatory networks and functional genes
4.2
Research on secondary metabolic pathways and functional genes in Kadsura species is burgeoning. For example, Liang et al. (2022) conducted targeted metabolomic analysis combined with transcriptomic sequencing of the roots, stems, and leaves of K. coccinea. The metabolomic analysis identified 51 lignans, while transcriptomic sequencing revealed 137 unique genes across 13 categories of lignan biosynthesis. It was found that the CCoAOMT, C3H, and SIDR gene families were primarily expressed in the roots and stems. Further integrated analysis of metabolomics and transcriptomics identified 11 key enzyme gene families closely associated with lignan synthesis, including HCT, DIR, COMT, CAD, SIDR, and PLR. These findings demonstrate that K. coccinea roots, stems, and leaves contain numerous lignans and their associated synthesis enzyme genes, but the content and expression levels of these lignans and enzymes vary significantly among plant parts. Similarly, Fu et al. (2024) used transcriptomic sequencing to analyze root, stem, and leaf samples of K. coccinea and identified CYP genes, and conducted phylogenetic analyses of these genes. They were distributed across 8 clades within 38 families.
The roots exhibited specific expression patterns for CYP genes, and sequence alignment identified 22 homologous single genes among these CYPs. Of these, six homologous genes of CYP719As and one gene of CYP81Qs were highly expressed in the roots. Wang et al. (2019) conducted transcriptome sequencing of the roots, stems, and leaves of K. coccinea, successfully cloning KcSQS and performing bioinformatics analysis. This provides a basis for further research into the biosynthesis of triterpenoids in K. coccinea. In addition, Li et al. (2022) studied the leaves of K. ananosma and obtained transcriptomic data, laying the foundation for exploring the biosynthetic pathways and functional genes of active metabolites in K. ananosma.
In conclusion, research on the transcriptional regulatory network and the functional genes of Kadsura plants is still in its infancy. The main research focus is on K. coccinea. People have only conducted preliminary screening and expression profiling studies on some key regulatory genes at the transcriptomic and metabolomic levels, and have not carried out further functional verification. However, relevant research on other species remains a blank.
Comparative genomic research
4.3
To date, there have been very few studies on the genomes of Kadsura plants (Xu et al., 2021). Estimated the genome sizes of four Kadsura species (K. interior, K. heteroclita, K. longipedunculata, and K. coccinea) using FCM. The comparison revealed minimal differences in genome size among K. interior, K. heteroclita, and K. longipedunculata, whereas K. coccinea showed a markedly distinct genome size. This indicates that genome size alone is insufficient for fully distinguishing these species. Dong (2022) studied a total of 107 individuals of K. interior, K. heteroclita, K. longipedunculata, K. oblongifolia, and K. coccinea to examine genetic diversity, phylogeny, and ecology using single-nucleotide polymorphisms (SNPs) generated via restriction site-associated DNA sequencing (RAD-seq). The results showed moderate differentiation among K. heteroclita, K. longipedunculata, and K. oblongifolia, and a high degree of genetic differentiation between K. interior and these species. The phylogenetic tree indicated that each species was monophyletic. Furthermore, the results of population genetic structure showed that there was admixture and gene flow among K. heteroclita, K. longipedunculata, and K. oblongifolia. Moreover, due to morphological similarities, “Flora of China” treated K. interior and K. heteroclita as conspecific. However, based on the above research, Dong (2022) proposed that K. interior should not be considered a synonym of K. heteroclita.
In addition, Lee et al. (2023) assembled the genomes of Schisandra repanda (Siebold and Zucc.) Radlk. and K. japonica using a combination of Nanopore and Illumina sequencing technologies. They employed these genomes for taxonomic studies and developed two InDel markers to differentiate S. repanda, K. japonica, and Schisandra chinensis (Turcz.) Baill. This methodology offers a new avenue for exploring the evolutionary relationships within the Schisandraceae family.
In summary, to date, only a preliminary assessment of genome size and degenerate genome sequencing analysis have been conducted on some plants of the Kadsura genus, and the resulting data have been used for species identification and phylogenetic analysis of the genus. Whole-genome sequencing and further research on molecular regulatory mechanisms, transcriptional regulatory networks related to plant growth and development, or secondary metabolites have not yet been carried out.
Conclusion and future prospects
5
From the above review, we can see that people consume the fruits of many Kadsura species, which have definite medicinal value, such as K. coccinea, K. heteroclita, and K. oblongifolia. Among them, K. coccinea is currently the most widely used, extensively studied, and largest cultivated species. In addition, most plants of the Kadsura genus have the effects of promoting qi circulation and relieving pain, activating blood circulation and removing blood stasis, dispelling wind and dampness. Meanwhile, some species also have unique medicinal properties, such as K. interior, K. japonica, K. oblongifolia, and K. ananosma—their unique chemical metabolites determine the distinctive medicinal value of these species. Previously, people have conducted relatively detailed research on the chemical metabolites of the genus Kadsura. However, to date, researchers have not conducted corresponding pharmacological activity verification and active metabolite analysis for the above-mentioned species with unique traditional medicinal value. Perhaps through these studies, novel active metabolites can be discovered, and data supporting the development of new medicines can be provided.
In terms of pharmacological activity, plants of the genus Kadsura not only exhibit common effects such as hepatoprotective, antibacterial, antioxidant, antitumor, and anti-HIV activities but also demonstrate diverse pharmacological properties, including lipid-regulating, antidiarrheal, antifertility, and insecticidal effects, among others. These findings highlight the potential therapeutic value of various metabolites in Kadsura plants, indicating broad prospects for their development and utilization. Furthermore, these research outcomes provide a robust theoretical foundation for future drug development based on the Kadsura species. A recapitulative summary is presented in Table 2 and Figure 2. It should be noted that the current research results indicate that plants of the Kadsura genus exhibit unique attributes in both traditional folk disease applications and modern metabolite composition and pharmacological activity studies. However, systematic comparative studies of different Kadsura species have not been conducted to elucidate the scientific significance of the unique attributes of individual species. It can be inferred that implementing such research will significantly advance the discovery of novel metabolites and the advancement of new drug development.
Furthermore, as can be seen from Tables 1, 3, many plants of the Kadsura genus have excellent medicinal value. However, the species that have been extensively studied in modern pharmacology are limited to a few, such as K. cocinea, K. heteroclita, and K. longipedunculata. As mentioned above, some Kadsura species have unique medicinal properties, such as K. interior, K. japonica, K. oblongifolia, and K. ananosma; however, pharmacological research on these species is relatively scarce. Therefore, we suggest that researchers shift their focus to these species to discover more valuable pharmacological activities and development potential.
Regarding propagation techniques, the most commonly used method for Kadsura plants is cutting propagation, and seed propagation is rarely adopted, which might be related to the fact that all Kadsura plants are vines (with longer growth cycles), and cutting propagation can shorten the growth cycle. Currently, research on seed propagation techniques has been reported only for K. coccinea. Research has shown that various methods can effectively break K. coccinea seed dormancy, promoting germination. However, whether these methods can also be applied to other Kadsura species requires further experimental verification. Research on tissue culture of the genus K. coccinea has just begun, and only partial work has been carried out on K. coccinea. Tissue culture is not only an effective method for enhancing breeding efficiency but also a fundamental tool for genetic transformation and functional gene research in plants. Therefore, it is imperative to conduct tissue culture for plants of the genus Kadsura.
In the field of molecular pharmacognosy, researchers have completed the sequencing assembly and structural analysis of multiple chloroplast genomes of Kadsura species, and have used these data to conduct partial phylogenetic evolution analysis of Kadsura and its close relatives, providing a good data foundation for the later development of DNA barcode labeling and molecular marker-assisted breeding of this genus. Meanwhile, research on the transcriptional regulatory network and the functional genes of Kadsura plants is still in its infancy; the main focus is on K. coccinea, and only preliminary screening and expression profiling studies have been conducted on some key regulatory genes at the transcriptome and metabolome levels. Furthermore, whole-genome sequencing and further research on molecular regulatory mechanisms underlying plant growth and development or secondary metabolite production have not yet been conducted. It is now extremely urgent to carry out whole-genome sequencing for Kadsura species with significant developmental value, as well as to verify functional genes and to develop molecular marker-assisted breeding based on this.
In conclusion, we propose three priority research directions: First, systematically validate species-specific pharmacological activities and characterize their bioactive metabolites; second, intensify research on tissue culture propagation techniques; third, accelerate whole-genome sequencing of priority Kadsura species, alongside gene screening, functional validation, and molecular regulatory network analysis, to lay a solid foundation for biosynthetic pathway elucidation and molecular marker-assisted breeding of elite cultivars.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ai Q. Li Y. (2005). Research progress on chemical metabolites and activities of Kadsura coccinea . Chem. Bioeng. (02), 7–9.
- 2Association G. O. A. A. Prevention C. C. F. D. (2022). Chinese guidelines for the diagnosis and treatment of AIDS (2021 edition). Peking Union Med. J. 13 (02), 203–226. 10.12290/xhyxzz.2022-0097 · doi ↗
- 3Ban N. K. Thanh B. V. Kiem P. V. Minh C. V. Cuong N. X. Nhiem N. X. (2009). Dibenzocyclooctadiene lignans and lanostane derivatives from the roots of Kadsura coccinea and their protective effects on primary rat hepatocyte injury induced by t-butyl hydroperoxide. Planta Med. 75 (11), 1253–1257. 10.1055/s-0029-1185537 19350485 · doi ↗ · pubmed ↗
- 4Cao L. Shehla N. Tasneem S. Cao M. Sheng W. Jian Y. (2019). New cadinane sesquiterpenes from the stems of Kadsura heteroclita . Molecules 24 (9), 1664. 10.3390/molecules 24091664 31035334 PMC 6539153 · doi ↗ · pubmed ↗
- 5Chen Y. (2015). Cultivation techniques of the wild fruit tree of Kadsura coccinea . Mod. Agric. (10), 35–36.
- 6Chen D. Zhang S. Kozuka M. Sun Q. Feng J. Wang Q. (2002). Interiotherins C and D, two new lignans from Kadsura interior and antitumor-promoting effects of related neolignans on epstein-barr virus activation. J. Nat. Prod. 65 (9), 1242–1245. 10.1021/np 0105127 12350139 · doi ↗ · pubmed ↗
- 7Chen Y. G. Qin G. W. Xie Y. Y. Cheng K. F. Lin Z. W. Sun H. D. (1998). Lígnans from Kadsura angustifolia . J. Asian Nat. Prod. Res. 1 (2), 125–131. 10.1080/10286029808039854 11261213 · doi ↗ · pubmed ↗
- 8Chen Y. Qin G. Cao L. Leng Y. Xie Y. (2001). Triterpenoid acids from Schisandra propinqua with cytotoxic effect on rat luteal cells and human decidual cells in vitro . Fitoterapia 72 (4), 435–437. 10.1016/s 0367-326x(01)00269-6 11395273 · doi ↗ · pubmed ↗