Biological Diversity and Parasitological Peculiarities of Myxosporea (Cnidaria, Myxozoa) Infecting Merluccius Merluccius (Linnaeus, 1758) in the Sea of Marmara
Derya Yadak, Cem Tolga Gürkanli, Sevilay Okkay, Yılmaz Çİftçİ, Ahmet Özer

TL;DR
This study explores myxozoan parasites infecting European hake in the Sea of Marmara, identifying new species and expanding their known geographic range.
Contribution
The study reports the first occurrence of Pseudalataspora vanderlingeni and a new Ceratomyxa taxon in Turkish waters.
Findings
Pseudalataspora vanderlingeni was identified in Turkish waters for the first time.
A new Ceratomyxa taxon was documented, differing from previously known species in spore morphology.
The findings suggest P. vanderlingeni is a common parasite of Merluccius species in the Mediterranean and adjacent seas.
Abstract
This study aimed to investigate the biodiversity and parasitological peculiarities of myxozoan parasites infecting European hake, Merluccius merluccius, one of the most commercially important demersal fish species, in the Sea of Marmara, Türkiye. A total of 47 M. merluccius specimens were collected from the eastern coasts of the Sea of Marmara and examined for myxozoan infections. Morphological observations, morphometric measurements, and phylogenetic analyses based on 18 S rDNA sequences were conducted to identify the parasites. Of the 47 fish specimens examined, 12 were infected by two distinct myxozoan species. One was identified as Pseudalataspora vanderlingeni based on morphology, morphometry, and molecular data. The second species, Ceratomyxa sp., which could be identified to the genus level based solely on spore morphology, was consistently found co-infecting fish hosts of the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
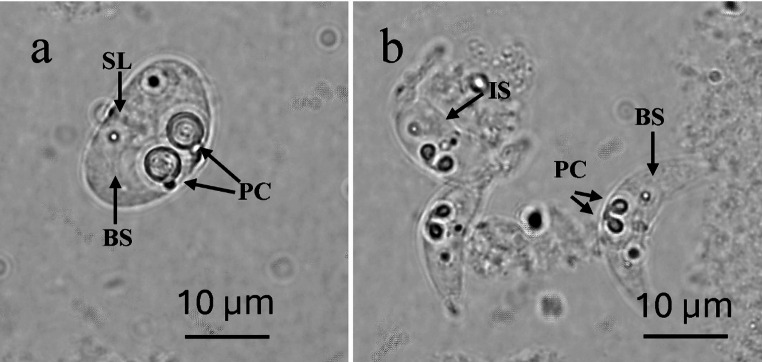 Figure 1
Figure 1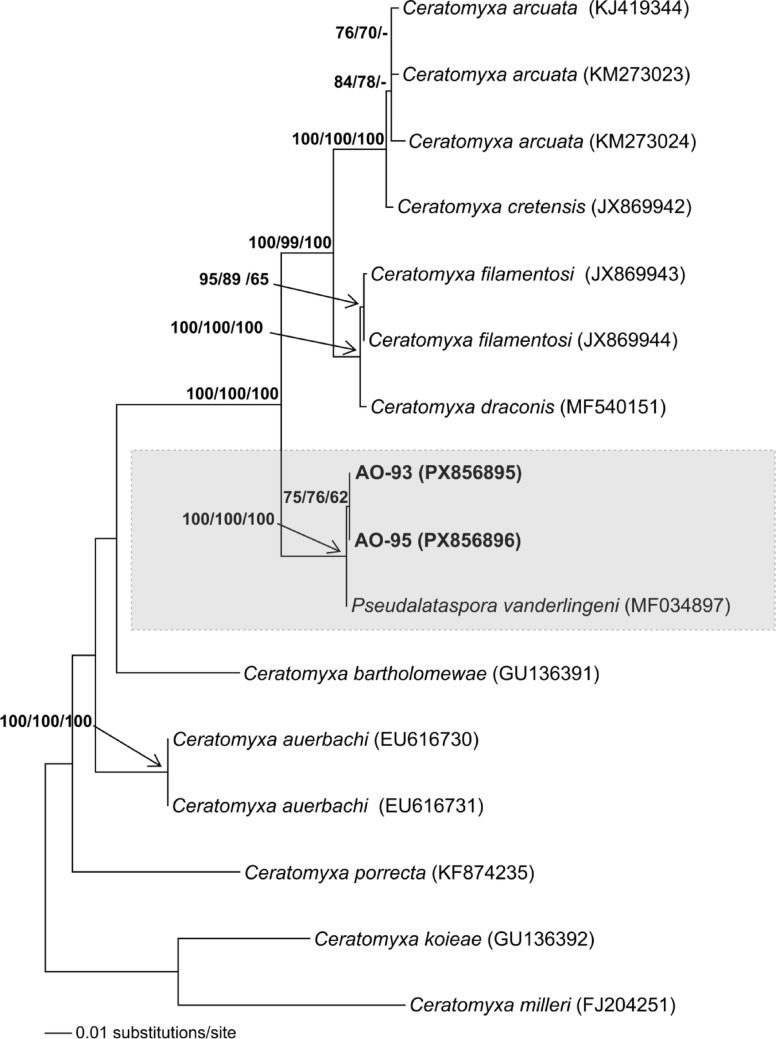 Figure 2
Figure 2- —Ordu University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMyxozoan Parasites in Aquatic Species · Parasite Biology and Host Interactions · Marine Invertebrate Physiology and Ecology
Introduction
The European hake, Merluccius merluccius (L., 1758), is one of the most economically important and therefore, one of the most heavily exploited demersal fish species in the Mediterranean Sea and its adjacent waters, including the Sea of Marmara [1, 2]. It is predominantly carnivorous and is generally found on muddy bottoms, with a wide depth distribution ranging from the coastline to 1000 m [3, 4]. It has a wide distribution across the Northeast Atlantic, extending from Norway to the Gulf of Guinea, the Mediterranean, and the Black Sea [1].
It has frequently been reported to be infected by metazoan parasites, including helminths and copepods [5–7]. In addition, members of another metazoan taxon, Myxozoa (Cnidaria), have also been reported as parasites of European hake and the other species of the genus Merluccius Rafinesque, 1810 [8–12]. Myxozoans are microscopic parasites with an extremely reduced body size and structure [13]. Currently, the class Myxozoa comprises more than 2,600 species across 65 genera and 17 families [14, 15]. Several myxosporean species have so far been reported from various hake species (Merluccius hubbsi-Argentine hake, M. capensis-Cape hake, M. paradoxus, M. australis-Southern hake, M. productus-Pacific hake, and M. gayi-Chilean hake) across countries with Atlantic and Pacific coastlines, including Argentina, Uruguay, South Africa, Canada, the USA, and the Mediterranean coastlines of France and Spain. These species include Kudoa rosenbuschi, K. thyrsites, K. paniformis, Myxoproteus meridianalis, Pseudalataspora vanderlingeni, Ceratomyxa globulifera, and Fabespora sp. Of these species, K. thyrsites and K. paniformis have been reported as the causative agents of post-mortem myoliquefaction (also known as milky, soft, or jelly flesh) in their hake hosts, resulting in significant economic losses [5, 7, 9–12, 16–21]. Despite its economic importance and relatively wide distribution, there are only two records of myxosporean infection in European hake M. merluccius: Ceratomyxa globulifera from the Mediterranean coast of France [17], and an unidentified coelozoic myxosporean parasite from the north-west Mediterranean coast of Spain [7].
Current literature lacks information on the diversity of myxosporean species infecting M. merluccius. Thus, this study aimed to elucidate the first biological diversity and infection characteristics of myxosporean species infecting M. merluccius in the Sea of Marmara, which lies adjacent to the Eastern Mediterranean Sea, as previously reported.
Materials and Methods
Fish Sampling and Parasitological Examination
Between January and December 2023, a total of 47 European hake, M. merluccius specimens (n: 17 in winter, n: 30 in summer), measuring 25.2–29.9 cm in length, were collected off the eastern coasts of the Sea of Marmara, specifically near Yalova and İzmit Bay (40°44′06′′ N, 29°′16′′ E), by local fishermen. Gills, fins, skin, urinary bladder, kidney, gallbladder, liver, intestine, smooth muscles, and gonads were microscopically examined for the presence of myxozoan parasites using a light microscope (Olympus BX51) equipped with a digital camera (DP71). Measurements of spore and polar capsule length and width were based on 30 fresh spores. All measurements were presented as mean values and min–max values in parentheses. Prevalence values (%) were calculated according to the definition by Bush et al. [22], while density values were determined using the density scale (1 + , 2 + , 3 + , 4 +) reported for myxosporean infections by Gürkanlı et al. [23].
Molecular Analyses
Species-level identification of the myxozoan morphotypes observed in the infected M. merluccius tissues was conducted using phylogenetic analyses based on nucleotide sequences of the small subunit of ribosomal DNA (18S rDNA hereafter). Two myxozoan specimens from each morphotype, representing different sampling seasons, were included in the molecular analyses. Total genomic DNA from myxozoa-infected tissues were extracted using the PureLink® Genomic DNA Mini Kit (USA), according to the manufacturer’s instructions. Extracted DNA was stored at − 20 °C before use. The PCR amplification of the 18S rDNA locus was performed using primers MyxospecF [24] and 18R [25]. For amplification, a 50 μl PCR reaction was prepared using genomic DNA (< 0.5 μg), 1.5 mM MgCl_2_ (Invitrogen), 1.25 U Taq polymerase (Invitrogen), 0.8 mM dNTP mix (Thermo Scientific), 1 × PCR buffer (Invitrogen), 0.4 pmol (final concentration) of each primer, and ddH_2_O (up to 50 μl). A Techne (TC-Plus, Staffordshire, UK) thermal cycler was used for the PCR amplification with the following conditions: an initial denaturation at 95 °C for 3 min, followed by 40 cycles of denaturation at 94 °C for 1 min, annealing at 51 °C (− 0.1 °C/Cyc) for 1 min, and extension at 72 °C for 2 min. The procedure was completed with a final extension at 72 °C for 10 min. Subsequently, the PCR products were electrophoresed on a 1% agarose gel (prepared in 1X TBE buffer) and visualized using a Vilber Lourmat photo-print imaging system (France). Nucleotide sequencing was performed commercially by Macrogen-Europe (Amsterdam, the Netherlands) on both strands, using the same primers as those employed for the PCR amplifications. Assemblage and editing of the nucleotide sequences from both strands were conducted using BioEdit v.7.0.5.3 [26]. For phylogenetic analyses, a dataset was assembled from BLAST (Basic Local Alignment Search Tool; https://blast.ncbi.nlm.nih.gov/Blast.cgi) search results and the available literature (as shown in the table provided in the Online Resource). Multiple nucleotide sequence alignment of the data set was performed using MAFFT v.7 [27, 28]. Software BioEdit v.7.0.5.3 and Gblocks v.0.91b [29] were used to edit the alignment file. Akaike information criterion [30] and Bayesian information criterion tests were performed using the jModelTest v. 0.1 [31] software package to determine the best-fitting evolutionary model(s) for the data set. Phylogenetic constructions were conducted using Maximum-Likelihood (ML), Neighbor-Joining (NJ), and Maximum-Parsimony (MP) algorithms. Software PhyML 3.0 [32]was used for the ML analyses, whereas PAUP* v. 4.0b10 [33] was employed for the NJ and MP analyses. A heuristic search approach with a TBR swapping algorithm (10 random repetitions) was implemented for the MP analysis. The reliability of the phylogenetic relationships were assessed using Bootstrap tests performed with 1000 replicates. The new 18S rDNA genotypes obtained in this study have been deposited in GenBank under accession numbers PX856895- PX856896.
Results
Morphology and Light Microscopy
Microscopic investigations of 12 infected fish hosts revealed two distinct myxospore morphotypes corresponding with the genera Pseudalataspora (Fig. 1a) and Ceratomyxa (Fig. 1b). The spore walls of all Pseudalataspora specimens observed consisted of two valve structures fused into an oval shape, with clearly visible and symmetrical margins. The valves exhibited a uniform, round-oval thickness and enclosed the binucleate sporoplasm (BS) and polar capsules (PC) evenly, indicating a symmetrical and regular valve architecture. Two polar capsules (PC) were oval with rounded ends, relatively large with a slight distance to one another, and were distinctly located near the center of the spore. They were positioned separately. The suture line (SL) between the valves was clearly defined, and two nuclei were present within the sporoplasm (Fig. 1a). The spores were measured, 14.0 μm (13.0–15.0 μm) in length, 19.6 μm (19.0–20.0 μm) in thickness, polar capsules 4.6 μm (4.0–5.0 μm) in length and 4.3 μm (4.0–5.0 μm) in width, polar capsule length: spore length = 1: 3.0–3.2; spore length: spore width = 1: 1.3–1.5. (Table 1). Due to similar spore morphologies and very close morphometric measurements, all Pseudalataspora specimens in this study were considered to belong to the same species, Pseudalataspora sp.-1.
Fig. 1. Spore morphology of myxosporeans (a) A fresh Pseudalataspora sp.-1 spore in valvular view, (b) Fresh Ceratomyxa sp.-1 spores in valvular view (PC: polar capsule; BS: binucleate sporoplasm; SL: suture line, IS: Immature spore)
Table 1. Site of infection, hosts, geographical localities, and dimensions (μm) of species of the genus Pseudalataspora and Ceratomyxa found in some marine fish (ND = not determined)SpeciesSpore BodyPolar capsuleSite of infectionHost speciesLocalityReferencesLengthThicknessLengthWidthPseudalataspora sp.-114.0(13.0–15.0)19.6(19.0–20.0)4.6(4.0–5.0)4.3(4.0–5.0)Gall bladder Merluccius merluccius Türkiye, Sea of MarmaraThis study Pseudalataspora vanderlingeni 14.2 ± 1.0(12.2–15.3)20.6 ± 1.8(18.2–23.4)5.0 ± 0.3(4.4–5.5)4.2 ± 0.3(4.8–0.3)Gall bladder Merluccius capensis South Africa[12]Ceratomyxa sp.-16.7(6.0–7.0)25.0(22.0–28.0)2.0(1.9–2.0)1.2(1.0–1.5)Gall bladder Merluccius merluccius Türkiye, Sea of MarmaraThis study Ceratomyxa globulifera 10.050.0NDNDGall bladder Merluccius vulgaris France, Mediterranean Sea[17] C. globulifera 8.0–10.052.0–58.04.0–5.0NDGall bladder Ophidion rochei Russia, Black Sea[34] C. scophthalmi 9.4(8.5–10.0)49.6(44.5–55.0)3.1(2.7–3.5)3.2(2.7–3.5)Gall bladder Scophthalmius maeoticus Türkiye, Black Sea[35] C. elegans 6.0–9.120.0–35.52.4–2.52.2–2.4Gall bladder Scorpaena porcus Ukraine, Black Sea[36] C. hepseti 6.0–8.010.0–15.06.0–8.0Gall bladder Atherina hepsetus Ukraine, Black Sea[37] C. inaequalis 5.0(5.0–6.0)28.0(28.0–31.0)2.5–3.5NDGall bladderSypmhodus spp.Ukraine, Black Sea[38] C. markewichi 4.8–6.016.0–20.42.0–2.51.3–1.8Gall bladder Trachurus mediterraneus Ukraine, Black Sea[39] C. merlangi 5.5(5.0–5.8)32.2(27.6–34.8)2.7(2.4–2.9)2.2(1.9–2.3)Gall bladder Merlangius merlangus Türkiye, Black Sea[40] C. peculiaria 6.5–8.521.0–29.32.4–2.71.9–2.4Gall bladder Spicara flexuosa Ukraine, Black Sea[41] C. allantoidea 6.0–6.629.3–38.62.0–2.71.0–1.5Gall bladder Ammodytes tobianus Bay of Biscay[42] C. leatherjacketi 5.8–6.524.5–31.02.5–3.0Gall bladder Aluterus monoceros West coast of Malaysia[43]
The second myxosporean species observed in this study was assigned to the genus Ceratomyxa based on its characteristic features, including two-valved, arcuate spores. Additionally, a distinctive symmetry was detected in the valves tapering to rounded tips. Moreover, sporoplasm binucleate (BS), polar capsules (PC) were identified as slightly pear-shaped toward the ends and located centrally within the spore (Fig. 1b). Developing immature spores (IS), often ‘crumpled’ in shape were observed (Fig. 1b). The measurements from 30 spore individuals revealed the following results: spore length was 6.7 μm (6.0–7.0 μm), spore thickness was 25.0 μm (22.0–28.0 μm), polar capsule length was 2.0 μm (1.9–2.0 μm), and polar capsule width was 1.2 μm (1.0–1.5 μm) (Table 1). Based solely on the significant similarities in spore morphology and morphometric measurements, all Ceratomyxa specimens in this study were classified as a distinct morphotype, Ceratomyxa sp.-1.
Phylogenetic Investigations
Specimens AO-93 and AO-95 were included in the molecular analyses as representatives of Pseudalataspora sp.-1. However, molecular analyses could not be conducted for Ceratomyxa sp.-1, because all specimens of this species were found mixed with Pseudalataspora sp.-1.
The 18S rDNA nucleotide sequences obtained from the selected Pseudalataspora sp.-1 specimen, AO-93 and AO-95, were approximately 1435 bp in length. The nucleotide sequence similarity between these two genotypes was 99.7%. The BLAST search identified both of these genotypes as the closest to Pseudalataspora vanderlingeni Reed, Kalavati, Mackenzie, Collins & Hemmingsen, 2018. Accordingly, a dataset comprising the related myxozoan species was established (see Online Resource). Phylogenetic analyses were performed on 1,165 aligned nucleotides, including 239 variable sites and 324 mutations. The GTR + I + G (I: 0.582; G: 0.617) and TIM3ef + I + G (I: 0.590; G: 0.642) evolutionary models were suggested by the AIC and BIC tests, respectively. The ML and NJ trees, derived from the initial model, were preferred because they had higher bootstrap support. The MP analysis, conducted using 162 synapomorphic sites, produced a single most parsimonious tree with 422 steps (CI: 0.767773; RI: 0.827768 and HI: 0.232227). In all phylogenetic trees constructed using the NJ, ML, and MP algorithms, specimens AO-93 and AO-95 appeared as sister taxa, with P. vanderlingeni (MF034897) as the closest binomial species to them. The nucleotide sequence identities of AO-93 and AO-95 with P. vanderlingeni were 99.5% and 99.7%, respectively. Additionally, the relationships within this lineage, comprising these three specimens, were supported by significant bootstrap values (62% ≤) in all phylogenetic trees (Fig. 2).
Fig. 2. The NJ tree, based on 18S rDNA nucleotide sequences of Pseudalataspora sp.-1 genotypes obtained in this study (AO-93 and AO-95) and closely related myxozoan species obtained from NCBI (GenBank accession numbers are indicated on the tree. Additional details are provided in the Online Resource). The tree is created using the GTR + I + G model, and Bootstrap values (≥ 50%) from NJ, ML, and MP analyses are displayed in the respective order at each relevant node. The tree is rooted with Ceratomyxa koieae and* C. milleri*
Parasitological Investigations
Of the 47 European hake (M. merluccius) samples investigated, 12 (25.5%) were infected with two myxosporean species. belonging to the genus Pseudalataspora Kovaljova & Gayevskaya, 1983 (Pseudoalataspora sp-1) and Ceratomyxa Thélohan, 1892 (Ceratomyxa sp-1) occurring either as sole species or co-infections in the bile of the gall bladder in the infected hosts.
Pseudoalataspora sp.-1 individuals were determined in all of the infected host fishes (n: 12), either as the sole parasite (n: 7) or co-occurring with Ceratomyxa sp.-1 spores (n: 5). The overall prevalence of infection and density of Pseudoalataspora sp.-1 were calculated as 25.5% and 3 + , respectively. Seasonal prevalence and infection density were also determined and are presented in Table 2. Ceratomyxa sp.-1, the second myxozoan species identified in this study, was found in 5 out of 47 host fishes, corresponding to a prevalence of 10.6%, together with an infection density of 3 + . Seasonal prevalence and density values of infection of Ceratomyxa sp.-1 were also determined and provided in Table 2. It is worth noting that Ceratomyxa sp.-1 was observed exclusively co-occurring with Pseudoalataspora sp.-1 rather than as a sole parasite species in the bile of host fishes.
Table 2. Overall and seasonal levels of infection prevalence (%) and density of Pseudalataspora vanderlingeni, and co-existing P. vanderlingeni + Ceratomyxa sp. in European hake, M. merluccius.N = number of fish examinedInfection indicesSeasonParasite Pseudalataspora vanderlingeni Co-infection(P. vanderlingeni + Ceratomyxa sp.)Prevalence (%)Winter (n = 17)29.411.8Summer (n = 30)23.310.0Overall (n = 47)25.510.6DensityWinter (n = 17)3 + 3 + Summer (n = 30)4 + 3 + Overall (n = 47)3 + 3 +
Discussion
This study presents valuable new information on the biodiversity and parasitological characteristics of Myxosporea infecting M. merluccius in the Sea of Marmara, Türkiye. Both microscopic and phylogenetic examinations revealed that two distinct spore types, belonging to the genera Pseudalataspora and Ceratomyxa, were present in the gallbladders of infected host fishes. Morphological and morphometric data, as well as phylogenetic analyses of the 18S rDNA gene with a high bootstrap value (100%) for Pseudalataspora sp.-1 spores, revealed that all samples belonged to the same species, Pseudalataspora vanderlingeni Reed, Kalavati, Mackenzie, Collins & Hemmingsen, 2018 (MF034897).
Morphology and Light Microscopy
When comparing the spore morphology and morphometric data of Pseudalataspora sp.-1 with the original description of P. vanderlingeni [12], it is evident that the two species are almost indistinguishable (see Table 1 for details), with only slight differences. The original description noted relatively small alate processes that joined at the proximal extremities of the valves; these were observed only in some developing spores in the present study. In the present study, the shape and dimensions of polar capsules were mostly round, with a uniform margin, whereas in the original spores, they were oval, with varying dimensions and margins. As a result, all evidence obtained from spore morphology, morphometry, and molecular phylogeny led us to assign Pseudalataspora sp.-1 spores to Pseudalataspora vanderlingeni. This species is relatively new, having been observed and identified in the gallbladder of the type host, the Cape hake, Merluccius capensis Castelnau, 1861, in South Africa. Additionally, M. paradoxus Franca, 1960, from the west coast of Africa, has been reported as another host of this parasite [12]. Since it’s the first identification, there have been no subsequent records of this parasite. Therefore, this study presents the second global record of P. vanderlingeni. So far, several myxozoan genera, including Myxobolus, Henneguya, Ortholinea, Kudoa, Ceratomyxa, and Myxidium, have been reported in various fish species along the Turkish coasts [23, 35, 40, 44–54]. However, although a single species, Pseudalataspora pontica Kovaljova, Donec & Kolesnikova, 1989 (ext. from Chelon aurata), has been identified in the northern Black Sea, there has been no previous record of infection by members of the genus Pseudalataspora in Türkiye. Therefore, this study provides the first record of both the species P. vanderlingeni and the genus Pseudalataspora in Turkish waters.
The second myxozoan genus identified in this study was Ceratomyxa, which was observed only in co-infection with P. vanderlingeni spores. The defining characteristics of the genus are crescent-shaped or arcuate spores and elongated shell valves that are generally thicker than they are long [55]. Homogeneity in morphometric measurements, coupled with the presence of characteristically arcuate and equal spore valves, indicated that all Ceratomyxa specimens in this study belong to the same morphotype: Ceratomyxa sp.-1. However, molecular data could not be obtained from these specimens to enable a precise species identification. To date, six binomial species of the Ceratomyxa and two unspecified specimens of the genus have been recorded from marine fishes in the Turkish seas. These include: Ceratomyxa sp. (extracted from Dicentrarchus labrax), Ceratomyxa beloneae (ex. from Belone belone), C. merlangi (ex. from Merlangius merlangus), Ceratomyxa sp. (ex. from Scophthalmus maeoticus), C. scorpaeni (ex. from Scorpaena porcus), C. scophthalmi (ex. from S. maeoticus), C. diplodae and C. labracis (ex. from D. labrax) [35, 48, 50–54]. Although all these species exhibited equal-valved spores, Ceratomyxa sp.-1 exhibited morphometric characteristics that did not correspond exactly to any previously recorded species in the Black Sea or other regions (see Table 1 for details), suggesting that it is likely a new taxon from Turkish waters. Additionally, when compared to other morphologically similar and equally valved Ceratomyxa species distributed worldwide (as reported by Eiras [56] and Eiras et al. [57], as well as by several authors in the Black Sea (see Table 1 for details), some species exhibited mostly overlapping spore and polar capsule dimensions, but different spore morphology. These species include C. merlangi, which has crescent-shaped spores with blunt, rounded ends very closely located polar capsules to each other; C. peculiaria, which has valves that are totally round-ended with an anterior margin that is slightly convex and a posterior margin that is straight or slightly convex; and C. leatherjacketi, which has spores that are crescent-shaped and transversely elongated with a posterior angle that is slightly concave, as well as valves that are equal in size or one that is slightly elongated, and spherical polar capsules. Some of the "crumpled" spores observed in the present study were also previously reported for C. scophthalmi [35]. It must be noted that the traditional taxonomic classification based on spore and polar capsule morphology may not be sufficient for a proper species identification because several myxosporeans with very similar spore morphology assigned to the same genera have been located in phylogenetically distantly related taxa. Given the lack of genetic data in the present study and the morphological differences among the above-mentioned species, our Ceratomyxa sp.-1 remains a distinct morphotype of a likely new taxon based on current data; however, further studies will be required to confirm the exact species identification.
Phylogenetic Investigations
The genus Pseudalataspora comprises 14 species; however, only two, P. kovalevae and P. vanderlingeni, have available genetic data. Phylogenetically, members of this genus have been found in different lineages within the Ceratomyxa clade, forming a paraphyletic group [12, 58, 59]. Therefore, the boundaries of genetic variation established within and between Ceratomyxa species are also applicable to Pseudoalataspora species. A low intraspecific 18S rDNA variation has been reported for the genus Ceratomyxa. For instance, this value was 0.15% (99.85% similarity) in C. nolani and 0.36% (99.64% similarity) in C. lunula [55]. Additionally, the genus has been characterised by high interspecific divergence in the 18S rDNA gene. Reported genetic similarities among closely related species include 98.5% between C. sultani and C. arabica, 97.6% between C. gunterae and C. dennisi, and 97.6% between C. robertsthomsoni and C. thalassomae [35, 55]. From this perspective, the high nucleotide sequence similarity (99.7% and 99.5%) between Pseudalataspora sp.-1 (AO-93 and AO-95) and P. vanderlingeni specimens supported the conclusion that these two taxa belonged to the same species.
Parasitological Investigations
All species of the genus Pseudalataspora, including P. vanderlingeni, are coelozoic parasites that have been found in the gall bladders of marine fishes [12]. In line with the original description of P. vanderlingeni, all specimens examined in this study were observed in the gall bladder of the host fish, M. merluccius. Consequently, the P. vanderlingeni spores examined in this study occupy the same ecological niche as those originally described. Similarly, the genus Ceratomyxa comprises more than 270 species that predominantly inhabit the gallbladder [57]. Consistent with this background, all Ceratomyxa sp.-1 specimens in this study were observed in the gall bladder of infected M. merluccius individuals.
The prevalence of P. vanderlingeni infections in the two originally reported host species, M. capensis and M. paradoxus, collected from the Southern Benguela off the West Coast of South Africa, was 100%. By contrast, the prevalence was 61.5% in M. capensis in the southern Benguela region [12]. The overall prevalence of infection in this study (25.5%) is lower than that reported in the aforementioned studies, but higher than the prevalence of P. misrae (3.8%) in Rastrelliger kanagurta [60] and within the range reported for P. lophii in Lophius piscatorius [35] and P. beryxi in Beryx splendens [61]. The overall prevalence of co-occurring P. vanderlingeni and Ceratomyxa sp. infections in the present study was 10.6%, which is double the reported prevalence (4.76%) of co-occurring P. lophii and C. lophii infections in L. piscatorius [35]. Despite limited data on infections involving members of the genus Pseudalataspora, there are more reports of single and co-infections involving members of the genus Ceratomyxa in fish hosts, with overall prevalence rates of up to 100% and marked seasonal variation [35, 53, 57, 62–65]. Myxosporean co-infections have been linked to biological, geographical, and environmental factors, as well as to similar life cycle strategies [53, 66]. Our results revealed very similar overall and seasonal (winter and summer) infection prevalence values for P. vanderlingeni and its co-infections with Ceratomyxa sp.-1. A two-stage life cycle strategy involving a myxosporean stage in fish and an actinosporean stage in invertebrate alternate hosts in freshwater environments, and fish-to-fish transmission in marine myxosporean parasites, has been reported [67–73]. Alama-Bermejo et al. [63] reported the presence of the invasive blood stage of C. puntazzo in sharpsnout seabream (Diplodus puntazzo) year-round, as determined by PCR. Based on their unsuccessful experimental transmission of different C. puntazzo developmental stages in seawater or via oral and intracoelomic injection, they hypothesized that an invertebrate host was involved in life-cycle dynamics rather than fish-to-fish transmission in aquaculture systems. Currently, no reports indicate how infections with P. vanderlingeni and Ceratomyxa sp.-1 progress. Based on the aforementioned reports on the life-cycle strategies of marine myxosporean parasites and the infection levels of the two parasite species identified in our study, it can be inferred that they likely share the same life-cycle strategy: either an alternate or a direct fish-to-fish transmission. According to Rocha et al. [74], temperature is the primary factor influencing the life cycle of myxosporean parasites, facilitating their successful development alongside specific physical, chemical, and biological conditions. The Benguela system, the locality where the original description of P. vanderlingeni was reported, is one of the four major eastern boundary upwellings of the World Ocean and is unique because its warm water currents with enhanced plankton production, which are requested by pelagic and demersal resources, including fish, for food [75, 76]. On the other hand, the biodiversity in the Sea of Marmara, where this study was conducted, is threatened by low dissolved oxygen levels, increasing eutrophication caused by human impacts, and marine litter on the seafloor.[77]. The differences in the overall infection prevalence values of P. vanderlingeni in the original description (61,5–100%) and in the present study (25.5%) could have resulted from these different characteristics of both sampling localities. Alama-Bermejo et al. [63] also reported temperature-related seasonal C. puntazzo infections in fish, coinciding with temperature values, and indicated that temperature appeared to regulate the density of actinospores of C. puntazzi in the environment in accordance with their hypothesized invertebrate–fish host life cycle strategy. Similar seasonal patterns observed for both parasite species in the present study may also be due to these factors being available year-round to the parasites or their alternate hosts.
Various myxozoan genera, including Kudoa, Myxoproteus, Fabespora, Pseudalataspora, and Ceratomyxa, have been reported to infect Merluccius species, some of which have either a widespread or a restricted global distribution. Of these, C. globulifera has only ever been found in M. merluccius off the Mediterranean coast of France, and P. vanderlingeni has only ever been found once in M. capensis and M. paradoxus along the west and south coasts of South Africa. However, the results of this study suggest that P. vanderlingeni and Ceratomyxa have a broader distribution, extending into the eastern Mediterranean Sea and neighbouring seas. This indicates that P. vanderlingeni could be a widespread parasite of Merluccius species, affecting populations not only off the African coast, but also in the Mediterranean Sea. Similarly, Ceratomyxa appears to be prevalent in Merluccius populations throughout the Mediterranean Sea. Nevertheless, further studies on myxosporean occurrences in a wider range of host species and geographical locations are required to confirm these presumptions.
The main findings of this study can be summarised as follows; (a) Members of two myxozoan taxa (P. vanderlingeni and Ceratomyxa sp.-1) were identified in M. merluccius caught in the Sea of Marmara in Türkiye; (b) This study presents the second global record of P. vanderlingeni since its original description, and the first record of the genus Pseudalataspora and P. vanderlingeni in Turkish waters; (c) Based on unique spore morphology, this study presents a likely new Ceratomyxa taxon in Turkish waters; (d) The results of this study, in conjunction with those of Reed et al*.* [12], suggest that P. vanderlingeni is a common myxozoan parasite of Merluccius species along the Southern Atlantic coasts of Africa, extending into the Mediterranean and surrounding seas; (e) Similarly, in conjunction with the results of Desportes and Theodorides [17], this study presents Ceratomyxa is one of the common myxozoan parasite of Merluccius species in the Mediterranean and adjacent waters.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Casey J, Pereiro J (1995) European hake (M. merluccius) in the North-east Atlantic. In: Alheit J, Pitcher TJ (eds) Hake; fisheries, ecology and markets. Fish and Fisheries Series, Vol. 15. Chapman and Hall, London, UK, pp. 125–147. 10.1007/978-94-011-1300-7
- 2Odabaş-Alver D, Ertürk Ö, Gürkanlı CT (2024) Biodiversity and parasitological characteristics of Myxozoa (Cnidaria) infecting European seabass, Dicentrarchus labrax (Linnaeus, 1758) in the Aegean Sea coasts of Türkiye. Trj FAS 24 TRJFAS 25882. 10.4194/TRJFAS 25882
- 3Özer A, Öztürk T (2011) First report of Ceratomyxa sp.(Myxozoa) from Dicentrarchus labrax in the Aegean Sea and Caligus minimus (Copepoda) in the Black Sea. In: 16th National Fisheries Symposium. pp 25–27
- 4Yurakhno VM, Güneydağ S, Özer A (2017) New species of myxosporeans of genus Ceratomyxa (Myxosporea: Ceratomyxidae) from gall bladder of the Black sea turbot Scophthalmus maeoticus. Ecologicheskaya, promishlennaya i energeticheskaya bezopasnost’ – 2017: sbornik statey po materialam nauchno-prakticheskoy conferentsii s mezhdunarodnim uchastiyem, 11 – 15 September 2017. In: Omelchuk YA, Lyamina NV, Kucheric GV (eds). Sevastopol. pp 1592–1595 (in Russian).