Salt stress responses and tolerance mechanisms in native desert plants of the UAE: growth, biochemical, and antioxidant perspectives
Sahara Abo Amin, Faisal Hayat, Mohammed Alyafei

TL;DR
This study examines how four native UAE desert plants respond to high soil salinity, identifying which species are most resilient and the mechanisms they use to survive.
Contribution
The study provides field-based insights into salinity tolerance mechanisms of native UAE desert shrubs under realistic conditions.
Findings
Salvadora persica and Lycium shawii showed the highest salinity tolerance with strong osmotic adjustment and antioxidant activity.
Calligonum comosum had limited high-salinity tolerance, with reduced survival and photosynthetic performance.
Ion regulation and oxidative protection were key factors in salinity tolerance across species.
Abstract
Soil salinization threatens productivity and ecosystem stability in arid regions, yet salinity-tolerance mechanisms of native UAE desert shrubs remain poorly resolved under field-like conditions. This study aimed to identify tolerance strategies and rank salinity resilience among Lycium shawii, Salvadora persica, Calligonum comosum, and Haloxylon salicornicum. Plants were grown in an outdoor pot trial under ambient desert conditions and irrigated with three salinity levels (ECw = 1.5, 10, and 25 dS m-¹). Severe salinity (25 dS m-¹) revealed clear interspecific divergence: S. persica and L. shawii maintained 100% survival, relatively stable water status and photosynthetic performance, and reduced membrane injury and lipid peroxidation. Their tolerance was associated with stronger osmotic adjustment (higher proline and soluble sugars), improved ion homeostasis (lower shoot Na+…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1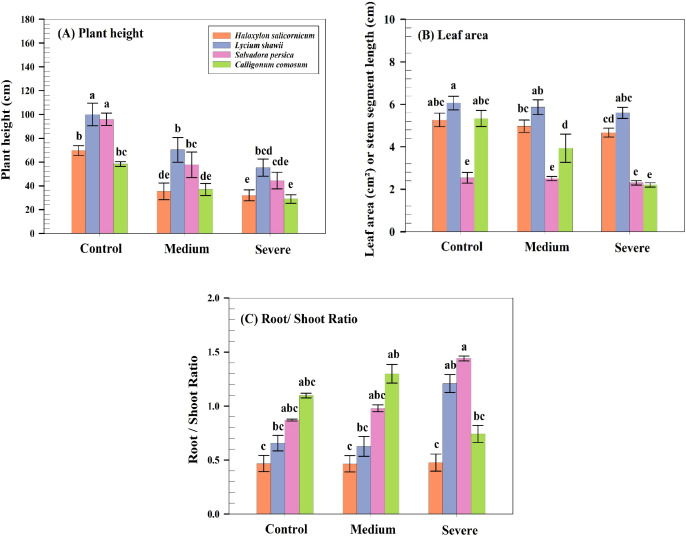 Figure 2
Figure 2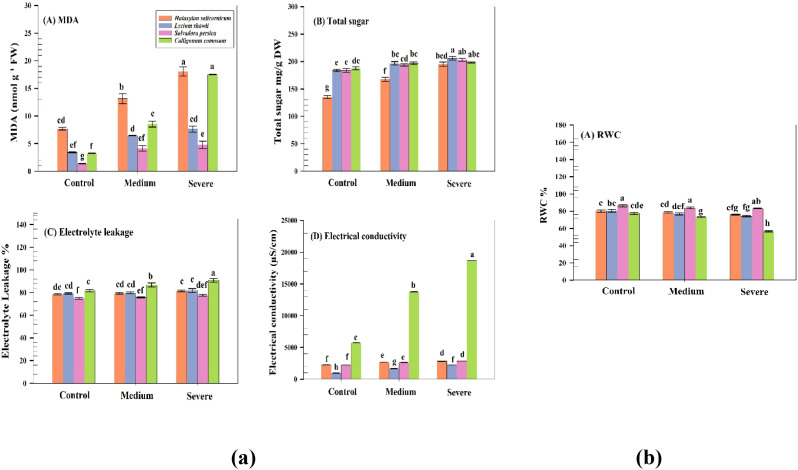 Figure 3
Figure 3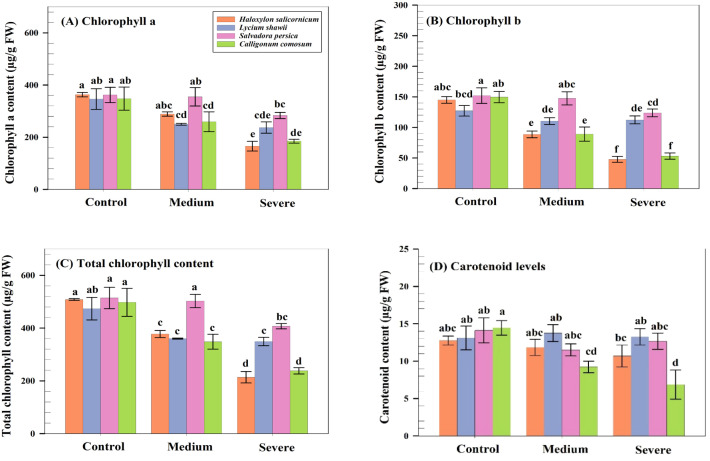 Figure 4
Figure 4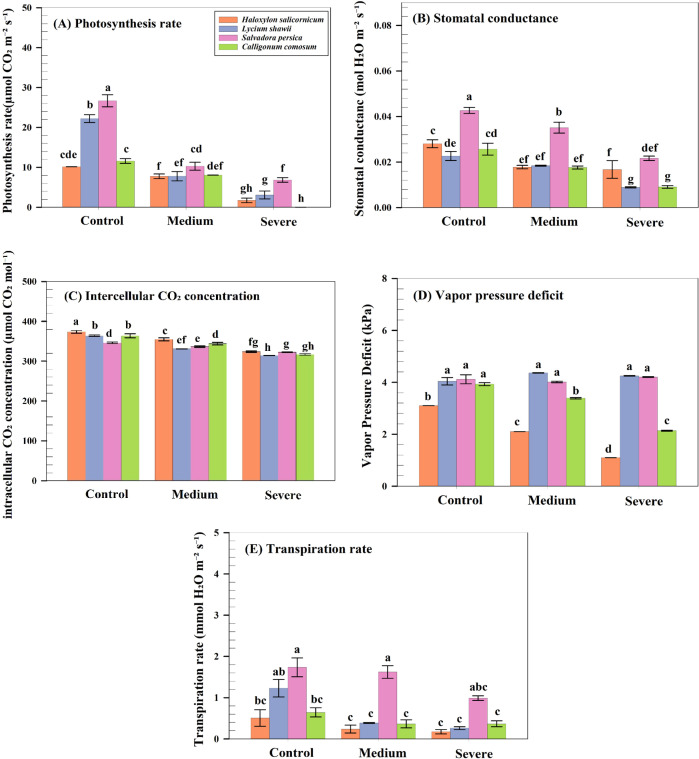 Figure 5
Figure 5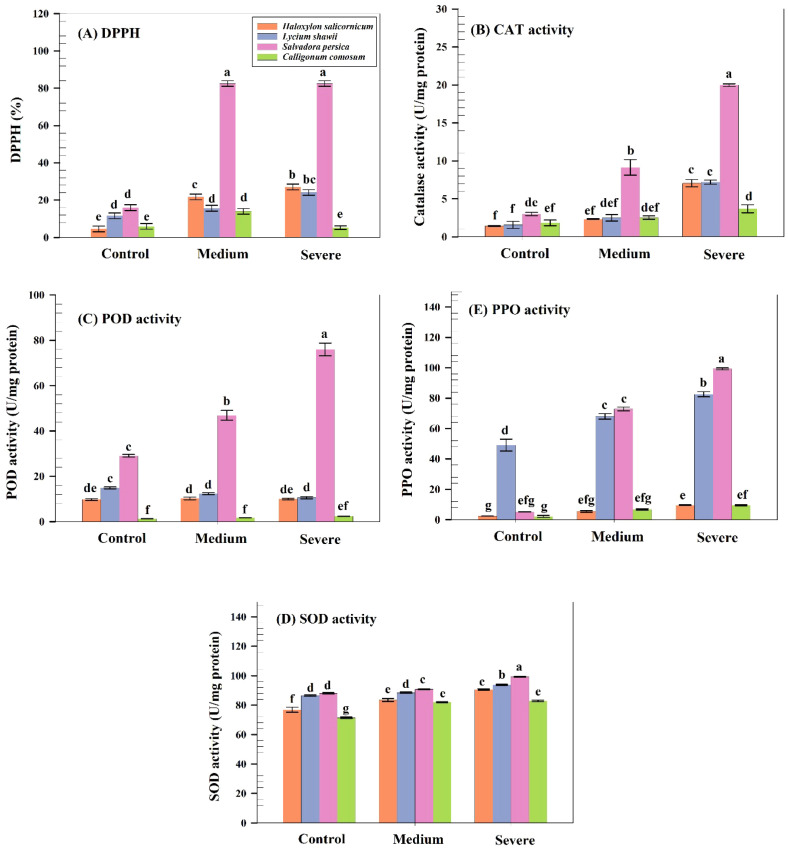 Figure 6
Figure 6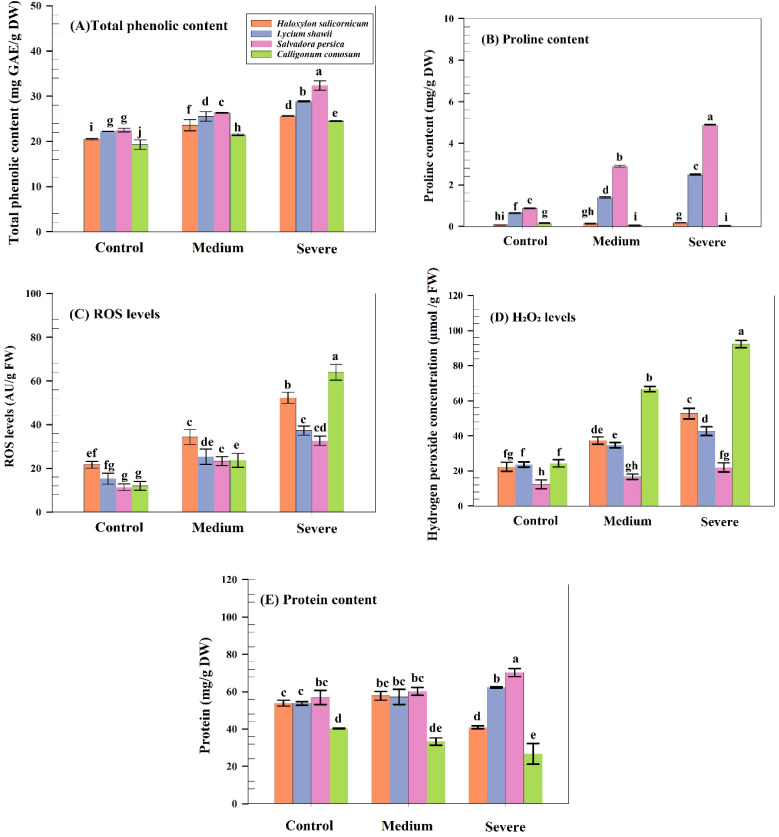 Figure 7
Figure 7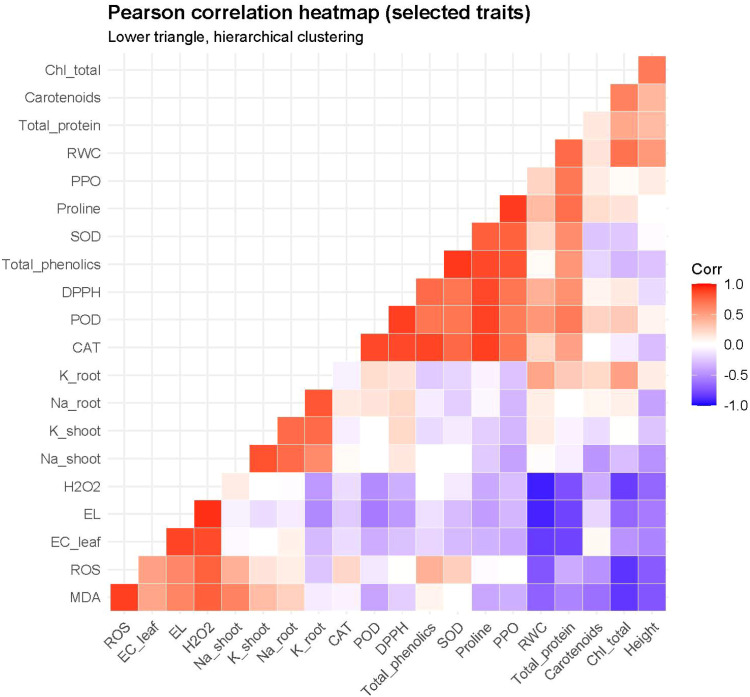 Figure 8
Figure 8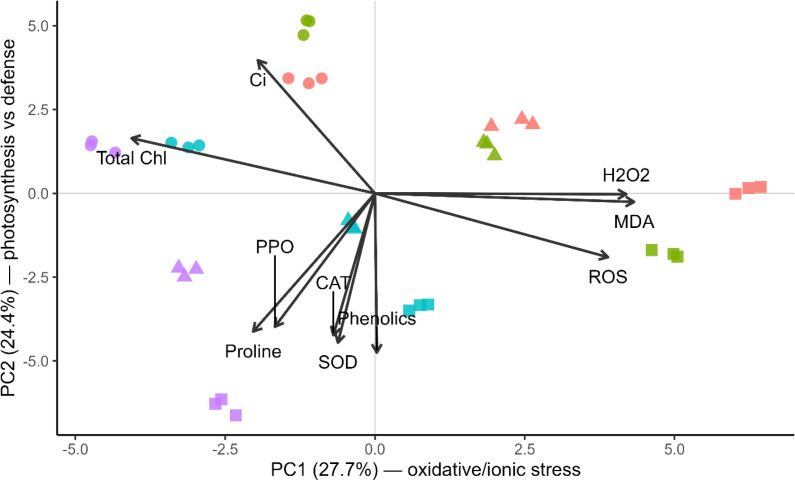 Figure 9
Figure 9| Species | Habitat and ecological role | Physiological traits supporting salinity adaptation | Known uses | References |
|---|---|---|---|---|
|
| Rocky deserts and saline wadis | Strong osmotic adjustment, efficient antioxidant system, high membrane stability | Antidiabetic, anti-inflammatory, forage, saline land rehabilitation |
|
|
| Coastal deserts and saline soils | High salt exclusion capacity, strong antioxidant defenses | Miswak (oral hygiene), antimicrobial, fodder, saline soil phytostabilization |
|
|
| Sandy and gravel plains with moderate salinity | Moderate tolerance, weak antioxidant response, early growth decline | Forage, sand fixation, traditional medicine |
|
|
| Highly saline arid zones and sabkha habitats | Moderate membrane stability, tolerates ionic stress | Fuelwood, rangeland rehabilitation, xerophytic landscaping |
|
| Parameter | Value | Method | Reference |
|---|---|---|---|
| Soil texture | Sandy (90% sand, 6% silt, 4% clay) | Hydrometer method |
|
| Water-holding capacity | Low | Visual/drainage observation |
|
| pH | 8.0 | pH meter |
|
| Electrical conductivity (ECe) | 1.1 dS m-¹ | EC meter |
|
| Organic matter content | <0.5% | Walkley-Black method |
|
| Nitrogen (mg kg-¹) | 28 | Kjeldahl digestion | |
| Phosphorus (mg kg-¹) | 7.4 | Olsen method |
|
| Potassium (mg kg-¹) | 93 | Flame photometry |
|
| Species × treatment × organ | Na | K | P | Fe | Zn | Mn |
|---|---|---|---|---|---|---|
| 2987.3 l | 4655.6 j | 1313.5 l | 1841.1 a | 64.5 g | 64.9 d | |
| 5307.2 k | 5461.2 h | 1480.8 j | 638.5 c | 108.3 c | 28.5 g | |
| 7987.7 j | 5186.3 i | 1584.1 k | 705.6 g | 93.9 b | 38.4 h | |
| 7225.7 j | 3225.9 k | 1400.6 k | 1379.7 a | 93.1 e | 77.4 b | |
| 6119.6 k | 3437.6 l | 2007.1 i | 926.7 e | 98.8 a | 36.1 j | |
| 7752.8 i | 3371 j | 2292.6 e | 739.7 b | 155.2 a | 33.9 f | |
| 12000.1 i | 10581.8 d | 2157.2 h | 440.1 j | 90.7 c | 24.7 k | |
| 14835.6 f | 6133.3 g | 2383.9 d | 97.8 j | 81.6 f | 23.9 j | |
| 21284.4 c | 12765.5 b | 2396.7 g | 212.2 l | 62.5 i | 23.1 l | |
| 22883.5 d | 15893.2 a | 3066 b | 93.3 k | 45.1 k | 17.1 k | |
| 19919.3 d | 7195.4 e | 1819.1 j | 1000.2 d | 58.5 j | 44 g | |
| 14174 g | 3822.6 i | 1654.4 i | 167.4 i | 50.1 j | 24.6 i | |
| 31479.3 a | 17553.3 a | 3416.7 b | 1561.7 b | 89.2 d | 158.2 a | |
| 26946.5 c | 13115.9 b | 2524.3 c | 280.5 e | 125.5 b | 68.7 d | |
| 24225.7 b | 12121.9 c | 3137.1 d | 1134.2 c | 62.85 h | 132.7 b | |
| 32139.9 b | 12551.4 c | 2099.9 g | 234.5 g | 101.4 d | 76.7 c | |
| 16972 f | 6690.5 f | 2857.5 e | 706.7 f | 36.5 l | 107.2 c | |
| 37333.2 a | 11986.8 d | 1675.5 h | 188.5 h | 77.2 i | 84.7 a | |
| 12582 h | 5328.5 h | 2598.5 f | 500.8 h | 75.5 f | 54.3 e | |
| 7759.1 i | 3361 j | 2297.4 e | 738.7 b | 154.2 a | 33.7 f | |
| 18413.4 e | 6024.3 g | 4562.5 a | 391.8 k | 84.9 e | 38.1 i | |
| 13960.2 h | 8343.9 e | 3278.3 a | 327.8 d | 77.9 h | 28.2 h | |
| 15467.4 g | 4548.2 k | 3156.4 c | 467.5 i | 56.4 k | 45.6 f | |
| 17450.2 e | 7551.5 f | 2240.3 f | 245.1 f | 78 g | 41 h |
| Species | Survival control (%) | Survival 10 dS m-¹ (%) | Survival 25 dS m-¹ (%) | Salt-Injury index control | Salt-Injury index 10 dS m-¹ | Salt-Injury index 25 dS m-¹ |
|---|---|---|---|---|---|---|
|
| 100 | 100 | 100 | 1 | 1 | 1.5 |
|
| 100 | 100 | 100 | 1 | 1 | 1.2 |
|
| 100 | 100 | 100 | 1 | 2 | 3 |
|
| 100 | 96.7 | 83.3 | 1 | 2.5 | 4 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Aluminum toxicity and tolerance in plants and animals · Silicon Effects in Agriculture
Introduction
1
Soil salinization is one of the most severe constraints to agricultural productivity, ecosystem stability, and sustainable development in arid and semi-arid regions. It threatens several United Nations Sustainable Development Goals (SDGs), particularly Zero Hunger (SDG 2), Clean Water and Sanitation (SDG 6), and Life on Land (SDG 15). Globally, more than one billion hectares of land are affected by salinity across more than 100 countries, and the extent of salt-affected soils continues to expand due to climate change, rising temperatures, seawater intrusion, and unsustainable irrigation practices. Beyond agriculture, soil salinization degrades biodiversity, alters ecosystem services, and limits the success of land rehabilitation and restoration programs (Singh, 2022). Salinity imposes two major stresses on plants: osmotic stress, which reduces water uptake and turgor, and ionic stress caused by excessive accumulation of Na^+^ and Cl^-^ in plant tissues. These stresses disrupt water relations, decrease stomatal conductance, impair CO_2_ fixation, and ultimately suppress photosynthetic efficiency and growth. Reduced carbon assimilation under salinity also enhances the generation of reactive oxygen species (ROS). Excess ROS can damage lipids, proteins, and DNA, leading to membrane instability, metabolic dysfunction, and cellular injury (Balasubramaniam et al., 2023).
To mitigate oxidative damage, plants activate complex defense systems that include enzymatic antioxidants such as superoxide dismutase (SOD), catalase (CAT), and peroxidases, as well as non-enzymatic antioxidants including carotenoids, phenolics, and other secondary metabolites. These systems work together with osmotic adjustment mechanisms (proline and soluble sugars accumulation) and ion regulation strategies (Na^+^ sequestration, K^+^ retention, and vacuolar compartmentalization) to maintain cellular homeostasis under salt stress (Abdulfatah, 2022; Mansoor et al., 2022; Kesawat et al., 2023; Sadiq, 2023; Bandodkar et al., 2025).
Plant species exhibit strikingly different responses to salinity. Glycophytic crops such as rice and wheat are highly salt-sensitive, whereas crops like barley and tomato show moderate tolerance. Tree crops, including olive and date palm, can tolerate salinity to some extent but still suffer yield reductions under high salt conditions (Sarkar and Sadhukhan, 2023; El Yamani and Cordovilla, 2024). In contrast, halophytes, including genera such as Salicornia, Suaeda, and Atriplex have evolved specialized adaptations that enable survival in highly saline environments. These include efficient Na^+^ sequestration in vacuoles, salt gland excretion, succulence, osmolyte accumulation, and enhanced antioxidant defenses. Consequently, halophytes are increasingly explored for biosaline agriculture, phytoremediation, and ecological restoration of degraded saline lands (Calone, 2022; Mehra et al., 2024; Soni et al., 2024). However, most mechanistic studies on salinity tolerance have focused on model species or a limited number of well-known halophytes, often under controlled greenhouse conditions. Moreover, greenhouse-based studies often overestimate salt tolerance because they exclude combined environmental constraints such as high radiation, temperature fluctuations, and evaporative demand, which strongly modify ion uptake, water relations, and oxidative stress responses in desert ecosystems. Therefore, tolerance rankings under controlled environments may not reflect plant performance under realistic arid-land conditions. As a result, there remains a critical knowledge gap regarding how native desert shrubs tolerate salinity under realistic environmental conditions.
In the United Arab Emirates (UAE), soil and groundwater salinity are intensifying due to seawater intrusion, over-extraction of groundwater, high evaporation rates, and the use of marginal-quality irrigation water. In several northern regions, groundwater salinity has been reported to reach up to 25 dS m^-^¹, posing serious risks to agriculture and natural ecosystems. Rising salinity has even affected traditionally salt-tolerant species such as date palms, with documented cases of dieback linked to salt accumulation (Elmahdy and Mohamed et al., 2023; Hammami et al., 2024). Field-based research in the UAE, including mangrove and halophyte trials irrigated with saline water, suggests that native shrubs can tolerate high ionic loads and may contribute to soil stabilization and salinity mitigation. Recent initiatives by the International Centre for Biosaline Agriculture (ICBA) and the Environment Agency- Abu Dhabi (EAD) have focused on screening native halophytes and implementing salinity-resilient revegetation programs across coastal and inland saline zones (Lyra et al., 2022; Choudhary et al., 2023; Friis and Killilea, 2023; Rhee et al., 2025). Within this context, four native UAE shrubs, Salvadora persica, Lycium shawii, Calligonum comosum, and Haloxylon salicornicum are of particular interest due to their ecological significance, traditional uses, and presumed salinity tolerance (Table 1). These species are well adapted to desert conditions and play important roles in habitat stabilization, biodiversity support, and potential biosaline agriculture (Brown and Feulner, 2023).
Despite growing interest in halophytes for biosaline agriculture and land rehabilitation, integrated mechanistic evidence under field-like desert conditions remains limited. Most existing studies either focus on model halophytes, single physiological traits, or greenhouse experiments, which do not fully capture plant responses under realistic environmental conditions (Todorović et al., 2022; Shahid and Alkandari, 2024). Consequently, there is insufficient knowledge to guide evidence-based species selection for large-scale saline land restoration in the UAE and similar arid regions. In particular, comparative multi-trait evaluations linking water status, ion regulation, and oxidative damage under field-like desert conditions remain largely unavailable for native Arabian shrubs. To address this gap, we tested the hypothesis that salt-tolerant native shrubs maintain higher water status and photosynthetic performance through coordinated ion regulation, osmotic adjustment, and antioxidant protection under realistic desert conditions. This study evaluates growth, water relations, ion regulation, photosynthetic traits, osmolytes, and oxidative stress responses of L. shawii, S. persica, H. salicornicum, and C. comosum under three irrigation salinity levels (ECw = 1.5, 10, and 25 dS m^-^¹) to identify species-specific tolerance strategies and guide saline-land rehabilitation in arid environments.
Materials and methods
2
Plant material and growth conditions
2.1
The experiment was conducted as an outdoor pot experiment under ambient desert conditions at Al Fo’ah Farm, United Arab Emirates University, in Al Ain, from October 2023 to May 2024. The site experiences a typical desert climate, with average daytime temperatures ranging from 28 to 35 °C and relative humidity between 30% and 50%. Uniform seedlings of four native desert species, including L. shawii, S. persica, H. salicornicum, and C. comosum, were obtained from Al Salamat Nursery, Al Ain. The soil at the site that used in the pots was classified as sandy (90% sand, 6% silt, 4% clay), with poor water-holding capacity and rapid drainage, typical of arid zones. The physicochemical properties of the experimental soil are summarized in Table 2. A completely randomized design (CRD) was implemented. Each species (15 plants per species) was assigned randomly to one of three irrigation salinity treatments: a control (EC_w_ = 1.5 dS m^-^¹), a moderate salinity level (EC_w_ = 10 dS m^-^¹), and a high salinity level (EC_w_ = 25 dS m^-^¹), with five biological replicates per treatment, where ECw refers to the electrical conductivity of the irrigation water. Salinity treatments were prepared by dissolving NaCl in the irrigation water as a standard salt source to impose controlled ionic stress and standardize treatments. Salinity was increased gradually over two weeks to minimize osmotic shock. Soil salinity was monitored weekly from the soil saturation extract (ECe) using a portable conductivity meter. Plants were grown individually in pots (50 cm height × 40 cm diameter) and arranged with sufficient spacing (≥1 m apart) to ensure uniform light penetration and to minimize edge effects. During the greenhouse stage (Acclimatation stage; August to September 2023), plants were grown under natural sunlight supplemented with LED lights (200 µmol/m²/s) to maintain a photoperiod of 14 h light/10 h dark at 28 ± 2 °C. In the field stage (Salinity stress stage; October 2023 to May 2024), light intensity ranged from 1200 to 2000 µmol/m²/s during daylight hours, with an average natural photoperiod of 11 to 13 hours across the winter–spring season.
Observations of morphological traits
2.2
Morphological and growth parameters were recorded under control and salinity stress treatments, including plant height, leaf area (or segment length for leafless species), root to shoot ratio and survival rate. Plant height was measured using a meter scale. For species with leaves, leaf area was determined from five representative plants per treatment using the manual tracing method on graph paper. For species without leaves, the mean stem segment length was recorded. Fresh weight was measured immediately after harvest, and samples were then wrapped in aluminum foil and oven-dried at 80 °C for 48 hours to determine dry weight. Plants were separated into shoots and roots before weighing, and root-to-shoot ratio was calculated as root dry weight divided by shoot dry weight. Survival rate was calculated as the percentage of healthy plants remaining in each treatment at the end of the experiment. In addition, a salt injury index was scored visually on a 1–5 scale (1 = healthy plants with no necrosis; 2 = 1–25% necrotic leaf area; 3 = 26–50%; 4 = 51–75%; 5 = 76–100% necrotic area or plant death), following Zhen et al (Zhen et al., 2010).
Photosynthetic pigments and gas exchange parameters
2.3
Total chlorophyll, chlorophyll a, chlorophyll b, and carotenoid contents were estimated from fully expanded leaves using 85% acetone extraction following Palta (Palta, 1990). Absorbance was recorded at 663, 645, and 440 nm using a spectrophotometer (Helios Alpha, Thermo Scientific), and pigment concentrations were calculated using the MacKinney equations (Marker, 1972), and values were expressed on a fresh weight basis (µg g^-^¹ FW).
Gas exchange parameters, including net photosynthetic rate (A), stomatal conductance (gs), transpiration rate (E), and vapor pressure deficit (VPD_leaf_), were measured using a portable infrared gas analyzer (LI-6400XT Portable Photosynthesis System., USA) on the first fully expanded leaf of each plant. Measurements were taken under ambient light and temperature conditions, with three biological replicates per treatment.
Relative water content, membrane stability, and lipid peroxidation
2.4
Leaf relative water content (RWC) was determined following Mullan and Pietragalla (Mullan and Pietragalla, 2012) using five leaves per treatment. Fresh weight (FW), turgid weight (TW), and dry weight (DW) were measured, and RWC was calculated using Equation 1.
Membrane stability was assessed by measuring electrolyte leakage (EL) following Dionisio-Sese and Tobita (Dionisio-Sese and Tobita, 1998). Fresh leaves (0.2 g) were incubated in deionized water for 2 h at 32 °C to record initial conductivity (EC1), then autoclaved and cooled to measure final conductivity (EC2). EL (%) was calculated using Equation 2.
Lipid peroxidation was measured by estimating malondialdehyde (MDA) content following the thiobarbituric acid (TBA) method (Schmedes and Hølmer, 1989). Absorbance was recorded at 532 and 600 nm, and MDA was calculated using an extinction coefficient of 155 mM^-^¹ cm^-^¹ and expressed as nmol/g fresh weight.
Biochemical parameters and antioxidant activities
2.5
Antioxidant enzyme activities, including catalase (CAT), peroxidase (POD), polyphenol oxidase (PPO), and superoxide dismutase (SOD), were analyzed from leaf extracts using standard spectrophotometric methods described by Beers and Sizer (Beers and Sizer, 1952), Chance and Maehly (Chance and Maehly, 1955), Mayer et al (Mayer et al., 1966), and Beauchamp and Fridovich (Beauchamp and Fridovich, 1971), respectively. Triplicate samples were homogenized in 0.1 M sodium phosphate buffer (pH 7.0) and centrifuged at 12,000×g for 15 minutes at 4 °C and absorbance was measured at appropriate wavelengths for each assay. Enzyme activities were expressed on a protein basis (U/mg protein). DPPH (radical scavenging activity) was determined using the method of Blois (Blois, 1958) with minor modifications. Absorbance was recorded at 517 nm, and the percentage of radical scavenging activity was calculated relative to a methanol blank. Total soluble proteins were estimated using the Bradford method (Bradford, 1976), with bovine serum albumin as the standard.
Ionic content and soluble sugars
2.6
Ionic concentrations were determined in leaf, stem, and root tissues using Inductively Coupled Plasma Mass Spectrometry (ICP-MS). Oven-dried samples (0.5 g) were digested with a 9:4 mixture of nitric and perchloric acids using a microwave digestion system (CEM-MARS 6), diluted to 50 mL with deionized water, and filtered. Macronutrients (K, P) and micronutrients (Fe, Mn, Zn, Na) were quantified using standard calibration curves and expressed as mg kg^-^¹ DW. Total soluble sugars were estimated following ethanol extraction and reaction with Anthrone reagent. Absorbance was measured at 620 nm, and sugar concentration was calculated against a glucose standard curve and expressed as mg/g dry weight.
ROS, H2O2, proline, and phenolic compounds
2.7
Hydrogen peroxide (H_2_O_2_) content was estimated following Velikova et al (Velikova et al., 2000). by reacting leaf extracts with KI and measuring absorbance at 390 nm. Reactive oxygen species (ROS) were quantified using the nitro blue tetrazolium (NBT) method as described by Jambunathan (Jambunathan, 2010), with absorbance recorded at 530 nm. ROS values were expressed as arbitrary absorbance units per g fresh weight (AU/g FW).
Proline concentration was determined using the acid ninhydrin method of Bates et al (Bates et al., 1973), and absorbance was measured at 520 nm after toluene extraction. Total phenolic content was quantified using the Folin–Ciocalteu method from methanol-extracted leaf samples, with absorbance recorded at 765 nm after incubation at 40 °C for 30 minutes. Total phenolic content was expressed as gallic acid equivalents (mg GAE g^-^¹ DW) using a gallic acid calibration curve.
Statistical analysis
2.8
Data were tested for normality and homogeneity of variance prior to analysis. Differences among all treatment combinations (species × salinity) were evaluated using one-way ANOVA, considering each combination as an independent group. When significant differences were detected, means were separated using Tukey’s HSD test at P ≤ 0.05. Statistical analyses were performed using R Studio (version 2022.07.1; R Core Team, Vienna, Austria), and graphical representations were prepared with SigmaPlot (version 12.0) and R Studio. Morphological and water-relation traits were measured using five biological replicates per treatment (n = 5). Gas-exchange parameters were measured on three biological replicates per treatment (n = 3) due to instrument measurement time limitations. For biochemical analyses, tissues from independent plants were analyzed separately (n = 6 biological replicates). All assays were performed with three technical replicates per biological replicate. Data are presented as means ± standard deviation (SD), unless otherwise stated. For mineral nutrient concentrations (Table 3), each organ was analyzed separately due to inherent physiological differences among plant organs. Therefore, nutrient concentrations were compared across the combined groups (species × salinity) using one-way ANOVA followed by Tukey’s HSD test (P ≤ 0.05).
Results
3
Survival rate
3.1
Survival differed significantly among species (P< 0.05) at the highest salinity level (25 dS m^-^¹). Under control conditions, all species exhibited 100% survival. Under severe salinity, L. shawii, S. persica, and H. salicornicum maintained full survival (100%), whereas C. comosum showed significantly lower survival under severe salinity (83.3%).
Plant height
3.2
Plant height differed significantly among species (P< 0.05), with the greatest reduction observed in C. comosum under severe salinity (from 58.3 to 29.0 cm), followed by H. salicornicum (69.7 to 32.0 cm), S. persica (96.0 to 44.3 cm), and L. shawii (100.0 to 55.3 cm; Figures 1, 2). The salt-injury index increased progressively with salinity in all species. Under severe salinity, C. comosum exhibited the highest salt injury index, H. salicornicum showed intermediate salt injury index, and S. persica and L. shawii displayed the lowest salt injury index levels (Table 4).
Representative photographs of four native UAE shrubs (Calligonum comosum, Haloxylon salicornicum, Lycium shawii, and Salvadora persica) under control, 10 and 25 dS m-¹ salinity treatments. Images document visual phenotypic responses only; quantitative differences are presented in subsequent figures.
Effects of salinity on morphological traits of the studied species. (A) Plant height (cm). (B) Leaf area in S. persica and L. shawii, and stem segment length in C. comosum and H. salicornicum (cm² or cm). (C) Root-to-shoot ratio, reflecting biomass allocation under salinity. Data are means ± SD (n = 5).
Leaf area and stem segment length
3.3
Leaf area or stem segment length varied significantly among species (p ≤ 0.05). In leafy species, S. persica showed a slight but significant reduction (2.53 to 2.30 cm²), whereas L. shawii exhibited a moderate decline (6.07 to 5.60 cm²). Among leafless species, C. comosum displayed a marked reduction in stem segment length (5.33 to 2.20 cm), whereas H. salicornicum showed a smaller decrease (5.27 to 4.67 cm). Tukey’s post-hoc comparisons confirmed significant treatment effects within each species (Figure 2B).
Biomass production
3.4
Biomass accumulation differed significantly among species (P< 0.05), with species-specific responses. L. shawii showed moderate reductions in shoot fresh weight (22.5 to 12.4 g) and a slight decrease in root dry weight (5.6 to 5.1 g) under severe salinity. In contrast, C. comosum experienced the greatest biomass loss, with root fresh weight declining from 30.7 to 3.9 g and root dry weight from 26.1 to 2.4 g. S. persica maintained relatively stable shoot and root biomass across treatments (P > 0.05; Figure 2C).
Root-to-shoot ratios
3.5
Root-to-shoot ratios varied significantly among species (P< 0.05), with contrasting patterns among species. Under severe salinity, L. shawii exhibited the highest root-to-shoot ratio. Conversely, C. comosum showed a significant decline in root-to-shoot ratio (from 1.10 to 0.74), resulting in the lowest value among all species under severe salinity (Figure 2C). These differences were confirmed by Tukey’s HSD comparisons.
Effects of salinity on relative water content
3.6
RWC differed significantly among species (P< 0.05), whereas within S. persica no difference was detected between control and severe salinity. C. comosum exhibited the largest decline (77.5 to 56.6%). L. shawii (80.5 to 74.0%) and H. salicornicum (80.2 to 75.9%) showed moderate reductions. In contrast, S. persica maintained relatively stable RWC (86.3 to 83.3%), with no significant difference between control and severe salinity (P > 0.05; Figure 3bA). A strong negative correlation was observed between RWC and electrolyte leakage (r = −0.92).
Effects of salinity on physiological and biochemical traits of four native plant species under control, moderate, and severe salinity conditions. (a) Oxidative and membrane damage–related parameters: (A) malondialdehyde (MDA), (B) total soluble sugars, (C) electrolyte leakage, and (D) leaf electrical conductivity. (b) Plant water status: (A) relative water content (RWC). Bars represent means ± SD (n = 5). Different lowercase letters indicate significant differences among treatments according to Tukey’s HSD test (P ≤ 0.05). Sample sizes are provided in each figure caption.
Effects of salinity on micro- and macronutrient content
3.7
Na^+^ accumulation varied significantly among species across all organs (Table 3). Under severe salinity, H. salicornicum showed the highest shoot Na^+^ concentration (37,333.2 mg kg^-^¹), whereas S. persica and L. shawii exhibited lower shoot Na^+^ levels. Root K^+^ declined significantly in L. shawii (from 4,655.6 to 3,437.6 mg kg^-^¹), whereas C. comosum and H. salicornicum maintained relatively high shoot K^+^ concentrations even under severe salinity. Phosphorus responses varied by species and organ. C. comosum roots showed a significant increase under moderate salinity (2,598.5 to 4,562.5 mg kg^-^¹), whereas H. salicornicum shoots exhibited a significant decrease under severe salinity. Micronutrient patterns also differed among species. L. shawii accumulated notably high Zn in shoots under severe salinity (155.2 mg kg^-^¹), while H. salicornicum maintained consistently high Fe and Mn concentrations in roots.
Effects of salinity on total soluble sugars, lipid peroxidation, and ionic homeostasis
3.8
Total soluble sugars differed significantly among species–salinity combinations (P ≤ 0.05). Under severe salinity, L. shawii showed the highest sugar content (206 mg g^-^¹ DW), followed by S. persica (203 mg g^-^¹), C. comosum (198 mg g^-^¹), and H. salicornicum (195 mg g^-^¹). Lipid peroxidation (MDA) varied significantly among species–salinity combinations (P ≤ 0.001). C. comosum exhibited the highest MDA levels (17.5 nmol g^-^¹ FW), whereas S. persica maintained the lowest (4.8 nmol g^-^¹ FW).
Electrolyte leakage (EL) differed significantly among species–salinity combinations, with the highest values recorded in C. comosum (91%). L. shawii (82%) and H. salicornicum (81%) showed intermediate values, whereas S. persica exhibited the most stable EL. Electrical conductivity (EC_leaf) varied significantly among species–salinity combinations, particularly in C. comosum (18,671 µS cm^-^¹; Figure 3a D). MDA was negatively correlated with plant height (Pearson r = −0.73) and RWC (r = −0.92), as confirmed by correlation analysis.
Effects of salinity on photosynthetic pigments
3.9
Chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids differed significantly among species (p ≤ 0.05). C. comosum exhibited the lowest total chlorophyll under severe salinity (237.3 µg g^-^¹ FW). H. salicornicum showed a marked reduction in chlorophyll a (165.7 µg g^-^¹ FW) and carotenoids (10 µg g^-^¹ FW). In contrast, L. shawii maintained relatively stable pigment levels, with only moderate declines in chlorophyll a and no significant change in carotenoids (13.1 to 13.3 µg g^-^¹ FW). S. persica displayed intermediate reductions in total chlorophyll (407.2 µg g^-^¹ FW; Figure 4).
Photosynthetic pigment contents of four native shrubs under different salinity levels. (A) Chlorophyll (a) (B) Chlorophyll (b) (C) Total chlorophyll (µg g-¹ FW). (D) Carotenoids (µg g-¹ FW). Data are means ± SD (n = 5).
Effects of salinity on photosynthetic parameter
3.10
All gas exchange parameters varied significantly among species (P< 0.05). Under severe salinity, S. persica exhibited the highest photosynthetic rate (7.18 µmol CO_2_ m^-^² s^-^¹), followed by L. shawii (4.08), while H. salicornicum (2.06) and C. comosum showed lower rates. Stomatal conductance was highest in S. persica (0.043 mol m^-^² s^-^¹). Intercellular CO_2_ concentration was significantly reduced in H. salicornicum and L. shawii. VPDleaf was higher in S. persica and L. shawii (>4.2 kPa), whereas H. salicornicum maintained lower VPDleaf (1 kPa). Transpiration rate followed a similar pattern, being highest in S. persica (2.20 mmol m^-^² s^-^¹) and lowest in H. salicornicum (0.33 mmol m^-^² s^-^¹; Figure 5).
Gas-exchange responses of four native shrubs under salinity. (A) Net photosynthetic rate [(A), µmol CO2 m-² s-¹]. (B) Stomatal conductance (gs, mol m-² s-¹). (C) Intercellular CO2 concentration (Ci, µmol mol-¹). (D) Leaf vapor pressure deficit (VPDleaf, kPa). (E) Transpiration rate (E, mmol m-² s-¹). Data are means ± SD (n = 3).
Effects of salinity on DPPH and antioxidant activity
3.11
Non-enzymatic antioxidant activity differed significantly among species (p ≤ 0.05). S. persica exhibited the highest DPPH scavenging activity (83.9%), followed by H. salicornicum (28.4%) and L. shawii (25.4%), whereas C. comosum showed minimal activity (6.3%). S. persica displayed the highest CAT (20.15 U mg^-^¹ protein), POD (84.3), SOD (99.5), and PPO (100) activities under severe salinity. L. shawii exhibited intermediate enzyme activities, whereas C. comosum consistently showed the lowest CAT, POD, and PPO activities (Figure 6).
Antioxidant responses of the studied species under control, moderate, and severe salinity. (A) DPPH radical scavenging activity (%). (B) Catalase (CAT) activity. (C) Peroxidase (POD) activity. (D) Superoxide dismutase (SOD) activity. (E) Polyphenol oxidase (PPO) activity. Values are means ± SD (n = 6).
Effects of salinity on proline, oxidative markers, total phenolic content, and protein levels
3.12
Total phenolic content varied significantly among species (p ≤ 0.05). S. persica showed the highest phenolic levels (32.3 mg GAE g^-^¹ DW), followed by L. shawii and H. salicornicum, while C. comosum had the lowest (24.5 mg GAE g^-^¹ DW). Proline increased significantly in S. persica (4.9 mg g^-^¹ DW) and L. shawii (2.5 mg g^-^¹ DW) but declined sharply in C. comosum (0.05 mg g^-^¹ DW). Total protein content increased under severe salinity in S. persica (72.6 mg g^-^¹ DW) and L. shawii (59.9 mg g^-^¹ DW) but decreased in C. comosum (25.4 mg g^-^¹ DW). ROS and H_2_O_2_ levels differed significantly among species–salinity combinations (P ≤ 0.05). C. comosum exhibited the highest ROS (64 AU/g FW) and H_2_O_2_ (92 µmol g^-^¹ FW), whereas S. persica and L. shawii showed lower oxidative stress levels (Figure 7).
Biochemical and oxidative stress indicators in four native shrubs under salinity. (A) Total phenolic content (mg GAE g-¹ DW). (B) Proline content (mg g-¹ DW). (C) Reactive oxygen species (ROS, AU/g FW). (D) Hydrogen peroxide (H2O2, µmol g-¹ FW). (E) Total soluble protein (mg g-¹ DW). Values are means ± SD (n = 6).
Correlation analysis and principal component analysis
3.13
Pearson correlation heatmap revealed clear associations among physiological and biochemical traits under salinity stress (Figure 8). Oxidative stress indicators (ROS, MDA, H_2_O_2_, electrolyte leakage, and EC_leaf) were strongly positively correlated with each other and negatively correlated with plant performance traits including plant height and relative water content. Notably, RWC showed a strong negative correlation with electrolyte leakage (r = −0.92), and plant height was negatively correlated with MDA (r = −0.73), confirming the impact of membrane damage and oxidative stress on growth reduction. Conversely, osmoprotectants and defense metabolites (proline, total phenolics, antioxidant enzymes, and protein content) showed strong positive relationships, with proline highly correlated with total phenolics (r = 0.86) and total protein (r = 0.72), indicating coordinated biochemical protection mechanisms. Ion homeostasis traits formed a distinct cluster: Na^+^ accumulation was associated with oxidative stress variables, whereas K^+^ retention was positively related to photosynthetic pigments and growth traits, suggesting its role in maintaining physiological stability under salinity. This pattern corresponded closely with PCA clustering, confirming that oxidative damage variables drive species separation under salinity stress.
Pearson correlation heatmap illustrating relationships among selected physiological, biochemical, and ionic traits under salinity. The heatmap represents the lower triangular correlation matrix with hierarchical clustering applied to variables using Ward’s method and Euclidean distance. Warmer colours (red) denote positive correlations, whereas cooler colours (blue) indicate negative correlations, with colour intensity reflecting the magnitude of correlation coefficients.
Principal component analysis (PCA) biplot (Figure 9) further integrated these relationships. The first two principal components explained 27.7% (PC1) and 24.4% (PC2) of total variance. PC1 represented an oxidative–ionic stress gradient, positively associated with ROS, MDA, and H_2_O_2_ and negatively associated with chlorophyll and internal CO_2_ concentration (Ci). PC2 represented biochemical defense activation, associated with proline accumulation, antioxidant enzymes (SOD, CAT, PPO), and phenolic compounds.
Principal component analysis (PCA) biplot showing relationships among physiological and biochemical traits under salinity stress. Arrows indicate variable loadings. PC1 represents an oxidative–ionic stress gradient, while PC2 represents biochemical defence activity. Coloured points represent plant species: Calligonum comosum, Haloxylon salicornicum, Lycium shawii, and Salvadora persica. Species separation reflects distinct salt-tolerance strategies, ranging from oxidative damage-dominated response to maintenance of photosynthetic stability and antioxidant protection. Colored symbols indicate species identity: C. comosum (purple triangles), H. salicornicum (green squares), L. shawii (cyan circles), and S. persica (red triangles).
Species separation reflected distinct tolerance strategies. In the biplot, species were distinguished by color: C. comosum (purple), H. salicornicum (green), L. shawii (cyan), and S. persica (red). C. comosum clustered within the oxidative stress region, indicating susceptibility to salinity. H. salicornicum occupied an intermediate ionic-adjustment region. L. shawii was positioned along the biochemical defense axis, showing osmotic and antioxidant adaptation. S. persica grouped with photosynthetic integrity traits, maintaining chlorophyll and lower oxidative damage.
Together, the heatmap and PCA demonstrate that salinity tolerance among species is governed by three coordinated adaptive strategies: oxidative stress avoidance, ionic regulation, and biochemical defense activation.
Discussion
4
Exposure to salinity resulted in species-specific responses that reflected contrasting tolerance strategies among the studied native shrubs. All species survived under control and moderate salinity conditions; however, under severe salinity, Calligonum comosum exhibited a reduced survival rate (83.3%), indicating comparatively lower tolerance (Latef et al., 2023). This reduced performance aligns with previous findings suggesting that C. comosum is primarily adapted to arid, sandy environments rather than highly saline soils, relying more on structural and mechanical adaptations than biochemical salt tolerance mechanisms (Soliman et al., 2018; Latef et al., 2023). In contrast, Salvadora persica and Lycium shawii maintained full survival with minimal salt injury index (SI: 1–1.5), demonstrating superior tolerance. Survival alone did not reflect physiological tolerance differences among species, as substantial sub-lethal damage occurred in less tolerant species and that physiological injury indices provide deeper insights into hidden salinity effects (Table 4).
Plant height differed significantly among species–salinity combinations (P< 0.05), with the greatest reduction observed in C. comosum under severe salinity (50.2%), likely due to combined osmotic stress and ion toxicity that inhibited cell expansion and shoot elongation. In contrast, S. persica and L. shawii exhibited better growth maintenance, possibly related to hormonal regulation (e.g., ABA and ethylene signaling) (Müller, 2021; Lim et al., 2022; Nazir et al., 2024). Leaf area and stem development were also differentially affected. S. persica showed the least reduction in leaf area, suggesting efficient osmotic protection and ion exclusion mechanisms, while C. comosum experienced marked stem reduction, consistent with its weaker tolerance (Mann et al., 2023). Biomass allocation patterns further supported species-specific adaptation strategies. S. persica and L. shawii increased root-to-shoot ratios under high salinity, indicating a strategic shift toward enhanced water and nutrient acquisition. This pattern is consistent with salt-tolerant species such as Saussurea salsa, which similarly increase root investment under salinity stress as an adaptive survival mechanism (Arif et al., 2020; Monson et al., 2022; Li et al., 2021; Bao et al., 2025).
Relative water content (RWC) declined in all species with increasing salinity, but to varying extents. S. persica maintained the most stable RWC (83.3%), likely due to deep rooting, effective osmotic adjustment, and strong biochemical defenses. H. salicornicum also maintained relatively high RWC, attributed to its succulent tissues that dilute internal salt concentrations and reduce transpiration. In contrast, C. comosum exhibited the sharpest RWC decline (56.6%), reflecting limited osmotic adjustment and ion exclusion capacity. These findings align with previous studies on Atriplex halimus, where populations with stronger osmotic regulation showed improved water-use efficiency under salinity (Martinez et al., 2003).
Salinity altered nutrient uptake and ion compartmentalization in all species. Sodium (Na^+^) accumulation increased across species but was particularly pronounced in H. salicornicum and S. persica, likely due to efficient vacuolar sequestration mechanisms that help maintain cytosolic ionic balance. L. shawii exhibited a decline in root K^+^ under severe salinity, reflecting Na^+^–K^+^ competition at uptake sites. However, S. persica and C. comosum maintained relatively high shoot K^+^ levels, suggesting selective K^+^ transport and Na^+^ compartmentalization (Malakar and Chattopadhyay, 2021; Sun et al., 2024). These findings are consistent with salt-tolerant crops such as cotton, where genotypes exhibiting better Na^+^ sequestration and higher K^+^ retention demonstrate enhanced salinity tolerance through improved ionic homeostasis (Sun et al., 2021).
Chlorophyll and carotenoid content varied significantly among species, particularly in C. comosum, likely due to activation of chlorophyll-degrading enzymes and downregulation of pigment biosynthesis genes (Wang et al., 2024). In contrast, L. shawii maintained pigment stability, reflecting stronger antioxidant protection (Li et al., 2022). Gas exchange parameters declined in all species. C. comosum showed almost complete inhibition of photosynthesis, whereas S. persica maintained the highest photosynthetic rate under severe salinity, likely due to efficient stomatal regulation and osmotic adjustment. H. salicornicum adopted a conservative water-use strategy through ABA-mediated stomatal closure, reducing water loss but limiting carbon assimilation. These patterns support previous findings in sorghum and other salt-tolerant species that rely on coordinated stomatal and osmotic regulation to maintain physiological function under salinity (Sakoda et al., 2021; Dourado et al., 2022).
Proline accumulation increased significantly in S. persica and L. shawii, indicating strong osmotic adjustment capacity, likely mediated by upregulation of P5CS activity. Proline likely contributed to membrane stabilization, ROS detoxification, and redox balance. In contrast, C. comosum showed a sharp decline in proline content, suggesting poor osmotic regulation (Maghraby and Alzalaty, 2025). Similar patterns of proline accumulation have been reported in turfgrass, where enhanced proline levels mitigate dehydration and oxidative damage (Jiang et al., 2023). Total soluble sugars increased in all species under salinity, with L. shawii exhibiting the highest accumulation. These sugars likely contributed to osmotic balance and acted as signaling molecules regulating stress-responsive pathways. However, in C. comosum, sugar accumulation was insufficient to counteract pigment degradation and metabolic disruption (Afzal et al., 2021; Nawaz et al., 2022).
Salinity led to increased ROS and H_2_O_2_ accumulation in all species, with C. comosum showing the highest levels due to weaker antioxidant defenses. S. persica exhibited the strongest enzymatic antioxidant response (high CAT and POD activity), effectively mitigating oxidative damage. This pattern aligns with findings in salt-tolerant maize genotypes that exhibit stronger ROS-scavenging systems (Qiao et al., 2021; Rohman et al., 2024).
Lipid peroxidation (MDA) was highest in C. comosum, reflecting membrane instability, while S. persica maintained the lowest MDA levels, indicating superior oxidative stress tolerance. Total phenolic content increased across all species, but was highest in S. persica, further supporting its strong non-enzymatic antioxidant capacity. These results are consistent with a previous study in Catharanthus roseus, which maintains low MDA under stress through enhanced antioxidant defense (Ali et al., 2021). Protein content increased in L. shawii and S. persica under severe salinity, likely due to synthesis of stress-responsive proteins such as LEA proteins, which protect cellular structures during dehydration (Abdul Aziz et al., 2021; Hsiao, 2024).
Correlation analysis confirmed that ionic toxicity and osmotic stress were major constraints under high salinity, as indicated by strong negative relationships between RWC and electrolyte leakage. Furthermore, multivariate analysis provided integrative evidence for the mechanistic interpretation of salinity tolerance. The correlation heatmap revealed two clearly opposing functional networks: a stress-damage cluster (ROS, MDA, H_2_O_2_, electrolyte leakage, and EC_leaf) and a protection cluster (proline, phenolics, antioxidant enzymes, and protein content). The strong negative associations between oxidative markers and growth traits confirmed that membrane injury and oxidative imbalance were the primary drivers of growth inhibition under salinity rather than osmotic limitation alone.
The PCA supported this interpretation by separating species along two principal functional axes. PC1 represented the oxidative–ionic stress gradient, distinguishing the highly damaged C. comosum from the more stable species, whereas PC2 represented biochemical defense activation associated with osmotic adjustment and antioxidant protection. S. persica clustered with photosynthetic stability traits, L. shawii with biochemical protection mechanisms, and H. salicornicum occupied an intermediate ionic-regulation position. This multivariate separation confirms that salinity tolerance in native desert shrubs is governed by coordinated but species-specific physiological strategies rather than a single tolerance mechanism. Positive correlations among proline, phenolics, and protein content highlight the coordinated role of osmotic adjustment and antioxidant defense in salt-tolerant species. C. comosum in its native habitat prefers dune slopes and semi-stabilized sands, where transient droughts occur but salinity remains moderate. Similar trends were reported in Calligonum caput-medusae under water deficit (Wang et al., 2024), highlighting the genus’s reliance on opportunistic growth rather than constitutive salt tolerance. These findings are consistent with earlier drought-focused assessments of UAE desert species, where L. shawii and S. persica showed superior tolerance through coordinated physiological, biochemical, and structural adaptations (Abo Amin et al., 2025).
The findings highlight S. persica and L. shawii as strong candidates for saline land rehabilitation and afforestation programs in arid regions. These species offer ecological benefits, including erosion control, biodiversity enhancement, and habitat provision, as well as agricultural benefits such as forage production and potential use in salt-tolerant crop breeding. However, large-scale implementation requires efficient propagation methods such as tissue culture and cost-effective restoration strategies.
Unlike previous studies that examined these species individually or under controlled greenhouse conditions, the present work demonstrates that salinity tolerance in UAE native shrubs is not governed by a single mechanism but by three coordinated strategies (ionic regulation, biochemical defense activation, and oxidative stress avoidance), and that species occupy distinct functional tolerance niches under real desert environmental conditions.
This study provides a comprehensive comparative assessment of physiological, biochemical, and ionic responses of four key UAE native shrubs as an outdoor pot trial under ambient desert conditions salinity treatments. The results advance understanding of species-specific salinity tolerance mechanisms and providing an ecological-physiological framework for selecting native species in desert salinity rehabilitation and biosaline agriculture programs.
Conclusion
5
Among the studied species, S. persica and L. shawii had the highest tolerance to salinity stress, maintaining better water status, photosynthetic performance, and antioxidant protection. In contrast, C. comosum showed pronounced physiological damage, indicating limited tolerance.
This study provides an integrated comparison of salinity adaptation mechanisms in these native species and highlights S. persica and L. shawii as promising candidates for saline land rehabilitation, biosaline agriculture, and ecological restoration in arid regions. These traits may provide useful targets for future breeding and molecular studies aimed at improving crop salinity tolerance. Future work should validate the underlying molecular mechanisms and support large-scale propagation to facilitate practical deployment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdul Aziz M. Sabeem M. Mullath S. K. Brini F. Masmoudi K. (2021). Plant group II LEA proteins: intrinsically disordered structure for multiple functions in response to environmental stresses. Biomolecules 11, 1662. 34827660 10.3390/biom 11111662 PMC 8615533 · doi ↗ · pubmed ↗
- 2Abdulfatah H. F. (2022). Non-enzymatic antioxidants in stressed plants: A review. J. Univ. Anbar. Pure. Sci. 16, 25–37. doi: 10.37652/juaps.2022.176435 · doi ↗
- 3Abo Amin S. Hayat F. Alyafei M. (2025). Drought tolerance strategies of some native plants in the UAE desert: growth, biochemical, and antioxidant insights. BMC Plant Biol. 25, 1374. doi: 10.1186/s 12870-025-07305-z, PMID: 41087857 PMC 12522444 · doi ↗ · pubmed ↗
- 4Afzal S. Chaudhary N. Singh N. K. (2021). “ Role of soluble sugars in metabolism and sensing under abiotic stress,” in Plant Growth Regulators: Signaling Under Stress Conditions. Eds. Iqbal M. Nazar R. Khan N. ( Springer, Cham), 305–334. doi: 10.1007/978-3-030-61153-7_13, PMID: · doi ↗
- 5Al-Busaidi A. Cookson P. Yamamoto T. (2005). Methods of p H determination in calcareous soils: use of electrolytes and suspension effect. Soil Res. 43, 541–545. doi: 10.1071/SR 04102, PMID: 41161682 · doi ↗
- 6Ali E. F. El-Shehawi A. M. Ibrahim O. H. M. Abdul-Hafeez E. Y. Moussa M. M. Hassan F. A. S. (2021). A vital role of chitosan nanoparticles in improvisation the drought stress tolerance in Catharanthus roseus (L.) through biochemical and gene expression modulation. Plant Physiol. Biochem. 161, 166–175. doi: 10.1016/j.plaphy.2021.02.008, PMID: 33610861 · doi ↗ · pubmed ↗
- 7Ali S. S. El-Zawawy N. A. Al-Tohamy R. El-Sapagh S. Mustafa A. M. Sun J. (2020). Lycium shawii Roem. & Schult.: A new bioactive antimicrobial and antioxidant agent to combat multi-drug/pan-drug resistant pathogens of wound burn infections. J. Tradit. Compl. Med. 10, 13–25. doi: 10.1016/j.jtcme.2019.01.004, PMID: 31956554 PMC 6957848 · doi ↗ · pubmed ↗
- 8Arif Y. Singh P. Siddiqui H. Bajguz A. Hayat S. (2020). Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 156, 64–77. doi: 10.1016/j.plaphy.2020.08.042, PMID: 32906023 · doi ↗ · pubmed ↗