BrDMC1-mediated tolerance during pollen meiosis under heat stress in Brassica rapa
Xulin Wang, Jialin Guo, Gongyao Shi, Weiwei Chen, Gangqiang Cao, Baoming Tian, Luyue Zhang, Fang Wei, Zhengyu Wang

TL;DR
This study shows how a gene called BrDMC1 helps Brassica rapa tolerate heat during pollen development, improving fertility and reducing cell damage.
Contribution
The study identifies BrDMC1 as a novel regulator of heat tolerance during pollen meiosis in Brassica rapa.
Findings
OE-BrDMC1 plants showed increased pollen fertility and reduced chromosomal abnormalities under heat stress.
Overexpression of BrDMC1 lowered ROS levels and increased antioxidant enzyme activity in heat-stressed plants.
Heat stress upregulated genes related to DNA repair and meiotic cell cycle in OE-BrDMC1 plants.
Abstract
High temperatures may have a substantial impact on cellular meiosis, and subsequently affects plant reproduction, development, and yield over time. In this study, using overexpressed transgenic lines, we show that BrDMC1, a gene involved in meiotic recombination, regulates heat tolerance during the early pollen development stage in Brassica rapa. According to the expression pattern analysis, BrDMC1.A03 was not discovered at the transcriptional level, whereas BrDMC1.A01 was highly expressed in young flower buds in B.rapa. The Cis-acting element prediction revealed that BrDMC1.A01 contains a low-temperature responsive element, and GUS histochemical analysis revealed an increased staining ability following temperature stress. Under normal conditions, there were no significant cytogenetic or molecular differences between wild-type (WT) and overexpressed-BrDMC1 (OE-BrDMC1).After 24 h of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
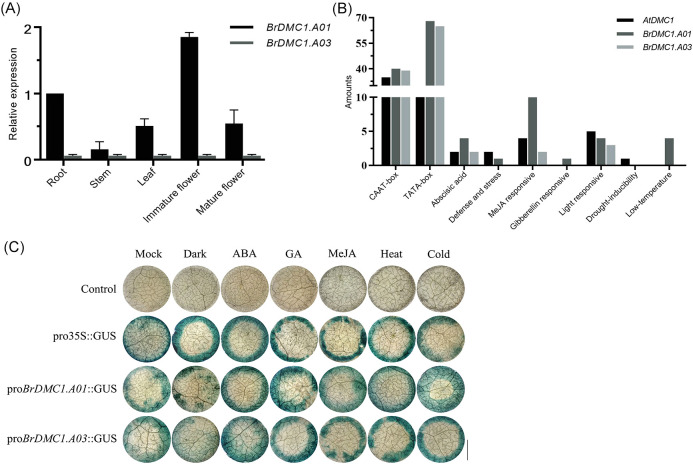 Figure 1
Figure 1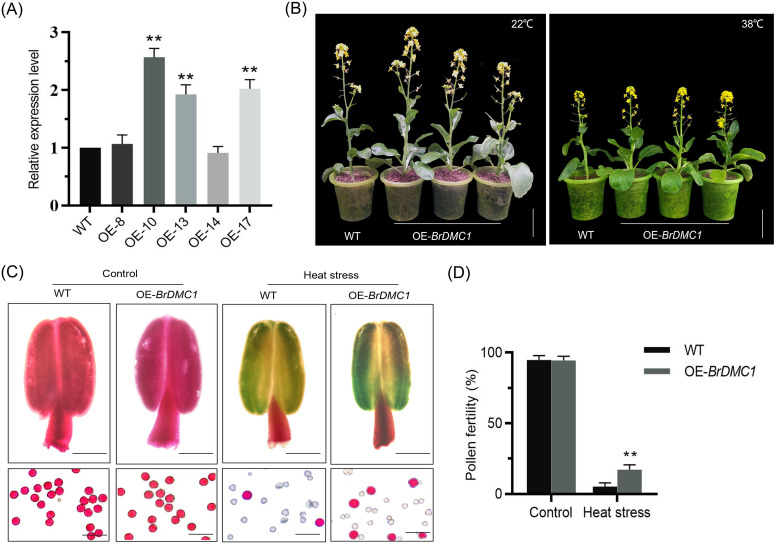 Figure 2
Figure 2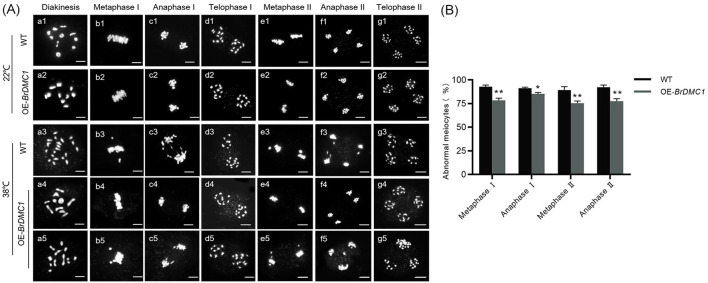 Figure 3
Figure 3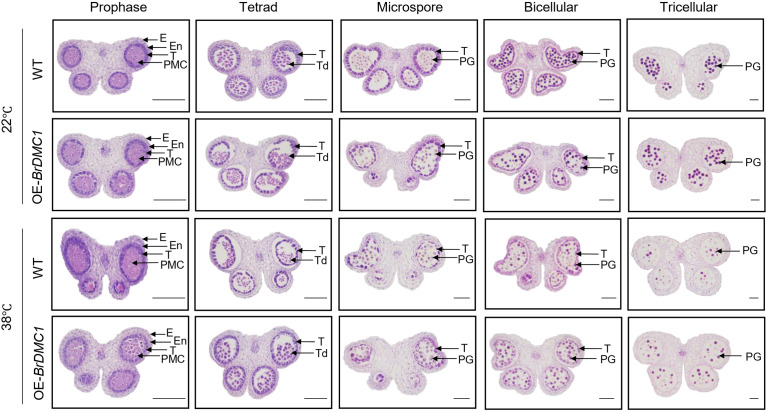 Figure 4
Figure 4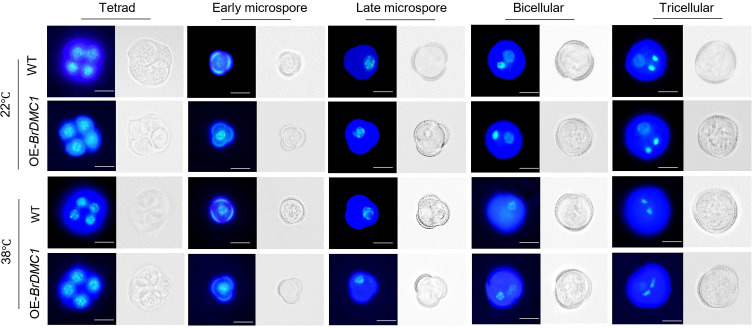 Figure 5
Figure 5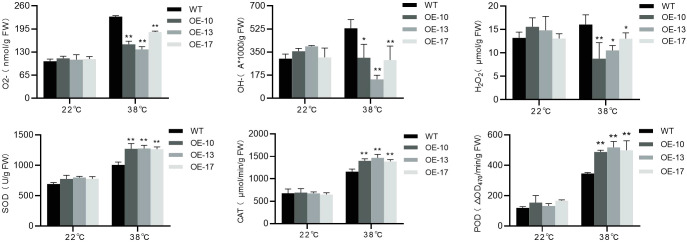 Figure 6
Figure 6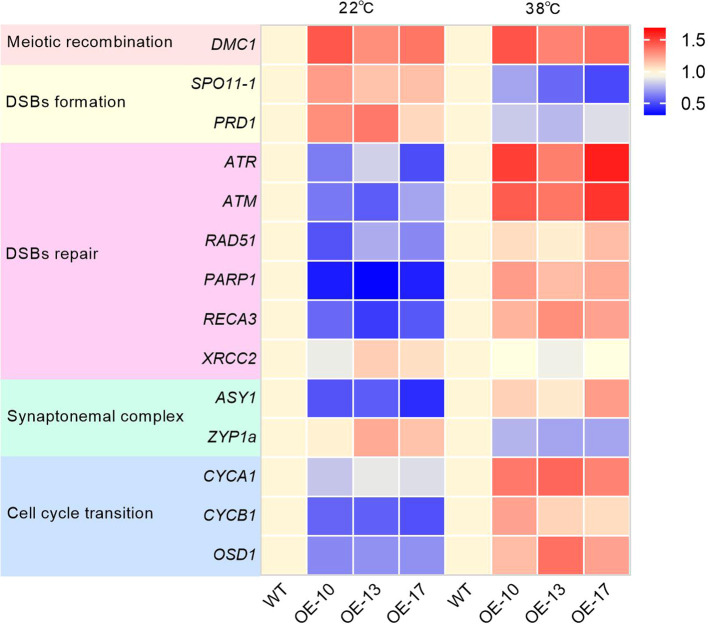 Figure 7
Figure 7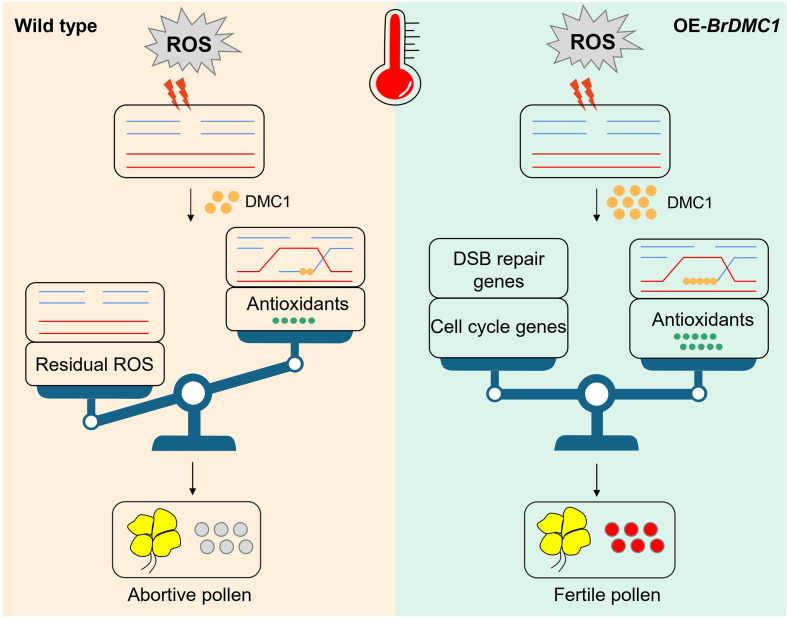 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · DNA Repair Mechanisms · Plant Molecular Biology Research
Introduction
1
Meiosis is an important phase in the sexual life cycle, influencing plant genetic diversity and ploidy stability. The highly conserved topoisomerase SPO11 and numerous other similar proteins facilitate the controlled production of DNA double-strand breaks (DSBs), which initiate meiotic recombination (Keeney et al., 1997; Grelon et al., 2001). SPO11 is endonucleolytically cleaved and removed by the MRX-N complex [MRE11-RAD50-XRS2 (Nbs1)] and Com1 (Sae2), followed by additional resection of the 5’ end by Exo1 to generate 3’ single-stranded DNA (ssDNA) (Borde, 2007). RPA binds to 3’ ssDNA ends to prevent degradation and increase recombinase loading (Ranjha et al., 2018). Subsequently, recombinases RAD51 or DMC1 replace RPA to bind to 3’ ssDNA ends, facilitating homologous search and single-strand invasion, ultimately forming crossover (CO) and non-crossover (NCO) products, with the former accompanied by genetic information exchange on chromosomes to generate genetic diversity and the latter resulting in limited genetic exchange (Bugreev et al., 2011; Kurzbauer et al., 2012; Yang et al., 2021). In plants, mutations in proteins involved in DSB generation and repair frequently lead to substantial meiotic abnormalities, defined by poor synaptonemal complex (SC) assembly, defective meiotic recombination (MR), and eventually reduced or lost plant fertility (Mercier et al., 2015).
High-temperature stress impacts several stages of plant reproduction, including meiosis, pollen formation, tapetal cells, reactive oxygen species (ROS) concentration, and fertilization (Endo et al., 2009; Parish et al., 2012; Djanaguiraman et al., 2018; Chen et al., 2021). Plant meiosis is quite sensitive to high temperatures. High temperatures (28°C) in the appropriate growth temperature range increase MR in Arabidopsis thaliana via increasing CO formation frequency (Modliszewski et al., 2018). Higher temperatures (32-34°C) could impact homologous synapsis in A. thaliana, reducing CO formation (De Jaeger-Braet et al., 2021). When temperatures reach the growth threshold (36-38°C), homologous chromosomal synapsis frequently fails, resulting in a dramatic decrease in MR (De Storme and Geelen, 2020). High temperature also disrupts chromosomal segregation and cytokinesis, resulting in abnormalities such as chromosome stickiness, lagging chromosomes, and micronucleus (Mai et al., 2019; Lei et al., 2020). Studies have demonstrated that ATM facilitates DSB repair under high temperature by acting downstream of the MRE11-RAD50-NBS1 (MRN) complex and functioning in a RAD51-independent but chromosomal axis-dependent way (Zhao et al., 2023). Furthermore, the tapetum plays an important role in the correct development of microspores and pollen grain maturation (Li et al., 2006, Li et al., 2011). In reaction to high temperatures during the microspore formation stage, tapetum-related genes in immature anthers are downregulated (Endo et al., 2009). Meanwhile, ROS are needed for normal cellular metabolic functions, but they are created and stored as harmful consequences of aerobic metabolism after plant exposure to abiotic stress. This causes significant intercellular biochemical damage (De Storme and Geelen, 2014; Mittler, 2017). Antioxidant enzymes and osmoprotectants can protect plants from oxidative stress (Choudhury et al., 2017; Khan et al., 2017; Wang et al., 2023).
The RecA-like recombinase family plays a key role in eukaryotic meiosis by facilitating homologous DNA search and strand exchange. Previous evolutionary analysis classified the RecA-like recombinase family into three subfamilies: Class I, the major recombinase subfamily RADα; Class II, the RADβ subfamily; and Class III, the RecA subfamily (Lin et al., 2006).DMC1 (disrupted meiotic control 1) is a member of the RADα subfamily that plays a role in meiotic recombination, DNA repair, and genomic stability, and repairs DSBs utilizing homologous chromosomes as templates during meiosis (Klimyuk and Jones, 1997; Crismani et al., 2013). Mutation or deletion of DMC1 causes failure in SC formation and incorrect segregation of homologous chromosomes, resulting in germ cell death (Pittman et al., 1998; Chen et al., 2016). Heat stress (36-38°C) inhibits MR by lowering SPO11-dependent DSBs and changing chromosomal axis shape, and DMC1 protein abundance decreases dramatically under high temperature (Ning et al., 2021). Studies have also found that DMC1 maintains seed germination under salt stress by regulating antioxidant enzyme activity and DNA repair gene expression, and BrDMC1-RNAi plants exhibit reduced antioxidant enzyme activity and germination rate (Wang et al., 2023). Previously, it was proposed in hexaploid wheat that the meiotic gene TaDMC1-D1 is most likely a candidate gene for maintaining synapsis and crossovers under low or high temperature conditions, and subsequent experiments confirmed this hypothesis, TaDMC1-D1 is responsible for maintaining normal crossover formation under low temperature and moderate high temperature (Draeger et al., 2020, Draeger et al., 2023). At 40°C, LIM15 (DMC1 homolog) in lilium is rapidly activated in anthers to stabilize the meiotic process and enhance crossover events, reflecting the functional conservation of DMC1 in responding to high temperature across different species (Ahn et al., 2025).
Meiotic recombination is a dynamic mechanism that is more sensitive to temperature fluctuations. However, how DMC1 influences the meiotic process under high-temperature stress is unknown. In this study, we show that DMC1 can respond to extreme temperature stress. Although both WT and OE-BrDMC1 lose pollen viability after high-temperature treatment, OE-BrDMC1 has much higher pollen viability than WT. Consistent with the pollen fertility phenotype, OE-BrDMC1 exhibit more normal chromosomal activities during meiosis than WT. Meanwhile, the tapetal cell and microspore growth processes in both WT and OE-BrDMC1 are significantly affected, with no significant difference. In comparison to WT, OE-BrDMC1 have considerably lower ROS levels and much higher antioxidant enzyme activity. Furthermore, high-temperature stress dramatically increases the expression levels of the SPO11-1, ATR, ATM, RECA3, CYCA1, and OSD1 in OE-BrDMC1. These findings suggest that DMC1 can positively influence heat tolerance during meiosis.
Materials and methods
2
Plant materials and growth conditions
2.1
Brassica rapa DH line CXL-45 and OE-BrDMC1 plants were cultured in an artificial climate chamber under conditions of 16 h light/22°C, 8 h dark/16°C, and relative humidity of around 60%. The Nicotiana benthamiana was cultured under 16 h light/25°C, 8 h dark/25°C, and relative humidity of around 60%.
High-temperature treatment
2.2
WT and OE-BrDMC1 blooming plants were placed in a growth cabinet at 38°C for 24 h, with 16 h of light and 8 h of darkness. All materials were treated between 10:00 and 11:00 a.m., and flower buds were collected for later studies.
Cis-acting element analysis of DMC1 promoter
2.3
The DMC1 promoter region was extracted using TBtools software and predicted using the Plant CARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/), with the results merged and analyzed. To create the proBrDMC1::GUS vector, a 1730 bp sequence upstream of BrDMC1 was cloned into the pEASY-T1 vector for amplification, then subcloned into the pCAMBIA3301 vector with XbaI and NcoI restriction sites, then transfected into Agrobacterium tumefaciens (EHA105) to complete vector synthesis. The created vector was infiltrated into tobacco leaves that had been grown for around four weeks. Exogenous hormones and stress treatments were applied after 48 h, and the results were seen 24 h later.
Cytology
2.4
Under normal conditions and high-temperature treatment, anthers from WT and OE-BrDMC1 were fixed in Carnoy’s solution (ethanol: glacial acetic acid, 3:1). Pollen was stained with Alexander’s stain for observation (Alexander, 1969). The procedure for observing the meiotic process by the squashing method was as previously described (Zhao et al., 2023). Microspore development was observed using 4’,6-diamidino-2-phenylindole (DAPI) staining (Caselli et al., 2019). Observation of tapetal development was carried out as previously described (Qian et al., 2021). Finally, the slides were examined and photographed with an upright fluorescence microscope.
Determination of ROS content
2.5
O_2_^•−^ combines with hydroxylamine to create NO_2_^-^, resulting in a pink azo color containing sulfanilic acid and α-naphthylamine. The highest light absorption of this dye is at 540 nm. The A540 value can be used to quantify the sample’s O_2_^•−^ concentration. H_2_O_2_ combines with a specific chromogenic agent to produce a colored material with a distinct absorption peak at 510 nm, revealing the sample H_2_O_2_ content. OH^•^ oxidizes 2-deoxyribose to a malondialdehyde analog, which then condenses with thiobarbituric acid (TBA) to produce a colorful product with a maximum absorption peak at 532 nm, revealing the OH^•^ concentration.
Determination of antioxidant enzyme activities
2.6
Anthers were thoroughly pulverized with a pre-cooled mortar and pestle in 10 mL of 50 mM sodium phosphate buffer (pH 7.8) containing 1% (w/v) polyvinylpyrrolidone. The homogenate was centrifuged at 15,000 g for 10 min at 4 °C. The supernatant from the crude enzyme extract was maintained in an ice bath (Ahmad et al., 2018). The photochemical approach was used to assess the activity of superoxide dismutase (SOD; EC 1.15.1.1). The activity of peroxidase (POD; EC 1.11.1.7) was determined using a substrate combination containing 50 mM sodium phosphate buffer (pH 6.0), 5 mM guaiacol, and 0.25 mM H_2_O_2_. POD activity was evaluated after adding 0.1 mL of enzyme extract by measuring the change in absorbance at 470 nm every 40 s. Catalase (CAT; EC 1.11.1.6) activity was assessed by measuring the decrease in absorbance at 240 nm every 30 s following H_2_O_2_ breakdown.
Quantitative reverse transcription PCR analysis
2.7
Total RNA was isolated from unopened flower buds of WT and OE-BrDMC1 plants grown under normal temperature conditions (22 °C) and after 38 °C treatment for 24 h, respectively. The Plant Total RNA Isolation Kit Plus (Foregene, Chengdu, China) was used to extract B.rapa total RNA, and the cDNA Synthesis Kit (TAKARA, Dalian, China) was used to create cDNA. We predicted the gene structure (using TBtools software) and functional domains (NCBI CDD database) of the meiotic gene copies provided for a focused analysis of the integrity of the core functional domains. Particular primers were made to solely amplify the target copies of the chosen copies. The LightCycler 480 System and qPCR Master Mix (Trans, Beijing, China) were used for qRT-PCR analysis. The following was the PCR program: 40 cycles of 95°C for 15 s, 60°C for 50 s, and a melting curve study from 72°C to 96°C were performed after 60 s at 95°C. As an internal control, the B.rapa β-ACTIN gene was employed. The 2^−ΔΔCt^ technique was used to calculate relative expression levels (Michaelidou et al., 2013). Using separate cDNA preparations, three biological replicates were carried out. The used primers are listed in Supplementary Table 1.
Statistical analyses
2.8
The experiment included at least three different biological and technological replicates. Data visualization and statistical analysis were carried out using GraphPad Prism 5. The significance of differences between the experimental and control groups was determined using the t-test and one-way ANOVA. Significant differences from the control are shown by **p < 0.01, and *p < 0.05.
Results
3
DMC1 promoter region shows response to different stress conditions
3.1
Bioinformatics analysis revealed that after the triplication event, BrDMC1 retained two copies in B.rapa, BrDMC1.A01 (ID: Bra023796) and BrDMC1.A03 (ID: Bra001890). Moreover, the genetic structure, conserved motifs, and conserved functional domains of BrDMC1.A01 were similar to those of AtDMC1, whereas BrDMC1.A03 retained only a partial structure (Supplementary Figure 1). The expression levels of different copies of BrDMC1 in different tissues of plants were analyzed. The results showed that BrDMC1.A01 was expressed in different plant tissues, and the expression level was the highest in young buds, followed by roots, leaves, and stems (Figure 1A). The expression level of BrDMC1.A03 is relatively low (Figure 1A), and it is speculated that its biological function is lost after replication events.
Analysis of the BrDMC1 promoter. (A) RT-qPCR analysis of the expression of BrDMC1 in various tissues of the WT. Three biological replicates were used for each sample. (B) Prediction of cis-acting elements of BrDMC1. (C) GUS staining of N. benthamiana leaves after different treatments to verify the activity of BrDMC1 promoter. Bar = 0.25 cm.
Cis-acting element analysis of DMC1 genes from A.thaliana and B.rapa revealed that both have light-responsive elements, defense and stress-responsive elements, GA-responsive elements, MeJA-responsive elements, and basic structural elements, CAAT-box and TATA-box, indicating promoter features (Figure 1B). Transient transfection of tobacco leaf cells was utilized to confirm the expression of the BrDMC1 promoter in order to see if the promoter fragment is active and how it reacts to various hormones and stress. The results showed that tobacco leaves injected with BrDMC1 promoter bacterial solution showed a blue color after GUS staining, indicating that BrDMC1 is an active promoter fragment (Figure 1C). To see how the BrDMC1 promoter reacted, several hormone and stress treatments were used. The result demonstrated that the blue color of leaves stained with GUS for the BrDMC1 promoter was deepened to varying degrees following the application of ABA, GA, MeJA hormones, and high-temperature and low-temperature treatments (Figure 1C). This is consistent with the prediction results of promoter cis-acting elements, all of which are induced by distinct hormones and stress conditions.
Pollen fertility of OE-BrDMC1 significantly increases under high-temperature stress
3.2
In order to investigate the role of DMC1 in plants, we created transgenic lines (OE-BrDMC1, OE-10/OE-13/OE-17) that expressed DMC1 much more than the WT (Figures 2A, and Supplementary Figure 2). Plant phenotypic observation revealed that WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) did not significantly differ in growth and development under normal conditions. Both plants displayed wilting following high-temperature stress (Figure 2B). Fertility studies demonstrated that under normal culture conditions, pollen grains of WT could be stained with Alexander’s stain, resulting in homogeneous pollen grain shape, and the same phenomena was found in OE-BrDMC1 lines (OE-10/OE-13/OE-17). The majority of pollen grains in both WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) showed abnormal discolored when exposed to high temperatures (Figure 2C). Further statistical examination of the stainability rate revealed that, under normal conditions, WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) had pollen stainability rates of 97.1% (n=1085) and 97.8% (n=1054), respectively. Under high-temperature stress, the pollen stainability rate of WT was 5.3% (n=1152), whereas that of OE-BrDMC1 lines (OE-10/OE-13/OE-17) was 17.3% (n=1023), which was significantly higher than that of WT (Figure 2D).
*Analysis of pollen fertility of BrDMC1. (A) Detection of DMC1 expression in WT and OE-BrDMC1 lines under normal temperature conditions. Three biological replicates were used for each sample. (B) The phenotypes of WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and high-temperature conditions. Bar = 7 cm. (C) Anther and pollen staining of WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. Bar = 100 mm. (D) Pollen staining rates of WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. *p < 0.01.
Abnormal meiotic chromosomal behaviors decrease in OE-BrDMC1 under high-temperature stress
3.3
To explore the effect of high temperature on B.rapa meiosis, we observed the complete process. The findings revealed that under normal culture conditions, the complete meiotic process in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) was normal. Despite the fact that both WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) displayed aberrations in the meiotic process after high-temperature treatment, OE-BrDMC1 lines (OE-10/OE-13/OE-17) continued to exhibit normal processes. At diakinesis, WT had mostly univalents, whereas OE-BrDMC1 lines (OE-10/OE-13/OE-17) had both univalents and bivalents (Figure 3A). Statistical investigation of abnormal meiotic processes revealed that at metaphase I, chromosomes were organized orderly on the equatorial plate in OE-BrDMC1 lines (OE-10/OE-13/OE-17), whereas lagging chromosomes and chromosomal bridges were identified in 78.5% (n=123) of pollen mother cells. During anaphase I, homologous chromosomes separated, and 85.3% (n=129) of pollen mother cells formed lagging chromosomes and chromosomal bridges. At metaphase II, 74.2% (n=117) of pollen mother cells contained lagging chromosomes. In anaphase II, 77.6% (n=126) of pollen mother cells had uneven chromosomal segregation and lagging chromosomes. Finally, uneven segregation at telophase caused the creation of three or four daughter cells (Figure 3B). Statistical analysis of WT showed that at different stages of meiosis, the proportion of pollen mother cells with abnormalities ranged from 88.1% to 94.8% (n=118~138) (Figure 3B), which was significantly higher than that of OE-BrDMC1 lines (OE-10/OE-13/OE-17), indicating that overexpression of BrDMC1 can enhance heat tolerance during meiosis.
*Observation of chromosome behavior during meiosis in BrDMC1 plants. (A) PI-stained chromosome spreads of male meiocytes during meiosis in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. Bar = 10 μm. (B) Percentages of abnormal meiocytes in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under heat-temperature conditions. **p < 0.01 and p < 0.05.
BrDMC1 does not affect tapetal development under high-temperature stress
3.4
The tapetum is essential for controlling the growth of pollen. Pollen abortion results from abnormal tapetal growth, which also prevents normal pollen grain development. We observed anthers in paraffin sections to see the morphology of the tapetum during anther development. The findings demonstrated that the epidermis, endothecium, middle layer, and tapetum of WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) did not significantly vary under normal conditions (Figure 4). After high-temperature stress treatment, the tapetal development process of OE-BrDMC1 lines (OE-10/OE-13/OE-17) and WT was nearly identical to that under normal conditions during the meiotic and tetrad stages. However, at the uninucleate microspore and binucleate stages, the tapetum color diminished (Figure 4), indicating premature degradation, and the tapetum was totally degraded by the trinucleate stage.
Observation of the development process of tapetum in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. (E) Epidermis layer, (En) Inner layer, (T) tapetum, (PMC) Pollen mother cell, (Td) Tetrad, (PG) Pollen grain. Bar = 100 μm.
BrDMC1 does not affect microspore development under high-temperature stress
3.5
DAPI was used to stain microspores at various stages of development in order to better observe the microspore formation process of WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17). The findings demonstrated that both WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) pollen mother cells underwent a typical microspore development process to produce trinucleate pollen grains under normal conditions (Figure 5). After high-temperature treatment, there were essentially no differences in microspore formation between WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) at the tetrad, early uninucleate, late uninucleate, binucleate, and trinucleate stages (Figure 5). Furthermore, we discovered a fascinating phenomenon: after high-temperature treatment, at the binucleate stage, the vegetative nucleus was distributed diffusely throughout the cell, whereas at the trinucleate stage, the vegetative nucleus vanished, implying that high temperature primarily affects the development of the vegetative nucleus rather than the generative nucleus of plants.
Observation of pollen development process in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. Bar = 10 μm.
BrDMC1 responds to high-temperature stress via anti-oxidative damage pathway
3.6
ROS, such as O_2_^•−^, H_2_O_2_, and OH^•^, build in plants during non-biological stress, causing oxidative damage to cells and interfering with genomic stability. Plants require antioxidant enzymes, osmoprotectants, and antioxidants to reduce the concentration of ROS and protect themselves from stress. Under normal conditions, the assessment of O_2_^•−^, H_2_O_2_, and OH^•^ contents reveals no substantial variation in the concentration of ROS between WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) (Figure 6). Following high-temperature stress, the contents of O_2_^•−^, H_2_O_2_, and OH^•^ in OE-BrDMC1 lines (OE-10/OE-13/OE-17) are much lower than those in WT, with extremely significant differences in O_2_^•−^ content and considerable differences in H_2_O_2_ and OH^•^ contents (Figure 6). After high-temperature stress, SOD, POD, and CAT activities in OE-BrDMC1 lines (OE-10/OE-13/OE-17) were significantly higher than in WT (Figure 6). The results show that OE-BrDMC1 lines (OE-10/OE-13/OE-17) exhibit lower oxidative damage and are more resistant to high-temperature stress.
*Determination of O2•−, H2O2, and OH• content and SOD, POD, and CAT activity in WT and OE-BrDMC1 lines (OE-10/OE-13/OE-17) under normal temperature and heat-temperature conditions. **p < 0.01 and p < 0.05.
BrDMC1 responds to high-temperature stress through DNA repair pathway and cell cycle transition
3.7
To investigate the molecular basis of BrDMC1 tolerance to high-temperature stress, we examined the expression of meiosis-related genes. Under normal conditions, the expression levels of SPO11–1 and PRD1 required for DSB formation increase, whereas the expression levels of ATR, ATM, RAD51, and RECA3 required for DSB repair decrease and that of XRCC2 remains steady. ASY1 and ZYP1a, two parts of the synaptonemal complex, are reduced and unchanged, respectively. Furthermore, there is a decrease in the expression levels of CYCA1, CYCB1, and OSD1, which regulate cell cycle transition (Figure 7). High temperature stress reduces the expression levels of SPO11–1 and PRD1, which are required for DSB formation, while increasing the expression levels of ATR, ATM, RAD51, and RECA3, which are required for DSB repair, as well as the expression levels of cell cycle transition genes CYCA1, CYCB1, and OSD1 (Figure 7). These findings suggest that BrDMC1 responds to high-temperature stress via the DNA repair pathway and cell cycle transition, which improves plant stress tolerance.
Expression pattern of genes related to DSBs formation, DSBs repair, synaptonemal complex, and cell cycle transitions during meiosis under normal temperature and heat-temperature conditions.
Discussion
4
With the acceleration of global climate change, regular severe temperature events have emerged as serious environmental threats to crop productivity. Brassica crops, which are key oil and vegetable crops, are highly susceptible to temperature fluctuations throughout growth and development. Meiosis, a critical mechanism in sexual reproduction, is very subject to temperature stress, which directly affects gamete formation and seed development quality (Lei et al., 2020).
DNA repair and antioxidant regulation mechanisms of DMC1
4.1
High temperature has a significant impact on plant reproductive development, particularly meiotic diseases, and BrDMC1 ensures fertility by maintaining the normal meiotic process. In this study, a large number of abnormalities such as lagging chromosomes, chromosome bridges, and unequal segregation were observed in WT pollen mother cells at high temperatures, whereas the incidence of these abnormal phenotypes was reduced in OE-BrDMC1 lines (OE-10/OE-13/OE-17), and pollen fertility was consistently improved (Figures 2, 3). This is consistent with wheat study results, where TaDMC1-D1 deletion mutants demonstrated lower meiotic crossovers and higher chromosome synapsis failure rate at 30°C (Draeger et al., 2023). In A.thaliana, high temperature suppresses DSB formation, lowers DMC1 activity, and impairs synaptonemal complex formation by altering the chromosomal axis (Ning et al., 2021). These findings suggest that DMC1, by regulating meiotic chromosomal processes, serves as an important barrier for B.rapa to survive high-temperature damage. Further cytological observations revealed that the tapetum of OE-BrDMC1 lines (OE-10/OE-13/OE-17) degraded prematurely under high temperatures (Figure 4), but pollen fertility and abnormal chromosomal behaviors were superior to those of WT (Figure 3), indicating that BrDMC1 selectively protects germ cells while prioritizing the protection of key meiotic processes over vegetative tissues. This is consistent with the result that LIM15, the DMC1 homolog in lilium, retains meiotic stability under high temperature (Ahn et al., 2025).
ROS eruption in plant cells under high-temperature stress causes membrane lipid peroxidation, protein denaturation, and genomic damage (Bokszczanin and Fragkostefanakis, 2013), and BrDMC1 also regulates the activity of the antioxidant system, which helps to maintain ROS balance. In addition to its role in homologous recombination and DNA repair, studies have discovered that RecA is also involved in the response to oxygen and heat stress, and the loss of RecA affects survival rate in response to heat shock and desiccation (Aranda et al., 2011). Overexpression of the ZmRAD51A in rice boosted the plant disease resistance, resulting to higher expression of defense-related genes (Liu et al., 2019). Under salt stress conditions, BrDMC1-RNAi plants exhibit decreased antioxidant enzyme activity, ROS generation, and seed germination rate (Wang et al., 2023). The study observed that OE-BrDMC1 lines (OE-10/OE-13/OE-17) had considerably lower levels of O_2_^•−^, H_2_O_2_, and OH^•^ following 38°C high-temperature treatment (Figure 6), suggesting that BrDMC1 may regulate intracellular ROS homeostasis through direct or indirect pathways to prevent oxidative damage. The activities of SOD, POD, and CAT in OE-BrDMC1 lines (OE-10/OE-13/OE-17) increased dramatically (Figure 6), demonstrating that these antioxidant enzymes work together to constitute the primary intracellular ROS scavenging mechanism. This is comparable to the prior observation in cabbage, which responds to a rise in ROS by boosting antioxidant defense under heat treatment (Soengas et al., 2018).
BrDMC1 creates a molecular defense network at high temperatures by activating DNA damage repair and cell cycle control genes. The qPCR research revealed that the expression level of SPO11–1 in OE-BrDMC1 lines (OE-10/OE-13/OE-17) plants was downregulated, while the expression levels of ATR, ATM, and RECA3 genes were dramatically increased(Figure 7). SPO11 is responsible for DSB creation, while ATR and ATM are involved in DNA damage repair (Chen and Weir, 2024). Furthermore, high temperatures increase the expression of the CYCA1 and OSD1. CYCA1 not only contributes in the transition from meiosis I to meiosis II, but also, coupled with OSD1, in the prophase transformation of meiosis I (d’Erfurth et al., 2010). The increased expression of these genes suggests that BrDMC1 may respond to high-temperature stress via these mechanisms, which include improving the formation and repair capacity of DNA double-strand breaks, activating DNA damage detection and signal transduction pathways, regulating cell cycle progression to ensure normal meiosis, and increasing homologous recombination capacity to maintain genome stability. In maize, HSP101 improves homologous recombination repair efficiency by boosting the loading of RAD51 at DSB sites, thus sustaining meiotic stability under high temperature (Li et al., 2022). Brassica napus transcriptome analysis under heat stress revealed that upregulated genes are primarily linked to heat shock proteins (HSPs) and other heat shock transcription factors, which play significant roles in heat tolerance (Kourani et al., 2025). Furthermore, HSPs can operate as molecular chaperones to prevent protein denaturation and aggregation, aid refold denatured proteins, and improve plant stress resistance (Zhang et al., 2022). Whether HSPs affect the function of DMC1 in meiosis under high temperature requires further research.
Functional conservation and evolution of DMC1
4.2
DMC1 is a gene discovered by screening a meiotic cDNA library in Saccharomyces cerevisiae. It is a meiosis-specific protein with structural similarities to the bacterial strand exchange recombinase RecA (Bishop et al., 1992). Meiotic recombination is initiated by programmed DSBs, generating single-stranded DNA ends at the break sites, DMC1 performs homology search by polymerizing on single-stranded ends to form nucleoprotein filaments, and conducts strand invasion and strand exchange between homologous chromosomes (Xu et al., 2023). DMC1 requires ATP to participate in homologous recombination, DNA repair, and genomic stability, and it executes DSB repair utilizing homologous chromosomes as templates during meiosis (Crismani et al., 2013; Klimyuk and Jones, 1997).
DMC1 homologs are found in a range of species. In yeast and mice, dmc1 mutants exhibit meiotic recombination anomalies, accompanied by chromosome synapsis problems, and cells halt at prophase, resulting in sterility (Bishop et al., 1992; Pittman et al., 1998; Yoshida et al., 1998; Bannister et al., 2007). In medaka dmc1 mutants, although synaptonemal complex formation is faulty and death occurs in testes, some spermatocytes complete meiosis and generate a modest number of sperm (Chen et al., 2016). In most diploid plant species, disruption of DMC1 causes sterility. A.thaliana possesses a single copy of DMC1, and synapsis failure occurs during meiosis I in Atdmc1 mutants, indicating aberrant crossovers and lower fertility (Couteau et al., 1999). Rice has two DMC1 homologs, DMC1a and DMC1b, and single mutants of either show no meiotic defects, however double mutants fail to generate bivalents, eventually resulting to full sterility (Wang et al., 2016). Barley has a single DMC1 homolog, HvDMC1, and mutation of this gene causes synapsis failure, chromosomal bridges, and chromosome fragments at anaphase and telophase of meiosis I and II (Colas et al., 2019; Szurman-Zubrzycka et al., 2019).
In addition to its roles in meiotic homologous recombination and DNA repair, DMC1 plays a role in abiotic stress responses. High temperature in wheat lowers CO formation, which is particularly obvious in Tadmc1 mutants, demonstrating that the meiotic gene TaDMC1 is responsible for maintaining proper crossover creation under high temperature (Draeger et al., 2023). Studies have also discovered that DMC1 maintains seed germination under salt stress by modulating antioxidant enzyme activity and DNA repair gene expression (Wang et al., 2023). In A.thaliana, the number of DMC1 foci reduces under heat stress, with no significant change in expression level, although RAD51 expression declines (Ning et al., 2021). This suggests that high temperatures may limit DMC1 accumulation by altering DMC1 affinity for chromatin via RAD51 (Cloud et al., 2012; Kurzbauer et al., 2012). The decreased number of DMC1 foci could also be a result of impaired DSB generation and/or unstable chromosomal axis (Sanchez-Moran et al., 2007; Ferdous et al., 2012). Unlike lilium, LIM15 (a DMC1 homolog) is promptly triggered in anthers at 40°C with dramatically elevated expression levels (Ahn et al., 2025). These discrepancies could be due to species-specific responses to heat stress; for example, dicotyledonous plants like A.thaliana may adapt to high temperatures by inhibiting DSBs rather than meiotic repair pathways. Although our study focused on transcriptional expression, subsequent protein-level studies would provide more information on individual responses.
Based on our findings, we propose a model in which DMC1-overexpressed plants respond to high-temperature stress via antioxidant mechanisms and cell cycle transitions (Figure 8). In this model, high temperatures cause elevated ROS in plants, resulting in a large number of DSBs. Plants engage fundamental antioxidant mechanisms, however due to poor repair effectiveness, unrepaired DSBs accumulate and residual oxidative damage persists, eventually resulting in abortive pollen. The OE-BrDMC1 restores homeostasis by activating antioxidant mechanisms and controlling DSB repair genes and cell cycles, resulting in viable pollen.
A DMC1 model in B.rapa at high temperatures. When plants are subjected to severe temperature stress, they produce a huge amount of ROS, which causes DSBs in the plant. In WT, the fundamental antioxidant repair system is active, but it is unable to properly remove excessive ROS, resulting in ROS accumulation in the plant and inadequate DNA repair, ultimately leading to aberrant pollen formation. In OE-BrDMC1, the highly expressed BrDMC1 not only significantly promotes antioxidant synthesis and efficiently removes ROS, but it also up-regulates the expression of DSB repair genes and cell cycle genes, achieving steady-state regulation of DNA damage repair and the cell cycle and, ultimately, producing fertile pollen.
In conclusion, this study found that DMC1 can positively modulate heat tolerance during meiosis. In natural environments, high temperatures are frequently accompanied by drought stress. Current research focuses primarily on single-temperature stress. In the future, detailed studies on combined stress can be carried out to give a theoretical foundation for crop breeding against abiotic stress.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmad H. Hayat S. Ali M. Liu T. Cheng Z. (2018). The combination of arbuscular mycorrhizal fungi inoculation (glomus versiforme) and 28-homobrassinolide spraying intervals improves growth by enhancing photosynthesis, nutrient absorption, and antioxidant system in cucumber (cucumis sativus L.) under salinity. Ecol. Evol. 8, 5724–5740. doi: 10.1002/ece 3.4112, PMID: 29938088 PMC 6010694 · doi ↗ · pubmed ↗
- 2Ahn Y.-J. Kang J.-Y. Lim K.-B. (2025). Heat stress-induced meiotic restitution and tissue-specific gene responses in the sterile lilium OT hybrid ‘yelloween.’ Sci. Hortic. 342, 114032. doi: 10.1016/j.scienta.2025.114032, PMID: 41783259 · doi ↗
- 3Alexander M. P. (1969). Differential staining of aborted and nonaborted pollen. Stain. Technol. 44, 117–122. doi: 10.3109/10520296909063335, PMID: 4181665 · doi ↗ · pubmed ↗
- 4Aranda J. Bardina C. Beceiro A. Rumbo S. Cabral M. P. BarbéJ. . (2011). Acinetobacter baumannii Rec A protein in repair of DNA damage, antimicrobial resistance, general stress response, and virulence. J. Bacteriol. 193, 3740–3747. doi: 10.1128/JB.00389-11, PMID: 21642465 PMC 3147500 · doi ↗ · pubmed ↗
- 5Bannister L. A. Pezza R. J. Donaldson J. R. de Rooij D. G. Schimenti K. J. Camerini-Otero R. D. . (2007). A dominant, recombination-defective allele of Dmc 1 causing male-specific sterility. Plo S Biol. 5, e 105. doi: 10.1371/journal.pbio.0050105, PMID: 17425408 PMC 1847842 · doi ↗ · pubmed ↗
- 6Bishop D. K. Park D. Xu L. Kleckner N. (1992). DMC 1: A meiosis-specific yeast homolog of E. coli rec A required for recombination, synaptonemal complex formation, and cell cycle progression. Cell 69, 439–456. doi: 10.1016/0092-8674(92)90446-J, PMID: 1581960 · doi ↗ · pubmed ↗
- 7Bokszczanin K. L. Fragkostefanakis S. (2013). Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 4. doi: 10.3389/fpls.2013.00315, PMID: 23986766 PMC 3750488 · doi ↗ · pubmed ↗
- 8Borde V. (2007). The multiple roles of the Mre 11 complex for meiotic recombination. Chromosome Res. 15, 551–563. doi: 10.1007/s 10577-007-1147-9, PMID: 17674145 · doi ↗ · pubmed ↗