Effects of exposure to Rickettsia bellii on reservoir competence of guinea pigs for Rickettsia rickettsii
Lina de Campos Binder, Carlos Eduardo Camargo Fanchini, Herbert Sousa Soares, Talita Beck Strabelli dos Santos, Sueli Akemi Taniwaki, Marcelly Bastos Nassar, Andrea Cristina Fogaça, Maria Carolina de Azevedo Serpa, Matheus Pasini Martins, Marcelo Bahia Labruna

TL;DR
Exposure to Rickettsia bellii in guinea pigs reduced their ability to spread Rickettsia rickettsii, a finding that could explain the uneven spread of the pathogen in tick populations.
Contribution
This study demonstrates for the first time that prior exposure to Rickettsia bellii reduces reservoir competence for Rickettsia rickettsii in guinea pigs.
Findings
Guinea pigs exposed to Rickettsia bellii had a 21% infection rate in ticks, compared to 54% in unexposed groups.
Only one guinea pig in the Rickettsia bellii group died, while most in the unexposed groups died.
Prior Rickettsia bellii exposure significantly lowers Rickettsia rickettsii transmission in ticks.
Abstract
Amblyomma sculptum is the main vector implicated in the transmission of Rickettsia rickettsii in southeastern Brazil, where capybaras are known to be the main amplifying hosts of this pathogen. Interestingly, many areas inhabited by large populations of capybaras and A. sculptum remain free of R. rickettsii, suggesting that other drivers may be involved in the pathogen perpetuation. A. sculptum frequently occurs in sympatry with Amblyomma dubitatum ticks, sharing the same hosts with them. Considering that A. dubitatum ticks are frequently infected with Rickettsia bellii, a possible interference of R. bellii in the reservoir competence of capybaras for R. rickettsii could explain the heterogeneous distribution of R. rickettsii in A. sculptum populations in southeastern Brazil. The present study aimed to evaluate the effect of primary exposure to R. bellii-infected A. dubitatum on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
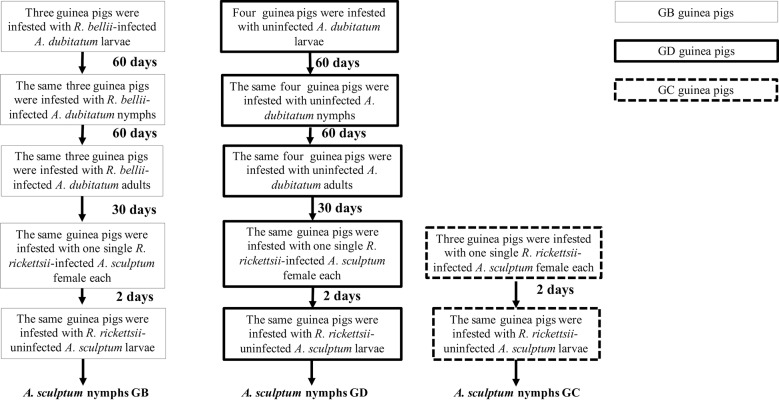 Fig. 1
Fig. 1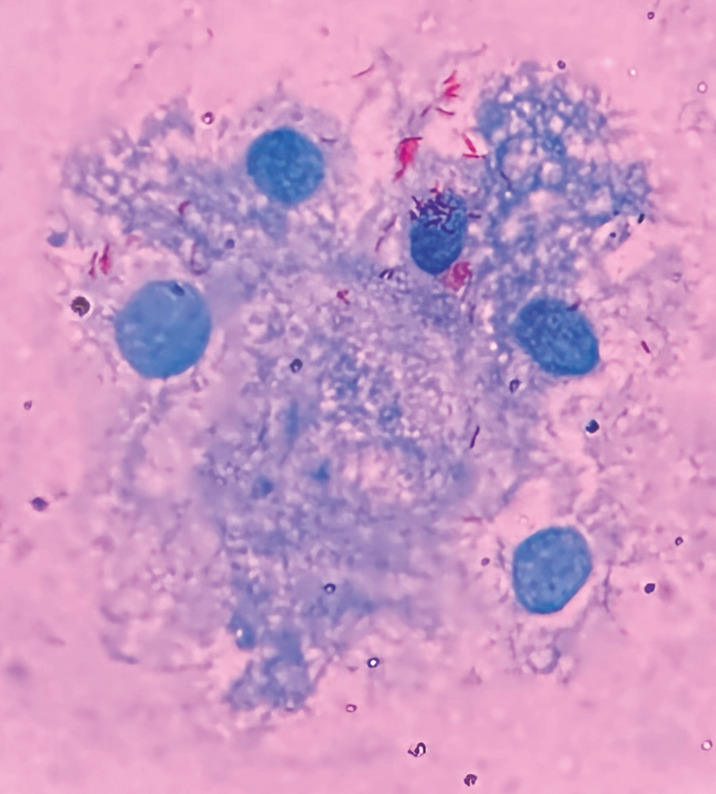 Fig. 2
Fig. 2 Fig. 3
Fig. 3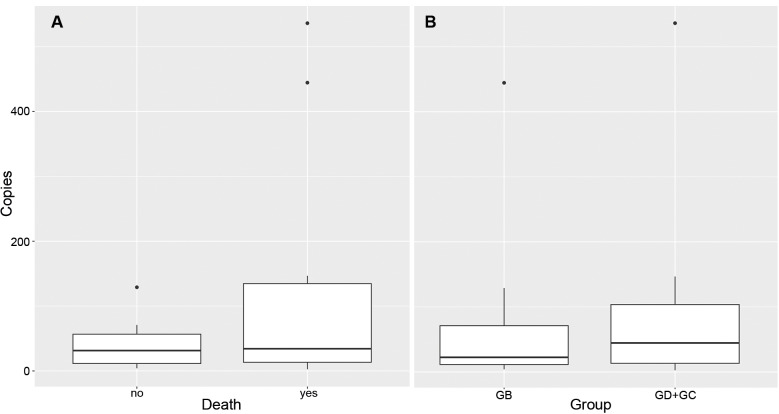 Fig. 4
Fig. 4| Experimental group | Guinea pig ID | Endpoint titre to | Endpoint titer to | Febrile period (DAI) | Death (DAI) |
| No. of bacteria/ng of extracted DNA* |
|---|---|---|---|---|---|---|---|
| GB | C10 | 4096 | 16184 | 7 - 11 | 8, 11 | 10.0 (7.0-12.9) | NO |
| C11 | NA | NA | 7 - 11 | 10, 11 | 235.7 (26.9-444.5) | 13 | |
| C12 | 4096 | 8192 | 6 - 14 | 9, 11, 12, 14 | 35.1 (4.1-129.0) | NO | |
| GD | C16 | 4096 | 4096 | 7 - 13 | 12, 13 | 57.8 (44.6-70.9) | NO |
| C17 | 512 | 1024 | 12 - 18 | 14, 15 | 81.8 (29-134.6) | 21 | |
| C18 | NA | NA | 7 - 10 | 9 | 6.4 | 13 | |
| C19 | NA | NA | 6 - 10 | 8, 10 | 285.2 (34.2-536.2) | 12 | |
| GC | C30 | NA | NA | 5 - 8 | 5, 6, 7, 8 | 42.0 (2.6-146.7) | 10 |
| C31 | <64 | <64 | NO | NO | NO | NO | |
| C32 | NA | NA | 6 - 10 | 7, 9 | 58.45 (13.3-103.6) | 12 |
| Experimental group | Guinea pig ID | Infection rate (No. of infected nymphs/No. of tested nymphs)*+ |
|---|---|---|
| GB | C10 | NA |
| C11 | NA | |
| C12 | 21% (17/80) | |
| Total | 21% (17/80)a | |
| GD | C16 | 0% (0/4) |
| C17 | NA | |
| C18 | 41% (7/17) | |
| C19 | 55% (42/76) | |
| Total | 50% (49/97)b | |
| GC | C30 | NA |
| C32 | 79% (11/14) | |
| Total | 79% (11/14)b | |
| GD + GC | Total | 54% (60/111)b |
- —FAPESP
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · T-cell and Retrovirus Studies · Trypanosoma species research and implications
INTRODUCTION
Rocky Mountain spotted fever, also known as Brazilian spotted fever (BSF), is caused by the bacterium Rickettsia rickettsii. BSF is the most relevant tick-borne disease in Brazil, where Amblyomma sculptum is the main vector implicated in the transmission of this pathogen to humans.1 2 3 4
In spite of being a competent vector, A. sculptum is partially refractory to R. rickettsii infection and less efficient than other tick species in maintaining R. rickettsii by transstadial perpetuation and transovarial transmission.5 6 7 These features suggest that vertical transmission alone is insufficient to perpetuate R. rickettsii infection over successive tick generations; thus, horizontal transmission through an amplifying vertebrate host is necessary.8 Indeed, mathematical models have predicted that after a 90% reduction in amplifying hosts birth rate, R. rickettsii infection tends to disappear within two years from A. sculptum populations.9
Capybaras (Hydrochoerus hydrochaeris) are efficient hosts for all A. sculptum feeding stages and act as amplifying hosts of R. rickettsii for this tick species.10 11 Since the 1980’s, this rodent population has been experiencing rapid growth in southeastern Brazil,12 13 14 and this has been associated with spatial expansion of BSF occurrence as well as higher incidence rates.1 8 15 However, it is noteworthy that many areas with high capybara and A. sculptum densities remain free of R. rickettsii in southeastern Brazil,16 17 18 indicating that other factors may be involved in the maintenance of the pathogen in vector populations.
Besides being hosts for A. sculptum, capybaras are also primary hosts of Amblyomma dubitatum 19 18 and these two ticks occur in sympatry in many areas across southeastern Brazil.3 18 20 21 22 23 Interestingly, natural infection of A. dubitatum with R. rickettsii has never been reported, while different studies have described Rickettsia bellii infection in A. dubitatum populations, often presenting high infection frequencies.21 24 25 26 27 28 29 30 31 32 33 34
Amblyomma sculptum and A. dubitatum were the only ticks found to infest capybaras in anthropised landscapes in southeastern Brazil.18 35 Nevertheless, Luz et al.18 noticed that A. sculptum greatly outnumbered A. dubitatum in BSF-endemic areas, while in BSF-non-endemic areas, abundances of these two species were similar, suggesting a negative correlation between R. rickettsii infection and higher A. dubitatum/A. sculptum abundance ratio.
Spotted fever group (SFG) Rickettsia are antigenically related bacteria and exhibit strong cross-reactivity in serological tests. In spite of not being part of SFG, R. bellii also presents antigenic cross-reactivity with SFG Rickettsia.36 Considering cross-reactivity between these two bacteria, it would be expected that host immune responses may affect the natural dynamics of these antigenically related organisms. Indeed, several experimental studies have shown that infection with a Rickettsia species elicits immune response in hosts reducing or preventing illness and fatal outcomes by a second infection with a pathogenic Rickettsia, besides reducing transmission of the pathogen to susceptible ticks.37 38 39 40 41
Since A. sculptum is not highly efficient in perpetuating R. rickettsii infection across generations, amplifying hosts are essential for the pathogen maintenance in vector populations. Therefore, it is plausible that reducing the reservoir competence (ability to transmit an infection to arthropod vectors) of capybaras would affect R. rickettsii perpetuation. Considering that capybaras are primary hosts for A. dubitatum, and these ticks are frequently infected with R. bellii, one can hypothesise that exposure of capybaras to R. bellii elicit an immunological response capable of reducing the reservoir competence of capybaras for R. rickettsii. Thereby, this study aimed to evaluate if exposure of amplifying hosts to R. bellii-infected A. dubitatum ticks could reduce or preclude R. rickettsii transmission to A. sculptum ticks, using guinea pigs as experimental models.
MATERIALS AND METHODS
Animals and ticks
Animals and ticks - All animals used along the study were kept in the same room, with temperature (23ºC), ventilation and photoperiod (12/12) control. They were maintained in individual cages during the whole experiment, preventing physical contact between them and were fed with a commercial guinea pig pellet diet, fresh grass and water ad libitum. Ticks were held in an incubator at 25ºC, 85-90% relative humidity, and total scotophase for off-host developmental stages.
Ticks used in this study derived from colonies maintained in our laboratory by feeding on tick-naïve guinea pigs (Cavia porcellus) or tick-naïve rabbits (Oryctolagus cuniculus). All the colonies derived from host-seeking adult ticks collected in 2022 in São Paulo municipality, State of São Paulo (-23.557, -46.718). Two colonies of A. dubitatum were used, one naturally infected with R. bellii and one naturally uninfected, as previously reported.42 For A. sculptum, also two colonies were used, however, one infected with R. rickettsii and one uninfected. For establishing the R. rickettsii-infected colony, guinea pig organs (liver and lung) infected with R. rickettsii strain Itu3 stored at -80ºC were thawed at room temperature, crushed with sterile phosphate buffered saline (PBS), and the resultant homogenate was inoculated intraperitoneally in guinea pigs, as previously described.5 7 One day after inoculation, A. sculptum larvae were allowed to feed on guinea pigs inside feeding chambers (white cotton sleeves) glued to the animals shaved back. Sleeves were daily opened and detached engorged larvae were removed and taken to an incubator for ecdysis. Thereafter, ticks were fed on tick-naïve guinea pigs.
In order to assess rickettsial infection status of the colonies, flat nymphs and adults were individually tested by different conventional polymerase chain reaction (PCR) protocols. To that end, genomic DNA was extracted from the ticks using the phenol-chloroform-guanidine-isothiocyanate method, as described elsewhere.43 All samples were initially submitted to a PCR reaction using primers CS-78 and CS-323, targeting a 401 bp fragment of the citrate synthase gene (gltA), common in all representatives of the genus Rickettsia.24 DNA samples positive for infection were further tested by two additional protocols: the first one using primers Rr190.7044 and Rr190.701,45 which amplifies a 631 bp fragment of the rickettsial 190-kDa outer membrane protein gene (ompA) in most of the SFG rickettsiae; and the second one, a R. bellii-specific assay, targeting a 338 bp fragment of the gltA gene, as previously described.46
From the R. bellii-infected colony, 100% (48/48) of the A. dubitatum individuals were shown to be positive for R. bellii and negative for other Rickettsia species, while 100% (50/50) of uninfected A. dubitatum ticks and 100% (33/33) of uninfected A. sculptum ticks were negative for any rickettsial infection. On the other hand, 67% (22/33) of A. sculptum ticks exposed to R. rickettsii in the larval stage were shown to be infected. Ticks from these four colonies were used in the present study.
Tick infestations
Tick infestations - Infestation design is summarised in Fig. 1. Ten tick-naïve guinea pigs were allocated in three experimental groups: three guinea pigs previously exposed to R. bellii-infected A. dubitatum ticks (group GB); four guinea pigs previously exposed to R. bellii-uninfected A. dubitatum ticks (group GD); and three guinea pigs that were not exposed to A. dubitatum ticks (group GC). Guinea pigs from GB and GD groups were initially exposed to R. bellii-infected and -uninfected larvae, respectively. Approximately 60 and 120 days after the larval infestation, the same animals were exposed to A. dubitatum nymphs and adults, respectively. Guinea pigs from group GC were kept free of ticks along this period.
Diagram illustrating experimental procedures with guinea pigs.Groups GB and GD were composed by three and four guinea pigs, respectively, each of them infested by Amblyomma dubitatum larvae, nymphs and adults at 60-day intervals. GB served as hosts for A. dubitatum ticks infected by Rickettsia bellii, whereas GD served as hosts for uninfected A. dubitatum. Thirty days later, all animals were exposed to Rickettsia rickettsii-infected Amblyomma sculptum females (one infected female per guinea pig), and two days later, by uninfected larvae of A. sculptum. The GC group, composed of three guinea pigs never exposed to A. dubitatum ticks, were submitted to the same protocols of infestation by A. sculptum females and larvae.
Thirty days after infestation with A. dubitatum adults (GB and GD), guinea pigs from all three groups (GB, GD and GC) were infested by A. sculptum ticks. Two feeding chambers were glued to each animal’s back. In the first chamber, one single R. rickettsii-infected A. sculptum female was allowed to feed for two days. In order to infest guinea pigs only with R. rickettsii-infected females, previous to the infestation, females had one leg cut off and submitted to DNA extraction by the boiling method.26 The respective DNA samples were tested by a quantitative polymerase chain reaction (qPCR) protocol (described below). Only females that yielded a positive result in this qPCR protocol were allowed to feed. After two days of feeding, females were detached from the host and submitted to the same qPCR protocol in order to confirm the infection status. One day after the removal of females, uninfected A. sculptum larvae (approximately 500 - 1000 larvae/animal) were released in the second chamber and were allowed to feed until repletion. Feeding chambers were opened daily and engorged detached larvae were taken to an incubator for ecdysis. All the procedures were done simultaneously for the three experimental groups.
All animals were monitored for 21 days after each infestation and had their rectal temperature daily measured. Guinea pigs were considered febrile when rectal temperature was > 39.5ºC.47 In all infestations, approximately 500 µL of blood was collected from the lateral saphenous vein or from the tarsal vein from each guinea pig48 0 and 21 day(s) after infestations (DAI). Additionally, febrile animals had blood samples collected daily during the febrile period. Blood samples collected at 0 and 21 DAI were centrifuged, and the respective serum samples were tested for the presence of antibodies against R. rickettsii and R. bellii, as described below, while blood samples collected during the febrile period were stored at -20ºC for molecular analysis, as described below.
Serological analyses
Serological analyses - Guinea pig serum samples were individually tested by the indirect immunofluorescence assay (IFA) using a commercial fluorescein isothiocyanate-labelled goat anti-guinea pig IgG (Sigma Diagnostics, Saint Louis, MO) diluted 1:400 and crude antigens derived from R. rickettsii strain Taiaçu49 and R. bellii strain Mogi.49 Each serum was diluted in two-fold increments with PBS from 1:64 to endpoint titre, as previously described.16 In each slide, a serum previously shown to be non-reactive (negative control) and a known reactive serum (positive control) from a previous study in our laboratory7 were tested at the 1:64 dilution.
Rickettsial detection
Rickettsial detection - After the last infestation with A. dubitatum ticks, two males from the R. bellii-infected colony, previous fed on each guinea pig from group GB, were dissected and had their salivary glands removed and washed in PBS, as described by Benatti et al.50 One fragment of the salivary gland was imprinted in a glass slide by gently touching the tissue to the slide in a vertical up-and-down motion. The imprints were left to air dry and then were stained by the Giménez staining.51 Imprints were evaluated under an optical microscope at a 1000x magnification. The second salivary gland fragment was submitted to DNA extraction using the phenol-chloroform-guanidine-isothiocyanate method.43 DNA samples were tested by a R. bellii specific PCR assay.46
DNA samples from A. sculptum females used in the infestations were tested by a Taqman qPCR assay targeting a fragment of the citrate synthase gene (gltA), using the primers CS-552 and CS-624 and an internal fluorogenic probe as described by Soares et al.6 Each reaction had a positive (Rickettsia vini DNA) and a negative control (molecular-grade water).
Blood samples from febrile guinea pigs were submitted to DNA extraction using the DNeasy Blood & Tissue kit (Qiagen, Germany), according to manufacturer instructions. For R. rickettsii detection, a probe-based qPCR assay was performed to amplify a gene encoding hypothetical protein A1G_04230 using primers RRi6_F and RRi6_R and an internal fluorogenic probe described by Kato et al.53 Reactions were performed in a 7500 Real Time PCR Systems apparatus (Applied BioSystems, Foster City, CA). For each reaction, 2.5 μL of DNA template was added to 10 μL of the GoTaq Probe qPCR Master Mix (Promega, Madison, WI), 1.8 μL of each primer at 10 μM, 0.8 μL of probe at 10 μM and molecular-grade water to a final volume of 20 μL. qPCR cycling conditions were 1 cycle of 2 min at 95ºC, followed by 45 cycles of 15 s at 95ºC, 20 s at 55ºC and 30 s at 72ºC.
For quantification of R. rickettsii in guinea pig blood samples, plasmids with RRi6_F and RRi6_R amplicons were used to generate a ten-fold dilution standard curve from 10^2^ to 10^6^ copies, which resulted in a linear equation (y = -3.464x + 45.06) with an R^2^ value = 0.998 and efficiency of 94.383%. The quantification limit of the assay was defined as 100 DNA copies of R. rickettsii. Samples and standard curve were run in duplicate. All qPCR results are expressed in bacterial copies per ng of extracted DNA, determined by dividing the calculated copy number per µL by the total DNA concentration (ng/µL) of each sample [quantified in a spectrophotometer (Nanodrop 2000, Thermo Fisher Scientific)].
Approximately 20 days after ecdysis, A. sculptum nymphs (fed in the larval stage on febrile guinea pigs) from all the experimental groups underwent individual DNA extraction by the boiling method.26 DNA samples were initially submitted to a PCR protocol using primers CS-78 and CS-323 targeting a fragment of the citrate synthase gene (gltA), common in all representatives of the genus Rickettsia.24 Samples yielding positive results by this first PCR protocol were further tested by a second assay using primers Rr190.7044 and Rr190.701,45 which amplifies a fragment of the rickettsial 190-kDa outer membrane protein gene (ompA) present in most of the SFG rickettsiae. Nymphs were considered to be infected by R. rickettsii only when shown to be positive for both PCR assays. Each reaction had a positive (Rickettsia vini DNA) and a negative control (molecular-grade water).
In order to validate DNA extraction, samples that yielded negative results in the rickettsial detection assays were tested by one additional PCR protocol. Tick samples underwent an assay targeting a fragment of the tick mitochondrial 16S rRNA according to Mangold et al.,54 while blood samples were tested by a PCR targeting a 359 bp fragment of the mitochondrial cytochrome b (cyt b) gene from vertebrates, as described elsewhere.55 Samples with negative results in these PCR protocols were excluded from the study.
Survey for R. bellii in tick saliva
Survey for R. bellii in tick saliva - Serological analyses showed that no guinea pig from group GB had detectable antibodies against R. bellii after exposure to infected A. dubitatum ticks (results are described below). Since R. bellii was detected in salivary glands from male A. dubitatum ticks fed on all animals from group GB (results are described below), we decided to investigate if A. dubitatum ticks secrete R. bellii in the saliva. For this purpose, A. dubitatum ticks were collected at the exact same locality, where ticks that originated the colonies used in the study were collected.
In order to use only R. bellii-infected females, they had one leg cut off and submitted to DNA extraction by the boiling method.26 The respective DNA samples were tested by a qPCR protocol which amplifies a 338 bp fragment of the gltA gene, using primers described by Szabó et al.46 and an internal fluorogenic probe, as described by Hecht et al.56 Only five females, those that yielded a positive result in this qPCR protocol, were allowed to feed on a tick-naïve guinea pig along with five A. dubitatum males. Females were allowed to feed for six to nine days, until they were partially engorged.
After partial engorgement, females were detached from the host, washed three times in ultrapure water and dried with filter paper. Each female was inoculated with pilocarpine (50mg/ml in 0.7M NaCL solution) using a 12.7 mm x 0.33 mm needle, as described by Esteves et al.57 Saliva was harvested every five to ten minutes until the female stopped to salivate. One microliter of each saliva sample was dripped onto a slide, left to air dry and then was stained by the Giménez staining.52 Slides were evaluated under an optical microscope at a 1000x magnification. The remaining saliva samples were stored at -20ºC for molecular analyses. Saliva samples were diluted 1: 2 in Tris-EDTA buffer solution and underwent DNA extraction by the boiling method.26 For R. bellii detection, a specific qPCR assay was performed, as described elsewhere.56
Statistical analyses
Statistical analyses - Rickettsia rickettsii concentration in blood samples were compared between different experimental groups (GB, GD and GC) by Kruskal-Wallis test and between animals that died and survived by Mann-Whitney test. Proportion of blood samples yielding positive results in qPCR assays for R. rickettsii detection as well as R. rickettsii infection rate among A. sculptum nymphs were compared between the different experimental groups (GB, GD and GC) using Fisher’s exact test. Analyses were performed using R58 and a 5% significance level was assumed.
Ethics statement
Ethics statement - This study has been approved by the Ethic Committee on Animal Use of the Faculty of Veterinary Medicine and Animal Science, University of São Paulo (CEUA No. 5319211021).
RESULTS
During all the infestations with A. dubitatum ticks, no guinea pig had fever or any other clinical manifestation. Further, no guinea pig seroconverted to R. bellii after any of the three consecutive exposures to R. bellii-infected A. dubitatum ticks (larvae, nymphs and adults). Despite the absence of seroconversion, R. bellii DNA was detected in salivary glands of A. dubitatum adult ticks that had fed on all guinea pigs from group GB and it was possible to visualise bacteria morphologically compatible with R. bellii in their salivary glands (Fig. 2). In addition, it was possible to visualise bacteria morphologically compatible with R. bellii (Fig. 3) and to detect R. bellii DNA in saliva samples from all tested females. These coupled results suggest that A. dubitatum are able to transmit R. bellii to hosts and that animals from the GB group were in fact exposed to R. bellii in spite of the absence of a humoral host response.
Optical microscopy analysis of the salivary glands of Amblyomma dubitatum ticks infected with Rickettsia bellii.Giménez staining, 1000x original magnification.
Optical microscopy analysis of saliva samples of Amblyomma dubitatum ticks infected with Rickettsia bellii.Giménez staining, 1000x original magnification.
After exposure to a single R. rickettsii-infected tick, all the guinea pigs [except for one guinea pig (C31) from group GC] presented fever and other clinical manifestations compatible with R. rickettsii infection, which started between five and 12 DAI. One guinea pig from group GB, three from group GD and two from group GC died 12 to 21 DAI and five to nine days after fever onset (Table I).
It was possible to detect R. rickettsii DNA in at least one blood sample from all animals, except for guinea pig C31. Noteworthy, R. rickettsii was detected only in one to four timepoints during the whole febrile period (Table I). Proportion of febrile days in which it was possible to detect R. rickettsii in blood samples was not different among experimental groups. Bacterial loads were slightly lower among animals from group GB and among animals that survived to infection, but no statistical significance was detected (Fig. 4).
Rickettsia rickettsii load in blood samples from guinea pigs.(A) R. rickettsii load in blood samples (No. of R. rickettsii genome copies/ng of extracted DNA) from guinea pigs that died after infection and guinea pigs that survived infection with R. rickettsii. (B) R. rickettsii load in blood samples (No. of R. rickettsii genome copies/ng of extracted DNA) from guinea pigs infested with R. rickettsii-infected Amblyomma sculptum females. GB: guinea pigs previously exposed to Rickettsia bellii-infected Amblyomma dubitatum ticks; GD: guinea pigs previously exposed to R. bellii-uninfected A. dubitatum ticks; and GC: guinea pigs not exposed to A. dubitatum ticks.
All guinea pigs that had blood samples collected at 21 DAI seroconverted to R. rickettsii and R. bellii, generally with higher titres for R. rickettsii (Table I). Guinea pig C31 (group GC), which was the only one to remain afebrile, was also the only one that did not seroconvert, indicating that this animal was in fact not exposed to the pathogen. For this reason, it was not considered in further analyses.
From the A. sculptum nymphs that fed as larvae on guinea pigs during their febrile period, 191 nymphs underwent molecular analyses, which showed that 77 (40%) of them were infected by R. rickettsii. When evaluating the different experimental groups separately, ticks fed on GB guinea pigs showed 21% (17/80) infection rate, while ticks from groups GD and GC had infection rates of 50% (49/97) and 79% (11/14), respectively. When analysing ticks from GD and GC together, an infection rate of 54% (60/111) was observed. R. rickettsii infection rate in GB ticks was significantly lower than the infection rates recorded on the GD and GC groups (both when analysed separately or together) (Table II).
DISCUSSION
In the present study, the reservoir competence of guinea pigs for R. rickettsii was evaluated by comparing the infection rates of A. sculptum nymphs after feeding as larvae on three groups of R. rickettsii-infected guinea pigs: one group previously exposed to R. bellii-infected A. dubitatum ticks (GB), one group previously exposed to uninfected A. dubitatum ticks (GD), and one group never previously exposed to A. dubitatum (GC). As the GB A. sculptum nymphs had a significantly lower R. rickettsii-infection rate than the GD and GC nymphs, it was demonstrated under experimental conditions that exposure to R. bellii-infected A. dubitatum ticks reduced the reservoir competence of guinea pigs for R. rickettsii.
After being exposed to R. rickettsii-infected females, all guinea pigs (except for C31) presented clinical illness and seroconverted to R. rickettsii or died during acute infection. Although it was possible to detect R. rickettsii DNA in blood samples from all animals, it was not possible to detect the pathogen in all blood samples collected daily during the febrile period. R. rickettsii infects endothelial cells and has a very low concentration in the bloodstream, which makes blood qPCR an unsuitable assay for R. rickettsii diagnosis, as reported by several other studies, in which blood samples from infected animals yielded negative results in blood qPCR assays.10 11 59 60
Santos et al.61 showed that among BSF human cases, R. rickettsii DNA was detected in serum samples from all patients that had fatal outcomes, while in nonfatal cases, qPCR yielded positive results only for one third of the samples. Fatal outcomes are probably related to a vaster endothelial injury leading to severe vascular alterations and therefore higher R. rickettsii concentration in the blood. Interestingly, herein no statistical difference was observed in R. rickettsii load between animals that died or survived infection, even though a slightly higher load was recorded in animals that had fatal outcomes.
Even though guinea pigs from group GB showed a slightly lower R. rickettsii load, no statistical difference was observed among different experimental groups, precluding association between qPCR results and reservoir competence, assessed by the number of ticks becoming infected after blood feeding. This could be due to our small sample of guinea pigs or to an unsuitability of blood qPCR to assess hosts infectivity to ticks. In this regard it is important to keep in mind that molecular assays reflect one single moment in time when the respective blood sample was collected, while ticks remain attached ingesting blood for days. Further, it is not possible to affirm that R. rickettsii DNA detected by qPCR corresponds to viable bacteria, precluding inferences on infectivity.
Guinea pig C31 was not exposed to R. rickettsii as verified by the absence of seroconversion. This was unexpected, since the A. sculptum female fed on this guinea pig was shown to be infected. One possible reason for the absence of R. rickettsii transmission is the lack of time for the female to transmit the agent. Magalhães62 demonstrated that A. sculptum ticks may need 36 h to efficiently transmit R. rickettsii to a susceptible host. In our study, feeding chambers were inspected two times a day until the female was seen to be attached to the host. Forty-eight hours after attachment, females were removed from the chambers. Nevertheless, during this 48 h period, feeding chambers were not opened, being not possible to state that ticks were continuously attached to the host during this entire period.
Despite being repeatedly exposed to R. bellii, which was confirmed by detection of this bacterium in salivary glands of ticks fed on all animals, no guinea pig from group GB seroconverted to R. bellii prior to exposure to R. rickettsii. This was also noticed by other studies29 42 and suggest that R. bellii exposure through tick bite is not able to elicit a humoral response in guinea pigs. These results are quite surprising, since many serosurveys have detected different wild and domestic animals with IgG titers for R. bellii higher than for other Rickettsia species, suggesting a homologous reaction to R. bellii.18 63 64 One hypothesis is that these seropositive animals were exposed to R. bellii through a different infection route (e.g., ingestion of infected ticks), which enabled a humoral response due to exposure to a higher bacterial load. Another hypothesis is that antibodies detected in these studies do not derive from a previous exposure to R. bellii, but to another agent that cross-reacts with this bacterium.
In the study by Pacheco et al.,16 guinea pigs were inoculated intraperitoneally with different species of Rickettsia (R. bellii, Rickettsia canadensis, Rickettsia monteiroi and R. rickettsii). While guinea pigs inoculated with R. bellii had titres to the homologous antigen ranging from 256 to 1024, animals inoculated with other Rickettsia showed much higher homologous titres, ranging from 2048 to 32768. This low humoral response to R. bellii, even after exposure to a high number of bacteria via intraperitoneal inoculation, may indicate a low ability of R. bellii to elicit antibodies production.
Further studies should be performed to elucidate these findings and to ascertain whether IFA is a suitable assay for determining a previous exposure to R. bellii via tick bite, or not.
Since Rickettsia spp. are obligate intracellular bacteria, cell-mediated immunity, instead of humoral immunity, is expected to be the main immune response involved in their clearance, which is corroborated by studies that evaluated passive transfer of T cells in infections by different species of Rickettsia.65 66 67 68 Further, it was observed that mice lacking CD8+ T cells showed reduced survival and increased bacterial load after infection with Rickettsia conorii and enhanced susceptibility to fatal outcomes after infection with Rickettsia australis.67 68 Thus, absence of detectable IgG against R. bellii does not necessarily mean that no effective adaptive immunity was induced. On the other hand, there is evidences that humoral response, specially antibodies to epitopes of OmpB, also play a relevant role in reinfections by Rickettsia, reducing the severity of the disease.69 70 71 This could suggest that the adaptive immunity elicited by R. bellii may not be as effective as immune responses induced by SFG Rickettsia in protecting hosts to a second infection with a more pathogenic Rickettsia.
Indeed, exposure to R. bellii did not prevent clinical illness or fatal outcomes after infection by R. rickettsii, however, the lethality rate was lower in group GB (1/3) than in the other experimental groups (5/6). These results are different from other studies in which exposure to different Rickettsia fully prevented death due to R. rickettsii infection.37 38 39 41 These studies, however, used Rickettsia species phylogenetically closer to R. rickettsii than R. bellii, possibly inducing a stronger cross-reaction to R. rickettsii than the one observed in this study and, therefore, being more effective in protecting the hosts from severe illness and death.
Even though exposure to R. bellii-infected ticks did not fully protect animals from illness nor prevented A. sculptum larvae from being infected by R. rickettsii, guinea pigs from group GB (previously exposed to R. bellii-infected ticks) were less efficient in infecting A. sculptum larvae, when compared to guinea pigs from the other experimental groups (not previously exposed to R. bellii). Considering that A. sculptum is not very efficient in maintaining R. rickettsii across successive generations, reduction in host reservoir competence would probably have a great impact on the perpetuation of this pathogen. Nevertheless, since A. sculptum larvae were fed on a few animals, it is not possible to completely rule out the hypothesis that the difference in reservoir competence between experimental groups is, in part, due to natural variability among individuals rather than to previous exposure to R. bellii.
Binder et al.42 showed that A. dubitatum ticks were competent vectors for R. rickettsii and also able to transmit the pathogen transovarially, suggesting that these ticks may be important in the enzootic cycle of R. rickettsii. On the other hand, the same study noticed that A. dubitatum females coinfected by R. rickettsii and R. bellii generated a low number of viable larvae. Taken together, these results suggest that R. bellii could negatively impact the perpetuation of R. rickettsii among A. sculptum populations by two mechanisms: first, by reducing capybaras’ reservoir competence for R. rickettsii, and second, by minimising the role of A. dubitatum in R. rickettsii enzootic cycle due to the negative impact of superinfection on tick viability with a consequent reduction in the population of R. rickettsii-infected A. dubitatum ticks. These mechanisms could explain the association between a higher A. dubitatum/A. sculptum abundance ratio and the absence of R. rickettsii noticed by Luz et al.,18 being important drivers associated to spatial heterogeneity of R. rickettsii-infected A. sculptum populations within southeastern Brazil.
In conclusion, our results demonstrated that prior exposure of guinea pigs to R. bellii reduced their reservoir competence for R. rickettsii. Under natural conditions, this kind of interference among capybaras may significantly influence the long-term persistence of R. rickettsii in A. sculptum populations, a condition yet to be confirmed in the capybara model. Therefore, R. bellii may represent a promising strategy for disrupting the amplifying role of capybaras in the R. rickettsii transmission cycle.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pinter A França AC Souza CE Sabbo C Nascimento EMM Santos FCP Febre maculosa brasileira Bepa 20118 Suppl. 1131
- 2Pinter A Sabbo C Leite R Spinola R Angerami R Informe técnico sobre febre maculosa brasileira Bepa 2021185478
- 3Krawczak FS Nieri-Bastos FA Nunes FP Soares JF Moraes-Filho J Labruna MB Rickettsial infection in Amblyomma cajennense ticks and capybaras (Hydrochoerus hydrochaeris) in a Brazilian spotted fever-endemic area Parasit Vectors 20147172438767410.1186/1756-3305-7-7PMC 3892071 · doi ↗ · pubmed ↗
- 4Labruna MB Krawczak FS Gerardi M Binder LC Barbieri ARM Paz GF Isolation of Rickettsia rickettsii from the tick Amblyomma sculptum from a Brazilian spotted fever-endemic area in the Pampulha Lake region, southeastern Brazil Vet Parasitol Reg Stud Reports 2017882853101464410.1016/j.vprsr.2017.02.007 · doi ↗ · pubmed ↗
- 5Labruna MB Ogrzewalska M Martins TF Pinter A Horta MC Comparative susceptibility of larval stages of Amblyomma aureolatum, Amblyomma cajennense, and Rhipicephalus sanguineus to infection by Rickettsia rickettsii J Med Entomol 200845115611591905864210.1603/0022-2585(2008)45[1156:csolso]2.0.co;2 · doi ↗ · pubmed ↗
- 6Soares JF Soares HS Barbieri AM Labruna MB Experimental infection of the tick Amblyomma cajennense, Cayenne tick, with Rickettsia rickettsii, the agent of Rocky Mountain spotted fever Med Vet Entomol 2012261391512200786910.1111/j.1365-2915.2011.00982.x · doi ↗ · pubmed ↗
- 7Gerardi M Ramírez-Hernández A Binder LC Krawczak FS Gregori F Labruna MB Comparative susceptibility of different populations of Amblyomma sculptum to Rickettsia rickettsii Front Physiol 2019106536533119135010.3389/fphys.2019.00653 PMC 6546895 · doi ↗ · pubmed ↗
- 8Labruna MB Moreira J Ferraz K Herrera E Macdonald D Brazilian spotted fever: the role of capybaras Capybara Springer New York 2013371383