Documenting coral spawning in East Africa: first in situ observations from Zanzibar for three reef-building species
Ashlee Lillis, Narriman Jiddawi

TL;DR
This study documents the first in situ coral spawning observations in Zanzibar for three reef-building coral species, providing insights into their reproductive patterns.
Contribution
The study provides the first in situ spawning records for three coral species in the Western Indian Ocean region.
Findings
Galaxea astreata spawned on the second night after the full moon in November and December.
Favites pentagona spawned on the fourth night in November, while Platygyra daedalea spawned on the fourth night in December and January.
Spawning behavior varied by species and individual, including gamete setting, release windows, and split spawning.
Abstract
Broadcast coral spawning is a vital reproductive event for many reef-building species and is essential to the resilience of coral reef ecosystems. Understanding spawning dynamics is key to assessing the reproductive health and resilience of these ecosystems. While extensively documented in the Indo-Pacific and Caribbean, coral spawning remains understudied in the Western Indian Ocean (WIO), a region facing rapid environmental change and increasing coral decline. This study presents the first in situ coral spawning records from Zanzibar for three Scleractinian species: Galaxea astreata, Favites pentagona, and Platygyra daedalea. Observations were made during monthly nighttime monitoring dives from November 2024 to February 2025, timed with lunar and solar cycles linked to peak spawning in other regions. Results showed evidence of species-specific spawning timing: an individual G.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
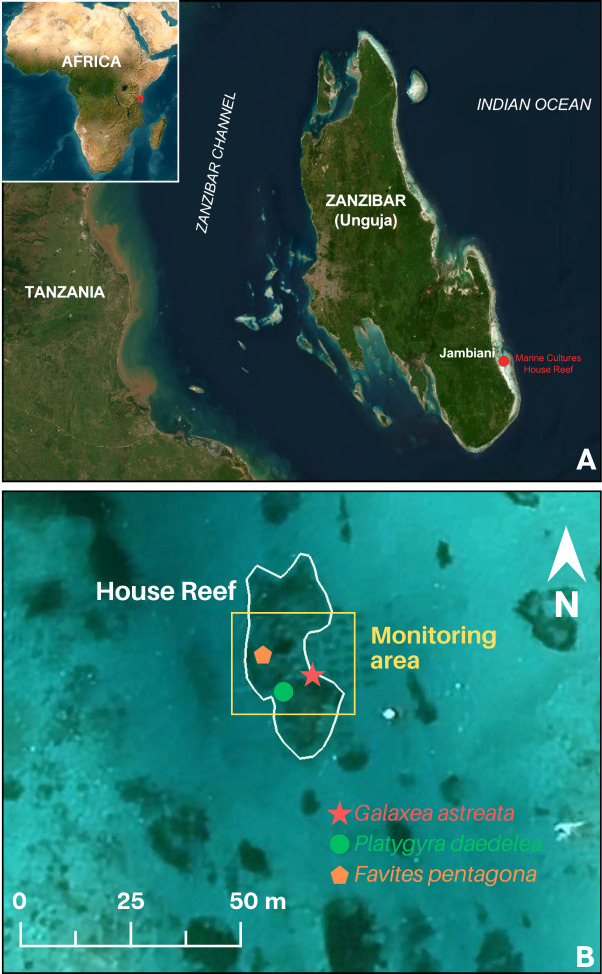 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoral and Marine Ecosystems Studies · Marine and fisheries research · Crustacean biology and ecology
Introduction
Coral reef ecosystems are among the most biodiverse on Earth and provide innumerable ecological services to coastal and island populations, from fish nursery habitat to shoreline protection (Bellwood et al., 2004; Burke & Spalding, 2022; Santavy et al., 2021). Coastal ecosystems, and coral reefs in particular, are in rapid decline due to myriad stressors both at local (e.g., coastal development, run-off, sedimentation, water quality, destructive fishing) and regional or global scales (e.g., climate change, invasive species). Coral reef degradation is predicted to continue unless these threats can be managed and recovery can be facilitated (Bellwood et al., 2004). The foundation of these critical ecosystems are reef-building stony corals, sessile invertebrate animals that live in symbiosis with photosynthetic organisms called zooxanthellae and produce a calcium carbonate skeleton.
East African coral reefs are an important yet understudied sub-region for coral diversity and connectivity (Mbije et al., 2002). In Zanzibar coral reefs are also a highly valuable coastal resource, increasingly experiencing widespread degradation due to various human stressors (Ussi et al., 2024). Though no comprehensive census has been completed for Zanzibar, there are currently estimated between 150–175 hard coral species (Johnstone, Muhando & Francis, 1998; Khamis, Kalliola & Käyhkö, 2017) and despite declines over the past 30 years healthy coral formations persist, especially in more remote areas protected from human impacts (Bravo et al., 2021; Muhando, 2009; Ussi et al., 2024). Central to continued persistence and the resilience potential of reefs is successful coral sexual reproduction. A substantial number of reef-building hard coral species are broadcast spawners (Gleason & Hofmann, 2011), simultaneously releasing eggs and sperm into the water column typically only once or twice per year, where the eggs are fertilized in surface waters and develop through a larval stage before settling and attaching to the seafloor (Babcock et al., 1986; Baird et al., 2021; Harrison & Wallace, 1990). These synchronized spawning events are known to occur across vast expanses of reef. Where dense coral populations exist, large surface slicks of coral gametes can be observed at the water surface following a spawning event (Harrison et al., 1984; Jamodiong et al., 2018).
Broadcast spawning is typically tightly synchronized to solar, lunar, and seasonal cycles, and is also related to environmental factors such as water temperature (Lin & Nozawa, 2023), light (Boch et al., 2011; Penland et al., 2004), wind (Van Woesik, 2010), and precipitation (Mendes & Woodley, 2002). For example, a given species may consistently spawn annually on the 10th night after the full moon in May between 45–60 min after sunset (Baird, Guest & Willis, 2009; Banaszak et al., 2023). Because gametes only remain viable for external fertilization over a few hours, it is critical to reproductive success that individuals release gametes in such close coordination. In the most well-studied regions (e.g., the Great Barrier Reef), mass spawning of dozens of species has been observed over just one or two nights in a single year (Baird et al., 2021; Harrison et al., 1984). However, variation in spawning frequency and timing, as well as inter-annual consistency within species, is known to occur across geographies (Lin & Nozawa, 2017; Lin & Nozawa, 2023; Sakai et al., 2020) and there are instances of populations with protracted non-overlapping spawning periods of several months (Mangubhai, Harrison & Obura, 2007; Mangubhai & Harrison, 2008; Mangubhai & Harrison, 2009; Monfared et al., 2023; Sheridan et al., 2025). Within ocean basins, reproductive timing and seasonality is known to vary across latitudinal gradients (Baird et al., 2021; Howells et al., 2014) and regional predictions of spawning do not reliably translate to other localities. Moreover, while several recent studies of coral reproductive phenology in the central Indian Ocean detail patterns in the Acropora genus (Monfared et al., 2023; Sheridan et al., 2025), most other genera have only a handful of observations from few locations, particularly in the Indian Ocean (Baird et al., 2021).
Increasing knowledge of the current reproductive status of reef-building corals is essential in the context of reef conservation, and baseline data can be used to assess, monitor, and predict coral population health and resilience. Specifically documenting coral reproductive phenology and spawning behaviour has become imperative in recent decades as scientists and conservation practitioners have increased efforts to stem declines in coral populations and to regenerate reef habitat through active restoration (Banaszak et al., 2023; Miller et al., 2024). Traditional restoration practices rely upon asexual propagation, cloning existing coral via fragmentation, which does not directly increase genetic diversity or create genetically new individuals. Because sexual reproduction can produce hundreds of millions of coral propagules at a single time from many parents, it provides new genetic diversity that promotes the population resilience required to replenish and sustain future reef communities (Banaszak et al., 2023). Restoration projects have expanded to include sexual propagation in many areas of the Caribbean and the Indo-Pacific (Banaszak et al., 2023; Miller et al., 2022), and sexually propagated recruits have recently shown higher thermal tolerance during bleaching events compared to adult and asexually propagated coral (Miller et al., 2024). Coral restoration efforts in the Western Indian Ocean, including Zanzibar and mainland Tanzania, are increasing rapidly; however, all currently known projects use only asexual propagation via fragmentation.
Coral reproductive patterns in East Africa remain largely undocumented, despite the ecological importance of its biodiverse nearshore reefs. In Zanzibar, establishing baseline data on spawning timing is critical for understanding the reproductive potential of local coral populations and informing restoration strategies that incorporate sexual propagation—a key element for enhancing reef resilience (Banaszak et al., 2023). Although over 6,500 coral spawning records exist across the Indo-Pacific (Baird et al., 2021), empirical data from the Western Indian Ocean remain extremely limited. To date, only two opportunistically observed spawning events have been reported from Zanzibar (Bronstein & Loya, 2011; CHICOP, 2017), and no direct in situ observations have been published for East Africa. The only prior studies in the East Africa coastal region inferred spawning timing indirectly through histological analyses of gametogenesis and post-settlement monitoring of coral spat in Kenya’s Mombasa Marine Reserve (Mangubhai, Harrison & Obura, 2007; Mangubhai & Harrison, 2008). To begin to address this significant knowledge gap, we conducted in situ observations of coral spawning at a diverse reef site in Zanzibar, documenting the timing and gamete release behaviours for members of three Scleractinian species for which no prior spawning records exist in the Western Indian Ocean.
Materials & Methods
Portions of this text were previously published as part of a preprint (Lillis & Jiddawi, 2025).
Study area: Jambiani, Zanzibar, Tanzania
Zanzibar is a semi-autonomous region of the United Republic of Tanzania, comprising two major tropical islands (Unguja and Pemba) and many small ones (Fig. 1A). The Zanzibar archipelago has substantial fringing and patch coral reefs, estimated to cover 90 km^2^, hosting high biodiversity and providing significant economic value to humans (Bravo et al., 2021; Khamis, Kalliola & Käyhkö, 2017). The reef systems are key to support local fisheries, tourism, and provide coastal protection (Bravo et al., 2021; Khamis, Kalliola & Käyhkö, 2017). The eastern coasts of the two major islands have continuous reefs fringing the coastline and creating protected shallow lagoons with wide beaches.
Study site map. Map data ©2025 Google.(A) Unguja Island, Zanzibar with location of Jambiani village and study reef site (red). (B) Satellite image of the Marine Cultures House Reef with monitoring area delineated. Locations of each coral species observed spawning are indicated.
This study was conducted at a shallow patch reef site in Jambiani lagoon. Jambiani, a coastal village bordering a shallow lagoon, is located on the southeast coast of Unguja Island (Fig. 1A). The lagoon is formed by a fringing coral reef system several kilometers offshore creating a barrier from the open ocean, while the lagoon comprises patches of coral reef, seagrass, and soft bottom. Traditionally a fishing village, the area has seen rapid coastal development and environmental change in the past 10–15 years. This lagoon system experiences large tidal fluctuations (2–4 m tidal range), seasonal sedimentation, and is hydrodynamically active. This work was carried out under Zanzibar Research Permit #2001709162242701335980.
Zanzibar-based marinecultures.org is a non-profit organization operating a coral farm and transplantation program in Jambiani lagoon (Fig. 1). Beginning in 2014 corals were fragmented, grown on nursery tables, and planted to their “house reef” (MC reef; Fig. 1B). The MC reef is approximately 750 m^2^ (50 m × 15 m), ranging from 2–9 m depth, with established large (>five m diameter) colonies of Porites spp., Galaxea astreata, and Lobophyllia sp., and many smaller (0.5–1 m diameter) colonies of a diversity of species including many Echinpora, Leptoria, and Favites. This patch reef, while relatively small, is estimated to contain individuals of >40 species of hard coral, including at least 30 genera. Like other reefs around the world, Zanzibar recently experienced a mass coral bleaching event during 2024, severely impacting the MC reef site where the majority of previously planted Acroporids perished. The coral cover is now estimated at 35–40%, Acropora spp. are rare, and some surviving coral colonies show partial mortality. Due to tidal range and limited access to shallow reefs with healthy coral for nighttime monitoring, Zanzibar is a challenging location in which to carry out spawning observations. This site was selected due to logistical feasibility for monitoring (shallow, accessible, nearshore), a high diversity of coral species available to monitor, and interest by marinecultures.org in integrating sexual propagation techniques in their restoration program.
Field observations
A series of monthly night dives on SCUBA, beginning just after sunset, were carried out during November and December 2024, January and February 2025 at the MC site, totaling 1,402 min of observation. Dives were scheduled each month during the 3–6 nights after the full moon (NAFM), based on the highest number of observations of coral spawning during these nights in other locales of similar latitudes (Baird et al., 2021; Baird et al., 2022). Lunar phases and local sunset timing for Zanzibar were obtained from http://www.timeanddate.com. Because no previous knowledge of coral spawning existed for this site, and because the aim was to document any and all species present, coral species to observe were not pre-determined but rather observers watched all corals in their area for indications of spawning, returning to the same area and corals for each observation.
On each observation night, two divers descended at sunset to the reef site, each patrolling a small area (∼50 m^2^) for the duration of the dive (1–2 h depending on sea conditions and air consumption) to consistently monitor the same coral colonies each dive. Divers monitored all corals within their zone to check for “setting”, when gamete bundles appear in polyp mouths prior to release, as well as “spawning”, when positively buoyant spherical gamete bundles are released into the water column and float towards the surface. While coral spawning can be obscured by high amounts of turbidity and light-attracted plankton in the water, gamete bundles observed were distinct and were confirmed to be originating from coral polyp mouths.
Time-stamped photos and video were taken to document any setting or spawning behaviours observed (beginning at first observed setting and/or spawning), as well as the conclusion of any spawning event (no egg bundles remaining in polyp mouths). For any individual spawning seen, observations included data on setting and spawning timing, duration, and synchrony (when multiple individuals of the same species were present). The same areas, and therefore same set of coral colonies, were observed each month of observation. For coral that spawned and are reported herein, species were identified using established guides including Indo-Pacific Coral Finder (Kelley 2022) and http://www.coralsoftheworld.org. To confirm coral identification, photos and video were assessed by regional experts (D. Obura, pers. comm., 2024).
Results
Representatives of three hermaphroditic coral species were recorded spawning during this initial monitoring interval at MC reef, each with distinct timing and release behaviour (Table 1). Despite limited replicates of each species, the focused and repeated monitoring of the same individual corals enabled uncommonly detailed descriptions of their specific gamete release patterns. Divers returned to the specific location for all monitoring dives and recognized the coral colonies by sight. Spawning events observed are described in detail in individual sections below.
Table 1: Summary data for coral spawning.
Species 1: Galaxea astreata (Family Euphylliidae)
A large (∼eight m diameter) colony of G. astreata was observed spawning on the 4th nights after the full moon in November and December 2024 (Table 1, Fig. 2). This colony had experienced severe bleaching in April 2024, leading to approximately 30–40% mortality. Spawning (gamete release) was detected 46 and 54 min after sunset and continued for 31 and 20 min in November and December, respectively. Gamete bundles were white in colour and seen rising from the large colony. Due to the coral’s morphology and long extended tentacles, it was not possible to see gamete bundles emerge from polyp mouths in this species. The December spawning event was observed to be smaller (i.e., fewer gamete bundles released) with gamete bundles appearing to be smaller in diameter than during the larger spawn in November. Two other colonies of this species were present in the monitoring area, none of which were observed to spawn during the monitoring period.
(A–D) Galaxea astreata November 2024 spawning event.White-coloured gamete bundles rising in the water column are circled in red. Recent mortality of the colony can be seen in C and D.
Species 2: Favites pentagona (Family Merulinidae)
On the 6th night after the full moon in November 2024, ripe pink gamete bundles were observed setting (visible in polyp mouths) at 19:13 in a single colony of F. pentagona (Fig. 3A). Divers searched the monitoring area for additional colonies of F. pentagona but none were found, therefore it was only possible to observe one individual. First gamete release began at 19:24 (Fig. 3C), 14 min after setting was noted, and continued until the final bundle was released at 19:35. To check if this F. pentagona colony would spawn again at the same lunar timing in December, it was monitored 3–6 nights after the full moon from 30–90 min after sunset. No spawning was observed.
Favites pentagona spawning sequence in November 2024.Gamete setting is seen in panel (A). First gamete bundles were released at 19:24 (B) and continued for 11 min (C–E), with the ûnal bundle released seen in (F).
Species 3: Platygyra daedalea (Family Merulinidae)
A brain coral species common to the MC reef, P. daedalea, was observed spawning in December 2024 and in January 2025, on the 6th night after the full moon, starting at 80 min after sunset in both months (Table 1, Figs. 4 and 5). Gamete release was remarkably rapid in this species, with pulses of spawning lasting only 2–3 min. Five adjacent colonies of this species are present in a small area (<five m^2^) and were able to be monitored simultaneously. In December, a single individual colony was observed spawning. Gamete release had already begun when the diver came across the spawning colony, thus initial setting was not observed. However, only part of the colony spawned in the first pulse at 80 min after sunset, while other parts of the colony began setting shortly after with spawning occurring again 30 min later (Table 1, Fig. 4).
Platygyra daedalea spawning recorded in December 2024.(A and B) The initial rapid release of gamete bundle cloud. Gamete setting is seen in (C and D) before the second rapid release observed (E). Panel (F) shows empty gaping polyp mouths following the release.
Platygyra daedalea spawning sequence in January 2025, observed in several colonies.Red circles indicate gamete bundles in the water column. (A–D) Gamete setting for four colonies, (E–F) gamete bundles are released.
One individual colony was observed spawning in December, while four were documented to spawn in January, including the individual that spawned the previous month (Fig. 5). In January, a small colony showed visible pink gamete bundles setting starting at 19:28 with other adjacent colonies exhibiting setting gamete bundles within the next 15 min. Setting lasted 38–40 min, and gamete bundles were rapidly released by each colony in less than 2 min (Figs. 5D, 5E).
Discussion
This study presents new in situ documentation of spawning timing and behaviour for colonies of G. astreata, F. pentagona, and P. daedalea in the Western Indian Ocean, specifically coastal Zanzibar. These findings provide a valuable first baseline for coral scientists, conservationists, and restoration practitioners, and support future monitoring efforts in the region. Though limited to one site over four months and few colonies, our observations contribute novel information on coral spawning in a region where such data have been scarce. They expand the known geographic range of spawning for these species, previously reported mainly in the Indo-Pacific, particularly Asia. For instance, F. pentagona and P. daedalea have been recorded spawning in Thailand, Singapore, and Taiwan (Baird et al., 2021; Lin & Nozawa, 2017), and G. astreata in Singapore (Baird et al., 2021). Our results partially align with these records but also reveal differences likely driven by local environmental conditions.
In Zanzibar, a G. astreata spawned on the fourth night after the full moon in November and December, within a consistent time window (46–54 min after sunset). Presenting similar seasonality, with reproductive peaks in the time of most rapid temperature increases, Singapore records show spawning in April, 3–5 nights after the full moon (Baird et al., 2021), but with greater variation in timing. In 2012 and 2014, the Singapore-documented spawning by Tun and Low (Baird et al., 2021) comprised 10 G. astreata individuals, with the spawning time ranging from 47 to 157 min after sunset (median = 103). While only one G. astreata individual was observed in Zanzibar, the consistent timing we observed suggests intra-individual regularity but highlights the need for broader sampling to assess inter-individual variation. F. pentagona was also represented in Zanzibar by a single colony, which spawned in November on the sixth night after the full moon, 63 min after sunset. No subsequent spawning was observed December through February. In Taiwan, where the only other observations of F. pentagona exist, this species has been documented spawning only twice, in April 2019 on the fourth and sixth nights after the full moon, several hours after sunset (Baird et al., 2021). These limited data and the variability in spawning nights and times observed between Zanzibar and Taiwan in these species for which we have limited data underscore the need for further monitoring to assess population-level synchrony.
P. daedalea spawning was observed in multiple individuals, offering insights into intra-population synchrony at our site. A single colony spawned in December, while four of six colonies observed spawned in January. No spawning was seen in February. All spawning occurred on the sixth night after the full moon, beginning around 80 min after sunset, with gamete release lasting just 1–3 min. This brief release window has implications for monitoring and gamete collection. In contrast, P. daedalea previously documented spawning in Taiwan, Thailand, and Singapore spawned 4–8 days after the full moon, at variable times past sunset, and none of the releases were recorded as rapidly occurring (Baird et al., 2021; Lin & Nozawa, 2017). Notably, Kenyan populations show peak spawning in February–March with a secondary peak in August–October (Mangubhai & Harrison, 2008), based on patterns in gametogenesis and not direct observations of spawning. These variations in seasonal timing might reflect spatial or temporal differences in environmental cues such as temperature, photoperiod, and tidal cycles, all of which can influence gametogenesis and spawning readiness (Lin & Nozawa, 2023). Further observation over multiple years and expanded spatial coverage is needed to understand the factors influencing reproductive timing.
Conclusions
Our findings reveal distinct species-specific spawning windows and behaviours at a single reef site, underscoring the complexity and ecological significance of coral reproductive strategies at local scales. Variation in gamete setting, release duration, and evidence of split spawning in individual colonies was observed. These behavioural nuances further emphasize the importance of localized monitoring, as within the same reef system, reproductive strategies can differ among individuals and species. The presence of split spawning—where not all colonies or an entire colony spawns simultaneously—may serve as a bet-hedging strategy, increasing the likelihood of successful fertilization under variable conditions. However, it also presents challenges for coral restoration practices that rely on synchronous gamete collection.
By establishing initial baseline reproductive data for corals in Zanzibar, this data report supports conservation strategies in a region facing growing environmental pressures. As spawning data are expanded, they can inform the timing of conservation and restoration interventions—such as closed areas, larval propagation, or managed breeding—ensuring they are aligned with natural reproductive windows to maximize success. Understanding this variability is vital for effective reef restoration and management, especially as reproductive timing may be sensitive to climate change. While this study provides an important first step, long-term, multi-site monitoring across more species is essential for a comprehensive understanding of reproductive dynamics in Zanzibar and across the WIO.
Supplemental Information
10.7717/peerj.20999/supp-1Supplemental Information 1Coral spawning monitoring logData log for coral spawning monitoring in Jambiani Lagoon, Zanzibar between November 2024 and February 2025.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Babcock RC Bull GD Harrison PL Heyward AJ Oliver JK Wallace CC Willis BL 1986 Synchronous spawnings of 105 scleractinian coral species on the Great Barrier Reef Marine Biology 90337939410.1007/BF 00428562 · doi ↗
- 2Baird AH Edwards AJ Guest JR Harii S Hatta M Lachs L Mera H Sinniger F Abrego D Ben-Zvi O Bronstein O Cabaitan PC Cumbo VR Eyal G Eyal-Shaham L Feldman B Figueiredo J Flot J-F Grinblat M Heyward A Hidaka M Hirose M Iguchi A Isomura N Kinzie RA Kitanobo S Kuba A Levy O Loya Y Mezaki T Mohamed AR Morita M Nojima S Nozawa Y Prasetia R Puill-Stephan E Ramirez-Portilla C Rapuano H Rosenberg Y Sakai Y Sakai K Shlesinger T Terraneo TI Yakovleva I Yamamoto HH Yamazato K 2022 A coral spawning calendar for Sesoko Station, Okinawa, Japan Galaxea, Journa · doi ↗
- 3Baird AH Guest JR Edwards AJ Bauman AG Bouwmeester J Mera H Abrego D Alvarez-Noriega M Babcock RC Barbosa MB Bonito V Burt J Cabaitan PC Chang CF Chavanich S Chen CA Chen CJ Chen WJ Chung FC Connolly SR Cumbo VR Dornelas M Doropoulos C Eyal G Eyal-Shaham L Fadli N Figueiredo J Flot JF Gan SH Gomez E Graham EM Grinblat M Gutiérrez-Isaza N Harii S Harrison PL Hatta M Ho NAJ Hoarau G Hoogenboom M Howells EJ Iguchi A Isomura N Jamodiong EA Jandang S Keyse J Kitanobo S Kongjandtre N Kuo CY Ligson C Lin CH Low J Loya Y Maboloc EA Madin JS Mezaki T Min C Mo · doi ↗ · pubmed ↗
- 4Baird AH Guest JR Willis BL 2009 Systematic and biogeographical patterns in the reproductive biology of Scleractinian corals Annual Review of Ecology, Evolution, and Systematics 40155157110.1146/annurev.ecolsys.110308.120220 · doi ↗
- 5Banaszak AT Marhaver KL Miller MW Hartmann AC Albright R Hagedorn M Harrison PL Latijnhouwers KR Mendoza Quiroz S Pizarro V Chamberland VF 2023 Applying coral breeding to reef restoration: best practices, knowledge gaps, and priority actions in a rapidly-evolving field Restoration Ecology 317e 1391310.1111/rec.13913 · doi ↗
- 6Bellwood DR Hughes TP Folke C Nyström M 2004 Confronting the coral reef crisis Nature 429699410.1038/nature 0269115215854 · doi ↗ · pubmed ↗
- 7Boch CA Ananthasubramaniam B Sweeney AM Doyle FJ Morse DE 2011 Effects of light dynamics on coral spawning synchrony The Biological Bulletin 220316117310.1086/BB Lv 220n 3p 16121712225 · doi ↗ · pubmed ↗
- 8Bravo H Cannicci S Huyghe F Leermakers M Sheikh MA Kochzius M 2021 Ecological health of coral reefs in Zanzibar Regional Studies in Marine Science 4810201410.1016/j.rsma.2021.102014 · doi ↗