Minutellins E-I and daldinone L, new secondary metabolites from different species of the Hypoxylaceae (Xylariales, Ascomycota)
Christopher Lambert, Mohammad Javad Pourmoghaddam, Esteban Charria-Girón, Frank Surup, Seyed Akbar Khodaparast, Hermann Voglmayr, Irmgard Krisai-Greilhuber, Marc Stadler

TL;DR
This study identifies new secondary metabolites from fungi in the Hypoxylaceae family and corrects a previous chemical structure misassignment.
Contribution
Discovery of a new daldinone derivative and five new minutellin-type azaphilones, along with a structural correction of daldinone A.
Findings
A new daldinone derivative and five new minutellin-type azaphilones were identified from Hypoxylaceae fungi.
Daldinone A was found to have been misassigned and is actually daldinone C.
The distribution of minutellin-type azaphilones across the Hypoxylaceae family was discussed.
Abstract
Fungi of the family Hypoxylaceae (Xylariales, Ascomycota) are ubiquitously distributed and fulfil important ecological roles as saprobes, pathogens, and endophytes. Members of this family tend to store large amounts of secondary metabolites in their carbonaceous stromatal tissue commonly formed on colonised wood. This feature is of both taxonomic and chemical value by serving as chemotaxonomic markers and sources of potential new and bioactive compounds. Despite tremendous progress in the characterisation of Hypoxylaceae both in terms of genomics and secondary metabolomics, many unknown metabolites remain to be identified or elucidated. Here, we report on the polyphasic, i.e. morphological, chemical, and genetical analysis of Hypoxylon and Annulohypoxylon spp. collected from the Iranian shore of the Caspian Sea and a chemotaxonomic study of the constituents of a concurrently studied…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
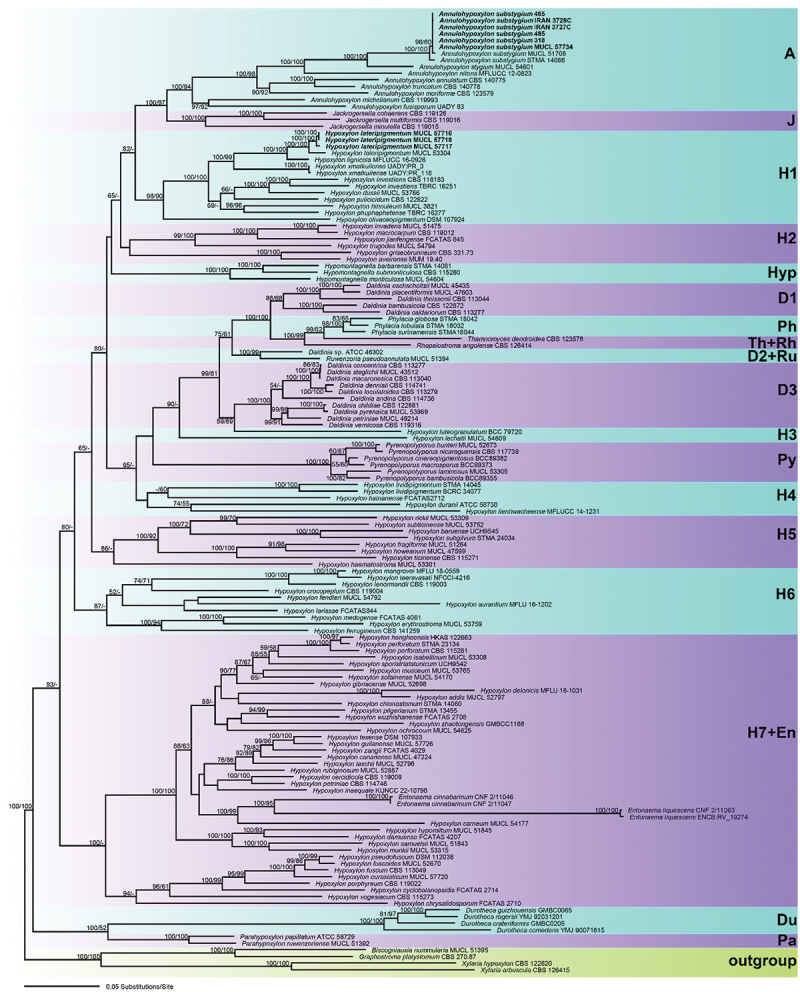 Figure 1
Figure 1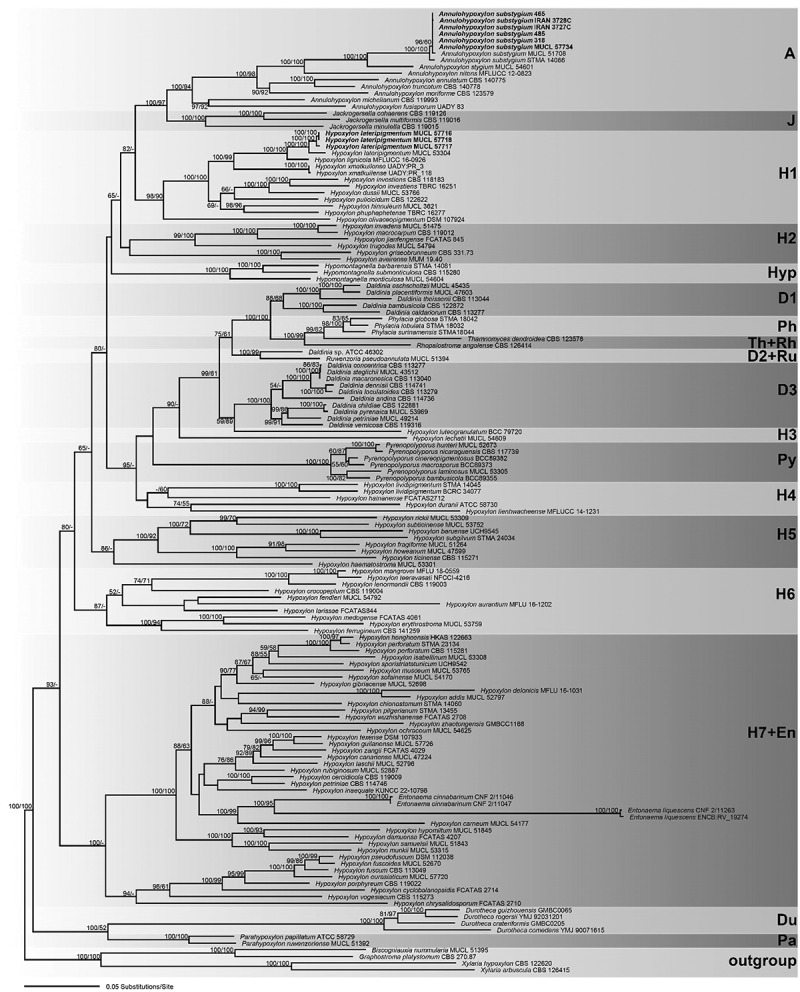 Figure 2
Figure 2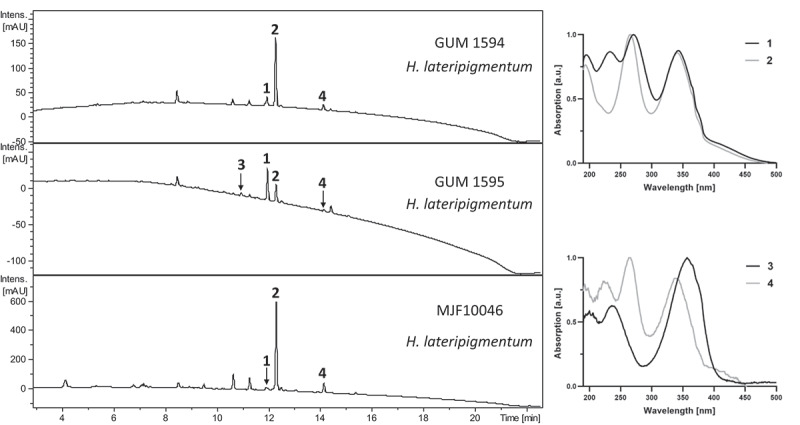 Figure 3
Figure 3 Figure 4
Figure 4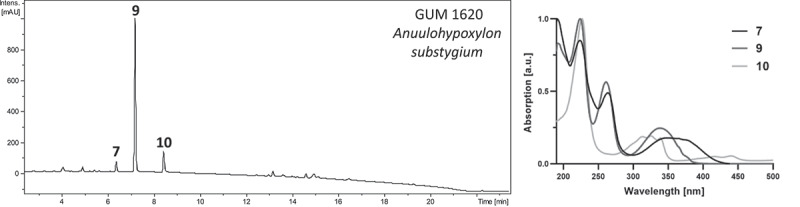 Figure 5
Figure 5 Figure 6
Figure 6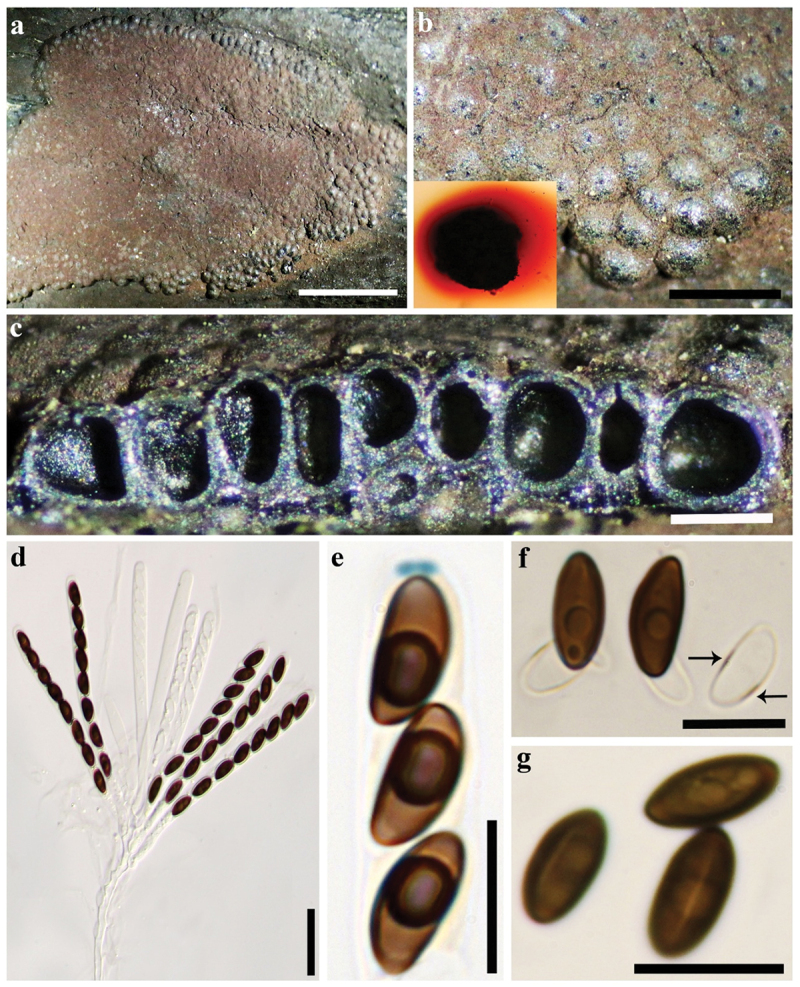 Figure 7
Figure 7 Figure 8
Figure 8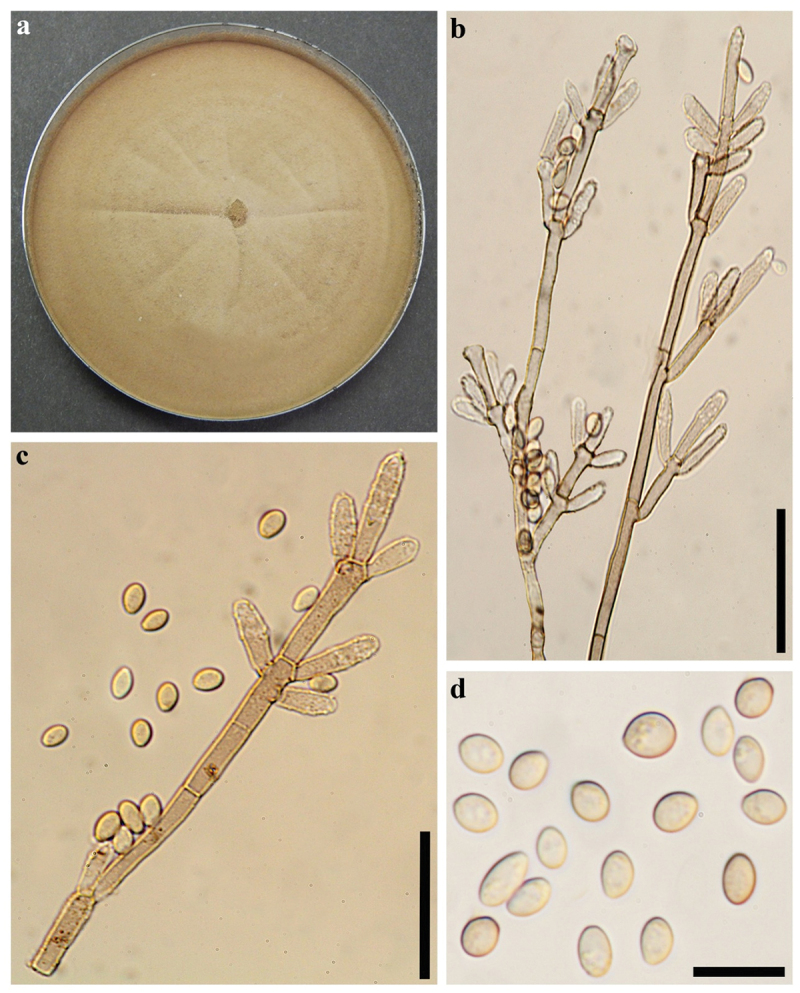 Figure 9
Figure 9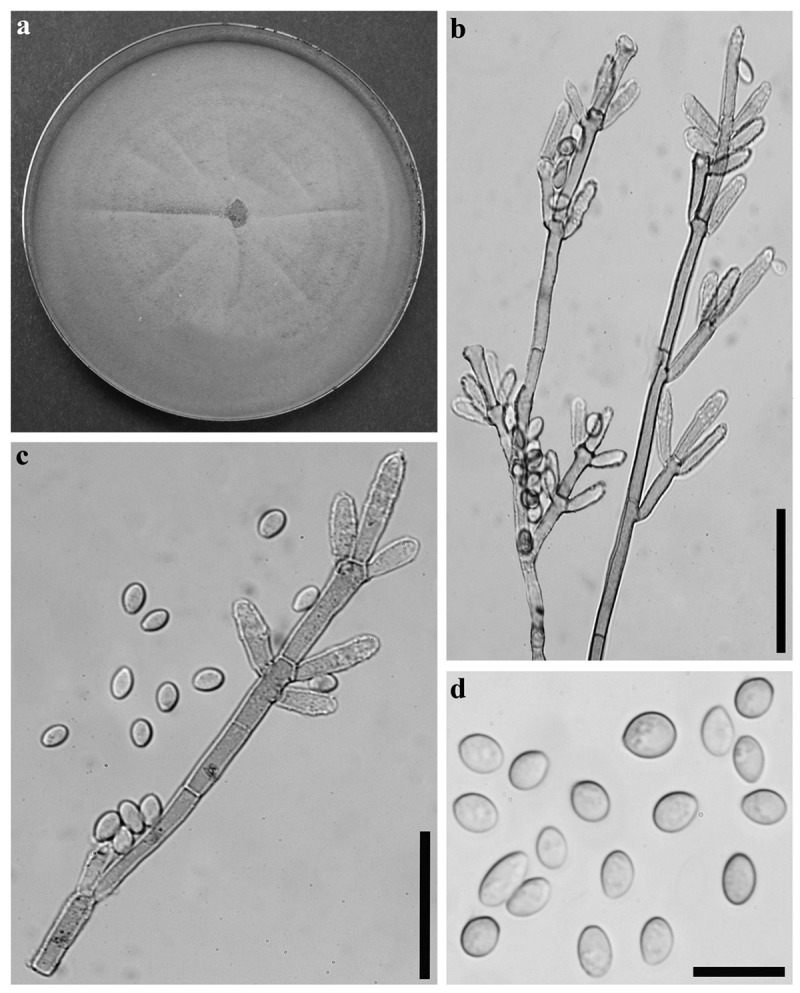 Figure 10
Figure 10 Figure 11
Figure 11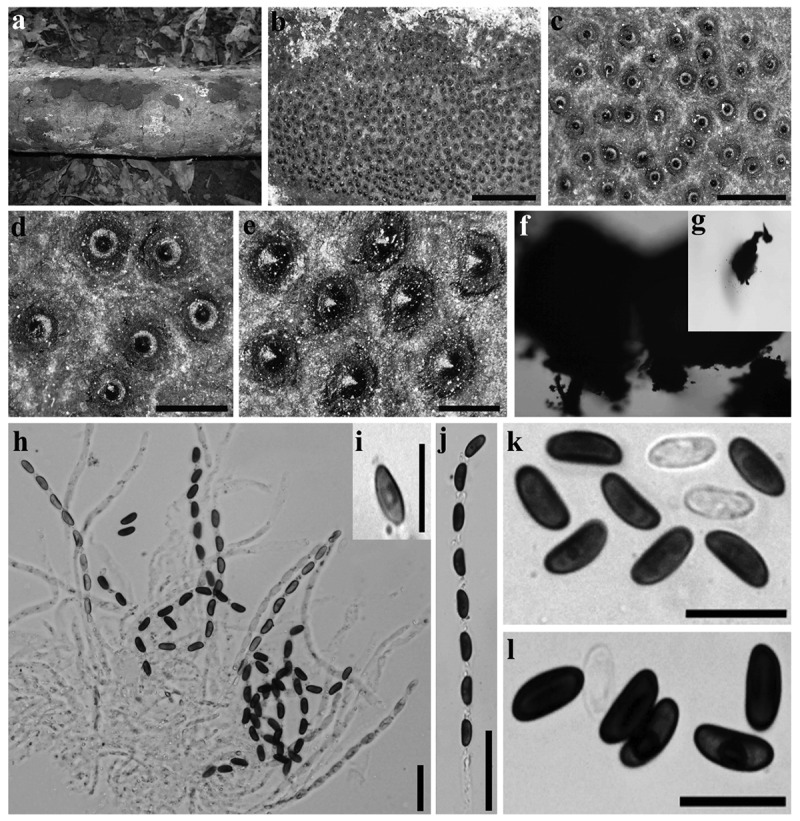 Figure 12
Figure 12 Figure 13
Figure 13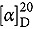 Figure 14
Figure 14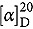 Figure 15
Figure 15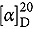 Figure 16
Figure 16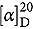 Figure 17
Figure 17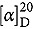 Figure 18
Figure 18- —Iran National Science Foundation (INSF)
- —DAAD10.13039/501100001655
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications
Introduction
Fungi of the family Hypoxylaceae (Xylariales, Ascomycota) are distributed worldwide and may occur as both, opportunistic plant pathogens and saprobes, in addition to their endophytic lifestyle (Helaly et al. 2018). They form a wide array of morphologically conspicuous, carbonised teleomorphs, along with several distinct types of anamorph structures in culture, which can occasionally also be observed in nature (Ju and Rogers 1996; Wendt et al. 2018). While teleomorph morphology has long been regarded as an important character in the systematics of the Hypoxylaceae, it became clear that this phenotypic feature alone is insufficient, as it is inconsistent with the phylogenetic evidence acquired over the years (Hsieh and Rogers 2005; Jaklitsch et al. 2016). Indeed, it has repeatedly been confirmed that the anamorph morphology first used by Ju and Rogers (1996) to segregate the “hypoxyloid” (nodulisporium-like) and “xylarioid” (geniculosporium-like anamorph) taxa is a better predictor of the phylogenetic affiliation of a given species. Still, teleomorph morphology repeatedly proves to have merit in delimiting individual species from each other. Nowadays, taxonomic assessment of members of the Hypoxylaceae involves a combinatorial analysis of both teleo- and anamorph morphology, genetic analysis, which is further complemented by chemical analysis within a chemotaxonomic framework (Wendt et al. 2018). Chemical analysis usually comprises an analysis of secondary metabolites in stromata present in almost all members of Hypoxylaceae. Together with secondary metabolites produced in culture, these chemical constituents repeatedly served as good indicators for phylogenetic affiliations (such as mitorubrins for the “red” Hypoxylon, Stadler et al. 2008; see also Stadler et al. 2014; Kuhnert et al. 2015, 2017a; Wendt et al. 2018; Lambert et al. 2019 for additional examples). Genetic analysis nowadays usually includes data derived from the internal transcribed spacer and large subunit ribosomal DNA (ITS and LSU rDNA), and the DNA-directed RNA polymerase II ribosomal and β-tubulin (rpb2, tub2) genes, both used in multi-locus genealogies for phylogenetic inference (Wendt et al. 2018). This polyphasic approach allowed the resurrection and segregation of the Hypoxylaceae, for example, from Xylariaceae based on phylogenetic and chemical evidence (Wendt et al. 2018). Another example comprises the recently described genus Parahypoxylon, which was erected to accommodate phylogenetically distinct specimens of Hypoxylon s. Ju and Rogers (Cedeño-Sanchez et al. 2023). In the latter study, sequences generated from an ascospore-derived isolate of a specimen collected in the Democratic Republic of Congo were shown to cluster with the aberrant Hypoxylon papillatum (now: Parahypoxylon papillatum), which typically resolved in a position basal to other Hypoxylaceae (Wendt et al. 2018; Lambert et al. 2019, 2021; Pourmoghaddam et al. 2020). State-of-the-art metabolomics were then used for the first time for Hypoxylaceae to propose an unprecedented chemical diversity of cohaerin-type azaphilone compounds in stromatal extracts, which are commonly found in the hypoxylaceous genus Jackrogersella (Quang et al. 2005, 2006; Surup et al. 2013). The collected specimen was described as Parahypoxylon ruwenzoriense sp. nov.; however, paucity of the available material prevented any in-depth chemical characterisation (Cedeño-Sanchez et al. 2023). Similar compounds were previously described to be extant in A. michelianum and we further noted similar compounds, besides other unidentifiable compounds, in a collection of Iranian specimens showing affiliations to Hypoxylon and Annulohypoxylon collected near the Caspian Sea. This motivated us to embark on studying the taxonomic affiliations of the Iranian specimen and to characterise these potentially new stromatal secondary metabolites.
Materials and methods
All scientific names listed throughout the manuscript follow the respective entries in Index Fungorum (http://www.indexfungorum.org) and are hence given without authorities or year or publication. Herbaria and culture collections are abbreviated as recommended by Index Herbariorum (http://sweetgum.nybg.org/science/ih).
Morphological observation
2.1.
The fungal specimens were collected in northern Iran (Guilan and Mazandaran Provinces). For light microscopy, fresh collections, single ascospore isolations, and cultures were examined for macro- and micromorphological characteristics, according to Pourmoghaddam et al. (2020). Dried specimens were deposited in the University of Guilan Mycological Herbarium (GUM). Living cultures were deposited in the culture collection MUCL (Louvain la-Neuve, Belgium) and in the Iranian Fungal Culture Collection, Iranian Research Institute of Plant Protection, Tehran, Iran (IRAN).
DNA extraction, PCR, and sequencing
2.2.
DNA extraction of fresh cultures and amplification of the ITS (nuc rDNA internal transcribed spacer region containing ITS1-5.8S-ITS2), LSU (5’ 1,200 bp of the large subunit nuc 28S rDNA), rpb2 (partial second largest subunit of the DNA-directed RNA polymerase II), and tub2 (partial β-tubulin) loci were carried out as described by Wendt et al. (2018).
Phylogenetic analyses
2.3.
Published sequences of a single accession for each Annulohypoxylon and Hypoxylon species served as basis for the sequence matrix. Information on all used strains, their corresponding sequences and GenBank accession numbers can be found in Table 1. To reveal the phylogenetic position of the Iranian Annulohypoxylon and Hypoxylon accessions, the newly generated sequences were aligned with the GenBank sequences. All alignments were produced with the server versions of MAFFT v. 7.490 (www.ebi.ac.uk/Tools/mafft or http://mafft.cbrc.jp/alignment/server/; Katoh et al. 2019) and checked and refined using BioEdit v. 7.0.4.1 (Hall 1999).Table 1.Isolation and accession numbers of sequences used in the phylogenetic analyses.SpeciesStrain numberOriginStatusGenBank accession numberReferencesITSLSUrpb2tub2Annulohypoxylon annulatumCBS 140775TexasETKY610418KY610418KY624263KX376353Kuhnert et al. (2017a; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Annulohypoxylon fusisporumUADY 83MexicoHTOR807998OR807987OR825472OR825468Reyes et al. (2024)Annulohypoxylon michelianumCBS 119993Spain KX376320KY610423KY624234KX271239Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Annulohypoxylon moriformeCBS 123579Martinique KX376321KY610425KY624289KX271261Kuhnert et al. (2017a; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Annulohypoxylon nitensMFLUCC 12-0823Thailand KJ934991KJ934992KJ934994KJ934993Daranagama et al. (2015)Annulohypoxylon stygiumMUCL 54601French Guiana KY610409KY610475KY624292KX271263Wendt et al. (2018)Annulohypoxylon substygiumMUCL 51708IranETKC968915N/AN/AKC977285Kuhnert et al. (2014)Annulohypoxylon substygiumSTMA 14066Argentina KU604575N/AN/AKU159526Sir et al. (2016; ITS), Kuhnert et al. (2017a; tub2)Annulohypoxylon substygiumMUCL 57734****Iran PV522280PV522289PV524395PV524404**This studyAnnulohypoxylon substygiumIRAN 3727C****Iran PV522281PV522290PV524396PV524405**This studyAnnulohypoxylon substygiumIRAN 3728C****Iran PV522282PV522291PV524397PV524406**This studyAnnulohypoxylon substygium318****Iran PV522283PV522292PV524398PV524407**This studyAnnulohypoxylon substygium465****Iran PV522284PV522293PV524399PV524408**This studyAnnulohypoxylon substygium485****Iran PV522285PV522294PV524400PV524409**This studyAnnulohypoxylon truncatumCBS 140778TexasETKY610419KY610419KY624277KX376352Kuhnert et al. (2017a; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Biscogniauxia nummulariaMUCL 51395FranceETKY610382KY610427KY624236KX271241Wendt et al. (2018)Daldinia andinaCBS 114736EcuadorHTAM749918KY610430KY624239KC977259Bitzer et al. (2008; ITS), Kuhnert et al. (2014; tub2), Wendt et al. (2018; LSU, rpb2)Daldinia bambusicolaCBS 122872ThailandHTKY610385KY610431KY624241AY951688Hsieh and Rogers (2005; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Daldinia childiaeCBS 122881FranceHTKU683757MH874773KU684290KU684129U’Ren et al. (2016; ITS, rpb2, tub2), Vu et al. (2019; LSU)Daldinia loculatoidesCBS 113279UKETAF176982KY610438KY624247KX271246Wendt et al. (2018)Daldinia petriniaeMUCL 49214AustriaETAM749937KY610439KY624248KC977261Bitzer et al. (2008; ITS), Kuhnert et al. (2014; tub2), Wendt et al. (2018; LSU, rpb2)Daldinia theisseniiCBS 113044ArgentinaPTKY610388KY610441KY624251KX271247Wendt et al. (2018)Daldinia vernicosaCBS 119316GermanyETKY610395KY610442KY624252KC977260Kuhnert et al. (2014; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Daldinia sp.ATCC 46302USA KY610389KY610443KY624253KX271248Wendt et al. (2018)Durotheca comedensYMJ 90071615Taiwan of ChinaHTEF026128N/AJX507793EF025613Ju et al. (2003)Durotheca crateriformisGMBC0205ChinaHTMH645426MH645425MH645427MH049441de Long et al. (2019)Durotheca guizhouensisGMBC0065ChinaHTMH645423MH645421MH645422MH049439de Long et al. (2019)Durotheca rogersiiYMJ 92031201Taiwan of ChinaHTEF026127N/AJX507794EF025612Ju et al. (2007)Entonaema cinnabarinumCNF 2/11046Croatia OQ863621OQ863622OQ877102OQ877113Pošta et al. (2023)Entonaema cinnabarinumCNF 2/11047Croatia OQ863735OQ864983OQ877103OQ877114Pošta et al. (2023)Entonaema liquescensCNF 2/11263USA OQ869784OQ865124OQ877106OQ877117Pošta et al. (2023)Entonaema liquescensENCB:RV_19274Mexico OR807997OR807993OR825474OR825466Reyes et al. (2024)Graphostroma platystomumCBS 270.87France JX658535DQ836906KY624296HG934108Zhang et al. (2006; LSU), Stadler et al. (2014; ITS), Koukol et al. (2015; tub2), Wendt et al. (2018; rpb2)Hypomontagnella barbarensisSTMA 14081ArgentinaHTMK131720MK131718MK135891MK135893Lambert et al. (2019)Hypomontagnella monticulosaMUCL 54604French GuianaETKY610404KY610487KY624305KX271273Wendt et al. (2018)Hypoxylon aveirenseMUM 19.40PortugalHTMN053021ON954142OP251028MN066636Vicente et al. (2021; ITS, tub2), Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon baruenseUCH9545PanamaHTMN056428ON954143PP732079MK908142Cedeño-Sanchez et al. (2020; ITS, tub2), Cedeño-Sanchez et al. (2023; LSU); Cedeño-Sanchez et al. (2025; rpb2)Hypoxylon canarienseMUCL 47224SpainPTON792787ON954140OP251029ON813073Cedeño-Sanchez et al. (2023)Hypoxylon carneumMUCL 54177France KY610400KY610480KY624297KX271270Wendt et al. (2018)Hypoxylon cercidicolaCBS 119009France KC968908KY610444KY624254KC977263Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon chionostomumSTMA 14060Argentina KU604563ON954144OP251030ON813072Sir et al. (2016; ITS), Cedeño-Sanchez et al. (2023; LSU, rpb2, tub2)Hypoxylon chrysalidosporumFCATAS 2710ChinaHTOL467294OL615106OL584222OL584229Ma et al. (2022)Hypoxylon crocopeplumCBS 119004France KC968907KY610445KY624255KC977268Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon cyclobalanopsidisFCATAS 2714ChinaHTOL467298OL615108OL584225OL584232Ma et al. (2022)Hypoxylon damuenseFCATAS 4207ChinaHTON075427ON075433ON093251ON093245Song et al. (2022b)Hypoxylon delonicisMFLU 16–1031ThailandHTMT215503MT386008N/AMT212215Perera et al. (2020)Hypoxylon duraniiATCC 58730MexicoHTPP718984PP729636PP732085PP721316Cedeño-Sanchez et al. (2025)Hypoxylon dussiiMUCL 53766GuadeloupeHTPP718981PP729635PP732081PP721315Cedeño-Sanchez et al. (2025)Hypoxylon erythrostromaMUCL 53759Martinique KC968910ON954154OP251031N/AKuhnert et al. (2014; ITS), Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon eurasiaticumMUCL 57720IranHTMW367851N/AMW373852MW373861Lambert et al. (2021)Hypoxylon fendleriMUCL 54792French Guiana KF234421KY610481KY624298KF300547Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon ferrugineumCBS 141259Austria KX090079N/AN/AKX090080Friebes and Wendelin (2016)Hypoxylon fragiformeMUCL 51264GermanyETKC477229KM186295MK887342KX271282Stadler et al. (2013; ITS), Daranagama et al. (2015; LSU), Wendt et al. (2018; tub2), Sir et al. (2019; rpb2)Hypoxylon fuscoidesMUCL 52670FranceHTON792789ON954145OP251038ON813076Cedeño-Sanchez et al. (2023)Hypoxylon fuscumCBS 113049FranceETKY610401KY610482KY624299KX271271Wendt et al. (2018)Hypoxylon gibriacenseMUCL 52698FranceHTKC968930ON954146N/AON813074Kuhnert et al. (2014; ITS), Cedeño-Sanchez et al. (2023; LSU, rpb2, tub2)Hypoxylon griseobrunneumCBS 331.73IndiaHTKY610402KY610483KY624300KC977303Kuhnert et al. (2014; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Hypoxylon guilanenseMUCL 57726IranHTMT214997MT214992MT212235MT212239Pourmoghaddam et al. (2020)Hypoxylon haematostromaMUCL 53301MartiniqueETKC968911KY610484KY624301KC977291Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon hainanenseFCATAS2712ChinaHTOL467296OL616132OL584224OL584231Ma et al. (2022)Hypoxylon hinnuleumMUCL 3621USAHTMK287537MK287549MK287562MK287575Sir et al. (2019)Hypoxylon howeanumMUCL 47599Germany AM749928KY610448KY624258KC977277Bitzer et al. (2008; ITS), Kuhnert et al. (2014; tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon hypomiltumMUCL 51845Guadeloupe KY610403KY610449KY624302KX271249Wendt et al. (2018)Hypoxylon inaequaleKUNCC 22–10798ChinaHTON329812ON329815N/AN/AJayawardena et al. (2022)Hypoxylon invadensMUCL 51475FranceHTMT809133MT809132MT813037MT813038Becker et al. (2020)Hypoxylon investiensCBS 118183Malaysia KC968925KY610450KY624259KC977270Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon investiensTBRC 16251Thailand OP856531OP856521OQ108848OQ144968Suetrong et al. (2023)Hypoxylon isabellinumMUCL 53308MartiniqueHTKC968935ON954155N/AKC977295Kuhnert et al. (2014; ITS, tub2), Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon jianfengenseFCATAS 845ChinaHTMW984546MZ029707MZ047260MZ047264Song et al. (2022a)Hypoxylon larissaeFCATAS844ChinaHTMW984548MZ029706MZ047258MZ047262Song et al. (2022a)Hypoxylon laschiiMUCL 52796France JX658525ON954147OP251027ON813075Stadler et al. (2014; ITS), Cedeño-Sanchez et al. (2023; LSU, rpb2, tub2)Hypoxylon lateripigmentumMUCL 53304MartiniqueHTKC968933KY610486KY624304KC977290Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon lateripigmentumMUCL 57716****Iran PV522286PV522295PV524401PV524410**This studyHypoxylon lateripigmentumMUCL 57717****Iran PV522287PV522296PV524402PV524411**This studyHypoxylon lateripigmentumMUCL 57718****Iran **PV522288PV522297PV524403**PV524412**This studyHypoxylon lechatiiMUCL 54609French Guiana KF923407ON954148OP251033KF923405Kuhnert et al. (2014; ITS, tub2), Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon lenormandiiCBS 119003Ecuador KC968943KY610452KY624261KC977273Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon lienhwacheenseMFLUCC 14-1231Thailand KU604558MK287550MK287563KU159522Sir et al. (2016; ITS, tub2), Sir et al. (2019; LSU, rpb2)Hypoxylon lignicolaMFLUCC 16-0926ChinaHTMK828609MK835808MN156534N/ALuo et al. (2019)Hypoxylon lividipigmentumBCRC 34077MexicoITJN979433N/AN/AAY951735Hsieh and Rogers (2005)Hypoxylon lividipigmentumSTMA 14045Argentina ON792788ON954149PP732080ON813077Cedeño-Sanchez et al. (2023; ITS, LSU, tub2), Cedeño-Sanchez et al. (2025; rpb2)Hypoxylon luteogranulatumBCC 79720ThailandHTPP955304PP955701PP968827PQ000351Wongkanoun et al. (2025)Hypoxylon macrocarpumCBS 119012Germany ON792785ON954151N/AON813071Cedeño-Sanchez et al. (2023)Hypoxylon mangroveiMFLU 18-0559ThailandHTMN047116MN017880N/AMN077053Dayarathne et al. (2020)Hypoxylon medogenseFCATAS 4061ChinaHTON075425ON075431ON093249ON093243Song et al. (2022b)Hypoxylon munkiiMUCL 53315Martinique KC968912ON954153OP251035KC977294Kuhnert et al. (2014; ITS, tub2), Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon musceumMUCL 53765Guadeloupe KC968926KY610488KY624306KC977280Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon ochraceumMUCL 54625MartiniqueETKC968937N/AKY624271KC977300Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; rpb2)Hypoxylon olivaceopigmentumDSM 107924USAHTMK287530MK287542MK287555MK287568Sir et al. (2019)Hypoxylon perforatumSTMA 23134USAETPP718982PP729634PP732084PP721314Cedeño-Sanchez et al. (2025)Hypoxylon perforatumCBS 115281France KY610391KY610455KY624224KX271250Wendt et al. (2018)Hypoxylon petriniaeCBS 114746FranceHTKY610405KY610491KY624279KX271274Wendt et al. (2018)Hypoxylon phuphaphetenseTBRC 16277ThailandHTOP856538OP856528OQ108849OQ144973Preedanon et al. (2023)Hypoxylon pilgerianumSTMA 13455Martinique KY610412KY610412KY624308KY624315Wendt et al. (2018)Hypoxylon porphyreumCBS 119022France KC968921KY610456KY624225KC977264Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon pseudofuscumDSM 112038GermanyHTMW367857MW367848MW373858MW373867Lambert et al. (2021)Hypoxylon pulicicidumCBS 122622MartiniqueHTJX183075KY610492KY624280JX183072Bills et al. (2012; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon rickiiMUCL 53309MartiniqueETKC968932KY610416KY624281KC977288Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon rubiginosumMUCL 52887GermanyETKC477232KY610469KY624266KY624311Stadler et al. (2013; ITS), Wendt et al. (2018; LSU, rpb2, tub2)Hypoxylon samuelsiiMUCL 51843GuadeloupeETKC968916KY610466KY624269KC977286Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon sofainenseMUCL 54170GuadeloupeHTPP718983PP729633PP732083PP721313Cedeño-Sanchez et al. (2025)Hypoxylon sporistriatatunicumUCH9542PanamaHTMN056426ON954150OP251036MK908140Cedeño-Sanchez et al. (2020; ITS, tub2); Cedeño-Sanchez et al. (2023; LSU, rpb2)Hypoxylon subgilvumSTMA 24034Panama PP718985PP729637PP732082PP721317Cedeño-Sanchez et al. (2025)Hypoxylon teeravasatiNFCCI-4216IndiaHTKY863509MF385274MG986895MG986894Phookamsak et al. (2019)Hypoxylon texenseDSM 107933USAHTMK287536MK287548MK287561MK287574Sir et al. (2019)Hypoxylon ticinenseCBS 115271France JQ009317KY610471KY624272AY951757Hsieh and Rogers (2005; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon trugodesMUCL 54794Sri LankaETKF234422KY610493KY624282KF300548Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon vogesiacumCBS 115273France KC968920KY610417KY624283KX271275Kuhnert et al. (2014; ITS), Kuhnert et al. (2017a; tub2), Wendt et al. (2018; LSU, rpb2)Hypoxylon wuzhishanenseFCATAS 2708ChinaHTOL467292OL615104OL584220OL584227Ma et al. (2022)Hypoxylon xmatkuilenseUADY:PR_3MexicoHTOR807999OR807990OR825476OR825467Reyes et al. (2024)Hypoxylon xmatkuilenseUADY:PR_118MexicoPTOR807996OR807995PP239283PP239284Reyes et al. (2024)Hypoxylon zangiiFCATAS 4029ChinaHTON075423ON075429ON093247ON093241Song et al. (2022b)Hypoxylon zhaotongensisGMBCC1168ChinaHTOP597690OP598100N/AN/AZhang et al. (2023)Jackrogersella cohaerensCBS 119126Germany KY610396KY610497KY624270KY624314Wendt et al. (2018)Jackrogersella minutellaCBS 119015Portugal KY610381KY610424KY624235KX271240Kuhnert et al. (2017a; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Jackrogersella multiformisCBS 119016GermanyETKC477234KY610473KY624290KX271262Kuhnert et al. (2014; ITS), Kuhnert et al. (2017a; tub2), Wendt et al. (2018; LSU, rpb2)Parahypoxylon papillatumATCC 58729USAHTKC968919KY610454KY624223KC977258Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Parahypoxylon ruwenzorienseMUCL 51392D. R. CongoHTON792786ON954156OP251039ON813078Cedeño-Sanchez et al. (2023)Phylacia globosaSTMA 18042Argentina OQ437889OQ437885OQ453168OQ453172Lambert et al. (2023)Phylacia lobulataSTMA 18032ArgentinaHTOQ437892OQ437882OQ453166N/ALambert et al. (2023)Phylacia surinamensisSTMA 18044Argentina OQ437891OQ437887OQ453167N/ALambert et al. (2023)Pyrenopolyporus bambusicolaBCC89355ThailandHTOP304856OP304876OP981624OQ101839Wongkanoun et al. (2023)Pyrenopolyporus cinereopigmentosusBCC89382ThailandHTOP304860OP304882OP981627OQ101843Wongkanoun et al. (2023)Pyrenopolyporus hunteriMUCL 52673Ivory CoastETKY610421KY610472KY624309KU159530Kuhnert et al. (2017a; tub2), Wendt et al. (2018; ITS, LSU, rpb2)Pyrenopolyporus laminosusMUCL 53305MartiniqueHTKC968934KY610485KY624303KC977292Kuhnert et al. (2014; ITS, tub2), Wendt et al. (2018; LSU, rpb2)Pyrenopolyporus macrosporusBCC89373ThailandHTOP304870OP304879OP981621OQ101844Wongkanoun et al. (2023)Pyrenopolyporus nicaraguensisCBS 117739Burkina Faso AM749922KY610489KY624307KC977272Bitzer et al. (2008; ITS), Kuhnert et al. (2014; tub2), Wendt et al. (2018; LSU, rpb2)Rhopalostroma angolenseCBS 126414Ivory Coast KY610420KY610459KY624228KX271277Wendt et al. (2018)Ruwenzoria pseudoannulataMUCL 51394D. R. CongoHTKY610406KY610494KY624286KX271278Wendt et al. (2018)Thamnomyces dendroideusCBS 123578French GuianaHTFN428831KY610467KY624232KY624313Stadler et al. (2010; ITS), Wendt et al. (2018; LSU, rpb2, tub2)Xylaria arbusculaCBS 126415Germany KY610394KY610463KY624287KX271257Fournier et al. (2011; ITS), Wendt et al. (2018; LSU, rpb2, tub2)Xylaria hypoxylonCBS 122620SwedenETKY610407KY610495KY624231KX271279Wendt et al. (2018)Type specimens are labelled with HT (holotype), ET (epitype), and PT (paratype). Isolates/sequences in bold were isolated/sequenced in present study.
For the phylogenetic analyses, 120 accessions from 107 species of Hypoxylaceae and four outgroup taxa from Graphostromataceae (Biscogniauxia nummularia, Graphostroma platystomum) and Xylariaceae (Xylaria arbuscula, X. hypoxylon) were included. The sequence matrices of ITS, LSU, rpb2, and tub2 were combined; after exclusion of ambiguously aligned and gap-rich regions, the resulting combined data matrix contained 3,863 alignment positions from four loci (547 from ITS, 942 from LSU, 1,045 from rpb2, and 1,329 from tub2).
Maximum likelihood (ML) analyses were performed with RAxML as implemented in raxmlGUI v. 2.0 (Edler et al. 2021) using the ML + rapid bootstrap setting with 1,000 bootstrap replicates and the GTRGAMMA substitution.
Maximum Parsimony (MP) analyses were performed with PAUP v. 4.0a169 (Swofford 2002). All molecular characters were unordered and given equal weight; analyses were performed with gaps treated as missing data; the COLLAPSE command was set to MINBRLEN. MP analysis of the combined multilocus matrix was done using 1,000 replicates of heuristic search with random addition of sequences and subsequent TBR branch swapping (MULTREES option in effect, steepest descent option not in effect). Bootstrap analyses with 1,000 replicates were performed in the same way, but using 10 rounds of random sequence addition and subsequent branch swapping during each bootstrap replicate. Bootstrap values ≤ 70% are considered low, between 70% and 90% intermediate, and ≥ 90% high.
HPLC profiling
2.4.
Stromata of Hypoxylon and Annulohypoxylon specimens were extracted essentially as described in Kuhnert et al. (2017a). Briefly, stromatal tissue was scraped with a spatula and collected in an Eppendorf tube, covered with acetone, centrifuged and 60–100 µL collected for subsequent HPLC-DAD-UV/Vis-ESI-MS analysis. System parameters and settings were identical to Kuhnert et al. (2014). Resulting spectra were analysed using DataAnalysis V4.4 (Bruker Daltonics) and compared with internal databases for identification of discernible compound peaks and stromata-fingerprints.
Extraction and isolation of secondary metabolites
2.5.
To study the chemical constituents identified by HPLC fingerprinting as described above, stromata of Hypoxylon lateripigmentum (GUM 1594, GUM 1595), Annulohypoxylon michelianum (KR-M-0002761), and A. substygium (GUM 1621, GUM 1619, GUM 1620, GUM 1624) were scraped and extracted by covering the material with acetone, followed by evaporation and transfer to a separate vial. This process was repeated three times, yielding in total 40 mg (GUM 1594; I), 26.1 mg (GUM 1595; II), 32.4 mg (STMA 16011; III), and 516.5 mg (GUM 1621, GUM 1619, GUM 1620, and GUM 1624; IV).
Extract I was purified on a reversed-phase (RP) HPLC system (Gilson, Middleton, WI, USA; GX-271 liquid handler, two pumps; 305 and 306, DAD wavelengths set to 220, 255, and 350 nm) using a C18-Gemini 10 µm; 250 × 21 mm column (A, Acetonitrile; B, H_2_O; flow rate set to 20 mL/min). The mobile phase was set to 50% B in isocratic conditions for 3 min, then increasing to 65% B in 45 min, another 2 min to 100% B and 100% B isocratic condition for 10 min. This yielded 3.9 mg of compound 2 at RT = 19.4–22.7 min and 0.3 mg of 4 at RT = 40.1–41.1 min.
Extract II was purified on the RP-HPLC system described above, with identical settings for the mobile phase. This yielded 0.3 mg of 3 at RT = 8.7–9 min, 0.8 mg of compound 1 at RT = 10.4 min, and 1.2 mg of 2 at RT = 11.4 min.
Extract III was purified using the same equipment, with the mobile phase set to 55% B in isocratic conditions for 3 min; increasing B to 85% in 40 min, then increasing to 100% in 3 min and 10 min of 100% B at again isocratic condition. This yielded 0.5 mg of compound 3 at RT = 13.2–14.5 min, 2.2 mg of 1 at RT = 19.1–20 min, 0.5 mg of 5 at RT = 27.7–28.4 min, and 2.6 mg of minutellin D (6) at RT = 33–34 min.
Extract IV was pre-purified by open normal-phase (NP) chromatography using 20 g of packed silica gel 60 (0.05–0.2 mm). Subsequent extraction with in total eight different solvent mixtures followed: 200 mL of 100% dichloromethane (DCM); 200 mL of 98% DCM and 2% acetone; 200 mL of 95% DCM and 5% acetone; 200 mL of 93% DCM and 7% acetone; 200 mL of 90% DCM and 10% acetone; 200 mL of 80% DCM and 20% acetone; 200 mL of 100% acetone and 50 mL of 100% methanol. This process yielded 40.1 mg of fraction V following extraction with 90% DCM and 10% acetone, which was further purified using the same RP-HPLC equipment as stated above. The mobile phase was set to 15% B at isocratic condition for 10 min; increasing to 25% in 15 min, remaining isocratic for 5 min; increasing to 40% B in 15 min and increasing to 80% B for 15 min. This yielded 0.7 mg of compound 7 at RT = 20.4–21.7 min and 1.7 mg of 8 at RT = 25–26 min.
Spectral data
2.6.
Minutellin E (1): yellow oil; +62 (c 0.1, acetonitrile); CD (acetonitrile): λ (Δε) 213 (+66.90), 275 (−42.40), 354 (+39.44), 417 (−25.80); UV-Vis (acetonitrile): λmax (log ε) 193 (0.62), 229 (0.58), 267 (0.64), 339 (0.56); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, acetone-d6) see Tables 1 and 2; HR-ESI-MS: m/z 495.2378 [M+H]^+^ (calcd. 495.5840 for C_29_H_35_O_7_^+^); tR = 15.26 min (HR-ESI-MS); C_29_H_34_O_7_ (494.5761 g mol^−1^).Table 2.^1^H data (700 MHz) of 1–5.Pos.1^a^2^b^3^b^4^a^5^a^18.77, br s8.81, br s6.87, br s8.74, s7.16, m46.40, s6.10, s6.08, s6.49, s6.42, s55.40 d (1.1)5.32 d (1.1)5.46 d (1.1)5.31, m5.44 d (1.1)8 3.32, dt (9.7, 1.9) 3.32, dt (9.5, 2.3)91.67, s1.73, s1.12, s1.67, s1.07, s10 2.97, d (12.1) 3.46, d (12.5) 122.73, dd(15.9, 3.7)2.54, dd(15.9, 7.3)2.91, ddd(13.5, 5.0, 2.0)2.46, dd (13.5, 11.6)2.81, m2.61, m6.05, dd(10.1, 2.2)6.82, d (8.1)134.33, m4.07, tt (11.5, 4.7)4.39, m7.18, m7.19, t (8.1)142.90, m2.63, dd (18.4, 6.1)2.35, m1.62, m2.81, m2.62, m2.63, m2.32, ddt(19.0, 10.8, 2.6)6.79, br d (8.1)15 2.13, m 2.56, m 162.11, s1.09, d (6.5)2.06, s1.06, d (6.7)2.23, s18 3.19, dd(17.2, 2.2)2.72, dd(17.2, 9.7) 3.12, dd(17.9, 2.3)2.99, dd(17.9, 9.5)203.11, ddd(18.3, 8.3, 6.3)2.88, m3.20, ddd(18.3, 9.0, 6.2)2.83, ddd(18.3, 8.9, 5.5)2.54, m3.09, m2.89, m2.65, t (7.4)211.60, m1.63, m1.56, m1.61, m1.59, m1.61, m22–251.20–1.35, m1.23–1.33, m1.23–1.35, m1.20–1.35, m1.26–1.34, m261.27, m1.24, m1.26, m1.26, m1.26, m271.29, m1.28, m1.30, m1.29, m1.29, m280.87, t (7.1)0.87, t (7.1)0.89, t (7.1)0.87, t (6.9)0.87, t (7.1)^a^ measured in acetone-d6, ^b^ measured in CHCl_3_-d.
Minutellin F (2): yellow amorphous solid; -39 (c 0.3, acetonitrile); CD (acetonitrile): λ (Δε) 213 (+42.90), 274 (−26.61), 349 (+30.38), 414 (−21.94); UV-Vis (a33cetonitrile): λmax (log ε) 262 (1.14), 336 (0.91); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, CDCl_3_-d) see Tables 1 and 2; HR-ESI-MS: m/z 497.2533 [M+H]^+^ (calcd. 497.599 for C_29_H_37_O_7_^+^), 529.2425 [M+Na]^+^ (calcd. 529.5817 for C_29_H_36_NaO_7_^+^); tR = 15.49 min (HR-ESI-MS); C_29_H_36_O_7_ (496.5919 g mol^−1^).
Minutellin G (3): yellow amorphous solid; -39 (c 0.1, acetonitrile); CD (acetonitrile): λ (Δε) 203 (+12.06), 249 (−6.80), 323 (+17.95), 422 (−1.54); UV-Vis (acetonitrile): λmax (log ε) 229 (0.299), 345 (0.30); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, CDCl_3_-d) see Tables 1 and 2; HR-ESI-MS: m/z 471.2743 [M+H]^+^ (calcd. 471.6057 for C_28_H_39_O_6_^+^); tR = 14.14 min (HR-ESI-MS); C_28_H_38_O_6_ (470.5977 g mol^−1^).
Minutellin H (4): yellow amorphous solid; -39 (c 0.1, acetonitrile); CD (acetonitrile): λ (Δε) 212 (+9.49), 272 (−8.15), 335 (+6.09), 415 (−3.83); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, CDCl_3_-d) see Tables 1 and 2; HR-ESI-MS: m/z 479.2434 [M+H]^+^ (calcd. 479.5846 for C_29_H_35_O_6_^+^), 501.2246 [M+Na]^+^ (calcd. 501.5664 for C_29_H_34_NaO_6_^+^); tR = 14.83 min (HR-ESI-MS); C_29_H_34_O_6_ (478.5767 g mol^−1^).
Minutellin I (5): yellow amorphous solid; -39 (c 0.1, acetonitrile); UV-Vis (acetonitrile): λmax (log ε) 348 (0.076); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, CDCl_3_-d) see Tables 1 and 2; m/z 453.2645 [M+H]^+^ (calcd. 453.5904 for C_28_H_37_O_5_^+^), 475.2453 [M+Na]^+^ (calcd. 475.5722 for C_28_H_36_NaO_5_^+^); tR = 14.15 min (HR-ESI-MS); C_28_H_36_O_5_ (452.5824 g mol^−1^).
Daldinone L (8): Brown amorphous solid; +275 (c 0.1, acetonitrile); NMR data (^1^H: 700 MHz, ^13^C: 175 MHz, CDCl_3_-d) see Table 4; m/z 335.0911 [M-H_2_O+H]^+^ (calcd. 335.3301 for C_20_H_15_O_5_^+^), 353.1016 [M+H]^+^ (calcd. 353.3454 for C_20_H_17_O_6_^+^), 375.0837 [M+Na]^+^ (calcd. 375.3272 for C_20_H_16_NaO_6_^+^); tR = 7.05 min (HR-ESI-MS); C_20_H_16_O_6_ (352.374 g mol^−1^).
Antimicrobial activity of compounds 1–5
2.7.
Compounds 1–5 were subjected to a serial dilution assay in 96-well microtiter plate format to determine the minimal inhibitory concentration (MIC) against Escherichia coli (DSM 1116), Bacillus subtilis (DSM 10), and Mucor hiemalis (DSM 2656) essentially as described by Surup et al. (2015).
Results
Molecular phylogeny
3.1.
Of the 3,863 characters of the combined matrix, 1,734 were parsimony informative (345 in ITS, 201 in LSU, 515 in rpb2, and 673 in tub2). The phylogram of the best ML tree (lnL = −107,247.1419) obtained by RAxML is shown as Figure 1. Maximum parsimony analyses resulted in five parsimonious tree [tree length = 24,727; Consistency Index (CI) = 0.158; Retention Index (RI) = 0.564 and Homoplasy Index (HI) = 0.842] (not shown). The alignment is provided as Table S1 in the Supporting Information. Figure 1.Phylogram of the best ML trees (lnL = −107,247.1419) revealed by RAxML from an analysis of the combined ITS-LSU-rpb2-tub2 matrix of selected Hypoxylaceae. Strains in bold were sequenced in the current study. ML and MP bootstrap support above 50% are given at the first and second positions, respectively, above or below the branches.
The phylogenies revealed a paraphyly of Hypoxylon, with the genera Annulohypoxylon, Daldinia, Durotheca, Entonaema, Jackrogersella, Hypomontagnella, Parahypoxylon, Phylacia, Pyrenopolyporus, Rhopalostroma, Ruwenzoria, and Thamnomyces embedded within the former, as reported in previous studies (e.g. Pourmoghaddam et al. 2020; Lambert et al. 2021; Cedeño-Sanchez et al. 2023). All of the latter genera appeared monophyletic except for Daldinia (Figure 1).
In our phylogenetic tree, Hypoxylon resolved into seven groups (H1–H7 in Figure 1). The Annulohypoxylon and Hypoxylon species examined in this study clustered in two subclades (A and H1). All major groups and deeper, highly supported nodes were consistent between the ML and MP analyses, but topologies of deeper unsupported nodes differed in the MP tree; as these differences are not relevant within the context of our species, they are not further considered here.
In subclade A, the sequences of all Iranian strains of Annulohypoxylon substygium clustered together with maximum support and were shown to be almost identical to those of the ex-epitype culture (MUCL 51708) and A. substygium (STMA 14066).
In subclade H1, the Iranian Hypoxylon species clustered as sister species to H. lateripigmentum (MUCL 53304) with maximum ML and MP BS support, and both form a sister group to H. lignicola (MFLUCC 16-0926), also with maximum ML and MP BS support (Figure 1).
HPLC profiling
3.2.
The HPLC chromatograms of Hypoxylon lateripigmentum specimens GUM 1594, GUM 1595 (shown in Figure 2), and GUM 1596 (not shown) appeared to be highly similar, containing severable compounds with UV/Vis absorption patterns resembling the minutellin-type azaphilone family. However, the compounds were not identifiable. The bottom panel of Figure 2 shows a stromatal acetone extract of the holotype of H. lateripigmentum, which confirms the phylogenetic affinities of the isolated strains. Figure 2.HPLC-UV/Vis chromatogram (210 nm) of an acetone stroma extract of specimen GUM 1594 and GUM 1595 (top and middle panel) and the holotype of Hypoxylon lateripigmentum, MJF10046 (below). Right panels: UV/Vis absorption spectra of designated compounds.
The HPLC chromatogram of Annulohypoxylon substygium specimens GUM 1620 (Figure 3), GUM 1618, GUM 1619, GUM 1621, GUM 1622, GUM 1624 (not shown) appeared practically identical, featuring two compounds (7 and 10) in minor and one compound (9) in higher abundance. Compounds 9 and 10 could be identified as daldinone A and BNT, respectively, while compound 7 showed affinities to the daldinone family. Figure 3.Representative HPLC-UV/Vis chromatogram (210 nm) of an acetone stroma extract of specimen Annulohypoxylon substygium (GUM 1620). Right panel: UV/Vis absorption spectra of designated compounds. Compounds 7, 9, and 10 were identified as daldinone F, daldinone A, and BNT, respectively.
As we had access to a considerable amount of stromata for chemical extraction, we next attempted to characterise the unknown components detected in the specimen identified as H. lateripigmentum and potential novel minor components in A. substygium. Concurrently, we analysed the stromatal constituents of a specimen of A. michelianum (KR-M-0002761) that was reported to feature similar minutellin-type azaphilones (Kuhnert et al. 2017a), which we hence included in this study.
Structure elucidation of minutellins 1–6
3.3.
Minutellin E (1) was isolated as a yellow oil; its molecular formula C_29_H_34_O_7_ was deduced from the molecular ion cluster at m/z 495.2378 in the HR-ESI-MS spectrum. ^1^H and HSQC NMR spectra revealed the presence of three olefinic methine, six resolved methylene and two methyl signals (Tables 2 and 3). Further unresolved methylenes in the region of δH 1.30/δC 30.0 ppm indicated a long fatty acid side chain. NMR data including chemical shifts of 1 were virtually identical to those of minutellin A, so a common scaffold including stereochemistry was assigned (Kuhnert et al. 2017b). However, the molecular formula of 1 entails two additional methylene carbons in the fatty acid side chain compared to minutellin A.Table 3.^13^C data (175 MHz) of 1–5.Pos.1^a^2^b^3^b^4^a^5^a^1154.4, CH153.5, CH145.1, CH153.4, CH146.5, CH3154.9, C156.7, C152.9, C158.3, C^x1^155.5, C4113.8, CH111.9, CH111.8, CH112.1, CH112.0, CH4a144.5, C143.1, C146.7, Cn.o.148.4, C5106.4, CH105.8, CH105.4, CH105.5, CH105.4, CH6190.7, C190.5, C199.2, C190.4, C199.0, C788.4, C87.7, C73.1, C87.8, C73.7, C8165.9, C165.0, C40.6, CH165.1, C41.6, CH8a112.0, C111.2, C120.5, C111.2, C120.9, C926.3, CH_3_26.3, CH_3_21.3, CH_3_25.6, CH_3_21.6, CH_3_10130.7, C64.5, CH130.8, C59.3, CH121.2, C11194.8, C202.8, C193.7, C194.6, C156.3, C1247.1, CH_2_50.4, CH_2_46.3, CH_2_128.7, CH114.4, CH1365.8, CH67.8, CH65.4, CH151.1, CH131.8, CH1441.9, CH_2_42.4, CH_2_40.9, CH_2_33.9, CH_2_122.3, CH15162.6, C30.6, CH160.1, C33.3, CH139.6, C1623.0, CH_3_20.5, CH_3_23.0, CH_3_19.4, CH_3_20.1, CH_3_17169.3, C168.0, C 167.8, C 18124.6, C123.7, C38.9, CH_2_124.0, C39.2, CH_2_19197.6, C197.2, C209.8, C197.0, C209.3, C2042.4, CH_2_42.2, CH_2_42.8, CH_2_41.8, CH_2_43.2, CH_2_2124.2, CH_2_23.3, CH_2_24.0, CH_2_23.6, CH_2_24.6, CH_2_2229.9, CH_2_29.0, CH_2_29.2, CH_2_29.0, CH_2_23–2529.2, 29.40, 29.4129.2, 29.38, 29.40**2632.7, CH_2_31.8, CH_2_31.8, CH_2_32.1, CH_2_32.7, CH_2_2723.4, CH_2_22.6, CH_2_22.6, CH_2_22.8, CH_2_23.4, CH_2_2814.4, CH_3_14.1, CH_3_14.1, CH_2_13.8, CH_3_14.4, CH_3_* overlapped by solvent signal, n.o.: not observed, x1: chemical shift extracted from HMBC crosspeak, ^a^ measured in acetone-d6, ^b^ measured in CHCl_3_-d.
For minutellin F (2), the molecular formula of C_28_H_40_O_6_, generated from the molecular ion cluster at m/z 497.2533 in the HR-ESI-MS spectrum, indicated the formal addition of two hydrogen atoms compared to 1. NMR data of 2 were similar to those of 1 with these key differences: two methines replaced the olefinic carbons C–10 and C–16, additionally the methyl CH_3_–16 multiplicity in the ^1^H NMR spectrum changed to doublet. Consequently, a 2-oxo-4-hydroxy-6-methylhexanyl- moiety was deduced for 2. The large coupling constant J10,15 = 12.1 hz indicated a trans configuration of these protons with pseudo-axial orientations, and the ROESY correlation between H–15 and H–13 confirmed H–13 to be also axial and on the same side as H–15.
Minutellin G (3) had the molecular formula C_28_H_38_O_6_, indicating the formal loss of one carbon and one oxygen atom and the formal addition of four protons compared to 1. 1D and 2D NMR data indicated the loss of C–17 and the replacement of olefinic carbons C–8 and C–18 by a methine and a methylene, respectively, as key differences.
Minutellin H (4) resembled 2 in its NMR data, with the exemption of the replacement of methylene C–12 and oxymethine C–13 by the δ^12,13^ double bond. This modification was confirmed by the molecular formula C_29_H_34_O_6_, denoting a loss of H_2_O.
The molecular formula C_28_H_36_O_5_ of minutellin I (5) indicated a formal loss of H_2_O compared to 3. The 2-hydroxy-6-methyl-benzyl moiety was confirmed by the aromatic signals for H–12 to H–14 in the ^1^H and C–10 to C–15 in the ^13^C spectrum.
Minutellin D (6) was identified based on comparison of its NMR and HRMS data to the literature (Kuhnert et al. 2017b). Analogous to other minutellins, the CD curves of 1–5 with local minima around 250 and 275 nm and a characteristic maximum around 350 nm confirm a 7R absolute configuration (Pyser et al. 2019).
Structure elucidation of daldinones 7–9
3.4.
Daldinone F (7) was identified by comparison of their NMR and HRMS data with previous reports (Wang et al. 2015). However, a close inspection of the NMR data of 7 revealed that various signals had been assigned incorrectly by Wang et al. (2015). Most notably, signals for H_2_-8 and H_2_-18 had been confused. However, chemical shift analysis as well as COSY and HMBC correlations (see Figure S32 for details) require the reassignment of the signals for C–1, C–4, C–8, H_2_–8, C–11, C–15, C–17, H_2_–17, C–20 (Table 4).Table 4.NMR data of daldinones F (7^b^; 700 MHz, CHCl_3_-d) and L (8; 700 MHz, CHCl_3_-d), comparison with data of Wang et al. (2015). 7^a^as assigned by Wang et al. (2015)7^b^as assigned in this study8^b^Pos.δC, mult.δH, mult.δC, mult.δH, mult.δC, mult.δH, mult.1142.8 120.3, C 115.9, C 2161.7OH: 12.39161.5, COH: 12.41, br s156.2, COH: 11.85, s3115.56.82, d (8.5)117.5, CH6.89, m118.5, CH6.77, d (9.0)4123.87.45, d (8.5)137.7, CH7.50, ps t (8.1)127.6, CH7.17, d (9.0)5117.3****6.87, d (8.5)120.3, CH7.20, dd (7.6, 1.0)148.5, COH: 8.79, br s6143.2 143.0, C 128.2, C 7203.2 203.2, C 204.4, C 827.2****2.42, dd (13.5, 4.0)2.24 (dd, 13.5, 4.9)36.1, CH_2_3.26, dd (17.4, 5.6)2.88, dd (17.4, 2.0)37.3, CH_2_3.27, dd (16.4, 5.0)2.85, dd (16.4, 2.6)956.22.46–2.5156.5, CH2.68, ddd (11.0, 5.6, 2.0)55.6, CH2.68, ddd (10.3, 5.0, 2.6)1079.8 79.5, C 82.0, COH: 5.45, br s11120.2 137.7, C 138.0, C 12123.87.55, d (8.0)123.9, CH7.54, d (8.2)135.2, CH8.17, d (8.6)13120.1****7.21, d (8.0)115.6, CH6.86, br d (8.2)116.1, CH6.76, d (8.6)14156.7 156.4, C 161.7, COH: 11.15, s15138.0 120.3, C 114.4, C 1669.85.08, q (8.5)69.5, CH5.10, m204.9, C 1735.8****3.24, dd (15.0, 5.5)2.85, d (15.0)33.8, CH_2_2.42, m38.6, CH_2_2.80, m2.64, m1833.7****1.69, q (8.0)1.47, q (8.0)27.3, CH_2_2.05, m1.46, m29.3, CH_2_2.49, m1.83, m1942.52.59 (dt, 11.0, 4.5)42.7, CH2.89, td (11.0, 4.5)43.1, CH2.89, ddd (11.2, 10.3, 4.5)20114.4 142.6, C 151.0, C ^a^ measured in CHCl_3_-d, ^b^ measured in acetone-d6. Previously misassigned signals are highlighted in bold.
The NMR data of compound 8 was highly similar to those of daldinone F, with the replacement of an aromatic methine by an oxygenised sp^2^ hybridised carbon as well as oxymethine CH–16 by a keton. Consequently, 8 was elucidated as (6bS,12aR,12bR)-4,6b,7,10-tetrahydroxy-1,2,6b,12,12a,12b-hexahydrobenzo[j]fluoranthene-3,11-dione, for which we propose the trivial name daldinone L.
^1^H and ^13^C NMR data of 9 matched those of not only daldinone A (Quang et al. 2002), but also daldinone C (Gu et al. 2007). This was surprising, since opposite stereo configurations of C–19 had been published for these metabolites. We addressed the stereochemistry of 9 by ROESY NMR data and observed a strong TOCSY artefact between H–9 and H–19. We conclude that Quang et al. (2002) misinterpreted this TOCSY artefact as a conventional cross peak, since they wrote: “NOE correlations were observed between … H–9 and H–19 … ” Based on this misinterpretation, a wrong relative stereochemistry was assigned between these protons. Gu et al. (2007) on the other hand assigned the correct structure, confirmed by X-ray data, but unfortunately did not realise that they had reisolated daldinone A and coined the name daldinone C instead of revising the structure of daldinone A. Additionally, the absolute stereochemistry of daldinone C was revised by Podlech and Gutsche (2023) by reinterpretation of CD data.
Taken together, we propose that daldinone A and C are identical and suggest only using the name daldinone A with its revised structure in the future.
Bioactivity of compounds 1–5
3.5.
Subsequent testing for antimicrobial activity of compounds 1–5 revealed moderate activity exerted by 5 against B. subtilis (MIC = 8.34 µg/mL). No growth inhibitory activity was recorded for compounds 1–5 against the other tested organisms.
Production of minutellins
3.6.
We realised that the stroma of the neotropical H. pulicicidum was reported to harbour similar minutellin-type azaphilones and since we had access to the holotype specimen, compared the stromatal constituents of the holotype with our pure standards. We were able to correlate in total three detectable compounds, i.e. minutellin A, C, and D with components of its stromatal extract (Figure S39). Table 5 summarises the distribution of the analysed minutellin-derivatives detected in the here studied specimen of Hypoxylon and Annulohypoxylon. Table 5 summarises the distribution of the analysed minutellin-derivatives detected in the here studied specimen of Hypoxylon and Annulohypoxylon and Figure 4 shows the structures of all discussed compounds in this section. Figure 4.Structures of the new minutellins E–I (1–5), minutellin D (6), daldinone F, and its new derivative L (7 and 8), revised structure of daldinone A (9), binaphthalene-tetrol (BNT; 10), and minutellins A and C (11 and 12) discussed in the present study.Table 5.Minutellins E–I, A, C, and D (1–5, 11, 12, 6) detected in Annulohypoxylon michelianum, Hypoxylon lateripigmentum, and H. pulicicidum stromatal extracts.SpecimenSpecies1234561112KR-M-0002761A. michelianum**+–+–++**––GUM 1594H. lateripigmentum++++–+**––GUM 1595H. lateripigmentum++++–+**––GUM 1596H. lateripigmentum++++–+**––MJF10046H. lateripigmentum++++–+**––MJF07147H. pulicicidum–––––+++**
Taxonomy
3.7.
Hypoxylon lateripigmentum J. Fourn., Kuhnert & M. Stadler, Fungal Diversity 64: 192 (2014). Figures 5 and 6. Figure 5.Hypoxylon lateripigmentum (GUM 1594). (a, b) Close-up view of stromatal surface, with stromatal pigments in 10% KOH. (c) Stroma in vertical section showing perithecia and ostioles. (d) Immature and mature asci in water. (e) Mature ascus tip in Melzer’s reagent. (f) Ascospores in 10% KOH with dehiscent perispore with thickened (black arrow). (g) Ascospore in water, with straight germ-slit. Scale bars: a = 5 mm, b = 1 mm, c = 0.5 mm, d = 20 µm, e – g = 10 µm. Figure 6.Culture and anamorphic structures of Hypoxylon lateripigmentum (MUCL 57716) on OA. (a) Surface of colony after 6 weeks of incubation. (b, c) General view of anamorph structure with periconiella-like branching patterns, conidiogenous cells, immature and mature conidia. (d) Conidia. Scale bars: b, c = 20 µm, d = 10 µm.
Teleomorph. Stromata superficial, effused-pulvinate, up to 2 cm long × 0.6–1.1 cm wide, surface Vinaceaous (57) to Dark Brick (60) becoming black with conspicuous perithecial mounds; Brown Vinaceaus (84) granules beneath the surface and between the perithecia, with KOH-extractable pigments Red (2) to Scarlet (5). Perithecia obovoid to tubular, 0.36–0.43 mm high × 0.23–0.35 mm wide. Ostioles umbilicate, inconspicuous. Asci 8-spored, cylindrical, with amyloid, discoid apical apparatus, 0.5–0.75 µm high × 1.5–2 µm wide, stipe up to 190 µm, and spore-bearing portion 60–85 × 4.5–6 µm. Ascospores smooth, unicellular, brown to dark brown, ellipsoid, inequilateral to slightly equilateral with narrowly rounded ends, 8–12(−13) × 3.5–5 µm, with straight to rarely sigmoid germ slit spore-length on convex side; perispore dehiscent in 10% KOH, smooth with a thickening on one or both side; epispore smooth.
Cultures and anamorph. Colonies on OA covering a 9 cm Petri dish in 2 wk, at first white, becoming pale brown, felty, zonate with diffuse margins; reverse becoming Hazel (88). Conidiogenous structure approaching a periconiella-like branching pattern as defined by Ju and Rogers (1996), (Figure 6b,c). Conidiophores hyaline, smooth to finely roughed. Up to three conidiogenous cells at the end of each terminus; hyaline, smooth to finely roughened, 11–16 × 2–3.5 µm. Conidia hyaline, smooth, ellipsoid to obovoid, 4–7 × 2–4 µm.
Secondary metabolites. New minutellin-type azaphilones minutellin E–H and minutellin D.
Specimens examined. Iran, Guilan Province, Shaft County, Rahimabad (Sefidab) forest, 36°34’10.94“N, 50°20’26”E, 400 m elev., on fallen branch of Parrotia persica, 21 September 2016, leg. M.J. Pourmoghaddam (GUM 1594; living culture MUCL 57716); Guilan Province, Shaft County, Rahimabad (Sefidab) forest, 36°34’10.94“N, 50°20’26”E, 400 m elev., on fallen branch of Pterocarya fraxinifolia, 21 September 2016, leg. M.J. Pourmoghaddam (GUM 1595; living culture MUCL 57717); Guilan Province, Rezvanshahr County, 37°37’52“N, 40°02’18”E, 7 m elev., on fallen branch of Gleditschia caspica, 6 October 2016, leg. M.J. Pourmoghaddam (GUM 1596; living culture MUCL 57718).
Note. These specimens show high similarities to H. lateripigmentum in teleomorphic and anamorphic appearance. Nonetheless, Iranian specimens have variations in the size of perithecia (0.36–0.43 × 0.23–0.35 vs. 0.6–0.7 × 0.35–0.5 mm) and ascospores (8–13 × 3.5–5 vs. 7.7–9.5 × 3.8–4.2 µm). A comparison of Iranian sequence data revealed differences between the three Iranian strains of H. lateripigmentum (MUCL 57716/MUCL 57717/MUCL 57718) and the ex-type strain of H. lateripigmentum [19/8/19 bp differences of 432 nucleotide characters in the ITS: 6/6/6 substitutions, 13/2/13 indels; 6/6/6 bp differences (substitutions) of 1,249 nucleotide characters in the LSU; 11/11/11 bp differences (substitutions) of 942 nucleotide characters in the rpb2; and 36/43/36 bp differences of 1,216 nucleotide characters in the tub2: 32/37/33 substitutions, 4/6/3 indels]. Nonetheless, chemotaxonomic analysis showed virtually identical chemotypes when compared with an ex-holotype-derived acetone extract, leading us to assign these specimens to H. lateripigmentum. This ought to be changed when a larger number of isolates becomes available to further delineate potential differences in population studies. Hypoxylon lignicola and H. phuphaphetense have recently been published according to asexual morphs and there is no evidence of a sexual morph in the latter species (Luo et al. 2019). Table 6 compares morphological characters of some other taxa that may be confused with this species.Table 6.Diagnostic characters of Hypoxylon lateripigmentum and closely related species.TaxonStromatal shapeStromatal surfaceKOH extractable pigmentsAscospores (µm)Germ slitPerispore in KOHAnamorphSecondary metabolitesHypoxylon croceumGlomerate to pulvinateFulvous when young, Sepia or Fuscous when agedIsabelline or Hazel8.5–12(−13.5) × 4–5Straight less than spore-lengthIndehiscentVirgariella-likeUnknown minutellin-type azaphilonesHypoxylon hinnuleumGlomerate to pulvinate or effused-pulvinateDark Vinaceous to Sepia, becoming BlackDark Brick(7.2–)7.6–9.5(−11.7) × (2.8–)3.7–4.5(−4.8)Straight, 2/3 spore lengthIndehiscentVirgariella-like, nodulisporium-like, and periconiella-likeBNT, minutellin D, various unknown cohaerinsHypoxylon investiensEffused-pulvinateBrown Vinaceous, Sepia, Dark Brick, or ChestnutGreenish Yellow, Dull Green, or Dark Green6.5–9.5(−10) × 3–4.5Straight less than to spore-lengthIndehiscentPericoniella-likeBNT, daldinone A, 5-methylmelleinHypoxylon lateripigmentumEffused-applanateDark VinaceousSienna to Bay7.7–9.5 × 3.8–4.2Straight spore-lengthDehiscentPericoniella-likeBNT, new minutellins 1–4 and minutellin D(6), 1–8-naphthol**Iranian Hypoxylon lateripigmentum specimens described hereEffused-pulvinateVinaceaous to Dark Brick, becoming blackRed to Scarlet8–12(−13) × 3.5–5Straight to rarely sigmoid spore-lengthDehiscentPericoniella-like1–4 and minutellin D (6)**Hypoxylon olivaceopigmentumGlomerate to pulvinate or effused-pulvinateBrown Vinaceous, becoming blackOlivaceous(9.3–)9.7–11.9(−15.7) × (4.5–) 5.0–6.6(−7.2)Straight 1/2 to 2/3 of spore lengthIndehiscentPericoniella-likeBNT, an unknown cohaerin-type compound with m/z = 498Hypoxylon pulicicidumIrregularly effused-pulvinate to elongateDark Vinaceous to Brown VinaceousOlivaceous Buff7.5–9.4 × 3.2–4.2Straight less than to spore-lengthIndehiscentNodulisporium-likeBNT, minutellins A, C, and DSpecimens are in bold described in present study.
Annulohypoxylon substygium Sir & Kuhnert, Fungal Diversity 85: 20 (2017). Figure 7 Figure 7.Annulohypoxylon substygium (GUM 1618). (a) Stromatal habit. (b, c) Close-up view of stromatal surface. (d, e) Close-up view on ostiolar discs. (f, g) Stromatal pigments in 10% KOH. (h) Immature and mature asci in water. (i) Ascus tip in Melzer’s reagent. (j) Mature ascus in water. (k, l) Ascospores in 10% KOH with dehiscent perispore showing thickening. Scale bars: b = 4 mm, c = 1 mm, d = 0.5 mm, e = 0.3 mm, h, j = 20 µm, i, k, l = 10 µm.
Teleomorph. Stromata superficial, effused-pulvinate; up to 20 cm long × 0.2–3 cm wide; with inconspicuous perithecial mounds; surface Vinaceous Gray (116) when young, becoming blackish with Brown Vinaceous (84) to Fawn (87), black when old, pruinose; blackish granules immediately beneath surface; with KOH-extractable pigments Greenish Yellow (16) or Fawn (87). Perithecia spherical to ovoid 0.3–0.65 mm high × 0.25–0.6 mm wide; ostioles conical papillate, encircled with a bovei-type disc 0.25–0.35 mm diam. Asci 8-spored, cylindrical, with weakly amyloid, discoid apical apparatus, 0.5 µm high × 1–1.5 µm wide, stipe up to 50 µm, and spore-bearing portion 65–75 × 4–5 µm. Ascospores light brown to brown, unicellular, ellipsoid nearly equilateral, with broadly rounded ends, 6.0–9.0(−10) × 2.5–3.5 µm, with straight germ slit spore-length on the more flattened side; perispore dehiscent in 10% KOH, smooth with a thickening on the more flattened side; epispore smooth.
Cultures and anamorph. Colonies on OA covering a 9 cm Petri dish in 2 wk, at first whitish, becoming Pale Green (19) in centre, Grey Olivaceous (107) and Flesh (37), whitish in margins, concentric zones, velvety to felty, with entire margins; finally, Isabelline (65) to Dark Brick (60). Conidiogenous structure branching periconiella-like as defined by Ju and Rogers (1996). Conidiophores brown to pale brown, smooth to finely roughened. Conidiogenous cell brown to pale brown, smooth to finely roughened, 12–17 × 2–3 µm. Conidia cylindrical to ellipsoid, hyaline to pale brown, smooth to finely roughened, 5–7 × 2–3 µm.
Secondary metabolites. BNT, daldinone A, daldinone F, the new daldinone L.
Specimens examined. Iran, Guilan Province, Lahijan County, Kuhbijar forest, 37°11ʹ26”N, 50°01ʹ46”E, 350 m elev., on fallen branch of Acer sp., 20 September 2016, leg. M.J. Pourmoghaddam (GUM 1618; living culture MUCL 57734); Guilan Province, Fouman County, Ghalerodkhan forest, 37°04ʹ05.02”N, 49°14ʹ40.08”E, 500 m elev., on fallen branch of Quercus castaneifolia, 29 October 2016, leg. M.J. Pourmoghaddam (GUM 1624; living culture IRAN 3727C); Guilan Province, Siahkal County, Deilaman forest, 36°57’25“N, 49°51’54”E, 1,100 m elev., on fallen branch of Quercus castaneifolia, 3 October 2016, leg. M.J. Pourmoghaddam (GUM 1619; living culture IRAN 3728C); Guilan Province, Rasht County, Saravan forest, 37°04’26“N, 49°38’13”E, 183 m elev., on fallen branch of Quercus castaneifolia, 3 October 2016, leg. M.J. Pourmoghaddam (GUM 1620); Guilan Province, Siahkal County, Ziaratgah forest, 37°08ʹ07”N, 49°55ʹ35”E, 263 m elev., on fallen branch of Quercus castaneifolia, 3 October 2016, leg. M.J. Pourmoghaddam (GUM 1622).
Note. Annulohypoxylon substygium was hitherto only reported from the northern hemisphere and the Neotropics (e.g. Argentina, France, Georgia, La Réunion) (Kuhnert et al. 2017a). Most of the characters of the Iranian specimens are in accordance with previous descriptions (Kuhnert et al. 2017a), aside from insignificant variations in the size of ascospores (6.0–10.0 × 2.5–3.5 vs. 6.5–8.5 × 2.8–3.5 µm) and in the colour of the KOH-extractable pigments (greenish yellow or fawn vs diluted fuscous black to fawn). A comparison of Iranian sequence data revealed similarities between the six Iranian strains of A. substygium (MUCL 57734/IRAN 3727C/IRAN 3728C/318/465/485) and the epi-type strain (MUCL 51708) [6/6/6/6/6/6 bp differences (indels) of 627 nucleotide characters in the ITS; and no differences of 1,346 nucleotide characters in the tub2]. We also included the newly sequenced LSU and rpb2 loci of this species for the first time. This species can be differentiated from A. stygium by morphological characters (larger ostiolar discs, larger perithecia, and KOH-extractable pigments), stromatal secondary metabolites (chemotaxonomy), and molecular phylogeny.
Discussion
In this study, we report on the identification and polyphasic characterisation of a collection of Iranian Annulohypoxylon and Hypoxylon spp. from near the Caspian Sea, and on a chemotaxonomic isolation campaign including both the newly collected specimens and a previously reported specimen of A. michelianum (Kuhnert et al. 2017a). A collection of Iranian Annulohypoxylon could unequivocally be assigned to A. substygium. The other specimen showed affinities to Hypoxylon lateripigmentum, but showed differences in the size of perithecia and ascospores (Table 6). Chemotaxonomic analysis revealed virtually identical chemotypes when compared with a holotype-derived acetone extract, which settled its identity as H. lateripigmentum.
During species identification, after thorough comparison with internal databases, featuring UV/Vis absorption maxima, retention time, and accurate mass of authentic standards (also called dereplication; see Bitzer et al. 2007), we noted the presence of several unidentifiable compounds. Concurrently, while examining the stromatal constituents of A. michelianum, we detected similar compounds, and decided to integrate these efforts into the current study. Subsequently, we isolated and herewith describe new azaphilones of the minutellin/cohaerin family from the stroma of A. michelianum and H. lateripigmentum. Structural key difference of the new minutellins 1–5 to their most closely known azaphilones relatives is the decanoyl fatty acid side chain moiety, in contrast to i.e. the 2-methyloctyl or the 2-methybutyl side chain of the cohaerins and multiformins, respectively (Surup et al. 2013; Jansen-Olliges et al. 2022). However, the decanoyl fatty acid side chain is also part of biscogniazaphilones A and B, which have been isolated from the endophytic fungus Biscogniauxia formosana (Cheng et al. 2012). Structural difference of 1–6 to them is the cyclohexyl moiety attached to C–3, in contrast to the propenyl residue of biscogniazaphilones A and B. Moreover, analogous variations of this cyclohexanyl ring structure with various patterns of hydrogenation or elimination/dehydration have been observed for cohaerins and multiformins, too.
Secondary metabolite production patterns, or chemotypes, have served as useful information for chemotaxonomic purposes and have also been shown to be specific for different lineages in the Hypoxylaceae. Orsellenic-acid derived mitorubrin-derivatives and rubiginosins, for instance, are widespread in the Hypoxylon rubiginosum complex (Stadler et al. 2008), and the H. fuscum complex features species producing BNT, daldinin derivatives, and other undescribed naphthalenes (Quang et al. 2004; Lambert et al. 2021). Moreover, members of Annulohypoxylon regularly include species that additionally contain truncatone-type derivatives, which are typically absent in its sister genus Jackrogersella (Kuhnert et al. 2017a; Wendt et al. 2018). The latter was even segregated from Annulohypoxylon by Wendt et al. (2018) based on phylogenetic and chemotaxonomic evidence, i.e. the presence of minutellin/cohaerin-type azaphilones, such as minutellins A–D reported from J. minutella (Kuhnert et al. 2017b) or cohaerins A–K from J. cohaerens (Quang et al. 2005, 2006; Surup et al. 2013). Occasionally however, chemotype and phylogenetic placement may differ. Recently, we proposed – based on comparisons with high-resolution HPLC-MS data – that the stroma of H. dussii contains mitorubrin-type derivatives (Cedeño-Sanchez et al. 2025). Concurrently, we sequenced an ex-type culture of this species, which then clustered in a phylogenetic lineage with H. investiens, H. pulicicidum and allies, in which these derivatives are absent. However, H. investiens was shown to contain naphthalene-type compounds, such as BNT and daldinone A usually encountered in Daldinia, or Annulohypoxylon (Stadler et al. 2014; Kuhnert et al. 2017a; Becker and Stadler 2021). Another interesting case is Annulohypoxylon michelianum, which had been shown to contain minutellin A, C, and D, identified based on a comparative approach (Kuhnert et al. 2017a). Morphologically, it shows typical characteristics of Annulohypoxylon and resolves in a basal position to other Annulohypoxylon. Together with the chemotaxonomic evidence, this taxon shows a rather intermediate phenotype between Annulohypoxylon and Jackrogersella (Wendt et al. 2018). It is currently not clear how the production of minutellin/cohaerin-type azaphilones has evolved and why this trait is missing in other Annulohypoxylon and Hypoxylon spp. phylogenetically related to H. investiens. A possible explanation could be the loss of secondary metabolite production capability of a former ancestor of the remaining Annulohypoxylon and a lineage that later evolved into H. investiens and related species. However, low bootstrap support, potentially due to a conflicting, underlying phylogenetic signal, for the final placement of H. investiens prevents any solid hypothesis associated with this phenomenon. It will be interesting in the future to study the recently sequenced genomes in detail to put these potential conflicts of phylogenetic signal into context, unravel evolutionary patterns of the secondary metabolism and further explore the evolutionary history of this clade.
Here, we report the results of an analysis and characterisation of stromatal constituents from environmental samples due to access to sufficient amounts for preparative work, but this is not always the case. In instances where stroma is scarce (or valuable types are studied), it is possible to assign compounds directly by comparison with a co-analysed authentic sample of a given natural product. This was, for example, done for stroma extracts derived from H. aeruginosum, Chlorostroma subcubisporum, and C. cyaninum, which were compared to lepraric acid derivatives (Læssøe et al. 2010). Unidentified, similar compounds to the ones reported here were previously reported from stromatal extracts of the predominantly endophytic H. pulicicidum (Bills et al. 2012). A comparison of the holotype with our compounds yielded identical UV/Vis and mass patterns, which lead us to the assignment of three distinct peaks to compounds 6, 11, and 12. Similar compounds had also been detected in H. croceum, H. hinnuleum, H. olivaceopigmentum (Sir et al. 2019) and Parahypoxylon (Cedeño-Sanchez et al. 2023), which can, in principle, be identified following a similar approach.
In a previous study, we tentatively assigned stromatal compounds of Parahypoxylon papillatum and P. ruwenzoriense to the minutellin azaphilone family based on molecular networking, which even suggested the presence of many other, not identified and never isolated derivatives (Cedeño-Sanchez et al. 2025). However, whether these compounds are indeed representing stable and isolatable compounds or merely represent degradation or extraction artefacts or instable intermediates is an open question. A full study based on stromatal material is unrealistic due to potentially requiring multi-gram scale of stroma material, but since candidate biosynthetic gene clusters are available (Kuhnert et al. 2021), the biosynthetic machinery can in future be studied complementarily by heterologous gene expression. Curiously, a recent study reported on the production of similar compounds from the new species Tengochaeta bulbillosa, accommodated in the Chaetomiaceae in culture, which should be an interesting candidate to further study the chemical space of these interesting azaphilones (Barrera-Adame et al. 2025). Until then, fresh specimens need to be collected to isolate and unambiguously assign the identity of individual compounds for chemotaxonomic purposes.
Supplementary Material
Supplemental Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barrera-Adame DA, Marin-Felix A, Wegener AK, Lalk M, Stadler M, Niedermeyer THJ. 2025. Bulbillosins A - E, azaphilones from Tengochaeta bulbillosa sp. nov. (Chaetomiaceae), a root endophyte of the Chinese medicinal plant Aster tataricus. IMA Fungus. 16:e 141036. doi: 10.3897/imafungus.16.141036.40052077 PMC 11882021 · doi ↗ · pubmed ↗
- 2Becker K, Lambert C, Wieschhaus J, Stadler M. 2020. Phylogenetic assignment of the fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and investigation of its secondary metabolites. Microorganisms. 8(9):1397. doi: 10.3390/microorganisms 8091397.32932875 PMC 7565716 · doi ↗ · pubmed ↗
- 3Becker K, Stadler M. 2021. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J Antibiot (Tokyo). 74(1):1–23. doi: 10.1038/s 41429-020-00376-0.33097836 PMC 7732752 · doi ↗ · pubmed ↗
- 4Bills GF, González-Menéndez V, Martín J, Platas G, Fournier J, Peršoh D, Stadler M, Figueras MJ. 2012. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLOS One. 7(10):e 46687. doi: 10.1371/journal.pone.0046687.23056404 PMC 3467290 · doi ↗ · pubmed ↗
- 5Bitzer J, Köpcke B, Stadler M, Hellwig V, Ju YM, Seip S, Henkel T. 2007. Accelerated dereplication of natural products, supported by reference libraries. Chimia. 51(6):332–338. doi: 10.2533/chimia.2007.332. · doi ↗
- 6Bitzer J, Læssøe T, Fournier J, Kummer V, Decock C, Tichy HV, Piepenbring M, Peršoh D, Stadler M. 2008. Affinities of Phylacia and the daldinoid Xylariaceae, inferred from chemotypes of cultures and ribosomal DNA sequences. Mycol Res. 112(2):251–270. doi: 10.1016/j.mycres.2007.07.004.18319146 · doi ↗ · pubmed ↗
- 7Cedeño-Sanchez M, Charria-Girón E, Lambert C, Luangsa-Ard JJ, Decock C, Franke R, Brönstrup M, Stadler M. 2023. Segregation of the genus Parahypoxylon (Hypoxylaceae, Xylariales) from Hypoxylon by a polyphasic taxonomic approach. Mycokeys. 95:131–162. doi: 10.3897/mycokeys.95.98125.37251992 PMC 10210297 · doi ↗ · pubmed ↗
- 8Cedeño-Sanchez M, Lambert C, Mejia LC, Ebada SS, Stadler M. 2025. Chemotaxonomic and molecular phylogenetic studies of selected Hypoxylon species from the Neotropics. Mycology. 16(1):250–265. doi: 10.1080/21501203.2024.2378071.40083408 PMC 11899249 · doi ↗ · pubmed ↗