Insights into the lifecycle of Cryptosporidium and compounds targeting developmental stages
Zoë Reynolds, Sumiti Vinayak

TL;DR
This review explores the lifecycle of the Cryptosporidium parasite and compounds that target its developmental stages to help combat disease.
Contribution
The paper provides new insights into stage-specific gene expression and molecular targets for intervention in Cryptosporidium.
Findings
Recent advances in understanding the Cryptosporidium lifecycle and stage-specific gene expression are highlighted.
The role of parasite proteins in invasion and reproduction is discussed.
Effective compounds targeting lifecycle stages are identified.
Abstract
The intestinal protozoan parasite, Cryptosporidium, is a leading cause of diarrhea-associated illness and death in young children, immunocompromised individuals, and neonatal ruminant animals. This apicomplexan parasite completes its entire lifecycle within a single host, involving a timely and coordinated progression through asexual and sexual developmental stages. With no fully effective drugs or vaccines available, a deeper understanding of the parasite’s lifecycle stages is crucial for identifying new molecular targets for disease intervention. In this review, we discuss recent advances in understanding the Cryptosporidium developmental lifecycle, stage-specific gene expression, and the role of parasite proteins in invasion, asexual proliferation, and sexual stages. We also discuss the lifecycle stages targeted by a few highly effective anticryptosporidial compounds.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
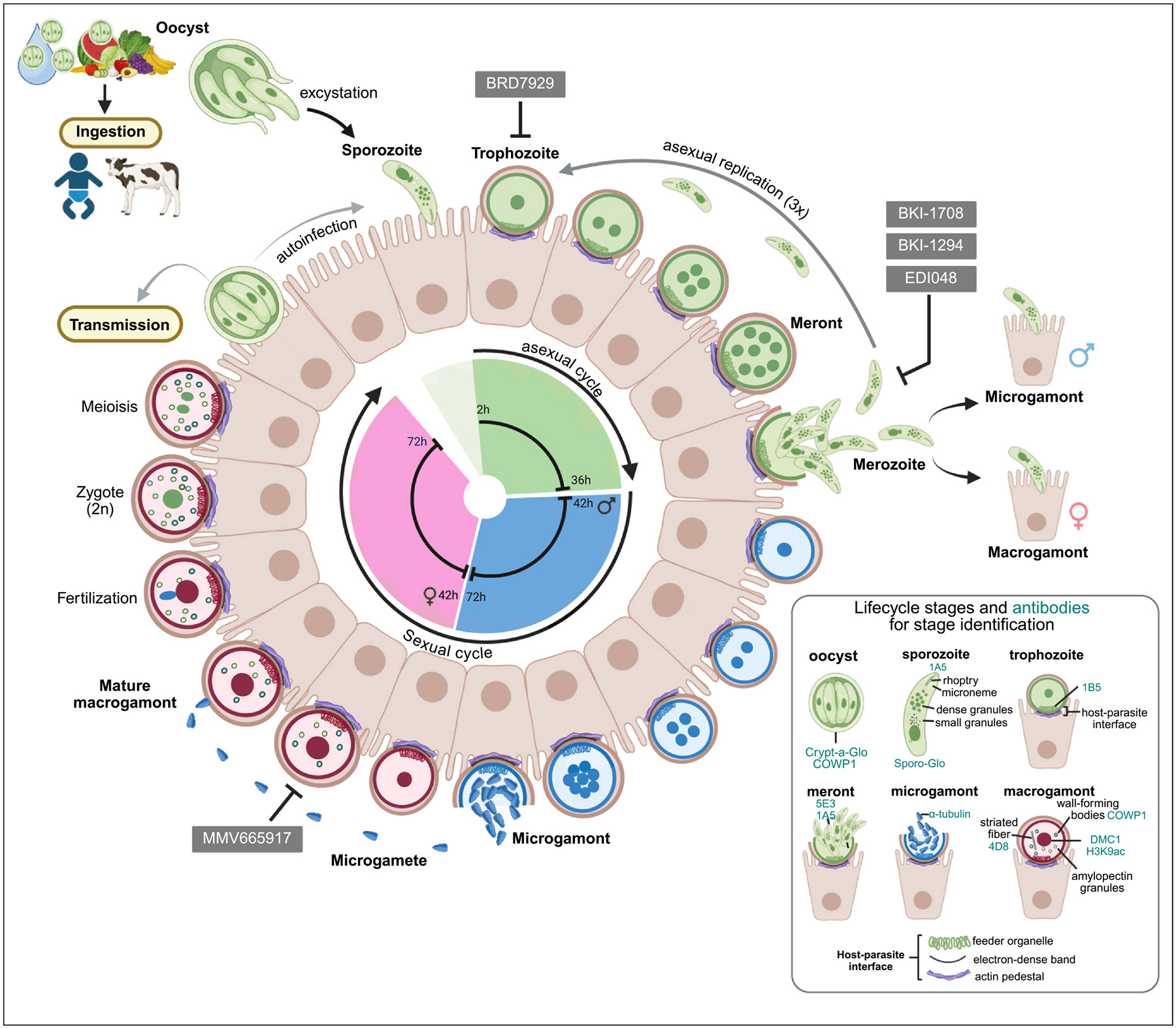 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Toxoplasma gondii Research Studies · Coccidia and coccidiosis research
Introduction
The protozoan parasite Cryptosporidium is a major cause of diarrheal disease and death among children under five years in low- and middle-income countries [1–4]. Repeated episodes of cryptosporidiosis are associated with chronic malnutrition, growth stunting, and developmental defects in young children [3,5–7]. Cryptosporidium species (C. parvum and C. hominis) are responsible for frequent global outbreaks caused by contamination of drinking and recreational water facilities with oocysts, as well as contact with infected animals [8,9]. Cryptosporidium parvum infects young ruminant animals (neonatal calves, young lambs, and goat kids) and humans, while C. hominis infection is limited to humans. Currently, no fully effective drugs exist to treat human or animal cryptosporidiosis or to eliminate oocyst shedding. The only US Food and Drug Administration (FDA)-approved drug, nitazoxanide, has poor efficacy in children and immunocompromised individuals, and there is no vaccine to prevent cryptosporidiosis in humans [6,10–12]. Cryptosporidium completes its entire lifecycle (including both asexual and sexual stages) within a single host and infects only the intestinal epithelial cells (enterocytes) of the small intestine. The development of CRISPR/Cas9 gene editing and mouse models of infection has advanced the field by enabling (i) the discovery of new aspects of the Cryptosporidium developmental cycle, (ii) the study of parasite gene functions, (iii) insights into host-parasite interactions, (iv) testing the effectiveness of new anticryptosporidial drugs, and (v) validation of drug targets [13–18]. The past decade has seen significant progress in the field, including the expansion of the genetic toolbox and the development of new intestinal organoids and air–liquid interface culture systems for growing the parasite in vitro [19–21]. For cell biological studies and phenotypic characterization of Cryptosporidium gene knockouts, researchers rely on a human ileocecal adenocarcinoma cell line (HCT-8) that is easy to grow and maintain in the laboratory. Infected HCT-8 cultures have been used for fixed- and live-imaging studies to understand the Cryptosporidium lifecycle and to generate multiple gene-expression datasets across different stages of parasite development. Although Cryptosporidium undergoes both asexual and sexual developmental stages in HCT-8 cells, the culture stalls at the fertilization stage with no new oocyst formation [22]. However, this block in fertilization is overcome in mice and organoid cultures, leading to oocyst production.
This review focuses on our current understanding of the Cryptosporidium lifecycle, gained through genetics, advanced microscopy, and transcriptomic approaches. We also highlight a few anticryptosporidial compounds that demonstrate efficacy in vitro and/or in animal infection models and inhibit different stages of parasite development, as evidenced by imaging studies. Figure 1 illustrates the various stages of the Cryptosporidium lifecycle and their timing, along with a small panel of antibodies and stains used to distinguish parasite stages.
Cryptosporidium developmental lifecycle: highly timed and coordinated
The lifecycle of Cryptosporidium begins when the thick-walled oocyst is ingested by a human or animal host. The oocyst excysts to release the four haploid sporozoites into the lumen of the small intestine. Changes in pH, bile salts, and the action of proteases have been reported as triggers for the induction of excystation [23]. These sporozoites have gliding motility and penetrate the mucus layer to attach to the intestinal epithelial cells. Upon contact with the cell, the parasite induces host actin polymerization and massive host cell remodeling through proteins it secretes into the host cell [24–27]. This host cell remodeling results in the microvillus membrane encapsulating the sporozoite, but the parasite never fully enters the host cell cytoplasm. Thus, the parasite establishes a unique epicellular (‘intracellular but extracytoplasmic’) niche for itself, where it resides and develops within a parasitophorous vacuole, yet remains separated from the host cell cytoplasm [28]. This unique epicellular localization is maintained throughout the parasite’s developmental cycle. Electron and fluorescence microscopy studies have shown that multiple structures separate the parasite from the host cell and restrict it to this extracytoplasmic location. These include a feeder organelle, a ring-like tight junction structure, an electron-dense band, and finally the actin pedestal at the base of the host–parasite interface [25,26]. After invasion, the sporozoite differentiates into a trophozoite, which then undergoes development, followed by three cycles of asexual replication to produce meronts (merogony). Merozoites released from the meront after the third replication cycle take an obligatory route to sexual development, differentiating into male and female sexual stages [22,29]. The male gametes (microgametes) released from the male gamont (microgamont) fertilize the female gamont (macrogamont). The resulting zygote (diploid) undergoes meiosis and sporogony to form an oocyst with four sporozoites. The oocyst is shed in the host’s feces or can initiate an autoinfection cycle within the same host (Figure 1).
As illustrated in Figure 1, the lifecycle of Cryptosporidium is highly programmed, proceeds in a coordinated manner, and takes 72 hours to complete. Infection of HCT-8 cells with fluorescent C. parvum reporter strains and time-lapse microscopy have revealed the dynamics and progression of the lifecycle [22,29]. A single cycle of asexual replication takes approximately 12 hours, and the parasite undergoes three rounds of replication. Live imaging of meronts has demonstrated that after the third round of asexual replication (36 hours), the merozoites released from a mature 8-nucleated meront are sexually committed and can develop into either male or female gamonts. The sex ratio is estimated to be twice the number of females to male gamonts for merozoites egressing from a single meront. Thus, there is no type-II meront, which was previously thought to be the precursor to the sexual stages [29]. Sexual development continues, with most parasites at 48 hours being male or female, and over 80% of sexual stages are seen at 72 hours post-infection [29].
In the following sections, we provide a concise overview of each stage of the Cryptosporidium life cycle and discuss the functions of selected proteins that have been functionally characterized or experimentally localized at different parasite stages. Table 1 summarizes the roles of the proteins discussed throughout the article. Additionally, we highlight potent compounds that have been shown to target various stages of parasite development.
Sporozoite: attachment and invasion of the host cell
Live-imaging studies have shown that host cell invasion by the sporozoite is rapid. Sporozoite attachment occurs within a few seconds of contact, followed by bending and straightening (1–2 minutes) for its complete encapsulation by the host membrane, and transforms into a trophozoite by 8 minutes [30]. The sporozoite releases secretory proteins for attachment and invasion of the host cell, but only a few have been experimentally characterized. Recently, spatial proteomics of the sporozoite has identified 154 secretory proteins that are stored in four secretory organelles: micronemes, rhoptries, dense granules, and small granules (an organelle unique to this parasite), but only a handful have been functionally characterized to date [31].
Regarding micronemal proteins, the localization and functional roles of a few Cryptosporidium proteins involved in parasite motility, attachment, and invasion have been described. These include the thrombospondin repeat-containing (TSP) family of proteins that are c-mannosylated and contribute to parasite attachment and invasion [32]. The C. parvum TSP1 (TRAP-C1) localizes to the apical end and surface of sporozoites, while TSP8 (MIC1) and TSP4 localize to micronemes, likely playing roles in gliding motility and adhesion [32–35]. Interestingly, C. parvum TSP4 is trafficked along two unique central microtubules and is secreted during sporozoite excystation, gliding, and invasion. Selective kinesin-5 inhibitors have been shown to block the secretion and transport of TSP4 [34]. Mucin-like glycoproteins such as GP60 (which is proteolytically cleaved into GP40/GP15), GP900, CP23, MUC1-MUC7, and apical complex glycoproteins AGP1 and AGP2 have been shown to play roles in parasite motility, attachment, and invasion [36–43]. Some of the immunogenic mucin-like glycoproteins (namely GP40, GP15, CP23, GP900, and MUC8) have been proposed as vaccine candidates because of their roles in parasite attachment and invasion, as well as the antibody responses they elicit, which are linked to protection against reinfection in both humans and animals [44,45]. Recently, the European Medicines Agency approved a recombinant vaccine (Bovilis Cryptium^®^) based on C. parvum GP40 for use in neonatal calves to provide protection against cryptosporidiosis. Inoculating pregnant cows with recombinant GP40 protein produces antibodies against this protein that are passively transferred through colostrum to newborn calves, and this has been shown to reduce the severity and duration of diarrhea, although it does not fully prevent infection [45]. Additionally, a C. parvum rhomboid protease, ROM1, which functions in invasion and proteostasis of the parasitophorous vacuole membrane and feeder organelle, along with four proteins (encoded by cgd3_980, cgd1_3550, cgd1_3680, and cgd2_1590 genes), has also been localized to the micronemes using immunoelectron microscopy [46].
Regarding rhoptry proteins, one rhoptry neck protein (PRP1), a homolog of Toxoplasma gondii RON1, localizes to the organelle in sporozoites, merozoites, and at the parasite–host cell interface in developing trophozoites [47]. The C. parvum rhoptry bulb protein ROP1 is injected into the host cell during invasion and interacts with the host cytoskeletal modulator LMO7. Genetic ablation of ROP1 has been shown to reduce infectivity in immunocompromised mice [30]. Additionally, eight rhoptry bulb proteins (ROPs) have been identified at the apical end of sporozoites that are secreted during invasion, but their functions remain uncharacterized [30]. Regarding dense granules, six proteins (DG1 to DG6) have been localized to this organelle; however, their molecular functions in host–parasite interactions are not known [31,48]. Recently, deletion of DG6 has been shown to reduce parasite virulence in vivo [48]. Concerning small granule proteins, SG1, SG2, SKSR1, and MVP1 have been localized to this organelle [31,49,50]. Genetic deletion of SKSR1 or a base insertion to change the reading frame has been shown to reduce parasite virulence [49]. MVP1 is an exported virulence factor that interacts with host EBP50 and CDC42 to drive elongation of intestinal microvilli during infection [50].
Overall, the mechanism by which Cryptosporidium invades to create its unique epicellular localization is particularly intriguing. The encapsulation of the parasite by host membranes likely serves as an adaptation that shields it from immune recognition and clearance. What other benefits the parasite derives from this unique epicellular positioning, and the molecular processes that prevent it from becoming fully intracellular, remain to be investigated. Further research on the role of Cryptosporidium secretory effector proteins in host cell manipulation would enhance our understanding of the molecular mechanisms of attachment and invasion.
Trophozoite: development and nutrient uptake
The trophozoite undergoes growth and development before DNA replication occurs. The feeder organelle is fully formed in the mature trophozoite and is also observed in all intracellular stages. This organelle is an elaborate network of tubular and bursiform membranes that has been historically described for its role in the uptake of essential nutrients from the host cell that the parasite cannot synthesize on its own [51]. Cryptosporidium has a very streamlined metabolism and salvages many metabolites, including purines, pyrimidines, fatty acids, amino acids, glucose-6-phosphate, and reduced glutathione from the host cell [41,52–54]. Furthermore, the parasite lacks an apicoplast and mitochondrion organelles but has a mitochondrial remnant (mitosome), does not have the TCA cycle, and relies on glycolysis for energy production [55].
Given the heavy dependency of Cryptosporidium on the host cell for metabolites, it is not surprising that the compact 9.1 Mb parasite genome encodes an extensive repertoire of 152 transporters [56]. However, the localization and function of most of these transporters remain unknown. So far, only four C. parvum transporters have been localized to the feeder organelle. These include the C. parvum ATP-binding cassette (ABC1) transporter and two glucose transporters, GT1 and GT2 [54,57]. Both GT1 and GT2 transport glucose-6-phosphate, whereas GT1 also transports glucose from the host cell [54]. Interestingly, a recent study has identified the role of the multidrug-resistant protein (CpMRP1), an ABC transporter, in exporting the toxic microbial metabolite deoxycholate to establish infection and contribute to virulence in mice [58]. This suggests that metabolites produced by the gut microbiota can restrict Cryptosporidium growth, but the parasite has evolved mechanisms to export microbial metabolites to thrive in the intestinal environment.
Regarding the inhibition of the trophozoite stage, BRD7929, a bicyclic azetidine, has been demonstrated to block its development. EdU (5-ethynyl-2’-deoxyuridine) incorporation and imaging indicate that BRD7929 treatment stalls the parasite at the early trophozoite stage, resulting in an expanded feeder organelle and a lack of DNA replication [59]. BRD7929 targets the phenylalanyl tRNA synthetase (PheRS), effectively killing Cryptosporidium in vitro and in immunocompromised mice [18]. In fact, aminoacyl-tRNA synthetases (aaRS) are conserved biological targets across protozoan parasites, and compounds targeting PheRS, methionyl tRNA synthetase (MetRS), as well as lysyl tRNA synthetase (KRS) in Plasmodium or Trypanosoma, have been optimized for Cryptosporidium and shown to kill the parasite both in vitro and in vivo [18,60–62].
Regarding the feeder organelle, many questions remain in the field. How the feeder organelle forms and its boundaries are still unclear. Is it open to the host cytoplasm? Which metabolites are transported to the parasite? Additionally, elucidating the role of transporters involved in compound efflux may have significant implications for understanding the development of resistance to new anticryptosporidial drugs.
Asexual proliferation (merogony)
During a single asexual proliferation cycle, the trophozoite undergoes three nuclear divisions to form an 8-nucleated meront. Transcriptomic data show that DNA replication and ribosomal genes are expressed during the meront growth phase, followed by expression of membrane and apical structure components, and finally the secretion machinery, which forms all four apical organelles for merozoite invasion [63]. Merozoites that egress from the meront are motile and invade new enterocytes [22,29]. Although the molecular mechanisms governing meront development and merozoite egress are not well understood, calcium signaling is reported to play an important role in this process [17]. The calcium-dependent protein kinase 1 (CDPK1) is a leading drug target, and selective bumped kinase inhibitors (BKI-1294, BKI-1708) have shown efficacy in killing Cryptosporidium in both immunocompromised mice and calf infection models [64–66]. CDPK1 is essential for parasite survival and is expressed in trophozoites, 8-nucleated meronts, and merozoites but not in the 4-nucleated meronts, indicating it is cell-cycle regulated [17]. Conditional knockdown of CDPK1 has been demonstrated to cause defects in merozoite development [17]. Notably, the promising clinical candidate, EDI048, a soft-drug that inhibits parasite PI(4)K (phosphatidylinositol-4-OH kinase), has been shown to block membrane development in merozoites, thus stalling them at the meront stage [67]. Additionally, the C. parvum cyclic GMP-dependent protein kinase G (PKG) and aspartyl protease 2 (ASP2) have been reported to play roles in merozoite egress [68,69]. Thus, targeting merozoite development and egress provides attractive targets for drug development since they can prevent both parasite proliferation and entry into the sexual cycle.
Together, these transcriptomics, cell biology, and drug discovery studies have enhanced our understanding of merozoite development and egress. However, many questions still need to be investigated. For example, the mechanisms by which Cryptosporidium counts three rounds of replication during merogony remain unclear. The entry of merozoites into the sexual cycle after the third replication is particularly intriguing. Environmental cues sensed by the parasite and transcriptional regulation might play a role in this highly programmed process, but further research is necessary to uncover these mechanisms.
Microgamont development and egress of male gametes
Microgamonts develop and undergo four rounds of nuclear division to produce 16 microgametes. Early microgamonts (~42 hours) have their eight nuclei arranged in a compact, rosette-shaped pattern, and as they develop, the nuclei become more rounded, ultimately resulting in bullet-shaped nuclei in the 16 non-flagellated male gametes [22,70]. The C. parvum single-cell transcriptomics atlas has mapped 389 genes to three clusters (early, mid, and late males), with 51 genes that are exclusively expressed only at this stage [63]. These 51 male-exclusive genes encode transcription factors, signaling molecules, cell cycle regulators, proteases, secretory proteins, and membrane proteins. To date, only two transcription factors have been functionally validated in male development. The Myb-M transcription factor is the earliest determinant of male fate, and its conditional genetic deletion results in the complete loss of male gametes in vitro and the absence of oocyst shedding in vivo [63]. The Apetala-2 transcription factor, AP2-M, is expressed exclusively in microgamonts during development and has been reported to be essential for parasite survival [71].
Furthermore, the C. parvum calcium-dependent protein kinase-5 (CDPK5) has been shown to play a role in the egress of male gametes and in parasite virulence, with substrates of this kinase identified using phosphoproteomics [70]. Additionally, ASP2, which is important for merozoite egress, has also been shown to be necessary for male gamete egress [68].
Two other male-specific genes that have been localized to male gametes encode for hapless 2 protein (HAP2) and the secretory family protein GGC1 [22,63]. HAP2 localizes to the apical pole of microgametes, while GGC1 localizes to a bar-like structure at the apex. This suggests that HAP2 and GGC1 may be components of the adhesion apparatus or fertilization machinery required for the fusion of male gametes with the female gamont.
Many questions about male gametogenesis and fertilization remain unanswered. Male gametes released from microgamonts are immotile, yet they somehow travel toward the macrogamont; how this occurs remains a mystery. Moreover, the molecular mechanisms underlying the adhesion of the male gamete to the female gamont for fertilization are yet to be discovered.
Macrogamont development
The single-nucleated macrogamont develops within the parasitophorous vacuole and does not undergo nuclear division or egress, but increases its size during development. Electron microscopy studies have reported that the macrogamont synthesizes the components of the oocyst wall, the wall-forming bodies 1 and 2, as well as amylopectin granules [72,73]. These amylopectin stores are degraded by the parasite glycogen phosphorylase, which releases phosphorylated glucose required for glycolysis and ATP generation [54]. Bulk and single-cell RNA sequencing of fluorescent reporter parasites in vitro and in vivo have identified genes that are highly expressed in the females, and these encode for oocyst wall synthesis, energy storage (amylopectin and trehalose metabolism), meiosis and DNA repair, glycosyltransferases, proteases, polyketide, and fatty acid synthesis [22,63]. Out of the 773 genes enriched in the female transcriptome, 187 genes are expressed exclusively at this stage.
Few proteins have been genetically characterized for their roles in macrogamont development. Conditional deletion of the essential female-specific AP2 transcription factor (AP2-F) has been shown to downregulate expression of genes encoding the DNA meiotic recombinase (DMC1), NIMA kinase 5, DNA polymerase, and crystalloid body proteins (an organelle of unknown function found in sporozoite), and significantly reduce oocyst shedding in IFN-γ KO mice [71]. Another protein reported to play a role in macrogamont development is the insulinase-like protease 1 (INS1), which belongs to the M16 family of metallopeptidases. INS1 localizes to small secretory vesicles near wall-forming bodies in the macrogamont, and genetic deletion of INS1, or its replacement with an active-site mutant, results in reduced macrogamont numbers in vitro and decreased oocyst shedding in vivo [74].
Two female-specific antibodies, DMC1 and 4D8, have proven useful for identifying compounds that specifically target macrogamont development. A DMC1 antibody-based high-content imaging assay identified nine potent small molecule compounds from a drug repurposing library that inhibited macrogamont differentiation [75]. Moreover, the piperazine-based inhibitor, MMV665917, a highly efficacious anticryptosporidial compound, was found to inhibit macrogamont development in DMC1 and 4D8-based imaging assays [59,76,77]. Interestingly, the fiber structure in the cytoplasm of macrogamonts stained with the 4D8 antibody was much shorter after treatment with this compound, although the composition of this fiber is not known [59].
Together, these studies show that the macrogamont is transcriptionally and translationally active and prepares for oocyst formation. Further research is required to understand the molecular mechanisms that regulate macrogamont maturation, as well as the formation of amylopectin granules and oocyst wall components.
Oocyst: forming the protective wall structure
The oocyst has a multilayered structure that protects the sporozoites inside and includes a glycocalyx, inner layers of acid-fast lipids, and an innermost layer of nine cysteine- and histidine-rich oocyst wall proteins (COWPs) [73,78–80]. COWP1 and COWP8 are expressed in the oocyst wall, as shown through immunolocalization and immunofluorescence studies, but COWP8 does not contribute to the strength of the oocyst wall nor is it required for transmission [73,80,81]. Interestingly, COWP2, COWP3, and COWP4 localize to the suture, which is the region where the sporozoite exits the oocyst [81]. Historically, thin-walled oocysts that easily excyst within the same host have been linked to the autoinfection cycle, while thick-walled oocysts have been associated with transmission [72].
Understanding how the oocyst wall is constructed would elucidate the oocyst’s resistance to disinfection and inform the development of compounds that target key wall components to reduce environmental contamination.
Concluding remarks
Overall, new insights into the Cryptosporidium lifecycle are enhancing our fundamental understanding of parasite biology and helping identify vulnerabilities that can be targeted for drug and vaccine development. Insights into the host–parasite interactions, characterization of transporter function, and identification of critical metabolites that the parasite scavenges from the host will open new avenues for both parasite- and host-targeted therapies. Future research on the molecular mechanisms underlying the development of sexual stages and the fertilization process will improve our understanding of the parasite’s lifecycle. Thus, targeting either the initial stages of infection, merogony, or the sexual development cycle to prevent transmission are promising strategies to combat cryptosporidiosis and reduce its significant public health burden.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, : Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 2013, 382:209–222.23680352 10.1016/S 0140-6736(13)60844-2 · doi ↗ · pubmed ↗
- 2Kotloff KL, Nasrin D, Blackwelder WC, Wu Y, Farag T, Panchalingham S, Sow SO, Sur D, Zaidi AKM, Faruque ASG, : The incidence, aetiology, and adverse clinical consequences of less severe diarrhoeal episodes among infants and children residing in low-income and middle-income countries: a 12-month case-control study as a follow-on to the Global Enteric Multicenter Study (GEMS). Lancet 2019, 7:e 568–e 584.10.1016/S 2214-109X(19)30076-2PMC 648477731000128 · doi ↗ · pubmed ↗
- 3Gilbert IH, Vinayak S, Striepen B, Manjunatha UH, Khalil IA, Van Voorhis WC, Cryptosporidiosis Therapeutics Advocacy Group CTAG, Cryptosporidiosis Therapeutics Advocacy CTAG: Safe and effective treatments are needed for cryptosporidiosis, a truly neglected tropical disease. BMJ Glob Health 2023, 8:e 012540.10.1136/bmjgh-2023-012540 PMC 1040737237541693 · doi ↗ · pubmed ↗
- 4Platts-Mills JA, Babji S, Bodhidatta L, Gratz J, Haque R, Havt A, Mc Cormick BJ, Mc Grath M, Olortegui MP, Samie A, : Pathogen-specific burdens of community diarrhoea in developing countries: a multisite birth cohort study (MAL-ED). Lancet Glob Health 2015, 3:e 564–e 575.26202075 10.1016/S 2214-109X(15)00151-5PMC 7328884 · doi ↗ · pubmed ↗
- 5Khalil IA, Troeger C, Rao PC, Blacker BF, Brown A, Brewer TG, Colombara DV, Hostos ELD, Engmann C, Guerrant RL, : Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: a meta-analyses study. Lancet Glob Health 2018, 6:e 758–e 768.29903377 10.1016/S 2214-109X(18)30283-3PMC 6005120 · doi ↗ · pubmed ↗
- 6Checkley W, White AC, Jaganath D, Arrowood MJ, Chalmers RM, Chen X-M, Upton SJ, Griffiths JK, Guerrant RL, Hedstrom L, : A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect Dis 2015, 15:85–94.25278220 10.1016/S 1473-3099(14)70772-8PMC 4401121 · doi ↗ · pubmed ↗
- 7Investigators: The MAL-ED study: a multinational and multidisciplinary approach to understand the relationship between enteric pathogens, malnutrition, gut physiology, physical growth, cognitive development, and immune responses in infants and children up to 2 years of age in resource-poor environments. Clin Infect Dis 2014, 59:S 193–S 206.25305287 10.1093/cid/ciu 653 · doi ↗ · pubmed ↗
- 8Gharpure R, Perez A, Miller AD, Wikswo ME, Silver R, Hlavsa MC: Cryptosporidiosis outbreaks — United States, 2009–2017. MMWR Morb Mortal Wkly Rep 2019, 68:568–572.31246941 10.15585/mmwr.mm 6825 a 3PMC 6597118 · doi ↗ · pubmed ↗