Characterization of a novel composite ICE in Streptococcus agalactiae conferring resistance to macrolides [erm(TR)] and cadmium (cadA)
Sida Yi, Chunli Shi, Liufan Yin, Xing Xu, Xueliang Wang

TL;DR
A new composite genetic element in Streptococcus agalactiae was found to carry genes for resistance to antibiotics and cadmium, aiding in the spread of these resistance traits.
Contribution
The discovery of a novel composite ICE, ICESag39, with a modular structure that promotes resistance gene dissemination.
Findings
ICESag39 has a nested structure with an internal Tn1806-like ICE carrying resistance genes erm(TR) and cadA.
ICESag39 transfers at a frequency of 8.2 × 10−9 and co-transfers both resistance determinants.
Database screening revealed 199 similar ICEs, 62.8% of which co-carried erm(TR) and cadA.
Abstract
Macrolide resistance genes (erm and mef families) and heavy metal resistance genes (cadA) are increasingly disseminated among streptococci via diverse mobile genetic elements. Through whole-genome sequencing of 16 Streptococcus agalactiae isolates resistant to both erythromycin and clindamycin, we identified 19 integrative and conjugative elements (ICEs), a type of self-transfer genetic elements, conferring antibiotic resistance. Among these, a novel composite ICE designated ICESag39 was identified in S. agalactiae Sag39 through comparative analysis with the NCBI database. ICESag39 measured 113,125 bp in length, and it featured a nested “Russian doll” structure comprising an ICESa2603 family backbone integrated with an internal Tn1806-like ICE. The embedded Tn1806-like ICE contained four variable regions (VR1-VR4) that serve as insertion hotspots; among these, VR3 and VR4 carry…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1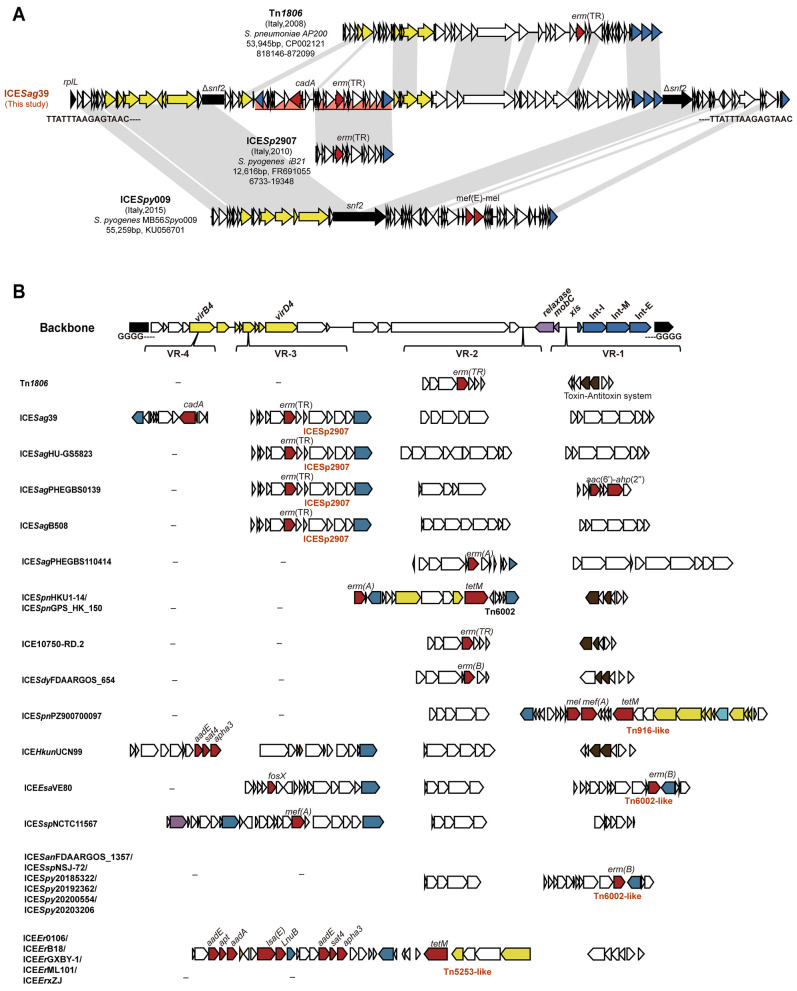 Figure 2
Figure 2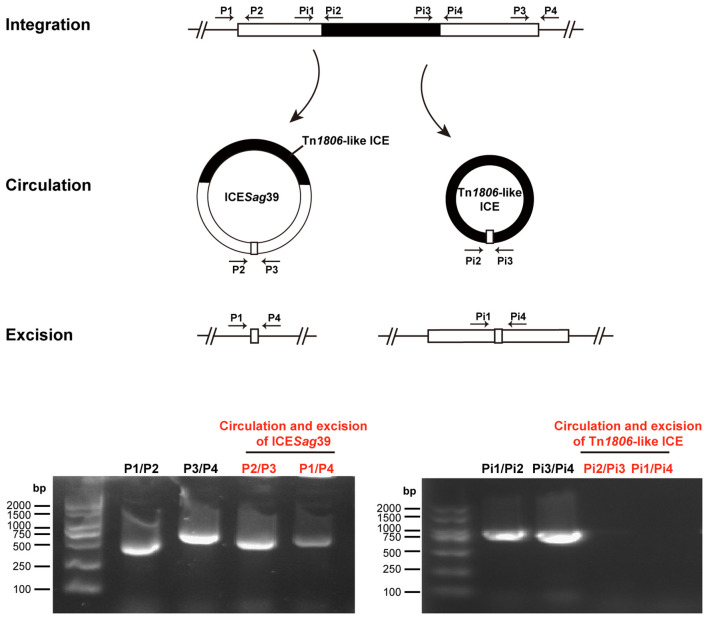 Figure 3
Figure 3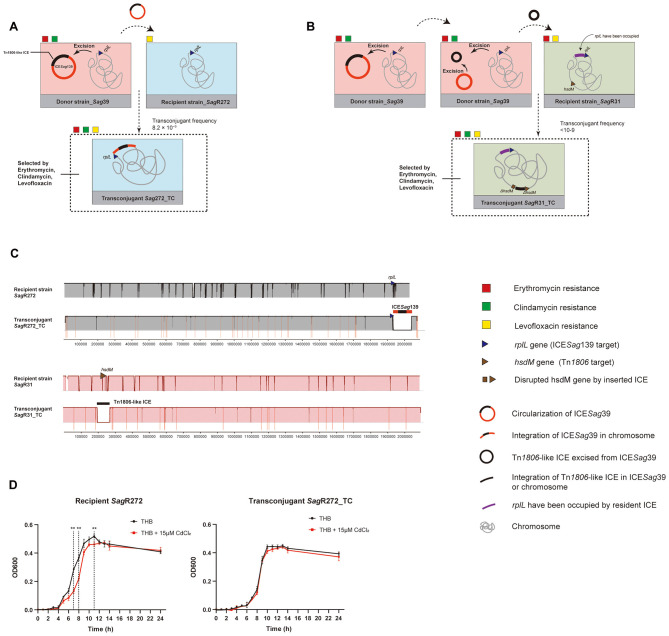 Figure 4
Figure 4|
|
|
|
|
|
|
| ||||
|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
| ||||||
| 2021 | >256 | >256 | 128 | ≤0.5 | ≤0.5 | Tn | ST1 | - | ||
| 2021 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2022 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2022 | 64 | >256 | 64 | 32 | 1 |
| ICE | ST19 | ||
| 2022 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2022 | 4 | 2 | 32 | ≤0.5 | ≤0.5 |
| Tn | ST23 | ||
| 2022 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2022 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2022 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | |||
| 2023 | >256 | >256 | ≤0.5 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
|
|
|
|
|
|
|
|
|
|
| |
| 2023 | >256 | >256 | 1 | ≤0.5 | ≤0.5 | Tn | ST10 | - | ||
| 2023 | >256 | 256 | >256 | ≤0.5 | ≤0.5 |
| ICE | ST17 | ||
| 2024 | >256 | >256 | ≤0.5 | 2 | ≤0.5 | Tn | ST10 | |||
| 2024 | 1 | 1 | >256 | ≤0.5 | ≤0.5 |
| Tn | ST10 | ||
| 2025 | 1 | 2 | 32 | 1 | 1 |
| Tn | ST10 | ||
|
|
|
|
|
|
| |||||
|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
| |||||||
|
| Donor |
|
| 32 | ≤0.5 | Sequence02, 12144-62118 | ||||
| Recipient | ≤0.5 | 1 | 128 |
| 8.2 × 10−9 |
| ICE | |||
| Transconjugant |
|
| 128 |
| ||||||
| Recipient | 4 | ≤0.5 | 4 |
| <10−9 |
| Tn | |||
| Transconjugant | >256 |
| 4 |
| ||||||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeonatal and Maternal Infections · Antimicrobial Resistance in Staphylococcus · Pneumonia and Respiratory Infections
Introduction
1
Macrolide resistance in streptococci is primarily mediated through two distinct mechanisms: active drug efflux and ribosomal target site modification (Leclercq, 2002; Varaldo et al., 2009). The first mechanism involves efflux pumps encoded by mef genes [such as mef (E), mef (A), and mef (I)], which reduce intracellular concentrations by expelling the antibiotic from the cell. This efflux-based strategy confers low-level resistance to 14- and 15-membered macrolides, and it is frequently observed in species such as Streptococcus pneumoniae and Streptococcus pyogenes. The second mechanism entails target site modification mediated by methylases encoded by erm family genes [including erm(A), erm(B), and erm(TR)]. These enzymes methylate the 23S rRNA of the bacterial ribosome, thereby inhibiting antibiotic binding. This modification confers high-level resistance to macrolides, lincosamides, and streptogramin B (the MLS_B_ phenotype), representing the most prevalent and clinically significant macrolide resistance determinants in streptococci and enterococci (Huang et al., 2016a).
The widespread dissemination of erm family genes in S. agalactiae and other streptococci is largely driven by integrative and conjugative elements (ICEs) (Morici et al., 2017; Chen et al., 2021; Yi et al., 2021). These mobile genetic elements excise from the chromosome, form a circular intermediate, and transfer between bacterial cells via conjugation machinery such as type IV secretion systems (T4SSs) and type IV coupling proteins (T4CPs) (Wozniak and Waldor, 2010). Following transfer, a site-specific integrase catalyses ICE integration into specific genomic sites, facilitating the efficient spread of antibiotic resistance genes (ARGs) (Partridge et al., 2018). Beyond antibiotic resistance, some ICEs can also transmit heavy metal resistance determinants, conferring related tolerance (Rodriguez-Blanco et al., 2012; Wang et al., 2025a).
Two groups of ICEs are currently recognized as common vehicles for the dissemination of erm genes in streptococci, namely the ICESa2603 and Tn1806 families. The ICESa2603 family, first identified in S. agalactiae 2603V/R, is the most frequently reported group (Gawron-Burke and Clewell, 1982; Huang et al., 2016a; Yi et al., 2021). These elements often carry multiple antibiotic resistance genes and integrate site-specifically 3′ rplL gene, which encodes the 50S ribosomal subunit protein L7/L12. Representative members include ICESsu32457 from Streptococcus suis, carrying aadE-aphA (conferring aminoglycoside resistance), erm(B) and tet(40)-tet(O/W/32/O) (tetracycline resistance) (Palmieri et al., 2012), as well as ICESpn529IQ from S. pneumoniae, which harbors catQ (chloramphenicol resistance), tetM (tetracycline resistance), mef (I), and erm(TR) (Mingoia et al., 2014). In addition, some ICESa2603-like family ICEs (characterized by distinct integrase modules and integration into alternative sites) also carry erm genes, such as ICESsuYZDH1 from S. suis (Yi et al., 2021) and ICESag37 from S. agalactiae (Zhou et al., 2017). The second group, Tn1806, was initially identified in S. pneumoniae, and it is characterized by its low-integration-site specificity and high transfer efficiency, promoting its dissemination across streptococci and enterococci (Camilli et al., 2008; Ambroset et al., 2016). However, ICEs are dynamic and modular. Genetic exchange and interaction can occur between different ICE groups, potentially giving rise to chimeric elements with novel characteristics and resistance profiles (Wang et al., 2025b). For instance, one ICE or mobile genetic element can integrate into a specific site within another, forming larger, more complex composite structures reminiscent of a “Russian doll” (Mingoia et al., 2016; Gawor et al., 2025). Although such composite elements/ICEs have rarely been documented, they can accumulate advantageous genes from multiple ICEs or other mobile genetic elements, thereby enhancing bacterial survival under diverse antibiotic pressures.
In this study, we analyzed the genetic context of antibiotic resistance genes in 16 macrolide-resistant isolates of S. agalactiae and identified a novel composite ICE. This ICE incorporates modules homologous to several previously characterized genetic elements, including an ICESa2603 family backbone, a Tn1806-like ICE, the erm(TR)-carrying element ICESp2907, and an uncharacterized cadA-carrying segment. These genetic elements are assembled at appropriate genomic locations according to their functional properties, resulting in a chimeric ICE that combines structural and functional features from distinct ancestral elements.
Materials and methods
2
Bacterial strains and susceptibility tests
2.1
This study included 47 non-duplicate clinical isolates of S. agalactiae collected and stored by the Shanghai Center for Clinical Laboratory (Shanghai, China) between 2021 and 2025. Among these, 16 isolates exhibited resistance to both erythromycin and clindamycin [minimum inhibitor concentration (MIC) > 1 mg/L]. Recipient strains for conjugation experiments were selected from the 47 S. agalactiae isolates. All bacterial strains were cultured in Todd–Hewitt (TH) broth at 37 °C for 8 h. Antimicrobial susceptibility testing was performed using the broth microdilution method, with breakpoints interpreted according to CLSI (the Clinical and Laboratory Standards Institute) guidelines. The reference strain S. agalactiae ATCC 13813 was obtained from the Huashan Hospital strain repository (Shanghai, China). The clinical isolates were partially collected through routine hospital laboratory procedures, and no additional personal information was involved in this study.
Genome comparative analysis and ICE identification
2.2
Genomic DNA was extracted from S. agalactiae using the QIAGEN Midi Kit (Qiagen, Hilden, Germany) and sequenced by next-generation sequencing (NGS) at BGI Genomics (Shenzhen, China) using HiSeq X (Illumina, San Diego, CA, USA). High-quality reads were assembled with SPAdes (Bankevich et al., 2012), and the assembled sequences were annotated through the RAST online server (rast.nmpdr.org) (Aziz et al., 2008). Contigs were ordered against the complete reference genome S. agalactiae 2603V/R using the “Order Contigs” function in Mauve v2.4.0 (Gawron-Burke and Clewell, 1982; Darling et al., 2004). The reordered genomes were then aligned and visualized against the reference using the “Comparison” function in Mauve, enabling preliminary localization of large genetic elements. Putative mobile genetic elements were identified by screening for genes encoding T4SSs, T4CPs, relaxase, and integrase. These candidate regions were further analyzed using ICEfinder to confirm their classification as ICEs.
Amplification experiments
2.3
PCR assays were performed for three objectives (Supplementary Table 1): to screen candidate recipient strains by detecting the presence of an unoccupied rplL gene; to amplify conserved ICE genes in trans-conjugants for preliminary confirmation of ICE integration; and to examine ICE integration, excision, and circularization using the primer pairs P1/P2 and P3/P4 to detect integration, the primer pair P1/P4 to detect excision, and the inverse primer pair P2/P3 to detect circular intermediates.
Conjugative transfer assays
2.4
Recipient strains were selected from clinical isolates based on two criteria: a resistance profile distinct from that of the donor strain and the presence of at least one unoccupied ICE integration site. These characteristics were initially assessed using antimicrobial susceptibility testing and PCR and subsequently confirmed by whole-genome sequencing.
Conjugation was performed using a filter mating method as previously described (Palmieri et al., 2012). Fifty mL Donor and 250 mL recipient strains were grown to the late-log phase, normalized to the same CFU, and mixed at a donor-to-recipient ratio of 1:5. Prior to mixing, 200 μL of the donor culture was sampled for CFU enumeration to calculate the conjugation frequency. A nitrocellulose membrane was cut to an appropriate size, treated with 10 μg/mL DNase to prevent transformation-mediated DNA transfer, and placed on a TH agar plate. The mixed culture was pelleted by centrifugation, resuspended, and evenly spread onto the DNase-treated membrane, followed by incubation at 37 °C for 4 h. After incubation, cells were harvested from the membrane, serially diluted, and plated onto TH agar containing 50 μg/mL erythromycin, 50 μg/mL clindamycin, and 20 μg/mL levofloxacin. Transconjugant colonies were counted after 2–3 days of incubation and randomly selected for validation.
The conjugation frequency was calculated as the number of transconjugants divided by the number of donor cells. Donor count: 100 μL culture was taken from a 50 mL donor culture and serially diluted 10-fold in 1 × PBS for 6-10 times. After thorough mixing, 100 μL dilution was spread evenly onto counting plates. Colony counts between 30 and 300 were considered valid. Transconjugant count: After mating, cells were collected from the nitrocellulose membrane using a sterile swab and resuspended. The suspension was then diluted 10-fold, 100-fold, or used undiluted, and plated onto transconjugant selective medium. Plates with 30-300 colonies were considered valid. If the undiluted suspension yielded transconjugants but fewer than 30 colonies, the transfer was considered successful, but the frequency was too low to be calculated accurately.
Growth curve analysis under cadmium stress
2.5
To evaluate the growth dynamics of the recipient and transconjugants under cadmium stress, growth curves were generated in TH broth with or without cadmium as previous described with minor modification (Parsons et al., 2017). The recipient and transconjugant strains were inoculated into two culture conditions: unsupplemented TH broth and TH broth supplemented with 15 μM CdCl2 to induce metal stress. Cultures were incubated at 37 °C with shaking at 160 rpm, and optical density at 600 nm was measured hourly using a microplate reader until the stationary phase was reached. Growth curves were plotted using GraphPad Prism (GraphPad, Boston, MA, USA), and the growth of transconjugants was compared with that of the recipient strain under cadmium stress. All experiments were performed in at least two independent biological replicates to ensure reproducibility.
Bioinformatics identification of ICEs similar to composite ICESag39
2.6
The core genes of ICESag39, including the tandem serine integrase, relaxase, and conjugation modules (including T4SS and T4CP cluster) of the Tn1806 family, along with the tyrosine integrase, relaxase, and conjugation modules of the ICESa2603 family, were used as templates for BlastN analysis in Nucleotide Collection and Whole-genome shotgun contigs databases from NCBI. According to the criteria defined by ICEFinder (db-mml.sjtu.edu.cn/ICEberg) and previous reports (Ambroset et al., 2016; Huang et al., 2019), a genomic region was considered to harbor an ICE similar to ICESag39 if it contained genes homologous to these core genes (>60% nucleotide identity); the maximum distance between these genes was < 100 kb; Minimum sequence coverage of BLAST hits >25%. In contigs database, a sequenced strain was considered to contain the composite ICE only when the complete element was identified on one scaffold/contig. Boundaries and attachment sites of ICEs were manually checked by terminal integrase and rplL genes. ICE-carried AMR genes were identified as previously described.
GeneBank accession number
2.7
Sequence data generated in this study have been deposited in the NCBI GenBank, including the NGS drafts of 16 erythromycin- and clindamycin-resistant S. agalactiae isolates, the complete genomes of recipient S. agalactiae SagR272 and SagR31, and the NGS draft of transconjugants SagR272_TC and SagR31_TC. Public data used in this study included Tn1806, ICESp2907, ICESpy009, ICESpy009, and the complete genome of S. agalactiae 2603V/R.
Results
3
Characterization of erythromycin- and clindamycin-resistant isolates
3.1
The NGS data of 16 erythromycin- and clindamycin-resistant S. agalactiae isolates revealed diverse phenotypic and genotypic profiles (Table 1). Ten carried the erm(B) gene, and one contained the erm(TR) gene. Three isolates harbored the macrolide resistance genes mef (E) and/or mel, whereas tetracycline resistance, associated with the tet(M) gene, was observed in seven isolates.
Multi-locus sequence typing identified ST10 as the most predominant sequence type (11 isolates), followed by ST19 (two isolates). ST1, ST17, and ST23 (one isolate each) were also detected.
Identification of antibiotic resistance gene-carrying genetic elements
3.2
To localize antibiotic resistance genes and identify their associated genetic elements, we compared the genomes of the 16 S. agalactiae isolates with that of the reference strain 2603V/R (Huang et al., 2016b). This analysis identified 35 major genomic regions of difference, 19 of which contained antibiotic resistance genes along with integrase genes, suggesting integration of mobile genetic elements into the genome (Figure 1). Comparative analysis using GenBank classified the antibiotic resistance gene-carrying elements into several known families: the Tn1806 family carrying erm(B), the Tn916 family carrying tet(M), and the ICESa2603 family carrying mef (E)-mel (Table 1). A novel composite genetic structure carrying erm(TR) was identified in S. agalactiae Sag39, and it was formed by the integration of Tn1806- and ICESa2603-like ICE 3′ rplL gene. This element also carried the cadmium resistance gene cadA, and because of the presence of multiple integrase and conjugation-related genes, it was classified as a composite ICE and designated ICESag39.
Linear comparison of 16 whole-genome shotgun assemblies of Streptococcus agalactiae against the reference strain 2603V/R (AE009948). Conserved syntenic regions are presented as solid colored lines. Discontinuities likely correspond to insertions of exogenous mobile genetic elements (MGEs). Resistance gene-carrying elements are indicated by red rectangles. Other MGEs (lacking known resistance genes or uncharacterized) are presented in white. ICESag39 is highlighted in purple, and its host genome is marked in red. The reference genome includes four well-characterized MGEs denoted by black rectangles (left to right): lambdaSa1 (558,765–559,346 bp), Tn916 (923,439–941,360 bp), ICESa2603 (1,256,480–1,311,228 bp), and lambdaSa2 (1,833,201–1,867,137 bp). Red vertical lines separate individual sequencing scaffolds.
Characterization of ICESag39 and the internal Tn1806-like ICE
3.3
ICESag39 measured 112,852 bp in length and featured 102 open-reading frames and 35% GC content. The entire ICE was flanked by direct repeats (TTATTTAAGAGTAAC–TTATTTAAGAGTAAC) 3′ rplL gene. Comparative genomic analysis revealed that ICESag39 comprises four modules (Figure 2A). The first module shared high similarity with ICESpy009, a member of the ICESa2603 family, but lacked the mef (E)-mel gene cluster present in S. pyogenes (Del Grosso et al., 2016). A Tn1806-like ICE inserted within the snf2 (methyltransferase encoding the sucrose non-fermenting 2 protein) gene of the ICESpy009 backbone constituted the second module. Notably, this internal Tn1806-like ICE itself carried two additional mobile genetic units: ICESp2907, which lacks a conjugation system but carries erm(TR) (Giovanetti et al., 2012), and an uncharacterized element harboring cadA.
Genetic context of ICESag39. (A) Comparative analysis of ICESag39 with known integrative and conjugative elements (ICEs) from the NCBI database, including Tn1806 (CP002121, 818,146–872,099 bp), ICESp2907 (FR691055, 6733–19,348 bp), and ICESpy009 (KU056701). The direct repeat sequences flanking the ICE are TTATTTAAGAGTAAC–TTATTTAAGAGTAAC. Additional regions are highlighted in different background colors. (B) Structure of internal Tn1806-family ICEs displaying conserved modules and variable regions (VR1–VR4) based on an alignment of 25 antibiotic resistance gene-carrying ICEs. Open-reading frames are colored by functional category: integrase/transposase/recombinase (dark blue), relaxase and MobC (purple), type IV secretion system-related genes (yellow), resistance genes (red), host gene interrupted by ICE insertion (black), and toxin-antitoxin systems (brown). Truncated genes are indicated by triangles. Light gray shading denotes regions sharing >60% sequence similarity.
To explore the structural organization and integration hotspots for exogenous DNA, we analyzed the relatively variable Tn1806-like ICE. This analysis involved 41 additional ICEs with high sequence similarity to Tn1806 from NCBI databases, and 25 of these ICEs carried at least one antibiotic resistance genes (Supplementary Table 2). These elements integrate with low specificity into a 4-bp direct repeat sequence (GGGG) within snf2 or diverse methyltransferase encoding genes, truncating the target gene upon insertion. Beyond the conserved integrase and conjugation modules, four variable regions (VR1–VR4) were identified as hotspots for the integration of foreign genetic elements (Figure 2B, Supplementary Table 3). VR1 and VR2 frequently harbored erm(A) and erm(B), along with the aminoglycoside resistance gene aac(6′)-aph(2″) in some cases. Entire mobile elements such as Tn5253 (Mingoia et al., 2014), Tn916 (Rice, 1998), and Tn6002 (Brenciani et al., 2007) were also identified within these regions. VR3 served as a specific integration site for ICESp2907, which carries erm(TR). VR4, uniquely identified in ICESag39, contained the cadA-carrying genetic element.
Excision and circulation of ICESag39 and the internal Tn1806-like ICE
3.4
Chromosomal excision and subsequent circularization are essential for the rolling circle replication and conjugative transfer of ICEs. To preliminarily assess the transfer potential of ICESag39 and the embedded Tn1806-like ICE, we designed PCR primers to detect these processes. Amplification with the primer pairs P1/P2 and P3/P4 as well as Pi1/Pi2 and Pi3/Pi4 (targeting ICE-flanking regions) yielded positive products for both ICESag39 and the internal Tn1806-like ICE (Figure 3), confirming their chromosomal integration. However, only ICESag39 produced faint PCR products with the primer pairs P1/P4 (excision) and P2/P3 (circular intermediate), whereas corresponding amplicons Pi1/Pi4 and Pi2/Pi3 were undetectable for the Tn1806-like ICE (Figure 3). These results suggest that ICESag39 is capable of excising and forming circular intermediates, whereas the internal Tn1806-like ICE appears incapable of autonomous excision, or its transfer frequency is too low to be detected by PCR.
Diagram and PCR detection of integration and excision/circularization of ICESag39 and an internal Tn1806-like integrative and conjugative element (ICE). Black rectangles represent the Tn1806-like ICE within ICESag39, and the combined white and black rectangles represent the composite ICESag39. The primers used are indicated by black arrows. The bottom panel presents the PCR results, and the PCR marker is Vazyme DL2000 DNA. The primer pairs P2/P3 and P1/P4 represent circulation and excision, the primer pairs P1/P2 and P3/P4 represent integration of ICESag39, the primer pairs Pi2/Pi3 and Pi1/Pi4 represent circulation and excision, and the primer pairs Pi1/Pi2 and Pi3/Pi4 represent integration of the internal Tn1806-like ICE.
Transferability of ICESag39
3.5
S. agalactiae Sag39 carrying ICESag39 was used as a donor (resistant to erythromycin, clindamycin, but sensitive to levofloxacin). The recipient strain, S. agalactiae SagR272 (sensitive to erythromycin, clindamycin, but resistant to levofloxacin), which possesses an unoccupied rplL gene for ICESag39, was selected (Figure 4A). In mating experiments, the complete ICESag39 was transferred from the donor to the recipient at a frequency of 8.2 × 10^−9^ (Table 2). Three randomly selected transconjugants acquired high-level resistance to erythromycin and clindamycin while retaining the recipient's resistance to levofloxacin. Under cadmium stress (15 μM CdCl2 in TH broth), transconjugants exhibited significantly better growth than the recipient (Figure 4D), suggesting that transfer of cadmium resistance. Whole-genome sequencing confirmed integration of ICESag39 downstream of the rplL gene in the transconjugants SagR272_TC comparing with recipient SagR272, as expected (Figure 4C).
*Conjugation transfer experiments of ICESag39. (A, B), Schematics of the experimental workflow for conjugative transfer of mobile genetic elements from donor S. agalactiae Sag39 to recipient strains SagR272 and SagR31, respectively. (C) Linear comparison of the recipient and transconjugant genomes confirming the specific integration of the target integrative conjugative element (ICE) into the recipient's genome. (D) Impact of ICESag139 (carrying cadA) on bacterial growth under cadmium stress. Growth curves of the recipient strain and transconjugant in Todd–Hewitt broth with or without 15 μM CdCl2 monitored over 24 h. The recipient strain displayed significant growth impairment in the presence of CdCl2, with notable differences observed at 7, 8, and 11 h (Repeated Measures ANOVA, *p < 0.001). The growth of the transconjugant remained unaffected by cadmium.
To assess whether the internal Tn1806-like ICE could excise and transfer independently, another isolate, S. agalactiae SagR31 (sensitive to clindamycin, but resistant to erythromycin and levofloxacin), was used as the recipient. This strain contains an hsdM gene (encoding type I methyltransferase), a known integration site for Tn1806 family ICEs, and the rplL site has been occupied by a resident ICE (Figure 4B). After three independent mating experiments, the Tn1806-like ICE transferred only once. The transconjugant counts obtained were 0, 0, and 7, respectively, none of which reached the minimum countable threshold of 30 and were therefore considered as < 30; the donor counts in the mating experiments were 3.4 × 10^10^ CFU, 4.1 × 10^10^ CFU, and 3.2 × 10^10^ CFU. Therefore, calculated conjugation transfer frequency was < 10^−9^. Whole-genome sequencing confirmed Tn1806-like ICE were compensatorily integrated into hsdM genes in recipient SagR31 (Figure 4C).
Prevalence of composite genomic elements resembling ICESag39
3.6
We screened the NCBI GenBank database using BlastN to identify composite ICEs structurally similar to ICESag39 in S. agalactiae. Details of the identified ICE, including the host strain, accession number, genomic location, length, and carried antibiotic resistance genes, are summarized in Supplementary Table 4. In total, 199 ICESag39-like ICEs were identified, including seven from complete genomes and 192 from whole-genome shotgun contigs, all integrated 3′ rplL gene. These strains were unloaded from different years (2012–2025), with 74.4% (148/199) from the United States, 16.6% (33/199) from China, 4.0% (8/199) from South Korea, 2.0% (4/199) from Brazil, 1.5% (3/199) from the United Kingdom, 1.0% (2/199) from Italy, and 0.5% (1/199) from Japan. Their sizes ranged from 90,473 to 138,541 bp. Analysis of resistance gene carriage revealed that 62.8% (125/199) of these ICEs harbored both erm(TR) and cadA, 22.1% (44/199) carried only cadA, and 4.5% (9/199) carried only erm(TR). Additionally, 1.5% (3/199) carried aac(6′)-aph(2″) together with erm(TR), whereas 7.0% (14/199) possessed erm(TR) along with catQ–mef (I). Four ICEs lacked any known resistance genes. Notably, nearly all resistance genes were located within the embedded Tn1806-like ICE module of these composite elements.
Discussion
4
Macrolide resistance determinants, such as erm family genes and the heavy metal resistance gene cadA, are widely recognized as common resistance factors. The erm(B) gene in particular exhibits a detection rate as high as 89.3% in S. agalactiae, posing a challenge of global significance (Moroi et al., 2019; Hatrongjit et al., 2025). In 2019, the US Centers for Disease Control and Prevention identified 21 antibiotic resistance threats, classifying clindamycin-resistant S. agalactiae and erythromycin-resistant S. pyogenes at the “concerning” level (CDC, 2019). Moreover, cadA has frequently been detected on plasmids from erm(B)- or mef (A)-positive isolates, indicating a potential linkage between these commonly co-occurring resistance genes (Costa et al., 2016).
This study identified a novel composite ICE characterized by a nested architecture, formed by an ICESa2603 family backbone integrated with a Tn1806-like ICE, spanning 112,852 bp in length. The embedded Tn1806-like ICE constitutes an autonomous genetic module featuring four variable regions (VR1–VR4) that act as insertion hotspots for additional resistance determinants. Specifically, ICESp2907 carrying erm(TR) and an uncharacterized cadA-carrying element were identified within VR3 and VR4, respectively (Giovanetti et al., 2012). Comparative genomic analysis further revealed that VR1 and VR2 might harbor resistance genes such as aac(6′)-aph(2″) (aminoglycoside), erm(B) (MLS_B_), and tet(M) and potentially accommodate entire mobile elements such as Tn6002, Tn916, and Tn5253. The backbone of ICESag39 represents a typical member of the ICESa2603 family, as evidenced by its tyrosine integrase, specific integration site rplL gene, and flanking 15-bp direct repeat sequences (Marini et al., 2015; Huang et al., 2016a). This backbone provides a stable genomic platform for the acquisition of exogenous DNA and rplL as the defined integration locus. By contrast, the embedded Tn1806-like ICE substantially expands the accessory gene content available to the ICESa2603 family by serving as an insertion site for smaller genetic elements. This Russian doll-like nesting of mobile genetic elements produced the formation of the composite ICE ICESag39, facilitating the co-localization and potential co-transfer of diverse resistance genes.
Through a systematic comparative analysis of Tn1806-like ICEs, this study first expanded the recognized diversity of this ICE group and elucidated the molecular basis for the formation of the composite ICE ICESag39. The Tn1806-like ICE identified in this study was integrated within the 6-kb (GGG–GGG) snf2 gene of ICESag39, an integration site that differs from the previously characterized hsdM (encoding a type I methyltransferase) target observed in Tn1806 and ICESpy10750 (Beres and Musser, 2007; Camilli et al., 2008; Ambroset et al., 2016). An expanded screening of Tn1806-like ICEs identified four additional Tn1806-like ICEs from Gen Bank also integrated within the snf2 gene, indicating that snf2 represents a previously overlooked integration site for this ICE family. Notably, snf2 has been reported as a conserved gene in the ICESa2603 family, and it is also present in enterococcal pheromone-responsive plasmids such as pBET_5 (Shan et al., 2022; Yang et al., 2022). The targeting of snf2 as an integration site strongly suggests that the composite structure of ICESag39, arising from the combination of ICESa2603 and Tn1806-like ICEs, is not an occasional event, but reflects a high-affinity interaction. This interpretation is further supported by the identification of 199 similar ICESag39-like ICEs in public databases. However, no composite plasmid-ICE structures associated with Tn1806-like ICEs or enterococcal pheromone-responsive plasmids were detected, despite the presence of the snf2 gene in such plasmids and prior evidence supporting the interspecies transferability of Tn1806-like ICEs. This absence might reflect specific genetic or physiological barriers that limit the formation of such composite, a possibility that remains to be investigated.
ICESag39 represents a novel class of composite ICEs that was demonstrated for the first time in this study to possess a modular architecture and to be transferable to other Sagalactiae strains. The embedded Tn1806-like ICE, which acts as a primary carrier of antibiotic and heavy metal resistance genes, is not a fixed component of the structure. Conjugation experiments confirmed that individual internal ICEs within ICESag39 can be excised from the composite and transferred independently. However, detection of circular intermediates and conjugation frequency indicated that the transfer efficiency of internal ICEs is remarkably low, suggesting that ICESag39 generally maintains its structural integrity under most conditions. In situations unfavorable for the transfer of the composite ICE, such as when its length imposes a high fitness cost or when suitable integration sites are absent in the recipient, the ability of internal Tn1806-like ICEs to transfer independently might provide an alternative dissemination pathway. This structural flexibility highlights a distinct adaptive advantage of composite ICEs over simple genetic elements. The assembly of erm(TR) and cadA within a single transferable ICE provides important insights into the evolution of ICE diversity and the horizontal dissemination of resistance determinants. Integrating these advantageous genes into a large, mobile composite ICE both increases the risk of multidrug-resistant “superbug” emergence and accelerates the diversification of bacterial genomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ambroset C. Coluzzi C. Guédon G. Devignes M. D. Loux V. Lacroix T. . (2016). New insights into the classification and integration specificity of Streptococcus integrative conjugative elements through extensive genome exploration. Front. Microbiol. 6:1483 doi: 10.3389/fmicb.2015.0148326779141 PMC 4701971 · doi ↗ · pubmed ↗
- 2Aziz R. K. Bartels D. Best A. A. Dejongh M. Disz T. Edwards R. A. . (2008). The RAST Server: rapid annotations using subsystems technology. BMC Genom. 9:75. doi: 10.1186/1471-2164-9-7518261238 PMC 2265698 · doi ↗ · pubmed ↗
- 3Bankevich A. Nurk S. Antipov D. Gurevich A. A. Dvorkin M. Kulikov A. S. . (2012). SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.002122506599 PMC 3342519 · doi ↗ · pubmed ↗
- 4Beres S. B. Musser J. M. (2007). Contribution of exogenous genetic elements to the group A Streptococcus metagenome. P Lo S ONE 2:e 800. doi: 10.1371/journal.pone.000080017726530 PMC 1949102 · doi ↗ · pubmed ↗
- 5Brenciani A. Bacciaglia A. Vecchi M. Vitali L. A. Varaldo P. E. Giovanetti E. (2007). Genetic elements carrying erm(B) in Streptococcus pyogenes and association with tet(M) tetracycline resistance gene. Antimicrob. Agents Chemother. 51, 1209–1216. doi: 10.1128/AAC.01484-0617261630 PMC 1855496 · doi ↗ · pubmed ↗
- 6Camilli R. Del Grosso M. Iannelli F. Pantosti A. (2008). New genetic element carrying the erythromycin resistance determinant erm(TR) in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 52, 619–625. doi: 10.1128/AAC.01081-0718070957 PMC 2224770 · doi ↗ · pubmed ↗
- 7CDC (2019). Centers for Disease Control and Prevention. Avaliable online at: http://www.cdc.gov/drugresistance/threat-report-2019. (Accessed December 01, 2023)
- 8Chen L. Huang J. Huang X. He Y. Sun J. Dai X. . (2021). Horizontal transfer of different erm(B)-carrying mobile elements among Streptococcus suis strains with different serotypes. Front. Microbiol. 12:628740. doi: 10.3389/fmicb.2021.62874033841355 PMC 8032901 · doi ↗ · pubmed ↗