First report of serological and molecular detection of influenza A (H1N1) in European bison (Bison bonasus)
Anna Didkowska, Víctor Martín-Santander, Anna Golke, Marlena Wojciechowska, Wanda Olech, Olga Szaluś-Jordanow, Tomasz Dzieciątkowski, Álex Gómez, Sergio Villanueva-Saz, Ewelina Kwiecień, Pablo Quilez, Krzysztof Anusz, Diana Marteles-Aragüés

TL;DR
This study reports the first evidence of H1N1 influenza in European bison, showing the virus can infect this wild species.
Contribution
First molecular and serological confirmation of H1N1pdm in European bison (Bison bonasus).
Findings
Seroprevalence of Influenza A virus was 0.3% in European bison samples.
A/H1N1pdm RNA was detected in heart and liver tissues of a 4-year-old male bison.
Findings suggest European bison may be susceptible to Influenza A virus spillover.
Abstract
Influenza A virus is a zoonotic pathogen capable of infecting a wide range of hosts, yet data on infections in wild ruminants are limited. The European bison (Bison bonasus), Europe’s largest terrestrial mammal, had not previously been identified as a host for Influenza A virus. This study aimed to provide the first molecular and serological evidence of H1N1 pandemic influenza A virus (A/H1N1pdm) infection in European bison, contributing to the understanding of Influenza A virus ecology at the wildlife–livestock interface. We retrospectively analysed 335 serum samples collected from European bison between 2017 and 2023 using a commercial ELISA detecting antibodies against all Influenza A virus subtypes. Tissue samples (heart and liver) from ELISA-positive animal were further tested by RT-qPCR targeting hemagglutinin (HA) and neuraminidase (NA) genes of A/H1N1pdm, A/H3N2, and A/H5N1.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
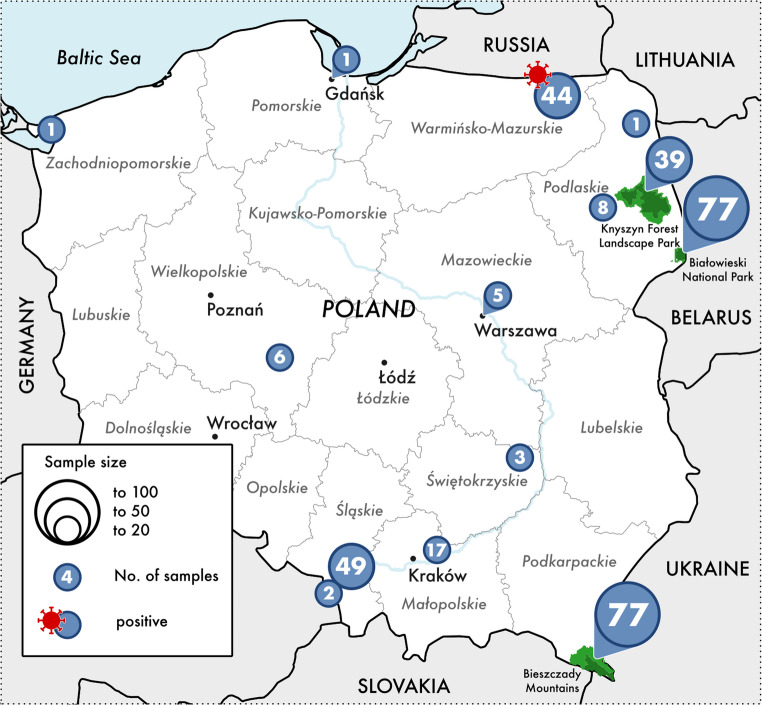 Figure 1
Figure 1- —Universidad de Zaragoza
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfluenza Virus Research Studies · Respiratory viral infections research · Infectious Encephalopathies and Encephalitis
Introduction
Highly pathogenic avian influenza (HPAI) viruses threaten wild birds and poultry globally. However, recently, some reports have appeared about spillover of these viruses to mammals (Burrough et al. 2024), including humans (Pulit-Penaloza et al. 2024). Wild waterfowl, particularly those inhabiting wetlands, serve as natural reservoirs for these viruses, facilitating their global distribution through migratory patterns. The genetic diversity of HPAI viruses, driven by reassortment events, has led to the emergence of novel strains capable of crossing species barriers (Ge et al. 2017).
The 2009 H1N1 pandemic underscored the potential for Influenza Avian Viruses (IAVs) to expose a wide array of hosts, including swine (Ryt-Hansen et al. 2025), turkeys (Berhane et al. 2016), cats (Villanueva-Saz et al. 2024), dogs (Duijvestijn et al. 2025), and various wildlife species (De Marco et al. 2022; Puryear and Runstadler 2024). Notably, instances of human-to-animal transmission have been documented, highlighting the bidirectional nature of IAV spread. While domestic ruminants have historically exhibited limited susceptibility to IAVs, serological evidence suggests sporadic infections, often coinciding with human influenza outbreaks (Sreenivasan et al. 2019).
The European bison (Bison bonasus), a free-ranging ruminant species, has not been extensively studied in terms of IAV susceptibility. Given the expanding host range of IAVs and the ecological significance of bison populations, investigating potential H1N1 infections in this species is imperative.
The aim of this study is to present a retrospective IAV-serological study in European bison, combined with the first serological and molecular confirmation of influenza in this species.
Materials and methods
Sampling
We have performed a retrospective serological study of 335 blood samples collected from European bison between 2017 and 2023. Samples were collected during planned chemical immobilisation or during necropsy (animals found dead or culled). No immobilization or culling was performed for the purpose of our study. In the case of post-mortem examination, tissue material (liver and heart fragments) was collected and frozen at -20 °C until further analysis.
Blood derived from 185 females and 150 males, aged from 0.25 to 25 years old (mean 6.7). Age was assessed based on dentition, horn appearance, and body mass according to previously described criteria (Krasińska and Krasiński 2002). Animals derived from both captive (living in captivity, primarily in breeding centers, but also in zoological gardens) (n = 157) and free-ranging (n = 178) herds (Fig. 1, five locations unknown). The captive herds comprised 82 females and 75 males, with an age range of 0.25–25 years. The free-living herds comprised 102 females and 75 males, and the age of animals ranged from one to 20 years.
Fig. 1. Location of European bison (Bison bonasus) tested serologically for influenza A
Blood was collected from the jugular vein into sterile tubes containing a clot activator (Medlab Products, Raszyn, Poland). The samples were transported to the laboratory at 4 °C. Next, centrifugation was performed and serum was separated. Serum samples were stored at -20 °C until further analysis.
Detection of influenza A antibodies by ELISA
Antibodies to Influenza A were detected by using a commercial test ID Screen Influenza A Antibody Competition Multi-species (Innovative Diagnostics, Grabels, France). The test detects antibodies against all Influenza A subtypes and antigenic variants due to the use of a monoclonal antibody against a highly conserved epitope of the Influenza A virus nucleoprotein. The test was performed according to the manufacturer’s instructions at 1/40 dilution of the serum sample. No other part of the instruction protocol was modified.
Additionally, the ID Screen^®^ Influenza H5 Antibody Competition 3.0 Multi-species kit (Innovative Diagnostics, Grabels, France) was performed according to the manufacturer’s instructions to detect antibodies against hemagglutinin H5 (H5). This assay is an upgraded version of the ID Screen^®^ Influenza H5 Antibody Competition ELISA kit, offering improved detection of emerging clades, including clade 2.3.4.4. For the detection of hemagglutinin H9 (H9), the ID Screen^®^ Influenza H9 Indirect kit (Innovative Diagnostics, Grabels, France) was modified by replacing the commercial conjugate with a conjugate consisting of protein A/G peroxidase diluted 1:10,000 in PBS containing 0.05% Tween 20 and1% dry skim milk. This conjugate binds to immunoglobulin G from various animal species, including European bison (Didkowska et al. 2025). No other part of the instruction protocol was modified. These assays were performed to serum samples that tested positive using the commercial ELISA for influenza A antibody detection, serving as complementary diagnostic techniques.
Detection of influenza A RNA by RT-qPCR
Molecular testing (RT-qPCR) was performed only on tissues available from the single ELISA-positive European bison. For this purpose, total RNA was isolated from 50 mg of heart and liver tissue using the Total RNA Mini Kit (A&A Biotechnology, Gdańsk, Poland), following the manufacturer’s instructions. The RNA was subsequently purified and concentrated to a final sample volume of 45 µl using the Clean-Up RNA Concentrator Kit (A&A Biotechnology, Gdańsk, Poland), according to the manufacturer’s instructions.
One-step reverse transcription real-time PCR (RT-qPCR) was conducted using the CFx96 system (Bio-Rad, Hercules, CA, USA) with an in-house method described by Stefańska et al. (2013) for IAVs. In this study, pairs of primers complementary to the genes encoding hemagglutinin and neuraminidase of the influenza subtypes A/H1N1pdm, A/H3N2, and A/H5N1 were used. Importantly, in this study, RT-qPCR was used as a qualitative detection method only; cycle threshold (Ct) values are reported only as contextual indicators of relative template abundance and not as calibrated viral load measurements.
Where RT-qPCR was positive, we attempted sequencing. Given known limitations of next-generation sequencing (NGS) in low-viral-load, degraded, or inhibitor-rich wildlife matrices, we prioritized Sanger sequencing of RT-qPCR amplicons. Sequencing was performed by Genomed (Warsaw, Poland) using the same primers as for RT-qPCR.
Sequencing of A/H1N1 RNA
Before RT-qPCR, which was performed for downstream sequencing of the amplification products, RNA extracted from the samples using the Total RNA Mini Kit (Total RNA Mini, A&A Biotechnology, Gdańsk, Poland) was additionally purified to remove DNA residues and concentrated (Clean-Up RNA Concentrator, A&A Biotechnology, Gdańsk, Poland). Partial Sanger sequencing was performed on RT-qPCR amplicons generated with primers described in Stefańska et al. (2013), at a certified commercial laboratory (Genomed, Warsaw, Poland). The raw sequences were manually checked and corrected using the Chromas software (version 2.6.6). A Basic Local Alignment Search Tool (BLAST) analysis was carried out on the National Center for Biotechnology Information (NCBI) web site (http://blast.ncbi.nlm.nih.gov) (Altschul et al. 1997).
Results
Seroprevalence of influenza A virus
The seroprevalence of Influenza A infection was determined to be 0.3% (1/335). The positive European bison was 4-years old male from a captive herd in Borecka Forest (Wolisko), culled and sampled in 2018 due to posthitis. The necropsy revealed poor body condition, pulmonary congestion and emphysema, and posthitis. European bison classified as seropositive by ELISA had an optical density (OD) of 0.2747. By contrast, the remaining serum samples were classified as seronegative by ELISA (mean ± standard deviation: 1.29 ± 0.13 OD). For the only seropositive sample detected by the commercial ELISA for influenza A antibody detection, the H5 and H9 ELISA kits yielded OD values similar to the negative control for each test.
RT-qPCR for influenza A
Genetic material of the influenza A virus subtype A/H1N1pdm was detected in heart and liver tissues. In the heart tissue, the Ct values for the H1 and N1 genes were 23.36 and 26.98, respectively. In the liver tissue, the Ct values for H1 and N1 were 28.83 and 37.32, respectively.
Sequencing of A/H1N1 RNA
Analysis of the 56-bp hemagglutinin gene sequence showed 100% identity and 100% coverage with influenza A(H1N1) virus sequences deposited in the GenBank database. Similarly, BLAST analysis of the 72-bp neuraminidase gene sequence obtained with the second primer pair revealed 100% identity and 100% coverage with influenza A/H1N1 neuraminidase gene sequences. The short sequence lengths precluded detailed genotypic classification. Analysed sequences were H1 (56 bp: GTGCTATAAACACCAGCCTCCCATTTCAGAATATACATCCGATCACAATTGGAAAA) and N1 (72 bp: TTGTGGTGTAAACAGTGACACTGTGGGTTGGTCATGGCCAGA GGTGCTGAGTTGCCATTTACCATTGACAA).
Discussion
The molecular and serological confirmation of H1N1 IAV infection in European bison represents a significant advancement in understanding the epidemiology of IAV among wild ruminants. This finding underscores the potential for European bison to be susceptible hosts for IAV.
The detection of H1N1 in European bison raises questions about the sources and routes of transmission. The detection of the virus in only one among 335 examined animals suggests a sporadic event in the population. The positive animal was a four-year-old captive male culled due to poor health conditions, which may have increased its susceptibility to infection or facilitated viral detection. The source of infection cannot be clearly identified. However, given the captive status of the animal, incidental exposure related to management conditions cannot be excluded. Possible transmission routes include contact with humans or indirect exposure via the shared environment, as H1N1 viruses are known to circulate primarily in human populations (Lakdawala and Brooke 2019).
The detection of H1N1 in a European bison suggests that similar mechanisms of interspecies transmission could be at play, especially in environments where wildlife and domestic animals interact closely. Given the ecological and conservation importance of European bison, understanding their susceptibility to IAVs is crucial for both wildlife management and public health. Especially, since the number of IAV reported cases is growing in both avian and mammalian species in Europe and Poland (Golke et al. 2024; Szaluś-Jordanow et al. 2023, 2024).
We used a serology-first, reflex molecular approach that fits routine wildlife surveillance and the realities of opportunistic sampling. Within this setup, qualitative RT-qPCR together with short-amplicon Sanger confirmation provides solid evidence of A/H1N1 infection in a European bison, while also showing why lineage-level typing is often not feasible in post-mortem wildlife material. Overall, our data point to susceptibility of large wild ruminants to IAV and support pragmatic diagnostics when complete respiratory sampling or high-yield sequencing cannot be obtained.
We do not infer causation between A/H1N1 detection and the European bison clinical condition. Heart and liver are not primary targets for IAV, yet viral RNA, and occasionally virus, can be detected in blood and extra-pulmonary organs in severe or disseminated infections, especially post-mortem, which aligns with our findings. Consistently, in our earlier reports, we detected IAV nucleic acid in the liver of a cat and in both the liver and heart of pet ferrets (Golke et al. 2024; Szaluś-Jordanow et al. 2023). The European bison’s captive setting may have lowered the barrier to spillover; nonetheless, confirming a natural A/H1N1 infection in a large ruminant, before the 2024 U.S. H5N1 dairy-cattle epizootic, provides an early One-Health data point for cross-species risk in Bovidae. This relevance is reinforced by growing retrospective evidence that cattle can be exposed to multiple IAV subtypes, including human and swine-origin viruses, documented by recent serological studies (Zedan et al. 2025).
In conclusion, this study provides the first molecular and serological evidence of H1N1 IAV infection in European bison, highlighting the need for ongoing surveillance and research into the dynamics of IAV in wildlife.